31 1Program Studi Biologi, Fakultas MIPA, Universitas Tanjungpura, Jl. Prof. Dr. H. Hadari Nawawi, Pontianak
Email korespondensi: Yusniawati_94@yahoo.com
Pertumbuhan Semai Bakau Putih (Bruguiera cylindrica (L.) BI.)
Pada Tingkat Salinitas Yang Berbeda
Yusniawati1, Mukarlina1,Elvi Rusmiyanto Pancaning Wardoyo1
Abstract
Salinity increase may effect the growths of several plants, for example (Bruguiera cylindrica (L.) BI.). This research aims to identify the growth of white mangrove seedling (B. cylindrica) on different salinity levels. Additionally, it attempts to recognise salinity that could be tolerated by white mangrove seedling (B.
cylindrica). This research was conducted since Oktober to December 2016. It used Completely Randomised
Design (RAL) consisting of six (6) treatments, namely control, NaCl 2,5% (S1) treatment, 3% (S2) treatment, 3,5% (S3) treatment, 4% (S4) treatment and 4,5% (S5) treatment. The result showed that NaCl treatments gave a clear effect toward seedling heights, leaf area, root lengths, wet crown weights,dry crown and root weights and chlorophyll content. However, such treatments appeared not to give an effect toward wet root weights. NaCl treatments started to effect growths with the fall of seedling heights, leaf area and root lengths at concentration of 3% and at 4% toward wet crown weights and dry crown and root weights. Meanwhile, the effect toward chlorophyll content was at concentration of 4,5%. Salinity which could be tolerated by B.
cylindrica growth stood around 2,5% (25‰) – 3,5% (35‰) and 4% (40‰) for chlorophyll content.
Keywords: Bruguiera cylindrica, Salinity, Growth PENDAHULUAN
Bakau putih (Bruguiera cylindrica (L.) BI) merupakan salah satu tumbuhan penyusun hutan mangrove. Mangrove merupakan kelompok tumbuhan berkayu yang tumbuh di sepanjang garis pantai dan memiliki adaptasi yang tinggi terhadap salinitas. Secara fisik mangrove dapat berperan dalam menjaga pantai dan pinggiran sungai dari proses abrasi (Arief, 2003).
Pertumbuhan B. cylindrica di hutan mangrove sangat dipengaruhi oleh lingkungan seperti salinitas. Menurut Bengen (2001) salinitas sangat menentukan perkembangan hutan mangrove, hal ini dapat terjadi karena pengaruh salinitas diantaranya dapat membagi hutan mangrove menjadi beberapa zonasi hutan, mulai dari zonasi terdekat atau berbatasan dengan laut (zona proksimal) hingga zonasi terjauh dari laut (zona distal).
Peningkatan salinitas dapat terjadi karena perubahan cuaca misalnya meningkatnya suhu dan kurangnya curah hujan. Keadaan tanah yang salin dapat mempengaruhi penyerapan air serta nutrisi oleh tumbuhan dan menyebabkan terjadinya gangguan osmotik, serta berakibat pada penurunan pertumbuhan tumbuhan (Gao et al., 2008). Salinitas tinggi dalam jangka waktu yang
lama juga dapat menurunkan kandungan klorofil per unit area daun (Robinson et al., 1983 dalam Marchner, 1986).
Menurut Tan (1991) garam-garam yang mendominasi pada lahan mangrove adalah natrium klorida (NaCl). Penelitian tentang pengaruh NaCl terhadap pertumbuhan tumbuhan mangrove telah dilakukan diantaranya pada jenis
Avicennia marina (Refik, 2005), Ceriops tagal
(Ramayani, et al., 2012) dan Rhizophora
apiculata (Prayunita et al., 2012).
B. cylindrica umumnya memiliki mekanisme
toleransi terhadap salinitas dengan mencegah masuknya NaCl (salt exclusion) melalui saringan atau ultra filter yang terdapat pada akar (Hutching & Saenger, 1987), sehingga tidak semua NaCl diserap ke dalam tubuh tumbuhan, karena itu penelitian ini dilakukan untuk mengetahui pertumbuhan semai bakau putih (B. cylindrica) pada tingkat salinitas yang berbeda dan mengetahui salinitas yang dapat ditoleransi oleh semai bakau putih (B. cylindrica).
