PENGUJIAN EFEKTIVITAS INOKULUM CENDAWAN MIKORIZA
ARBUSKULA (CMA) DENGAN MEDIA TANAM DAN TANAMAN
INANG BERBEDA PADA RUMPUT Brachiaria humidicola
SKRIPSI
NINDYAR HARUMI SUKMA
PROGRAM STUDI NUTRISI DAN MAKANAN TERNAK FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR 2006
ABSTRACT
A Study of Effectivity Utilization of Arbuscular Mycorrhizal-Fungae Inocullum in Different Media and Host Plant on Brachiaria humidicola Grass
Sukma, N. H., P. D. M. H. Karti, and Soedarmadi
Forage supply is one effort to increase livestock productivity. The importance of supply forage must be followed by increasing the production and quality intensively. However, the problem in forage plantation is high anorganic fertilizer. The use of anorganik fertilizer is relatively expensive and cause negative effects on the environment, but use of biofertilizer is safe for environment. One of the alternative biofertilizer that can be used is arbuscular mycorrhizal fungae. Many informations have reported the advantages of arbuscular mycorrhizal fungae symbioses in green house scale, but its application in open field scale is rare and must be examined. Therefore, the objective of this experiment is to obtain a potential arbuscular mycorrhizal fungi inocullum which is useful for forage growths in open field scale.
Arbuscular mycorhhizal fungae inocullum was applied in latosol soil and
Brachiaria humidicola as host plant. A completely randomized design was used,
with 7 treatments (sand-Pueraria javanica, sand-Centrosema pubescens,
soil-Pueraria javanica, soil-Centrosema pubescens, soil-Pueraria javanica, zeolit-Centrosema pubescens and control) and 3 replications. The data were analyzed by
analysis of variance (ANOVA), and differences among the treatments were examined with Duncan Range Multiple Test.
It is concluded that the addition of mycorrhizal inocullum did not significantly increase root infection, spore number, stolon length, and shoot dry weight in period I and II, but soil-Centrosema pubescens treatment significantly (p<0.05) the increase stolon numbers in period I.
Keywords: arbuscular mycorrhizal fungae, zeolit, Pueraria javanica, Centrosema pubescens, Brachiaria humidicola
RINGKASAN
NINDYAR HARUMI SUKMA. D24102020. 2006. Pengujian Efektivitas Inokulum Cendawan Mikoriza Arbuskula (CMA) dengan Media Tanam dan Tanaman Inang Berbeda pada Rumput Brachiaria humidicola. Skripsi. Program Studi Nutrisi dan Makanan Ternak. Fakultas Peternakan. Institut Pertanian Bogor.
Pembimbing Utama : Dr. Ir. Panca Dewi M.H.K.S., MSi. Pembimbing Anggota : Prof. Dr. Soedarmadi H., MSc.
Mikoriza merupakan bentuk asosiasi antara tanaman dan cendawan yang memiliki sifat dan peran unik bagi tanaman. Asosiasi ini diketahui memiliki beraneka fungsi, misalnya meningkatkan serapan hara, meningkatkan ketahanan terhadap kekeringan, tahan terhadap serangan patogen akar dan mampu menghasilkan zat pengatur tumbuh. Tanaman yang dikolonisasi cendawan mikoriza arbuskula (CMA) akan lebih baik pertumbuhannya dan produksinya.
Penelitian yang telah dilakukan sebelumnya oleh Ariyanti (2005) dalam skala rumah kaca menghasilkan tanaman inang Pueraria javanica (Pj) dan Centrosema
pubescens (Cp) dengan media tanam zeolit dan Centrosema pubescens dengan media
tanam tanah latosol merupakan kombinasi terbaik untuk produksi massal cendawan mikoriza arbuskula. Namun, pengujian hasil inokulumnya dalam skala lapang belum dilakukan. Tujuan dari penelitian ini adalah untuk mendapatkan inokulum cendawan mikoriza arbuskula (CMA) dengan media tanam dan tanaman inang berbeda yang cocok bagi perkembangan hijauan makanan ternak khususnya Brachiaria humidicola di lahan tanah latosol sehingga dapat meningkatkan produktivitasnya.
Rancangan Acak Lengkap (RAL) dengan 7 perlakuan kombinasi media tanam pasir (P), tanah (T), zeolit (Z) dan tanaman inang berbeda (PPj, PCp, TPj, TCp, ZPj, ZCp, dan Kontrol) dengan 3 ulangan, digunakan untuk pengujian pertambahan jumlah stolon, pertambahan panjang stolon, berat kering tajuk, persentase infeksi akar dan jumlah spora. Pengaruh perlakuan terhadap peubah yang diamati dianalisis dengan menggunakan sidik ragam (ANOVA), dan jika berbeda nyata dilanjutkan dengan uji Duncan.
Tahap pengolahan tanah dilakukan dengan pembajakan, pemupukan dan pembersihan gulma. Kemudian dilakukan penanaman rumput B. humidicola bersamaan dengan inokulum CMA. CMA yang digunakan hanya empat jenis yaitu
Gigaspora margarita, Glomus manihot, Glomus etinucatum, dan Acaulospora sp.
Pemeliharaan dilakukan selama 3 bulan, periode I selama ±60 hari dan periode II ±45 hari. Setelah pemanenan dilakukan pengambilan sampel berat segar tajuk dan sampel tanah yang tercampur akar untuk penghitungan persentase infeksi akar dan jumlah spora.
Hasil penelitian menunjukkan semua inokulum CMA yang digunakan memberikan efektivitas yang sama dalam peningkatan produksi hijauan pakan ternak. Perlakuan tidak berpengaruh nyata terhadap pertambahan jumlah stolon periode I, pertambahan panjang stolon, berat kering tajuk, persentase infeksi akar dan jumlah spora pada periode I dan II. Namun, perlakuan inokulum TCp memberikan pertambahan jumlah stolon tertinggi pada periode I.
Kata-kata kunci: cendawan mikoriza arbuskula, zeolit, Pueraria javanica, Centrosema pubescens, Brachiaria humidicola
PENGUJIAN EFEKTIVITAS INOKULUM CENDAWAN MIKORIZA
ARBUSKULA (CMA) DENGAN MEDIA TANAM DAN TANAMAN
INANG BERBEDA PADA RUMPUT Brachiaria humidicola
SKRIPSI
NINDYAR HARUMI SUKMA
PROGRAM STUDI NUTRISI DAN MAKANAN TERNAK FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR 2006
PENGUJIAN EFEKTIVITAS INOKULUM CENDAWAN MIKORIZA
ARBUSKULA (CMA) DENGAN MEDIA TANAM DAN TANAMAN
INANG BERBEDA PADA RUMPUT Brachiaria humidicola
Oleh
NINDYAR HARUMI SUKMA D24102020
Skripsi ini telah disetujui dan disidangkan di hadapan Komisi Ujian Lisan pada Tanggal 9 Juni 2006
Pembimbing Utama Pembimbing Anggota
Dr. Ir. Panca Dewi M. H. K. S., MSi. Prof. Dr. Soedarmadi H., MSc. NIP. 131 672 157 NIP. 130 345 018
Dekan Fakultas Peternakan Institut Pertanian Bogor
Dr. Ir. Ronny R. Noor, MRur.Sc. NIP. 131 624 188
PENGUJIAN EFEKTIVITAS INOKULUM CENDAWAN MIKORIZA
ARBUSKULA (CMA) DENGAN MEDIA TANAM DAN TANAMAN
INANG BERBEDA PADA RUMPUT Brachiaria humidicola
NINDYAR HARUMI SUKMA D24102020
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
PROGRAM STUDI NUTRISI DAN MAKANAN TERNAK FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR 2006
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 16 Juni 1984 di Rembang, Jawa Tengah, sebagai anak kedua dari empat bersaudara pasangan Bapak Purwanto dan Ibu Eny Sakdiatuz Zahrok.
