TINGKAT KEBERHASILAN PRODUKSI EMBRIO SECARA IN VITRO
MENGGUNAKAN SEMEN BEKU SAPI BALI (Bos javanicus)
DAN ONGOLE (Bos indicus)
G ANDRI HERMAWAN
FAKULTAS KEDOKTERAN HEWAN INSTITUT PERTANIAN BOGOR
BOGOR 2014
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Tingkat Keberhasilan Produksi Embrio secara In Vitro Menggunakan Semen Beku Sapi Bali (Bos javanicus) dan Ongole (Bos indicus) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2014
G Andri Hermawan NIM B04100118
ABSTRAK
G ANDRI HERMAWAN. Tingkat Keberhasilan Produksi Embrio secara In Vitro Menggunakan Semen Beku Sapi Bali (Bos javanicus) dan Ongole (Bos indicus). Dibimbing oleh YANYAN SETIAWAN dan ARIEF BOEDIONO.
Upaya peningkatan produktivitas dan populasi sapi lokal salah satunya adalah produksi embrio metode fertilisasi in vitro, metode ini memanfaatkan oosit ovarium sapi betina dari rumah potong hewan yang difertilisasi menggunakan semen beku sapi bali (Bos javanicus) dan sapi ongole (Bos indicus). Pengamatan dilakukan terhadap pembelahan embrio pada kultur hari ke-2 setelah fertilisasi, kecepatan pembelahan dan jumlah total blastosis yang terbentuk pada kultur hari ke-6, 7, 8, dan 9. Spermatozoa sapi bali lebih baik (p<0.05) dibandingkan sapi ongole dalam memfertilisasi dan mendukung kecepatan pembelahan embrio, serta total blastosis yang terbentuk. Embrio yang membelah pada hari ke-2 dari oosit yang difertilisasi semen sapi bali (39.40%) lebih tinggi dari embrio yang difertilisasi semen sapi ongole (25.22%). Tingkat perkembangan embrio tahap blastosis juga lebih tinggi pada embrio dari oosit yang difertilisasi semen sapi bali (47.05%) dari pada embrio yang berasal dari oosit yang difertilisasi semen sapi ongole (22.09%). Berdasarkan hal tersebut, spermatozoa sapi bali lebih baik digunakan dalam produksi embrio secara in vitro dari pada spermatozoa sapi ongole.
Kata kunci: produksi embrio in vitro, semen sapi bali, sapi ongole
ABSTRACT
G ANDRI HERMAWAN. The Successfulness of Embryo Production by In Vitro Fertilization Using Frozen Semen of Bali Cattle (Bos javanicus) and Ongole (Bos indicus). Supervised by YANYAN SETIAWAN and ARIEF BOEDIONO.
The one solution for increasing the population and productivity of local cattle is the introduction of embryo biotechnology especially in vitro embryo production (IVP). The aims of this study was to evaluate the embryo production rates of collected oocytes from slaughterhouse fertilized using frozen semen of bali cattle (Bos javanicus) and ongole cattle (Bos indicus). The cleavage rate was evaluated on day 2 after fertilization and blastocyst formation rate was on day 6, 7, 8, and 9, as well as an evaluation of the total bastocyst formed. The spermatozoa of bali cattle have a better ability (p<0.05) than ongole cattle at cleavage rate, blastocyst formation rate, and total of blastocyst formation. The cleavage rate of oocytes fertilized by semen of bali cattle (39.40%) was higher than by semen of ongole cattle (25.22%). The development of the embryo in to the blastocyst rate appeared to be better and rapid on oocytes which were fertilized by semen of bali cattle than by semen of ongole cattle. The total blastocyst formed up to day 9 was higher when the oocytes was fertilized by semen of bali cattle (47.05%) than by semen of ongole cattle (22.09%). These study concluded that the using frozen semen of bali cattle for in vitro embryo production was better than frozen of ongole cattle.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Kedokteran Hewan
pada
Fakultas Kedokteran Hewan Institut Pertanian Bogor
G ANDRI HERMAWAN
FAKULTAS KEDOKTERAN HEWAN INSTITUT PERTANIAN BOGOR
BOGOR 2014
TINGKAT KEBERHASILAN PRODUKSI EMBRIO SECARA IN VITRO
MENGGUNAKAN SEMEN BEKU SAPI BALI (Bos javanicus)
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wata’ala atas segala karunia-Nya sehingga skripsi ini berhasil diselesaikan. Judul skripsi ini ialah Tingkat Keberhasilan Produksi Embrio secara In Vitro Menggunakan Semen Beku Sapi Bali (Bos javanicus) dan Ongole (Bos indicus).