BAHAN DAN METODE Waktu dan Tempat Penelitian
Penelitian dilaksanakan selama 3 bulan, penanaman propagul B. cylindrical dengan
berbagai tingkat salinitas dilakukan pada bulan Oktober 2016 – Desember 2016. Penanaman dan pemeliharaan dilakukan di rumah kasa Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Tanjungpura, Pontianak.
Bahan
Bahan yang digunakan yaitu air ledeng, air laut, bubuk garam NaCl, media tanah dari hutan mangrove desa Sungai Bakau Besar Laut dan propagul B. cylindrica yang sehat dan matang.
Metode Penelitian
Penelitian dilakukan dengan menggunakan Rancangan Acak Lengkap (RAL) yang terdiri dari 6 perlakuan yaitu kontrol, perlakuan NaCl 2,5% (S1); 3% (S2); 3,5% (S3); 4% (S4) dan 4,5% (S5). Masing-masing perlakuan dilakukan pengulangan sebanyak 4 kali sehingga diperoleh 24 unit percobaan.
Prosedur Kerja
Persiapan Media Tanam
Media tanam yang digunakan adalah tanah dari hutan mangrove. Tingkat salinitas 2,5% (S1); 3% (S2); 3,5% (S3); 4% (S4) dan 4,5% (S5) dibuat dengan melarutkan bubuk garam NaCl masing-masing sebanyak 28,33; 34; 39,66; 45,33 dan 51 g dalam 1 liter air. Sedangkan pada perlakuan kontrol dilakukan penyiraman tanaman dengan air hujan dan air laut sesuai dengan kondisi lingkungan pada umumnya.
Pemilihan dan Penanaman Propagul
Propagul B. cylindrica yang digunakan merupakan propagul yang telah tua. Pemilihan propagul dapat dilihat secara morfologi dengan warna propagul hijau keunguan (Rusila et al., 1999). Propagul yang telah dipilih dibersihkan pada air yang mengalir kemudian ditanam pada
polybag yang telah berisi media tumbuh sesuai
dengan perlakuannya masing-masing dan diberi label.
Analisis Media Tanah
Media tanah dianalisis di Laboratorium Analilis Kimia Tanah, Fakultas Pertanian, Universitas Tanjungpura Pontianak. Pengukuran dilakukan sebelum tanah digunakan. Kandungan kimia tanah yang dianalisis meliputi Unsur hara N, P dan K.
Parameter Pengamatan
Parameter Pengamatan adalah parameter pertumbuhan dan parameter lingkungan. Adapun parameter pertumbuhan yang diamati adalah tinggi semai (cm), luas daun (cm2), panjang akar (cm), berat basah akar dan tajuk (g), berat kering
akar dan tajuk (g) dan kandungan klorofil daun (spad/unit). Parameter lingkungan yang diamati meliputi salinitas dan pH tanah, kelembapan tanah, suhu udara dan kelembapan udara rumah kasa.
Analisis Data
Data hasil pengamatan variabel tinggi semai, luas daun, panjang akar, berat basah akar dan tajuk, berat kering akar dan tajuk serta kandungan klorofil daun dianalisis menggunakan Analisys of
Variance (ANOVA). Kemudian dilakukan uji
lanjut menggunakan Duncan’s Multi Range Test (DMRT) pada taraf 5% terhadap tinggi semai, luas daun, panjang akar, berat basah tajuk, berat kering akar dan tajuk serta kandungan klorofil daun (Walpole, 1995).
HASIL DAN PEMBAHASAN Hasil
Berdasarkan hasil analisis anova perlakuan salinitas (NaCl) berpengaruh nyata terhadap tinggi semai B. cylindrica (F5,23 = 24,792, p = 0,000). Uji lanjut Duncan menunjukkan perlakuan S1 tidak berbeda nyata dengan kontrol sedangkan S2, S3, S4 dan S5 berbeda nyata (Gambar 1).