Pendidikan dasar dimulai di SD Negeri II Pamotan, Rembang pada tahun 1990-1996, selanjutnya diterima di SLTP Negeri 1 Pamotan, Rembang dan lulus pada tahun 1999. Penulis melanjutkan pendidikan menengah atas di SMU Negeri 2 Rembang dan lulus pada tahun 2002.
Pada tahun 2002, penulis diterima sebagai mahasiswa Program Studi Ilmu Nutrisi dan Makanan Ternak, Jurusan Ilmu Nutrisi dan Makanan Ternak, Fakultas Peternakan, Institut Pertanian Bogor melalui jalur Ujian Seleksi Masuk IPB (USMI). Penulis selama menjadi mahasiswa aktif di Kelompok Pecinta Alam Fakultas Peternakan IPB (KEPAL-D) dan Himpunan Keluarga Rembang Bogor (HKRB).
KATA PENGANTAR
Alhamdulillahi robbil’alamin
Puji syukur kehadirat Allah SWT atas segala limpahan nikmat, kasih sayang, dan ijinNya sehingga penulis dapat menyeleseikan skripsi yang berjudul ”Pengujian Efektivitas Inokulum Cendawan Mikoriza Arbuskula (CMA) dengan Media Tanam dan Tanaman Inang Berbeda pada Rumput Brachiaria humidicola”. Skripsi ini ditulis berdasarkan penelitian yang dilakukan penulis pada bulan Mei 2005 sampai dengan bulan Februari 2006 di Laboratorium Lapang Agrostologi dan Laboratorium Agrostologi, Program Studi Nutrisi dan Makanan Ternak, Fakultas Peternakan, Institut Pertanian Bogor, serta di Laboratorium Bioteknologi Hutan dan Lingkungan, Pusat Penelitian Sumber Daya Hayati dan Bioteknologi, Institut Pertanian Bogor.
Penyediaan hijauan makanan ternak (HMT) sebagai komponen utama pakan ternak ruminansia merupakan salah satu usaha yang dilakukan untuk kelangsungan peningkatan produktivitas ternak. Usaha peningkatan produksi dan kualitas hijauan makanan ternak secara intensif dilakukan melalui pemanfaatan spesies rumput yang cocok, disamping itu juga pemberian input yang tinggi antara lain pupuk buatan (Urea, KCl, dan SP36) untuk merangsang pertumbuhan. Mahalnya pupuk buatan merupakan salah satu alasan penulis mencari alternatif pupuk yang lebih murah. Pemanfaatan pupuk hayati berupa inokulum cendawan mikoriza arbuskula (CMA) bagi tanaman merupakan alternatif yang tepat. Selain mudah dan murah, CMA juga dapat meningkatkan produktivitas hijauan pakan.
Semoga skripsi ini bermanfaat bagi pembaca, terutama bagi peminat atau pemerhati percobaan.
Bogor, Juni 2006
DAFTAR ISI
Halaman
RINGKASAN... ii
ABSTRACT... iii
RIWAYAT HIDUP ... vi
KATA PENGANTAR ... vii
DAFTAR ISI... viii
DAFTAR TABEL... x
DAFTAR GAMBAR... xi
DAFTAR LAMPIRAN... xii
PENDAHULUAN ... 1 Latar Belakang ... 1 Perumusan Masalah ... 2 Tujuan ... 2 Manfaat ... 2 TINJAUAN PUSTAKA ... 3 Mikoriza... 3
Cendawan Mikoriza Arbuskular ... 3
Tanaman Inang ... 6 Brachiaria humidicola ... 6 Centrosema pubescens... 7 Pueraria javanica ... 7 Media Tanam ... 8 Latosol ... 8 Zeolit ... 9 Pasir ... 10 METODE... 11
Lokasi dan Waktu ... 11
Materi... 11
Rancangan... 12
Peubah... 12
Prosedur ... 14
HASIL DAN PEMBAHASAN ... 16
Pengamatan Umum... 16
Rekapitulasi Analisis Ragam ... 17
Pengaruh Perlakuan terhadap Pertambahan Jumlah Stolon... 17
Pengaruh Perlakuan terhadap Pertambahan Panjang Stolon ... 19
Pengaruh Perlakuan terhadap Persentase Infeksi Akar... 22
Pengaruh Perlakuan terhadap Jumlah Spora... 24
KESIMPULAN DAN SARAN ... 27
Kesimpulan ... 27
Saran ... 27
UCAPAN TERIMA KASIH ... 28
DAFTAR PUSTAKA ... 29
DAFTAR TABEL
Nomor Halaman
1. Analisis Tanah Latosol ... 9
2. Rekapitulasi Analisis Ragam ... 17
3. Rataan Pertambahan Jumlah Stolon (buah per minggu)... 18
4. Rataan Pertambahan Panjang Stolon (cm per minggu) ... 19
5. Rataan Berat Kering Tajuk (g)... 20
6. Rataan Persentase Infeksi Akar (%) ... 22
DAFTAR GAMBAR
Nomor Halaman 1. Gambar Vesikula... 4 2. Gambar Arbuskula ... 4 3. Penampang Memanjang Anatomi Mikoriza yang Disederhanakan 5 4. Rumput Brachiaria humidicola di Lapangan ... 16 5. Akar Terinfeksi Mikoriza... 24 6. Akar Tidak terinfeksi Mikoriza... 24
DAFTAR LAMPIRAN
Nomor Halaman
1. Sidik Ragam Pertambahan Jumlah Stolon Periode I... 33
2. Sidik Ragam Pertambahan Jumlah Stolon Periode II ... 33
3. Sidik Ragam Pertambahan Panjang Stolon Periode I ... 33
4. Sidik Ragam Pertambahan Panjang Stolon Periode II ... 33
5. Sidik Ragam Berat Kering Tajuk Periode I ... 34
6. Sidik Ragam Berat Kering Tajuk Periode II ... 34
7. Sidik Ragam Persentase Infeksi Akar Periode I ... 34
8. Sidik Ragam Persentase Infeksi Akar Periode II ... 34
9. Sidik Ragam Jumlah Spora Periode II ... 35
PENDAHULUAN Latar Belakang
Penyediaan hijauan makanan ternak (HMT) sebagai komponen utama pakan ternak ruminansia merupakan salah satu usaha yang dilakukan untuk kelangsungan peningkatan produktivitas ternak. Ketersediaan hijauan makanan ternak, didukung oleh produksi hijauan yang berkualitas, berkecukupan, dan berkelanjutan.
Usaha peningkatan produksi dan kualitas hijauan makanan ternak dilakukan melalui pemanfaatan spesies rumput yang cocok. Disamping itu juga pemberian input yang tinggi antara lain pupuk buatan (Urea, KCl, dan SP36) untuk merangsang pertumbuhan. Penggunaan pupuk buatan untuk meningkatkan produktivitas hijauan makanan ternak memiliki beberapa kendala yaitu memerlukan biaya dan energi yang relatif tinggi, serta menimbulkan dampak negatif terhadap lingkungan apabila digunakan secara berlebihan.
Pemanfaatan cendawan mikoriza arbuskula (CMA) dalam meningkatkan produktivitas hijauan makanan ternak merupakan salah satu cara yang tepat. Kolonisasi sistem perakaran cendawan mikoriza arbuskula menghasilkan manfaat langsung bagi tanaman inang yaitu meningkatkan serapan hara khususnya fosfat, meningkatkan ketahanan terhadap kekeringan, tahan terhadap serangan patogen akar dan mampu menghasilkan zat pengatur tumbuh misalnya auksin, sitokinin, dan giberelin (Imas et al., 1989). Zat pengatur tumbuh ini sangat diperlukan untuk proses pembelahan sel, memacu pertumbuhan serta mencegah atau memperlambat proses penuaan sehingga memperlama fungsi akar sebagai penyerap unsur hara dan air (Setiadi, 1989). Meningkatnya serapan hara akibat kolonisasi CMA disebabkan sedikitnya oleh tiga hal, yaitu 1) CMA mampu mengurangi jarak yang harus ditempuh permukaan akar tanaman untuk mencapai unsur hara, 2) meningkatnya serapan unsur hara dan konsentrasi pada permukaan penyerapan, 3) mengubah secara kimia sifat-sifat unsur hara kimia sehingga memudahkan penyerapan unsur hara tersebut ke dalam akar tanaman (Sylvia, 1999).