Terima kasih penulis ucapkan kepada ayah tercinta Iwik Wikana, ibu tercinta Munah, kakak tercinta Lilis Y. Sopyah dan Wiwit Juwita serta segenap keluarga atas kasih sayang tiada henti. Ucapan terima kasih juga diucapkan kepada Prof Drh Arief Boediono, PhD PAVet (K) dan Bapak Yanyan Setiawan, SPt MSi selaku pembimbing dan Ir Tri Harsi, MP selaku kepala Balai Embrio Ternak (BET) Cipelang beserta stafnya yaitu Ibu Laelatul Choiriyah, AMd, Bapak Fahrudin Darlian, SPt, dan Drh Nurwidayati yang membantu selama penelitian. Tidak lupa ucapan terima kasih kepada teman satu penelitian yaitu Muhammad Faris Firdaus, Alif Iman Fitrianto, Anisa Hasby Fauzia, dan Ika Septiana Anggun Puspita yang telah membantu selama penelitian, tidak lupa kepada Drh Vetnizah Juniantito, PhD APVet selaku dosen pembimbing akademik, Drh Amrozi, PhD beserta teman seperjuangan di SRC IPB, himpro ornithologi dan unggas, dan paguyuban beasiswa karya salemba empat IPB. Semoga skripsi ini bermanfaat baik bagi penulis maupun pembaca.
Bogor, Agustus 2014
DAFTAR ISI
DAFTAR TABEL ... xii
DAFTAR GAMBAR ... xii
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan ... 2
METODE ... 2
Lokasi dan Waktu ... 2
Analisis Data ... 4
HASIL DAN PEMBAHASAN ... 4
Standar Operasional Produksi Embrio In Vitro ... 4
Tingkat Perkembangan Embrio In Vitro ... 5
SIMPULAN DAN SARAN ... 8
Simpulan ... 8
Saran ... 8
DAFTAR PUSTAKA ... 9
DAFTAR TABEL
1 Tingkat perkembangan embrio tahap pembelahan pada kultur hari ke-2 setelah fertilisasi ... 5 2 Tingkat perkembangan embrio tahap blastosis pada kultur hari
ke-6, ke-7, ke-8, dan ke-9 setelah fertilisasi ... 6 3 Total embrio tahap blastosis yang terbentuk pada kultur hari ke-6,
ke-7, ke-8, dan ke-9 setelah fertilisasi ... 7
DAFTAR GAMBAR
1 Berbagai kualitas oosit hasil aspirasi ... 3 2 Perkembangan embrio secara in vitro pada sapi ... 8 1
PENDAHULUAN
Latar BelakangPeningkatan jumlah penduduk Indonesia meyebabkan permintaan daging sapi di pasaran meningkat pula. Tingginya permintaan tersebut tidak diimbangi dengan pasokan sapi potong nasional yang mencukupi, sehingga harga daging sapi di pasaran terus meningkat. Solusi yang dilakukan pemerintah adalah dengan melakukan impor, tahun 2013 saja pemerintah mengimpor sapi bakalan sebanyak 267.000 ekor dan daging sapi beku sebanyak 32.000 ton (Syaputri 2013). Impor yang dilakukan dalam jangka pendek cukup efektif memenuhi permintaan pasar, tetapi jika impor dilakukan dalam jangka panjang, maka Indonesia akan ketergantungan kepada negara lain dan semakin menurunnya peternakan sapi lokal di Indonesia (Diwyanto 2002).