Gambar 1. Rerata tinggi semai bakau putih (B.cylindrica) pada perlakuan NaCl
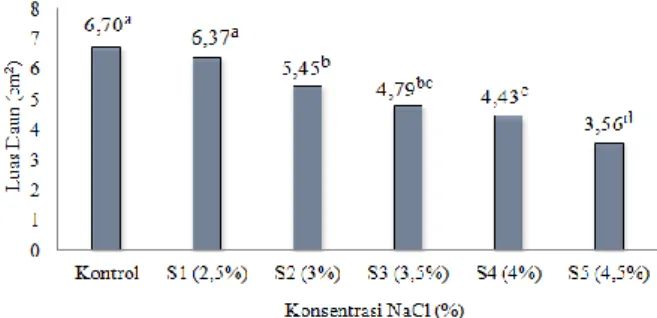
Hasil analisis anova perlakuan NaCl berpengaruh nyata terhadap luas daun (F5,23 = 17,863, p = 0,000). Uji lanjut Duncan menunjukkan perlakuan S1 tidak berbeda nyata dengan kontrol, tetapi berbeda nyata dengan perlakuan lainnya. Rerata luas pada perlakuan S2, S3, S4 dan S5 berbeda nyata dengan kontrol dan S1 (Gambar 2).
Gambar 2. Rerata luas daun semai bakau putih (B.cylindrica) pada perlakuan NaCl
33 Hasil analisis anova menunjukkan perlakuan NaCl
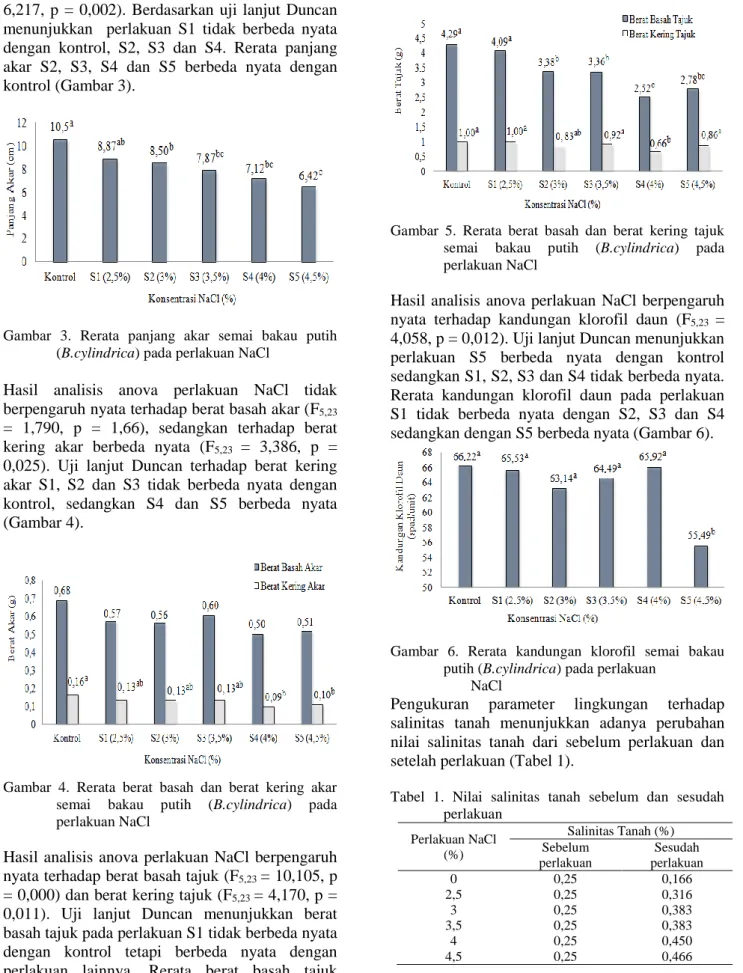
berbengaruh nyata terhadap panjang akar (F5,23 = 6,217, p = 0,002). Berdasarkan uji lanjut Duncan menunjukkan perlakuan S1 tidak berbeda nyata dengan kontrol, S2, S3 dan S4. Rerata panjang akar S2, S3, S4 dan S5 berbeda nyata dengan kontrol (Gambar 3).