Efektivitas cendawan mikoriza arbuskula sangat tergantung pada jenis CMA, tanaman inang, dan media tumbuh. Pada penelitian sebelumnya telah diketahui bahwa Pueraria javanica dan zeolit merupakan tanaman inang dan media tanam yang baik untuk CMA (Sulistyaningsih, 2003). Pueraria javanica dan Centrosema
2
pubescens dengan media tanam zeolit, dan Centrosema pubescens dengan media
tanam tanah latosol merupakan kombinasi terbaik untuk produksi massal cendawan mikoriza arbuskula (Ariyanti, 2005).
Penelitian pemanfaatan cendawan mikoriza arbuskula untuk produksi massal CMA dengan menggunakan tanaman inang dan media tanam yang sesuai dalam skala rumah kaca telah banyak dilakukan, sedangkan pengujian hasil inokulum CMA dalam skala lapang masih jarang informasinya. Oleh karena itu perlu dilakukan penelitian lanjutan pengujian inokulum cendawan mikoriza arbuskula pada kondisi lapang.
Perumusan Masalah
Penelitian Ariyanti (2005) dalam skala rumah kaca menghasilkan inokulum CMA dengan persentase infeksi akar dan jumlah spora yang tinggi. Dari hasil diatas dilakukan penelitian lanjutan untuk menguji efektivitas inokulum CMA yang ditumbuhkan pada media tanam dan tanaman inang yang berbeda pada skala lapang dengan menggunakan Brachiaria humodicola sebagai tanaman inang, sehingga diharapkan dapat meningkatkan produksi hijauan pakan ternak dan mengurangi biaya produksinya.
Tujuan
Penelitian ini bertujuan untuk mendapatkan inokulum cendawan mikoriza arbuskula (CMA) dengan media tanam dan tanaman inang berbeda yang cocok bagi perkembangan hijauan makanan ternak khususnya Brachiaria humidicola di lahan tanah latosol sehingga dapat meningkatkan produktivitasnya.
Manfaat
Penelitian ini diharapkan bermanfaat dalam menggunakan inokulum cendawan mikoriza arbuskula (CMA) sebagai pupuk alternatif, guna meningkatkan produktivitas hijauan makanan ternak tanpa menimbulkan efek negatif terhadap lingkungan.
TINJAUAN PUSTAKA Mikoriza
Mikoriza adalah suatu bentuk hubungan simbiosis mutualistis antara cendawan (mykes) dan perakaran (rhiza) tumbuhan tinggi. Adanya bentuk asosiasi antara cendawan mikoriza dan akar, sebenarnya adalah suatu bentuk parasitisme, dimana cendawan menyerang sistem perakaran, tetapi tidak sebagaimana halnya parasit yang berbahaya (patogen). Dalam hal ini cendawan tidak merusak atau membunuh tanaman inangnya, tetapi memberikan keuntungan kepada tanaman inang (host) dan sebaliknya cendawan dapat memperoleh karbohidrat dan faktor pertumbuhan lainnya dari tanaman inang (Setiadi, 1989). Berdasarkan struktur tubuh dan cara infeksi terhadap tanaman inang, Fakuara (1988) mengelompokkan mikoriza dapat ke dalam tiga kelas yaitu :
1) Endomikoriza: perakaran yang terinfeksi tidak membesar dan tidak memiliki selubung cendawan. Adanya hifa-hifa cendawan yang menembus akar secara intraseluler yang membentuk vesikula dan arbuskula, jenis ini lebih dikenal sebagai cendawan mikoriza arbuskula.
2) Ektendomikoriza: mempunyai ciri-ciri antara ekto dan endo mikoriza, tetapi kepentingan ekologisnya lebih sedikit dibandingkan dengan kelas lainnya. Ektendomikoriza mempunyai penyebaran terbatas pada tanah-tanah hutan dan ditemukan pada pohon hutan yang secara normal membentuk ektomikoriza. 3) Ektomikoriza: mudah dikenali karena perakaran yang terinfeksi membesar dan
struktur hifa seperti jala. Hifa cendawan tidak menembus sel, tetapi hanya berada di dinding-dinding sel jaringan korteks.
Cendawan Mikoriza Arbuskula
Cendawan mikoriza arbuskula termasuk ke dalam famili Endogonaceae, ordo Mucorales, kelas Zygomucetes. Cendawan ini merupakan simbion obligat dan tidak dapat ditumbuhkan pada medium sintetik yang tidak ada tanaman inangnya (Imas et
al., 1989). Gerdemann dan Trappe (1974) mengklasifikasikan cendawan mikoriza
vesikula-arbuskula menjadi empat macam yaitu Glomus, Gigaspora, Acaulospora, dan Sclerocystis. Mycofer adalah jenis mikoriza campuran yang terdiri dari
4
Acaulospora sp. merupakan spora yang dihasilkan secara tunggal di dalam
tanah atau di dalam sporakarp yang panjangnya dapat mencapai beberapa sentimeter. Spora melekat secara lateral pada hifa yang ujungnya menggelembung, biasanya tumbuh pada kisaran pH kurang dari 5. Spora Gigaspora sp. dihasilkan secara tunggal di dalam tanah, ukurannya besar dan bervariasi (Gunawan, 1993). Gigaspora
Margarita banyak tersebar pada tanah yang masam, pada pH 4-6. Sedangkan spora Glomus sp. dapat tumbuh pada semua tipe tanah (Gai dan Lui, 2003).
Gunawan (1993) menyatakan mikoriza arbuskula termasuk dalam endomikoriza yang dicirikan oleh adanya hifa cendawan yang menembus akar secara intraseluler, ada dua fase miselium pada perkembangan mikoriza arbuskula, yaitu: 1. Miselium eksternal yang ada di dalam tanah dengan spora yang dibentuknya dan
tersebar di sekitar akar.
2. Miselium internal yang ada dalam akar tanaman bermikoriza, terdiri atas hifa tidak bercabang yang intraseluler, hifa interseluler, arbuskula, dan vesikula, serta hifa yang melingkar-lingkar (hifa gelung).
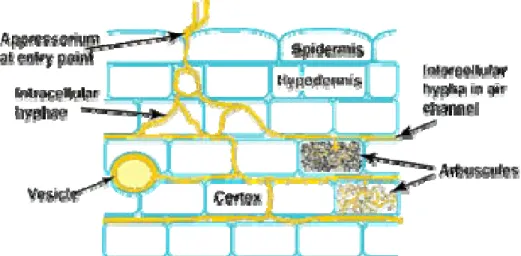
Mikoriza vesikula-arbuskula mempunyai dua macam organ yang terdapat di dalam jaringan akar yang terinfeksi, yaitu vesikula dan arbuskula (Gambar 1 dan 2).
Gambar 1. Vesikula Gambar 2. Arbuskula
(Sumber: Brundrett, 1999) (Sumber: Brundrett, 1999)
Pada fase perkembangan eksternal, perkembangan dan penyebaran cendawan arbuskula sangat berbeda-beda tergantung pada jenis tanah, tanaman dan cendawannya. Spora berasal dari miselium akar yang telah dikolonisasi oleh cendawan mikoriza arbuskula. Pada fase miselium internal, hifa yang telah terbentuk akan menembus korteks melalui epidermis atau rambut akar secara interseluler atau intraseluler. Struktur mikoriza pada fase miselium internal terdiri atas hifa interseluler yaitu hifa yang menembus sel korteks. Hifa interseluler yaitu hifa gelung
5 atau cabang-cabang hifa yang terletak di bagian tengah sel korteks, arbuskula, yang merupakan unit kolonisasi percabangan hifa yang kompleks tampak seperti pohon kecil yang mempunyai cabang-cabang, dan vesikula yang memiliki bentuk bulat yang berasal dari hifa yang membengkak (Gunawan, 1993).