Solusi yang tepat untuk memenuhi tingginya permintaan akan daging sapi di pasaran yaitu dengan cara meningkatkan populasi dan produktivitas sapi potong lokal. Sapi potong lokal tersebut sebagai pasokan sapi nasional untuk memenuhi permintaan daging sapi di pasaran (Sondy et al. 2009). Peningkatan produktivitas sapi lokal salah satunya bisa dilakukan pada sapi bali (Bos javanicus). Sapi bali merupakan plasma nutfah Indonesia yang didomestikasi dari banteng liar sekitar tahun 3.500 SM (Rollinsonn 1984). Penyebaran sapi bali cukup luas karena sapi ini memiliki potensi genetik dan ekonomi yang unggul jika dikembangkan sebagai sapi potong (Sobari et al. 2012). Sapi bali dikatakan unggul karena memiliki tingkat fertilitas tinggi, daya adaptasi baik, dapat memanfaatkan hijauan yang kurang bergizi, dan memiliki persentase karkas tinggi yaitu mencapai 52-57.7% (Handiwirawan dan Subandriyo 2004; Samberyet al. 2010). Peningkatan populasi dan produktivitas juga bisa dilakukan pada sapi peranakan ongole (PO) (Bos javanicus). Sapi PO merupakan sapi hasil persilangan pejantan sapi ongole (Bos indicus) dengan sapi jawa betina putih. Sapi PO banyak dipelihara karena bukan hanya sebagai sapi potong saja, tetapi juga sebagai sapi pekerja penarik gerobak dan pembajak sawah (Diwyanto 2002). Keunggulan yang dimiliki sapi PO yaitu memiliki tingkat adaptasi yang baik yaitu tahan terhadap panas, tahan terhadap ektoparasit dan endoparasit, pertumbuhan relatif cepat, dan kualitas daging baik (Hartati et al. 2005).
Prinsip bioteknologi fertilisasi in vitro (FIV) sendiri adalah pembuahan sel telur oleh spermatozoa di luar tubuh hewan betina yang direkayasa untuk memproduksi embrio dalam jumlah banyak, dan dapat meningkatkan mutu genetik (Vicanco dan Mackie 2001). Aplikasi produksi embrio secara in vitro bisa dilakukan pada sapi bali dan PO dengan memanfaatkan ovarium sapi betina yang terlanjur dipotong di rumah potong hewan (RPH). Ovarium sapi sapi betina dari RPH masih bisa dimanfaatkan karena bagian korteks ovarium merupakan tempat perkembangan oosit yang merupakan salah satu bahan dalam produksi embrio secara in vitro (Kaiin et al. 2008; Pujo et al. 2004). Metode FIV dalam produksi embrio mulai dari koleksi dan maturasi oosit, preparasi dan kapasitasi spermatozoa, fertilisasi dan kultur embrio secara in vitro (Susilowati et al. 1998). Adanya pasokan embrio kedua bangsa sapi tersebut bisa membantu mewujudkan cita-cita Indonesia menjadi negara yang swasembada daging.
2
Tujuan
Penyusunan skripsi ini untuk mengetahui tingkat fertilitas spermatozoa sapi bali dan ongole secara in vitro, dan juga mengetahui tingkat keberhasilan pembentukan embrio tahap pembelahan dan tahap blastosis dari oosit yang difertilisasi semen sapi bali dan ongole secara in vitro.
METODE
Lokasi dan WaktuPenelitian ini dilaksanakan di laboratorium produksi embrio Balai Embrio Ternak (BET) Cipelang Bogor Kementerian Pertanian Republik Indonesia pada bulan Juli 2013 dan Februari 2014. Penelitian meliputi kegiatan produksi embrio secara in vitro dan pengambilan data skunder produksi embrio secara in vitro pada tahun 2011 dan 2012.
Produksi embrio secara in vitro merupakan rangkaian proses biologi yang berkaitan satu sama lain. Rangkaian tersebut meliputi koleksi ovarium dan oosit, maturasi oosit secara in vitro, kapasitasi spermatozoa, fertilisasi dan kultur embrio secara in vitro.