Gambar 3. Rerata panjang akar semai bakau putih (B.cylindrica) pada perlakuan NaCl
Hasil analisis anova perlakuan NaCl tidak berpengaruh nyata terhadap berat basah akar (F5,23 = 1,790, p = 1,66), sedangkan terhadap berat kering akar berbeda nyata (F5,23 = 3,386, p = 0,025). Uji lanjut Duncan terhadap berat kering akar S1, S2 dan S3 tidak berbeda nyata dengan kontrol, sedangkan S4 dan S5 berbeda nyata (Gambar 4).
Gambar 4. Rerata berat basah dan berat kering akar semai bakau putih (B.cylindrica) pada perlakuan NaCl
Hasil analisis anova perlakuan NaCl berpengaruh nyata terhadap berat basah tajuk (F5,23 = 10,105, p = 0,000) dan berat kering tajuk (F5,23 = 4,170, p = 0,011). Uji lanjut Duncan menunjukkan berat basah tajuk pada perlakuan S1 tidak berbeda nyata dengan kontrol tetapi berbeda nyata dengan perlakuan lainnya. Rerata berat basah tajuk perlakuan S2, S3, S4 dan S5 berbeda nyata dengan kontrol dan S1. Uji Duncan terhadap berat kering tajuk menunjukkan perlakuan S4 berbeda
nyata dengan kontrol sedangkan S1, S2, S3 dan S5 tidak berbeda nyata (Gambar 5).
Gambar 5. Rerata berat basah dan berat kering tajuk semai bakau putih (B.cylindrica) pada perlakuan NaCl
Hasil analisis anova perlakuan NaCl berpengaruh nyata terhadap kandungan klorofil daun (F5,23 = 4,058, p = 0,012). Uji lanjut Duncan menunjukkan perlakuan S5 berbeda nyata dengan kontrol sedangkan S1, S2, S3 dan S4 tidak berbeda nyata. Rerata kandungan klorofil daun pada perlakuan S1 tidak berbeda nyata dengan S2, S3 dan S4 sedangkan dengan S5 berbeda nyata (Gambar 6).
Gambar 6. Rerata kandungan klorofil semai bakau putih (B.cylindrica) pada perlakuan NaCl
Pengukuran parameter lingkungan terhadap salinitas tanah menunjukkan adanya perubahan nilai salinitas tanah dari sebelum perlakuan dan setelah perlakuan (Tabel 1).
Tabel 1. Nilai salinitas tanah sebelum dan sesudah perlakuan Perlakuan NaCl (%) Salinitas Tanah (%) Sebelum perlakuan Sesudah perlakuan 0 0,25 0,166 2,5 0,25 0,316 3 0,25 0,383 3,5 0,25 0,383 4 0,25 0,450 4,5 0,25 0,466 Pembahasan
Hasil penelitian menunjukkan bahwa pertumbuhan tinggi semai, luas daun dan panjang
akar B. cylindrica pada konsentrasi NaCl 2,5% tidak berbeda nyata dengan pertumbuhan pada kontrol, sedangkan mulai konsentrasi NaCl 3% menunjukkan adanya beda nyata (Gambar 1, Gambar 2, Gambar 3). Keadaan ini menunjukkan bahwa konsentrasi NaCl 2,5% tidak mempengaruhi pertumbuhan tinggi semai, luas daun dan panjang akar B. cylindrica, sedangkan konsentrasi NaCl 3% mulai mempengaruhi pertumbuhan B. cylindrica dengan adanya penurunan pertumbuhan.
Menurunnya pertumbuhan B. cylindrica dapat terjadi akibat ion natrium (Na+) dan klorida (Cl-) menjadi lebih tinggi dalam media tumbuh sehingga mempengaruhi penyerapan air yang dibutuhkan untuk pertumbuhan tanaman. Gangguan penyerapan air dapat terjadi karena unsur Cl- yang tinggi akan meningkatkan tekanan osmotik dan membuat potensial air di luar sel menjadi negatif sehingga menganggu penyerapan air dan hara pada tanaman (Kurban, et al., (1998); Kabir et al., (2004) & Anonim (2011). Potensial air tanah yang negatif akan memacu air keluar dari jaringan dan menyebabkan tekanan turgor sel menurun, penurunan tekanan turgor ini akan mempengaruhi pemanjangan sel (Djukri, 2009). Peningkatan ion Na+ dan Cl- dalam media tumbuh dapat menurunkan penyerapan unsur hara esensial lain, sebagaimana menurut Rogers, et al., 2003; Hu & Schmidhalter (2005), terjadinya peningkatan salinitas akan menyebabkan ketidakseimbangan hara dengan meningkatnya penyerapan unsur Na+ dan Cl- sedangkan penyerapan unsur esensial lainnya, seperti Nitrogen (N), kalsium (Ca2+) dan kalium (K) menurun.