Gambar 3. Penampang Memanjang Anatomi Mikoriza yang Disederhanakan (Sumber: Brundrett, 1999)
Kolonisasi sistem perakaran mikoriza arbuskula menghasilkan manfaat langsung bagi tanaman inang yaitu meningkatkan serapan hara khususnya fosfat, meningkatkan ketahanan terhadap kekeringan, tahan terhadap serangan patogen akar dan mampu menghasilkan zat pengatur tumbuh misalnya auksin, sitokinin, dan giberelin (Imas et al., 1989). Zat pengatur tumbuh ini sangat diperlukan untuk proses pembelahan sel, memacu pertumbuhan serta mencegah atau memperlambat proses penuaan sehingga memperlama fungsi akar sebagai penyerap unsur hara dan air (Setiadi, 1989). Meningkatnya serapan hara akibat kolonisasi CMA disebabkan sedikitnya oleh tiga hal, yaitu 1) CMA mampu mengurangi jarak yang harus ditempuh permukaan akar tanaman untuk mencapai unsur hara, 2) meningkatnya serapan unsur hara dan konsentrasi pada permukaan penyerapan, 3) mengubah secara kimia sifat-sifat unsur hara kimia sehingga memudahkan penyerapan unsur hara tersebut ke dalam akar tanaman (Sylvia, 1999).
Intensitas infeksi VA dipengaruhi oleh berbagai macam faktor, meliputi pemupukan, nutrisi tanaman, pestisida, intensitas cahaya, musim, kelembaban tanah, pH, kepadatan inokulum, dan tingkat keretanan tanaman. Jumlah spora dapat dihubungkan dengan jumlah infeksi akar, pada umumnya pada waktu spora
6 membentuk miselium di sekeliling akar yang menghambat perkembangan miselium bagian luar atau pertumbuhan akar dihambat oleh miskinnya suplai hara. Spora lebih banyak pada tingkat fosfat sedang daripada tingkat fosfat rendah, jika kekurangan fosfat membatasi pertumbuhan dan mempengaruhi keseluruhannya (Fakuara, 1988).
Tanaman Inang
Cendawan mikoriza arbuskula membutuhkan tanaman inang yang dapat tumbuh dengan baik pada media dan sesuai dengan spesies cendawan yang diinokulasikan. Terdapat beberapa syarat untuk inang agar perkembangan cendawan mikoriza arbuskula mencapai optimal yaitu (1) Mikotropik, (2) Dapat beradaptasi pada keadaan iklim tempat asal CMA, (3) Tumbuh baik pada medium tumbuh, (4) Tahan terhadap kekeringan dan penyakit (Sieverding, 1991). Selain itu juga toleran terhadap sifat kimia tanah yaitu asam dan basa dan memiliki perakaran yang banyak (Gunawan, 1993).
Brachiaria humidicola
Rumput ini terkenal dengan nama rumput Koronivia dan rumput Creeping signal, yang berasal dari Afrika Selatan dan kemudian menyebar ke daerah Fiji dan Papua New Guinea. Rumput ini merupakan rumput tahunan, perkembangan vegetatif dengan stolon yang begitu cepat sehingga bila ditanam di lapangan segera membentuk hamparan. Batang yang berkembang dapat mencapai tinggi 20-60 cm. Helai daun berwarna hijau terang ( Bright green ), lebar 5-6 cm dan panjang 12-25 cm. Daunnya tidak berbulu dan umumnya menggulung untuk menahan penguapan air (Jayadi, 1991).
Tanaman ini tahan kekeringan dan cukup tahan genangan akan tetapi tidak setahan Brachiaria mutica. Rumput ini tahan terhadap penggembalaan berat dan mempunyai ketahanan yang tinggi terhadap invasi gulma, tetapi kurang cocok bila dilakukan tanaman campuran dengan leguminosa karena cepat sekali menutup tanah sehingga akan menekan pertumbuhan leguminosa. Tanaman ini tahan terhadap tanah yang mengandung Aluminium tinggi dan sangat responsive terhadap pemupukan Nitrogen yang tinggi. Tanaman ini tidak tahan terhadap pembakaran, dan dapat tumbuh dengan baik dibawah pohon kelapa. Rumput ini sangat efektif untuk menahan erosi (IPB, 2002). Kapasitas produksi mencapai 20 ton/ha (Jayadi, 1991)
7 Centrosema pubescens Benth
Legume Centrosema pubescens termasuk sub familia Papilionaceae dari
familia Leguminoceae. Spesies ini berasal dari Amerika Selatan. Spesies legum ini tumbuh baik di daerah-daerah tropik dan sub tropik (Reksohadiprodjo, 1981).
Tanaman ini merupakan tanaman berumur panjang (perennial), yang tumbuh kuat dengan cara merambat dan memanjat yang tumbuh menyebar sehingga cepat menutupi tanah dengan tinggi antara 40 - 45 cm dalam waktu 4 - 8 bulan setelah ditanam. Tanaman ini berdaun banyak, batangnya agak berbulu dan batangnya tidak berkayu sampai umur 18 bulan. Legume ini tahan terhadap kekeringan karena sistem perakaran yang dalam dan tahan penggembalaan, akan tetapi jangan digembalakan sebelum berumur 1 tahun. Tanaman ini akan dapat tumbuh pada tanah yang cukup luas dari tanah berpasir sampai tanah liat. Respon terhadap pemupukan posfor yaitu 100 - 200kg P2O5 per ha. Produksi bahan kering dapat mencapai 12 ton/ha di
Australia dari kultivar Belalto. Produksi biji berkisar 300 - 600 kg/ha/tahun. Kandungan nutrisi dari tanaman ini yaitu protein kasar berkisar 11 - 24 %, mengandung asam oksalat 2,22 %, kecernaan bahan kering 53,5 %, kecernaan bahan organic 53.2 % dan kecernaan protein kasar 62,4 %. Serat kasar berkisar 33 % (IPB, 2002).
Pueraria javanica Benth
Genus Pueraria adalah legume dari sub familia Papilioneceae berasal dari Asia bagian timur dan kepulauan Pasifik, bersifat membelit, merambat, dapat membentuk semak yang rimbun dengan perakaran yang berbentuk tuber yang kuat dengan pokok akar yang disebut mahkota (crown). Kudzu tropik atau Puero (Pueraria phaseoloides Benth, Pueraria javanica Benth disebut pula Kacang ruji (Jawa), Fuo banga (Ternate) (Reksohadiprodjo, 1981).
Menurut Reksohadiprodjo (1981), Puero berbatang kuat, berbulu, sedang varietas baru yang mempunyai batang tak berbulu terdapat di Puerti Rico hasil mutasi Puero di daerah tersebut, mutant Puero ini lebih disukai ternak dibanding yang berbulu. Puero mempunyai batang-batang stolon yang dapat mengeluarkan akar dari tiap ruas batangnya yang bersinggungan dengan tanah. Perakarannya dalam dan bercabang-cabang, sehingga Puero dapat berfungsi sebagai pencegah erosi. Puero tahan pula terhadap asam dan tanah yang kekurangan kapur dan phosphor, tahan
8 permukaan air tinggi dan dapat hidup di tanah-tanah yang berat maupun berpasir. Akan tetapi, Puero tidak tahan terhadap penggembalaan berat atau pemotongan yang dilakukan sedemikian sehingga sisa tanaman hanya tinggal sedikit di atas tanah. Kudzu tropika ini mengandung 23% bahan kering, 34,6% serat kasar, 19,2% protein kasar (Hartadi et al., 1993), 0,25-0,65% fosfor, dan 0,46-0,88% Ca (Blasco dan Bohorguez, 1968).