Koleksi Ovarium dan Oosit
Ovarium yang berasal dari sapi betina yang tidak diperhatikan bangsanya (biasanya sapi PO, sapi madura, dan sapi persilangan brahman) yang terlanjur dipotong di rumah potong hewan. Ovarium dibawa ke laboratorium in vitro di dalam termos berisi ringer laktat yang disuplementasi antibitoik. Antibiotik tersebut yaitu streptomycin (Sigma Cat.#S1277) 100 mg/L dan penicillin (Sigma Cat.#4687) 100.000 IU/L dengan suhu ±37.5 °C, dengan waktu transportasi tidak lebih dari 5 jam (Boediono et al. 2000). Folikel ovarium (kecuali folikel de Graaf) diaspirasi menggunakan spuit 5 mL dengan needle 18-G, media yang digunakan untuk aspirasi adalah phosphate buffered saline (PBS, Sigma) yang disuplementasi dengan fetal calf serum (FCS, Sigma Cat. #F 2442) sebanyak 3%, streptomycin 100 mg/L dan penicillin 100.000 IU/L. Oosit hasil aspirasi diletakkan di cawan petri bergaris ukuran 90x90x10 mm untuk memudahkan pencarian oosit menjelang maturasi.
Maturasi Oosit
Oosit hasil aspirasi dari cawan petri bergaris dipindahkan ke cawan petri bulat berdiameter 35 mm yang berisi tissue culture medium-199 (TCM-199; Gibco Cat.#11150-042; Sigma Chemical Co., USA) yang disuplementasi FCS 10%, streptomycin 100 mg/L dan penicillin 100.000 IU/L. Oosit kemudian dicuci dengan TCM-199 dan dikelompokan menjadi 4 kelompok, yaitu kualitas A, B, C, dan D. Pengelompokan tersebut berdasarkan ada tidaknya cumulus oophorus dan kualitas ooplasma oosit. Menurut Sobari et al. (2012) oosit kualitas A adalah oosit dengan cumulus oophorus tebal berlapis dan ooplasma homogen, oosit kualitas B adalah oosit dengan cumulus oophorus tipis dan ooplasma homogen, oosit kualitas C adalah oosit dengan ooplasma homogen tetapi tidak terdapat cumulus
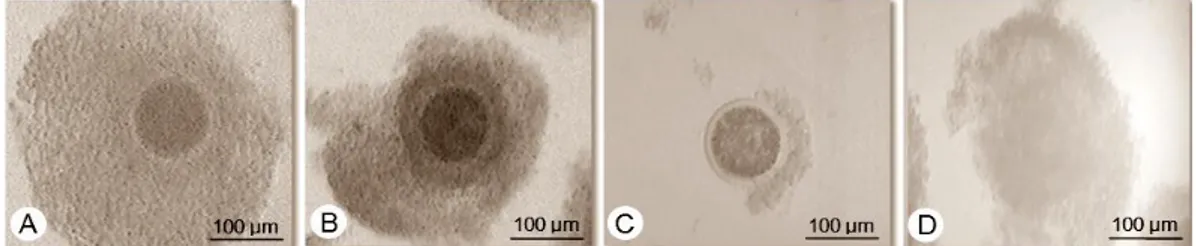
3 oophorus, oosit kualitas D adalah oosit dengan ooplasma tidak homogen, cumulus oophorus expand dan dikelilingi granul fibrin (Gambar 1).
Gambar 1 Kualitas oosit hasil koleksi dari ovarium: A) Oosit A, memiliki ooplasma yang homogen dan adanya sel-sel cumulus oophorus yang mengelilingi oosit secara penuh; B) Oosit B; memiliki ooplasma homogen, namun hanya sebagian oosit yang dikelilingi sel-sel cumulus oophorus; C) Oosit C, memiliki ooplasma homogen dan tidak terdapat sel-sel cumulus oophorus yang mengelilingi oosit; D) Oosit D, memiliki ooplasma yang tidak homogen dan sel-sel cumulus
oophorus telah menyebar atau expand. (Dokumentasi Balai Embrio Ternak).
Oosit kualitas A dan B dimaturasi selama 18-24 jam dalam media TCM-199 yang disuplementasi FCS 10%, streptomycin 100 mg/L dan penicillin 100.000 IU/L serta diequilibrasi di dalam inkubator CO2 dengan kadar CO2 5%, suhu
38.5°C, dan kelembaban 90%. Tiap drop (200 µL) berisi 40 oosit yang ditutup dengan mineral oil (Sigma Cat.#M.8410) untuk mencegah penguapan media dan menjaga kestabilan pH media.