NaCl dapat masuk ke daun tanaman dan banyak terakumulasi pada daun tua. Akumulasi NaCl pada daun ini akan menyebabkan tanaman memperkecil ukuran daunnya sebagai upaya untuk menjaga tekanan turgor, sehingga menyebabkan luas daun tanaman menurun. Menurut Hutching & Saenger (1987), mekanisme toleransi B. cylindrica terhadap NaCl yang berlebih adalah dengan mencegah masuknya NaCl (salt exclusion) melalui saringan atau ultra filter pada akar dan sebanyak 90% – 95% NaCl dapat dikeluarkan dari akar sedangkan 5% – 10% NaCl dapat masuk ke daun (Atkinson et al., 1967). NaCl yang masuk ke daun tidak dapat dikeluarkan melalui daun atau organ lain karena B. cylindrica termasuk jenis tumbuhan mangrove dengan adaptasi non-secretor, yaitu tidak memiliki sistem pengeluaran NaCl, sehingga peristiwa kehilangan
NaCl hanya akan terjadi ketika daun atau bagian tumbuhan lain gugur (Clogh et al., 1982).
Perlakuan salinitas juga mempengaruhi berat akar dan tajuk, konsentrasi NaCl 2,5% – 4,5% tidak berpengaruh nyata terhadap berat basah akar, namun berpengaruh nyata terhadap berat kering akar (Gambar 4). Sedangkan perlakuan NaCl terhadap berat basah dan berat kering tajuk menunjukkan adanya pengaruh nyata (Gambar 5). Berat basah tajuk mulai menurun pada konsentrasi NaCl 3%, sedangkan berat kering akar dan tajuk mulai menurun pada konsentrasi 4%.
Perlakuan NaCl berpengaruh nyata terhadap pertumbuhan panjang akar mulai konsentrasi 3% (Gambar 3), namun tidak berpengaruh nyata terhadap berat basah akar. Hal ini diduga bahwa konsentrasi NaCl yang sudah cukup tinggi menyebabkan akar mensintesis senyawa metabolit sekunder berupa prolin sebagai upaya penyesuaian dalam perubahan osmotik, sehingga meskipun potensial air tanah negatif akar tetap dapat menyerap air dari tanah. Morgan (1984), menyatakan dalam peranannya sebagai pengaturan osmotik, prolin akan menyebabkan penurunan potensial air dalam sel, sehingga memungkinkan pengambilan air dari lingkungan ke dalam sel tanaman.
Air yang diserap oleh akar diduga masih mengandung NaCl terlarut dengan potensial air yang negatif, sehingga dapat menganggu proses pengangkutan air dari akar ke tajuk tanaman. Hal ini ditunjukkan dengan adanya pengaruh nyata perlakuan NaCl terhadap berat basah tajuk (Gambar 5). Pergerakan air ke bagian atas tanaman dapat berlangsung dari potensial air yang tinggi ke potensial air rendah, sehingga jika potensial air tanah lebih rendah dari potensial air sel tanaman maka pergerakan air ke bagian atas tanaman menjadi terganggu dan menyebabkan berkurangnya air yang sampai ke tajuk tanaman (Lakitan, 2011).
Penurunan berat kering akar dan tajuk mulai pada salinitas 4%, hal ini dapat terjadi karena adanya gangguan berbagai reaksi biokimia dalam sel seperti reaksi fotosintesis akibat defisit air. Terganggunya proses fotosintesis akan mempengaruhi hasil fotosintat, sehingga jumlah fotosintat yang diterima tanaman menjadi lebih sedikit dan mempengaruhi berat kering tanaman. Air berpengaruh pada kinerja stomata dalam proses fotosintesis, apabila kekurangan air stomata akan menutup dan menyebabkan CO2 tidak dapat berdifusi melalui daun dan proses
35 fotosintesis menjadi terganggu (Salisbury & Ross,
1995).