Media Tanam
Respon tanaman tidak hanya ditentukan oleh karakteristik tanaman dan cendawan, tetapi juga oleh kondisi tanah dimana percobaan dilakukan. Efektivitas mikoriza dipengaruhi oleh faktor lingkungan tanah yang meliputi faktor abiotik (konsentrasi hara, pH, kadar air, temperatur, pengolahan tanah, dan penggunaan pupuk atau pestisida) dan faktor biotik (interaksi mikrobial, spesies cendawan, tanaman inang, tipe perakaran tanaman inang, dan kompetisi antar cendawan mikoriza). Adanya kolonisasi mikoriza tapi respon tanaman yang rendah atau tidak ada sama sekali menunjukkan bahwa cendawan mikoriza lebih bersifat parasit (Solaiman dan Hirata, 1995).
Latosol
Menurut Buckman dan Brady (1982), tanah latosol dibentuk dari pelapukan yang dipengaruhi oleh curah hujan yang besar dan suhu tinggi dari tropik dan semi tropik. Latosol mempunyai ciri warna merah atau kuning, terutama di horison B dan dapat meningkatkan drainase dalam (internal drainage). Lempung hidrat oksida tidak memiliki sifat liat dan kohesi yang mencirikan lempung silikat di suatu daerah tertentu. Tanah latosol yang digunakan untuk pertanian di bawah curah hujan besar akan mengakibatkan tanah menjadi berkersik dan sukar sekali dikerjakan.
Latosol khusus kecil kemampuan pertukaran kationnya dibandingkan dengan tanah daerah sedang yang mewakili. Hal ini sebagiab disebabkan oleh kurangnya bahan organik dan sebagian oleh sifat hidrat oksida. Mereka umumnya sangat kekurangan basa yang dapat tertukar dengan unsur hara yang tersedia. Oleh sebab itu, tanah darat akan cepat habis kesuburannya jika dikerjakan tidak dengan usaha pencegahan. Tanah ini biasanya memerlukan penutupan berat agar usaha penanaman berhasil baik (Buckman dan Brady, 1982).
9 Tabel 1. Analisis Tanah Latosol
Jenis Pengukuran Nilai Keterangan pH C Organik N P K Ca Mg
Kapasitas Tukar Kation
4-5 1,23% 0,11% 0,5 ppm 0,10 me/100g 2,10 me/100g 0,76 me/100g 13,44 me/100g Asam Rendah Sangat Rendah Rendah Rendah Rendah Rendah Rendah (Maryani, 2000) Zeolit
Zeolit merupakan kelompok mineral senyawa-senyawa aluminosilikat terhidrasi yang mengandung kation alkali dan alkali tanah, terutama Na, K, dan Ca yang mengisi rongga-rongga kerangkan aluminosilikat. Bentuk dari kerangka zeolit seperti spon dan mempunyai rongga-rongga yang berhubungan satu dengan yang lain dan merupakan saluran-saluran kosong ke segala arah dengan ukuran saluran tergantung dari garis tengah alkali atau alkali tanah yang terdapat pada strukturnya. Susunan struktur zeolit yang porous dan mempunyai banyak saluran serta rongga yang berkesinambungan tersebut menyebabkan mineral zeolit mempunyai karakter yang spesifik (Mumpton, 1984).
Mumpton (1984) juga menjelaskan bahwa zeolit mempunyai nilai kapasitas tukar kation (KTK) yang tinggi. Kapasitas tukar kation dari zeolit ini terutama merupakan fungsi dari tingkat penggantian Si oleh Al dalam struktur kerangka. Makin besar penggantian maka makin besar pula kekurangan muatan positif sehingga makin banyak pula jumlah kation-kation alkali dan alkali tanah yang dibutuhkan untuk menetralkan muatan listriknya. Kation-kation tersebut tidak terikat secara kuat dalam kerangka kristal zeolit sehingga dapat dipisahkan atau dipertukarkan secara mudah dengan cara pencucian dengan larutan kation yang lain. Kemampuan zeolit dalam menjerap maupun menukarkan kation akan dapat mengurangi kehilangan hara dalam tanah, karena proses pencucian.
10 Pasir
Pasir merupakan tanah yang bertekstur antara 2 mm-50 μ. Pasir mempunyai luas permukaan yang kecil sehingga sulit menyerap (menahan) air dan unsur hara. Selain itu, pasir juga mempunyai rasa kasar sangat jelas, tidak melekat, dan tidak dapat dibentuk bola dan gulunga (Hardjowigeno, 1992).
Pasir telah digunakan secara luas sebagai media perakaran stek karena media ini relatif murah, mudah tersedia, bersih serta memiliki daya rekat tinggi. Pasir tidak menyimpan kelembaban sehingga membutuhkan frekuensi penyiraman yang lebih tinggi. Penggunaan tunggal tanpa campuran dengan media lain membuatnya sangat kasar sehingga tidak memberikan hasil yang baik (Hartmann dan Kester, 1983).
METODE Lokasi dan Waktu
Penelitian ini dilaksanakan di Laboratorium Lapang Agrostologi dan di Laboratorium Agrostologi, Program Studi Nutrisi dan Makanan Ternak, Fakultas Peternakan, Institut Pertanian Bogor untuk penanaman, pemanenan, analisis berat kering, dan perhitungan persentase infeksi akar. Perhitungan jumlah spora dilakukan di Laboratorium Bioteknologi Hutan dan Lingkungan, Pusat Penelitian Sumber Daya Hayati dan Bioteknologi, Institut Pertanian Bogor. Penelitian berlangsung dari bulan Mei 2005 sampai dengan bulan Februari 2006. Masa penanaman periode I mulai 6 Juni sampai dengan 6 Agustus 2005 (±60 hari) dan 7 Agustus sampai 24 September 2005 (±45 hari) untuk periode II.
Materi
Materi yang digunakan dalam penelitian ini adalah inokulum CMA yang ditumbuhkan pada media tanam (pasir, latosol, zeolit) dan tanaman inang (Pueraria
javanica dan Centrosema pubescens) hasil penelitian Ariyanti (2005). Cendawan
mikoriza arbuskula yang digunakan yaitu mikofer terdiri atas Gigaspora margarita,
Glomus manihot, Glomus etinucatum, dan Acaulospora sp. diperoleh dari
Laboratorium Bioteknologi Hutan dan Lingkungan. Tanaman inang yang digunakan untuk penanaman berupa Brachiaria humidicola dan ditanam di lahan tanah latosol dengan luas 2x2 m2 per petak, sebanyak 21 petak. Pupuk yang digunakan yaitu Urea, SP36, dan KCl.
Peralatan yang digunakan pada saat penanaman dan pemeliharaan adalah cangkul, timbangan, dan selang air. Peralatan yang dipakai dalam analisis adalah gelas objek beserta kaca penutup, gelas ukur, gelas piala, pinset, gunting, oven, mikroskop, sentrifuse, saringan bertingkat, cawan petri, dan botol film. Bahan-bahan kimia yang digunakan KOH 2,5%, HCl 2%, sukrosa 60%, aquadest, dan larutan staining.
12 Rancangan
Rancangan percobaan pada penelitian ini adalah Rancangan Acak Lengkap (RAL) dengan 7 perlakuan dan 3 ulangan, yaitu:
PPj = inokulum CMA pasir-P. javanica; PCp = inokulum CMA pasir-C. pubescens; TPj = inokulum CMA tanah-P. javanica; TCp = inokulum CMA tanah-C. pubescens; ZPj = inokulum CMA zeolit-P. javanica; ZCp = inokulum CMA zeolit-C. pubescens; K = kontrol (tanpa inokulum CMA)
Penelitian ini menggunakan 21 petak percobaan, setiap petak terdapat 16 tanaman Brachiaria humidicola sebagai tanaman inang, yang ditanam di lahan tanah latosol dengan luas 2x2 m2 per petak dengan jarak tanam 40 cm. Bagan percobaan dan pengacakannya terlampir pada lampiran 10.