Kapasitasi Spermatozoa
Langkah pertama sebelum fertilisasi adalah pembuatan drop fertilisasi. Drop fertilisasi dibuat di dalam cawan petri yang berisi oocyte washing solution (OWS) sebanyak 5µL yang merupakan campuran brackett oliphant (BO) solution 40 mL dan bovine serum albumin (BSA, Sigma Cat. #A8806) sebanyak 400 mg, drop tersebut ditutup mineral oil. Kapasitasi sepermatozoa dimulai dengan thawing semen beku dengan air suhu 37oC, kemudian dimasukan ke dalam 6 mL semen washing solution (SWS) yang merupakan campuran dari BO solution, Na caffeine benzoate 0.942g dan heparin (Sigma Cat. #D4776) sebanyak 50µL. Campuran semen dan SWS selanjutnya disentrifugasi dengan kecepatan 1.800 rpm, suhu 38°C selama 5 menit kemudian dibuang SWS tanpa membuang endapannya dan diisi kembali SWS baru dan dilakukan sentrifugase kembali. Buang kembali SWS dan ambil sebanyak 50 µL endapan semen dan masukan ke dalam 4950 µL NaCl 3%, lalu simpan di neubaur chamber dan amati motilitas, abnormalitas dan konsentrasi spermatozoa sebagai acuan dalam pengenceran.
Pengenceran endapan dengan semen dilution solution (SDS) yang merupakan campuran dari BO solution 10 mL dan BSA 200 mg, pengenceran dilakukan hingga konsentrasi spermatozoa sebanyak 12.5x106/mL. Endapan semen yang telah diencerkan dengan SDS selanjutnya diambil sebanyak 95 µL dan dimasukan ke dalam drop fertilisasi OWS yang sebelumnya telah dibuat, dengan demikian volume tiap drop berisi 100 µL. Media fertilisasi yang telah dibuat selanjutnya diinkubasi selama 1 jam dalam inkubator CO2.
Fertilisasi dan Kultur Embrio In Vitro
Oosit yang telah matur dan terseleksi dicuci media OWS, kemudian oosit tersebut dipindahkan ke dalam drop fertilisasi. Tiap drop diisi 20-25 oosit dengan
4
proses fertilisasi dilakukan selama 5 jam di dalam inkubator CO2. Oosit yang telah difertilisasi selama 5 jam dicuci dengan charles rosenkrans (CR) 1aa yang telah disuplementasi FCS 10%. pencucian dilakukan untuk menghilangkan spermatozoa yang menempel pada zona pelusida. Selanjutnya oosit yang telah difertilisasi disebut sebagai zigot, selanjutnya zigot dikultur menggunakan media CR1aa yang dilapisi mineral oil, kultur dilakukan di dalam inkubator CO2. Hari saat dilakukan fertilisasi dihitung sebagai hari ke-0, hari ke-2 kultur dilakukan pengamatan dan penghitungan embrio yang membelah, dan pengamatan serta penghitungan embrio tahap expanded blastosis yang terbentuk pada kultur hari ke-6, ke-7, ke-8, dan ke-9.
Analisis Data
Peubah yang diamati adalah embrio tahap pembelahan yang terbentuk pada hari ke-2 setelah fertilisasi, dan jumlah embrio tahap blastosis yang terbentuk pada hari ke-6, ke-7, ke-8, dan ke-9 setelah fertilisasi. Tingkat pembelahan embrio dihitung dari jumlah embrio yang membelah terhadap oosit yang difertilisasi, dan tingkat pembentukan blastosis dihitung dari jumlah blastosis yang terbentuk terhadap embrio yang membelah pada hari ke-2.
Data pembentukan blastosis diolah dengan analisis data chi-kuadrat. Menurut Gaspersz (1991) chi-kuadrat disebut dengan istilah uji Khi-kuadrat (Chi-square test), yaitu data hasil penelitian merupakan nilai frekuensi pengamatan yang diperoleh bukan melalui hasil pengukuran yang menggunakan alat ukur tertentu, melainkan dihitung.