Karbohidrat sebagai fotosintat dari hasil fotosintesis banyak digunakan sebagai energi untuk pembentukan ATP dalam mekanisme pengeluaran NaCl dari sitoplasma. Menurut Onrizal (2005), NaCl yang dikeluarkan dari sitoplasma sel akan ditimbun dalam vakuola daun agar tidak mengganggu aktivitas enzim dan metabolisme dalam sitoplasma. Mekanisme tersebut dilakukan dengan cara memompakan NaCl dari sitosol ke vakuola. Pemompaan NaCl ini memerlukan energi metabolik yang pembentukannya membutuhkan banyak karbohidrat, akibatnya karbohidrat pada tanaman akan berkurang dan mempengaruhi hasil berat kering tanaman (Lakitan, 2011).
Mekanisme penyimpanan NaCl tidak hanya terjadi pada pada vakuola sel daun tapi juga pada batang maupun organ lainnya. Menurut Percival & Womersley (1975); Onrizal (2005), tumbuhan mangrove yang tidak memiliki kelenjar garam seperti B. cylindrica memiliki kulit luar pada batang maupun daun yang jauh lebih tebal dibandingkan dengan jenis mangrove yang memiliki kelenjar garam, penebalan kulit ini terjadi sebagai mekanisme penyimpanan NaCl yang berlebih. Penyimpanan NaCl pada kulit luar ini dapat menyebabkan bobot tanaman menjadi lebih besar, sehingga diduga tanaman B.
cylindrica pada konsentrasi NaCl 4,5% telah
menyimpan NaCl pada kulit luar batang maupun daun karena meskipun pertumbuhannya mengalami penurunan, berat kering tajuknya tidak berbeda nyata dengan kontrol (Gambar 5). Perlakuan NaCl terhadap kandungan klorofil daun pada konsentrasi 4,5% menunjukkan hasil yang berbeda nyata dengan kontrol (Gambar 6). Penurunan kandungan klorofil dapat disebabkan oleh unsur hara nitrogen yang menurun pada konsentrasi NaCl tinggi (Lakitan, 2011). Kurangnya ketersediaan nitrogen dapat menyebabkan sedikitnya pembentukan asam glutamat yang dapat berperan dalam sintesis klorofil (Salisbury & Ross, 1995; Robinson, 1995). Meskipun terjadi peningkatan salinitas, namun kandungan klorofil pada konsentrasi NaCl 3,5% dan 4% tidak berbeda nyata dengan kontrol (Gambar 6). Hal ini menunjukkan toleransi klorofil daun cukup tinggi terhadap salinitas, menurut Haryadi & Yahya (1988), Na+ dan Cl -tidak akan menghentikan sintesis klorofil secara langsung tetapi pada konsentrasi sangat tinggi
dapat mempengaruhi struktur kloroplas dan membran sel.