Semua data yang diperoleh dalam penelitian dianalisis dengan menggunakan sidik ragam (ANOVA), dan jika berbeda nyata dilanjutkan dengan uji Duncan (Mattjik dan Sumertajaya, 2002). Model statistik yang digunakan:
Yij = μ + זi + εij Keterangan: i = 1,2, … ,t
j = 1,2, … ,r
Yij = Pengamatan pada perlakuan ke-i dan ulangan ke-j
μ = Rataan umum
זi = Pengaruh perlakuan ke-i
= μi - μ
εij = Pengaruh acak pada perlakuan ke-i dan ulangan ke-j
Peubah yang Diamati
Peubah yang diamati pada penelitian ini adalah sebagai berikut: 1) Pertambahan jumlah stolon (buah)
Pertambahan jumlah stolon diperoleh dari selisih jumlah stolon antara minggu terakhir pengukuran dengan minggu sebelumnya.
13 2) Pertambahan panjang stolon (cm)
Pertambahan panjang stolon diperoleh dari selisih panjang stolon antara minggu terakhir pengukuran dengan minggu sebelumnya.
3) Berat kering tajuk (gram)
Berat kering tajuk dihitung dengan mengeringkan tajuk dioven pada suhu 700C
selama 24 jam.
4) Persentase infeksi akar (%)
Persentase infeksi akar dilakukan teknik pewarnaan akar (Phyllip dan Hayman, 1970 yang dimodifikasi oleh Koske dan Gemma, 1989). Pewarnaan akar dilakukan dengan cara akar dicuci kemudian dipotong-potong dan dimasukkan dalam tabung film, lalu ditambahkan 2,5% KOH dan tabung ditutup, dibiarkan selama 1-2 minggu. Setelah itu, akar dicuci dan diberi dengan larutan HCl 2%, direndam selama 24 jam. Kemudian diganti dengan larutan staining dibiarkan selama 24 jam dan simpan pada tabung film. Untuk menghitung infeksi akar, ambil potongan akar dengan panjang ±1 cm sebanyak 5 buah per perlakuan, kemudian diletakkan di gelas preparat dan ditutup dengan cover glass. Penghitungan persentase infeksi akar dilihat dengan menggunakan mikroskop yaitu dengan memberi tanda positif bila terdapat struktur mikoriza arbuskula (hifa, vesikula, arbuskula) setiap jarak pandang. Setiap 1 cm akar memberikan jarak pandang 7-10. Bila belum dapat dihitung akar yang terinfeksi dapat disimpan di kulkas. Persentase jumlah akar yang terinfeksi dihitung dengan rumus sebagai berikut:
Jumlah akar yang terinfeksi
Persentase infeksi = --- x 100 % Total jumlah akar
5) Jumlah spora (per 50 g sampel tanah)
Menurut Gerdemann dan Nicolson (1963), penghitungan jumlah spora dapat dilakukan dengan cara menimbang 50 g tanah, kemudian ditambah air, diaduk, dan didiamkan selama 5 menit sehingga terbentuk suspensi. Setelah itu dituangkan suspensi tersebut ke dalam saringan bertingkat dengan ukuran 710; 410; dan 45 μm. Tanah yang terendap saringan 45 μm ditambahkan aquadest dan sukrosa 60% secukupnya, kemudian disentrifuse selama 5 menit dengan kecepatan 2500 rpm. Supernatan yang diperoleh diambil dan ditampung dengan saringan 45 μm,
14 kemudian dibilas dengan aquades agar sukrosanya hilang, setelah itu ditampung dalam cawan petri. Jumlah spora dapat dilihat dan dihitung dengan mikroskop.
Prosedur Pembuatan inokulum CMA
Persiapan tanaman inang (Pueraria javanica dan Centrosema pubescens) dan media tanam (pasir, tanah, zeolit). Media tanam disterilisasi dan dimasukkan dalam polybag sebanyak 2 kg. Kemudian ditimbang mikofer CMA sebesar 50 g tiap perlakuan. Penanaman dan pemeliharaan dilakukan selama tiga bulan meliputi penyiraman, pemberantasan gulma, dan pemupukan. Setelah tiga bulan pemeliharaan dilakukan stressing yaitu tanaman tidak disiram selama 2 minggu, kemudian baru dipanen (Ariyanti, 2005). Setelah tanaman dipanen tajuknya, sisa media tanamnya digunakan sebagai inokulum CMA pada skala lapang.
Persiapan bahan dan alat
Inokulum CMA ditimbang sebanyak 100 g per pols dengan jumlah tanaman seluruhnya 336 tanaman. Kemudian ditimbang pupuk SP36 dan KCl masing-masing sebanyak 40 g (100 kg/ha) dan urea 80 g per petak (200 kg/ha). Selain itu juga penyediaan pols Brachiaria humidicola sebanyak 336 pols, setiap petak ditanam 16 pols Brachiaria humidicola.
Tahap Pengolahan Tanah
Pengolahan tanah dilakukan sebelum penanaman. Tanah dibajak dan digemburkan dengan traktor dan cangkul disertai pembersihan vegetasi yang dapat mengganggu penanaman. Petakan dibuat dengan luas 2x2 m2 per petak sebanyak 21
petak percobaan. Setelah itu dilakukan pemupukan awal sebanyak 40 g KCl dan SP36 per petak, kemudian tanah disiram. Pupuk urea diberikan 1 minggu setelah tanam.
Tahap Penanaman
Pols rumput Brachiaria humidicola ditanam sedalam ±10 cm dari permukaan tanah, yang sebelumnya telah diberi inokulum cendawan mikoriza arbuskula sebanyak 100 g per pols rumput. Satu pols terdiri atas 2 individu rumput. Setiap petak percobaan ditanam sebanyak 16 pols rumput Brachiaria humidicola.
15 Tahap pemeliharaan
Pemeliharaan dilakukan selama tiga bulan meliputi penyiraman, pemberantasan gulma dan pemupukan dengan urea 1 minggu setelah penanaman, serta pengukuran panjang penyebaran stolon dan perhitungan jumlah stolon yang dilakukan secara intensif selama penelitian setiap minggu.
Tahap Pemanenan
Pemanenan dilaksanakan dua kali yaitu periode I dilakukan setelah tanaman berumur ±60 hari dan periode II dilakukan setelah tanaman berumur ±45 hari. Tanaman inang dipanen dengan menimbang berat segar total yang kemudian diambil sampel sebanyak 100 g per perlakuan dan dikeringkan dalam oven pada suhu 70oC selama 48 jam untuk mendapatkan berat kering tajuk, kemudian dilakukan pengambilan sampel akar untuk mengetahui persentase infeksi akar dan tanah sebanyak 50 g pada akhir penelitian untuk menghitung jumlah spora.
KESIMPULAN DAN SARAN Kesimpulan
Hasil secara keseluruhan menunjukkan tidak ditemukan inokulum cendawan mikoriza arbuskula yang spesifik untuk meningkatkan produktivitas B. humidicola dalam skala lapang. Semua inokulum yang digunakan memberikan efektivitas yang sama dalam peningkatan produksi hijauan pakan ternak. Namun, perlakuan inokulum TCp memberikan pertambahan jumlah stolon terbanyak pada periode I.
Saran
Diperlukan penelitian lanjutan mengenai aplikasi inokulum CMA dalam skala lapang dengan masa penanaman lebih lama dan pemberian dosis pupuk P yang berbeda, serta prosedur penelitian yang lebih teliti sehingga dapat memperkecil pengaruh faktor lingkungan.
UCAPAN TERIMA KASIH
Alhamdulillahi robbil’alamin
Puji syukur kehadirat Allah SWT atas segala limpahan nikmat, kasih sayang, dan ijinNya sehingga penulisan skripsi ini dapat diselesaikan.