DAFTAR PUSTAKA
Anonim, 2011, Salinity Management Handbook, Quieensland Goverment, Departmen of Environment and Resource Management,
Diakses 15 Januari 2013,
http://www.derm.qld.gov.au./salinity
Arief, A, 2003, Hutan Mangrove Fungsi dan
Manfaatnya, Kanisius, Yogyakarta
Atkinson, MR, Findly ,AB, Hope, MG. Pitman, HDW, Sadler & West, KR, 1967, ‘Salt Regulation in the Mangrove Rhizhophora mucronata Lam. Bengen, DG, 2001, Pengenalan dan Pengelolaan
Ekosistem Mangrove, Pusat Kajian Pesisir dan
Lautan, Institut Pertanian Bogor, Bogor Clough, BF & Attiwill, PM, 1982, Primary
Productivity of Mangrove In B.F. Clough (Ed.). Mangrove Ecosystem in Australian: structure, fungtion and management,
Australian University Press, Canberra. Djukri, 2009, ‘Cekaman Salinitas terhadap
Pertumbuhan Tanaman’, Prosiding Seminar
Nasional Penelitian, Pendidikan dan Penerapan MIPA, Universitas Negeri Yogyakarta, Yogyakarta
Gao, S, Ouyang, C, Wamg, S, Xu, Y, Tang, L, & Chen, F, 2008,‘Effect of salt stress on growth, antioxidant enzyme and phenylalanine ammonialyase activities in Jatropha curcas L. Seedlings’, Plant Soil Environ, vol. 54, no. 9, hal. 374-381
Haryadi, SS & Yahya, S, (1988), Fisiologi Stress Tanaman, PAU IPB, Bogor
Hu, Y & Schmidhalter, U, 2005, ‘Drought and Salinity: A Comparison of their Effects on Mineral Nutrition of Plant’, J. Plant Nutr. Soil Sci., no. 168, hal. 541-549
Hutching, P & Saenger, P, 1987, Ecology of Mangrove, University of Queensland Press, Australia Kabir, ME, Karim, MA & Azad, MAK, 2004, ‘Effect
of Potassium on Salinity Tolerance of Mungbean (Vigna radiata L. Wilczek)’, J. Of
Biol. Sci., vol. 4, no. 2, hal. 103-110
Kurban, H, Saneoka, H, Nehira, K, Adilla, A & Fujita K, 1998, ‘Effect of Salinity on Growth and Accumulation of Organic and Inorganic Solutes in the Leguminous Plants Alhagi
pseudoalhagi and Vigna radiata’, Soil Sci. Plant Nutr., vol. 44, no. 4, hal. 589-597
Lakitan, B, 2011, Dasar-dasar Fisiologi Tumbuhan, Rajawali Pers, Jakarta
Marchner, H, 1986, ‘Mineral Nutrition of Higher
Plant’, Academic Press Harcourt Brace Jovanovich Publisher, London
Morgan, JM, 1984, ‘Osmoregulation and Water Stress in Higher Plants’ Annual Review of Plant
Physiology and Plant Molecular Biology, no.
35, hal. 299-319
Onrizal, 2005, ‘Adaptasi Tumbuhan Mangrove pada Lingkungan Salin dan Jenuh Air’, e-USU
Repository, Universitas Sumatera Utara,
Medan
Percival, M & Womersley, JS, 1975, ‘Floristics and Ecology of the Mangrove Vegetation of Papua New Guinea’, Bot. Bull, no. 8, hal. 1-96. Prayunita, Basyuni, M, & Agustina, L, 2012, ‘Respon
Pertumbuhan dan Biomassa Semai Rhizopora
apiculata BI Terhadap Salinitas dan Kandungan Lipidanya pada Tingkat Pohon’, Fakultas Pertanian Universitas
Sumatera Utara, Medan
Refik, 2005, ‘Pengaruh Kadar Garam NaCl (Salinitas) terhadap Pertumbuhan Bibit Api-api Putih (Avicennia marina (Forsk.) Vierh.) pada Media Pembibitan’, Skripsi, Fakultas Kehutanan Universitas Tanjungpura, Pontianak
Ramayani, Basyuni, M, & Agustina, L, 2012, ‘Pengaruh Salinitas terhadap Pertumbuhan dan Biomassa Semai Dan Kandungan Lipida Pohon Non-Sekresi Ceriops tagal’, Fakultas
Pertanian Universitas Sumatera Utara,
Medan.
Robinson, T, 1995, Kandungan Organik Tumbuhan
Tinggi, Institut Teknologi Bandung, Bandung
Rogers, ME, Grieve, CM, and Shannon, MC, 2003, ‘Plant Growth and Ion Relations in Lucerne (Medicago sativa L.) in Responses to the Combined Effect of NaCl and P’, Plant Soil, no. 253, hal. 187-194
Salisbury, FB & Ross, CW, 1995, Fisiologi Tumbuhan
Jilid II, ITB, Bandung
Tan, KM, 1991, Dasar-dasar Kimia Tanah, UGM. Press., Yogyakarta
Walpole, Ronald E., 1995, Pengantar Statistika Edisi
ke-3, Penertbit PT. Gramedia Pustaka Utama