Penulis menyadari bahwa skripsi ini tidak akan tercapai tanpa bantuan dan dukungan dari berbagai pihak. Penulis mengucapkan terima kasih kepada Dr. Ir. Panca Dewi M.H.K.S., MSi. dan Prof. Dr. Soedarmadi H., MSc. atas segala bimbingan, semangat, dan ilmu yang diberikan selama penelitian hingga penulisan skripsi. Penulis mengucapkan terima kasih kepada Dr. Ir. Dewi Apri Astuti, MS. sebagai dosen penguji seminar, Ir. Anita S. Tjakradidjaja, MRur.Sc. dan Dr. Ir. Henny Nuraeni, MSi. sebagai dosen penguji sidang. Terima kasih kepada Staf Laboratorium Agrostologi dan Staf Laboratorium Bioteknologi Hutan dan Lingkungan, Institut Pertanian Bogor.
Penulis mengucapkan terima kasih untuk rekan sepenelitian Yanti Fitriyati atas persahabatan, kerjasama, dukungan dan pengertiannya. Widiarti, Tanti P., Arindiyah, Suprayitno, Heri, Fitri serta temen-temen lainnya atas semangat, persahabatan dan kerjasamanya dalam pelaksanaan penelitian. Penulis mengucapkan terima kasih untuk teman-teman di INMT’39, THT’39, TPT’39, SEIP’39, atas keceriaan dan kebersamaannya selama ini.
Kepada Bapak dan Ibu tersayang terima kasih atas kasih sayang, untaian doa, bimbingan, dan kesabaran yang tak pernah putus menyertai perjalanan hidup Penulis. Kepada ketiga saudaraku tersayang Irma Nuranthy Purnama, Pandu Satria Laksono, dan Harum Fadhilatunnur terima kasih atas keceriaan dan hari-hari indah yang telah dijalani selama ini. Terima kasih kepada semua keluarga besar di Pamotan atas doa dan dukungannya kepada Penulis.
Saran dan kritik yang membangun senantiasa Penulis harapkan. Semoga skripsi ini bermanfaat bagi pembaca, terutama bagi peminat dan pemerhati percobaan. Amin.
Bogor, Juni 2006
DAFTAR PUSTAKA
Arifanti, V. B. 1999. Pengaruh pemberian media tumbuh, pupuk NPK dan cendawan endomikoriza Glomus etunicatum terhadap pertumbuhan Tectona grandis L. Skripsi. Fakultas Kehutanan. Institut Pertanian Bogor. Bogor.
Ariyanti, G. J. 2005. Alternatif jenis tanaman inang dan media tanam untuk produksi massal cendawan mikoriza arbuskula (CMA) pada hijauan pakan. Skripsi. Fakultas Peternakan. Institut Pertanian Bogor. Bogor.
Bagyaraj, D. J. 1992. Vesicular-arbuscular mycorrhiza: Application in Agriculture. Dalam: J. R. Norris, D. Read dan A. K. Varma. Techniques for Mycorrhizal Research. p. 819-833. Academic Press, London.
Blasco, L. M. dan A. N. Bohorquez. 1968. Pasture species in the Amazon Region. Analysis of Some Chemical Components. Agricultura Trop. 24:175-177.
Brundrett, M. 1999. Arbuscular Mycorrhizas. http://www.ffp.csiro.au/research/myco rrhiza/vam.html. [2 Februari 2006]
Buckman, H. O., dan N. C. Brady. 1982. Ilmu Tanah. Terjemahan: Soegiman. Bhratara Karya Aksara. Jakarta.
Fakuara, M. Y. 1988. Mikoriza, Teori dan Kegunaan dalam Praktek. Pusat Antar Universitas. Institut Pertanian Bogor. Bogor.
Feniara. 2001. Efektivitas cendawan mikoriza arbuskula (CAM), pupuk P dan N terhadap pertumbuhan dan produksi rumput Gajah (Pennisetum purpureum
Schum). Skripsi. Fakultas Peternakan. Institut Pertanian Bogor. Bogor.
Ferguson, J. J. dan S. H. Woodhead. 1982. Production of endomycorrhizal. A. increase and maintenance of vesicular-arbuscular mycorrhizal fungi. Dalam: Gunawan, A. W. 1993. Mikoriza Arbuskula. Pusat Antar Universitas Ilmu Hayat. Institut Pertanian Bogor. Bogor.
Gai, J. Dan R. Lui. 2003. Effect of soil factors on arbuscular mycorrhizae (AM) fungi around roots of wild plants. Article in Chinese. Ying Yong Sheng Tai Xue Bao. 14(3): 470-472. http: //www.ncbi.nlm.nih.gov./pubmed. [10 Juli 2005].
Gerdemann, J. W., dan J. M. Nicolson. 1963. Spore of mycorrhizal endogone species extracted from soil by wet sieving and decanting. Trans. Brit. Mycol. Soc. 46 : 235-244.
Gerdemann, J. W., dan J. M. Trappe. 1974. The Endogoneceae in The Pacific Northwest. Mycologia Memoir No.5. 76 p.
30 Giovannetti, M., A. Schubert, M. C. Cravero, dan L. Salutini. 1988. Spore production
by the vesicular arbuscular mycorrhizal fungus Glomus monosporum as related to host species, root colonization and plant growth enhancement. Biol. Fert. Soils. 6:120-124.
Gunawan, A. W. 1993. Mikoriza Arbuskula. Pusat Antar Universitas Ilmu Hayat. Institut Pertanian Bogor. Bogor.
Hardjowigeno, S. 1992. Ilmu Tanah. Akademi Pressindo. Jakarta.
Harisuseno.1979. Fisiologi Tumbuhan Dasar. Institut Pertanian Bogor. Bogor.
Hartadi, H., S. Reksohadiprodjo, dan A. D. Tillman. 1993. Tabel Komposisi Pakan untuk Indonesia. Gadjah Mada University Press. Yogyakarta.
Hartmann, H. T., dan D. E. Kester. 1983. Plant Propagation, Principle and Practices. 4th ed. Printice Hall Inc. Englewood Cliffs. New Jersey. 727p.
Howeler, R. H., E. Sieverding dan S. Saif. 1987. Practical aspects of mycorrhizal technology in some tropical crops and pastures. Plant and Soil. 100:249-283. Imas, T., R. S. Hadioetomo, A. W. Gunawan, dan Y. Setiadi. 1989. Mikrobiologi
Tanah II. Pusat Antar Universitas Bioteknologi. Institut Pertanian Bogor. Bogor.
IPB. 2002. Agrostologi. http://www.iel.ipb.ac.id/sac/hibah/2002/agrostologi/ Brachiaria%20humidicola.htm. [7 Januari 2006].
Jarstfer, A. G. dan D. M. Sylvia. 1992. Sheared-root Inocula of VA Mycorrhizal Fungi. Dalam: Gunawan, A. W. 1993. Mikoriza Arbuskula. Pusat Antar Universitas Ilmu Hayat. Institut Pertanian Bogor. Bogor.
Jayadi, S. 1991. Tanaman Makanan Ternak Tropika. Fakultas Peternakan. Institut Pertanian Bogor. Bogor.
Karti, P. D. M. H. 2003. Respon morfofisiologi rumput toleran dan peka alumunium terhadap penambahan mikroorganisme dan pembenah tanah. Disertasi. Program Pasca Sarjana. Institut Pertanian Bogor. Bogor.
Karti, P. D. M. H. 2004. Pengaruh pemberian cendawan mikoriza arbuskula terhadap pertumbuhan dan produksi rumput Setaria splendida stapf yang mengalami cekaman kekeringan. Journal. Vol. 28 No. 2.
Koske, R. E., dan J. H. Gemma. 1989. A modified procedure for staining roots to detect vesicular arbuskular mikoriza. Mycol. Res. 92 (4): 486-505.
Lukiwati, D. R. 1996. Peningkatan produksi dan nilai nutrisi legum pakan dengan pemupukan batuan fosfat dan inokulasi mikoriza vesikular-arbuskular. Disertasi. Program Pasca Sarjana. Institut Pertanian Bogor. Bogor.
31 Maryani, Y. 2000. Pengaruh inokulasi cendawan mikoriza arbuskula terhadap
pertumbuhan dan produksi rumput tropika. Skripsi. Fakultas Peternakan. Institut Pertanian Bogor. Bogor.
Mattjik, A. A. dan I. M. Sumertajaya. 2002. Perancangan Percobaan dengan Aplikasi SAS dan Minitab. Jilid I (Edisi kedua). IPB Press. Bogor.
Mitiku-Habte and R. L. Fox. 1993. Effectiveness of VAM fungi in on sterile soils before and after optimization of P soil solution. Plant and Soil, 151(2):219-226.
Mumpton, F. A. 1984. The role of natural zeolites in agriculture and aquaculture. J. Animal Sci. 3-24.
Philips J. M., dan D. S. Hayman. 1970. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions of the British Mycological Soc 55 : 158-160.
Reksohadiprodjo, S. 1981. Produksi Tanaman Hijauan Makanan Ternak Tropik. Fakultas Peternakan. Universitas Gadjah Mada. Yogyakarta.
Setiadi, Y. 1989. Pemanfaatan Mikro Organisme dalam Kehutanan. Pusat Antar Universitas Bioteknologi. Institut Pertanian Bogor. Bogor.
Setiadi, Y., I. Mansur, S. W. Budi dan Achmad. 1992. Petunjuk Laboratorium: Mikrobiologi Tanah Hutan. Pusat Antar Universitas Bioteknologi. Institut Pertanian Bogor. Bogor.
Sieverding, E. 1991. Vesicular-Arbuscular Mycorrhiza Management in Tropical Agrosystems. Eschborn. Germany.
Smith, S. E. dan D. J. Read. 1997. Mycorrhizal Symbiosis. Academic Press. London. Smith, F. A. dan S. E. Smith. 1995. Nutrient transfer in vesicular-arbuscular
mycorrhizas: A new model based on the distribution of ATP uses on fungal and plant membrans. Biotropia. (8): 1-10.
Solaiman, M. Z. dan H. Hirata. 1995. Effect of indigenous arbuscular mycorrhizal fungi in paddy fields on rice growth and NPK nutrition under different water regimes. Soil Sci. Plant Nutr., 41(3):505-514. Dalam: I. G. M. Subiksa. 2002. Pemanfaatan Mikoriza untuk Penanggulangan Lahan Kritis. http://rudyct.tripod.com/sem2_012/igm_subiksa.htm. [10 Juli 2005].
Sulistyaningsih, E. 2003. Penentuan tanaman inang dan media pertumbuhan yang sesuai untuk perkembangan cendawan mikoriza arbuskula. Skripsi. Fakultas Peternakan. Institut Pertanian Bogor. Bogor.
Sylvia, D. M. 1999. Mycorrhizal Symbioses: Principles and Aplications of Soil Mycrobiology. http://cropsoil.psu.edu/sylvia/mycorrhiza.htm.[22 April 2006].
33 Lampiran 1. Sidik Ragam Pertambahan Jumlah Stolon Periode I
SK db JK KT Fhit F0,05 F0,01
Perlakuan 6 4,791374476 0,798562413 2,88730792* 2,848 4,456
Galat 14 3,872075333 0,27657681
Total 20 8,66344981 0,43317249
Keterangan : * berbeda nyata (p<0,05) Coeff Var = 13,95751
Lampiran 2. Sidik Ragam Pertambahan Jumlah Stolon Periode II
SK db JK KT Fhit F0,05 F0,01 Perlakuan 6 2,385714286 0,397619048 1,02047052 2,848 4,456 Galat 14 5,455 0,389642857 Total 20 7,840714286 0,392035714 Coeff Var = 26,40179
Lampiran 3. Sidik Ragam Pertambahan Panjang Stolon Periode I
SK db JK KT Fhit F0,05 F0,01
Perlakuan 6 28,25030781 4,708384635 1,253110704 2,848 4,456
Galat 14 52,603002 3,757357286
Total 20 80,85330981 4,04266549
Coeff Var = 7,882502
Lampiran 4. Sidik Ragam Pertambahan Panjang Stolon Periode II
SK db JK KT Fhit F0,05 F0,01
Perlakuan 6 8,138095238 1,356349206 0,365557588 2,848 4,456
Galat 14 51,945 3,710357143
Total 20 60,08309524 3,004154762 Coeff Var = 11,66738
34 Lampiran 5. Sidik Ragam Berat Kering Tajuk Periode I
SK db JK KT Fhit F0,05 F0,01 Perlakuan 6 224285,58 37380,93001 0,858429256 2,848 4,456 Galat 14 609640,2426 43545,73161 Total 20 833925,8226 41696,29113 Coeff Var = 31,85769
Lampiran 6. Sidik Ragam Berat Kering Tajuk Periode II
SK db JK KT Fhit F0,05 F0,01 Perlakuan 6 1726465,052 287744,1753 0,696523927 2,848 4,456 Galat 14 5783603,833 413114,5595 Total 20 7510068,885 375503,4442 Coeff Var = 26,11408
Lampiran 7. Hasil Sidik Ragam Pengaruh Perlakuan terhadap Persentase Infeksi Akar Periode I SK db JK KT Fhit F0,05 F0,01 Perlakuan 6 146,1505179 24,35841965 0,57485916 2,848 4,456 Galat 14 593,2198693 42,37284781 Total 20 739,3703872 36,96851936 Coeff Var = 6,87420
Lampiran 8. Hasil Sidik Ragam Pengaruh Perlakuan terhadap Persentase Infeksi Akar Periode II SK db JK KT Fhit F0,05 F0,01 Perlakuan 6 247,2260059 41,20433432 1,65620557 2,848 4,456 Galat 14 348,3025853 24,8787561 Total 20 595,5285912 29,77642956 Coeff Var = 5,192739
35 Lampiran 9. Sidik Ragam Jumlah Spora Periode II
SK db JK KT Fhit F0,05 F0,01
Perlakuan 6 389,1428571 64,85714286 0,927792916 2,848 4,456
Galat 14 978,6666667 69,9047619
Total 20 1367,809524 68,39047619
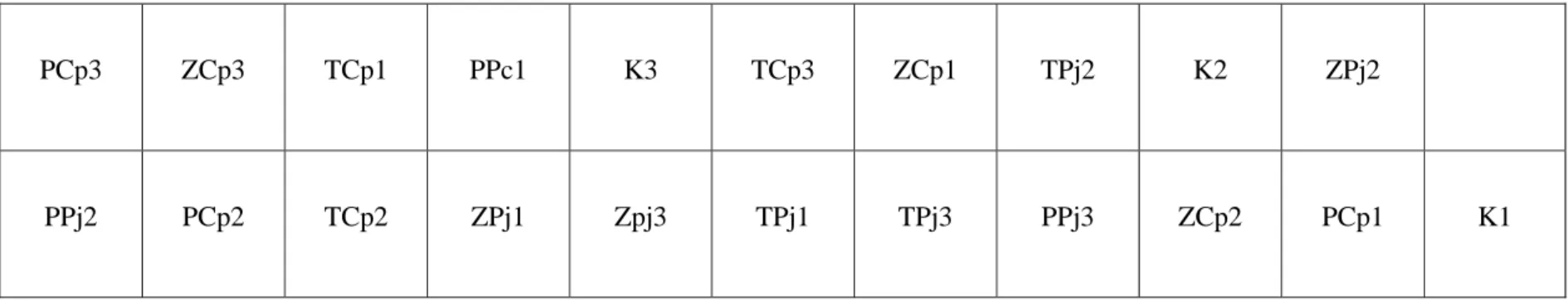
36 Lampiran 10. Bagan Percobaan dan Pengacakan Penelitian
BAGAN PENGACAKAN Penelitian dengan 7 perlakuan 3 ulangan, meliputi;
1. PCp dengan 3 ulangan 2. TCp dengan 3 ulangan 3. ZCp dengan 3 ulangan 4. PPj dengan 3 ulangan 5. TPj dengan 3 ulangan 6. ZPj dengan 3 ulangan 7. Kontrol
Gambar bagan pengacakan:
PCp3 ZCp3 TCp1 PPc1 K3 TCp3 ZCp1 TPj2 K2 ZPj2