HASIL DAN PEMBAHASAN
Standar Operasional Produksi Embrio In VitroTahapan penting sebelum fertilisasi yaitu preparasi oosit dan spermatozoa, preparasi oosit dan spermatozoa sangat penting agar proses fertilisasi dapat terjadi. Preparasi oosit yaitu maturasi oosit, langkah ini penting karena salah satu syarat keberhasilan fertilisasi adalah oosit yang matang, yaitu oosit yang sudah mencapai tahap metafase II. Menurut Boediono et al. (2000) fertilisasi hanya bisa terjadi pada oosit yang telah mencapai tahap metafase II. Preparasi spermatozoa meliputi pencucian spermatozoa dari pengencer dan kapasitasi spermatozoa. Kapasitasi spermatozoa secara in vitro dilakukan untuk meningkatkan motilitas dan kemampuan spermatozoa dalam pelepasan tudung akrosom dan asam hyaluronidase. Fungsi asam hyaluronidase untuk memudahkan spermatozoa menembus zona pelusida dan membran vitelin oosit.
Tahapan maturasi oosit, kapasitasi sepermatozoa, fertilisasi in vitro, dan kultur embrio dilakukan di dalam inkubator CO2 dengan kadar CO2 5%.Kadar tersebut berguna untuk menjaga pH media tetap stabil. Kondisi pH yang terlalu tinggi atau rendah akan menyebabkan kestabilan media terganggu, dan bersifat toksik terhadap embrio. Suhu inkubator selama maturasi adalah 38.5oC, suhu tersebut merupakan suhu yang optimal untuk maturasi oosit dan kultur embrio (Lequaere et al. 2003). Waktu maturasi oosit dilakukan selama 18-24 jam, waktu
5 tersebut berguna untuk mencegah oosit membelah secara partenogenesis (Sirard dan Blodin 1996).
Media yang digunakan di setiap tahap produksi embrio in vitro berbeda-beda. Media maturasi oosit yang digunakan adalah TCM-199, media ini memiliki kemampuan yang baik untuk pematangan oosit secara in vitro, dan merupakan standar media yang digunakan untuk pematangan oosit kambing dan sapi (Takahashi et al. 1996).
Kapasitasi spermatozoa dilakukan di dalam media SWS, media tersebut merupakan BO solution yang ditambah heparin dan kafein yang berfungsi untuk kapasitasi spermatozoa. Menurut Im et al. (1995) BO solution yang ditambah heparin dan kafein berguna sebagai agen kapasitasi spermatozoa agar dapat memfertilisasi oosit. Media kultur embrio adalah CR1aa, media tersebut merupakan semi defined medium yang telah digunakan untuk embrio pada sapi (Rosenkrans et al. 1993; Rosenkrans dan First 1994). Media CR1aa mengandung media minimum dasar penting yang merupakan campuran dari asam amino, vitamin, garam-garam anorganik, ribonucleosides dan deoxyribonucleosides (Sagirkaya et al. 2004).
Media yang digunakan untuk maturasi, fertilisasi, dan kultur embrio ditambah FCS, fungsi FCS sendiri merupakan faktor pertumbuhan yang berfungsi mendukung pematangan oosit dan perkembangan embrio dengan cara
meningkatkan proliferasi dan diferensiasi sel (Margawati 1999). Selain ditambah FCS, media juga ditambah antibotik. Antibiotik yang ditambahakan yaitu
penicillin dan streptomycin yang berguna untuk mencegah kontaminasi mikroorganisme di dalam media. Kontaminasi mikroorganisme yang akan mengganggu perkembangan oosit dan embrio dengan cara menghabiskan nutrisi yang dibutuhkan oleh oosit dan embrio, dan menghasilkan sisa-sisa metabolisme yang dapat meracuni oosit dan embrio.
Tingkat Perkembangan Embrio In Vitro
Pengamatan tingkat keberhasilan produksi embrio in vitro meliputi tingkat perkembangan embrio tahap pembelahan, tahap blastosis, dan total blastosis yang terbentuk secara in vitro antara embrio dari oosit yang diferilisasi semen sapi bali dan ongole.
Perkembangan Embrio Tahap Pembelahan
Kultur embrio hari ke-2 dilakukan pengamatan dan penghitungan embrio yang membelah (Gambar 2). Embrio yang membelah merupakan gambaran tingkat fertilitas spermatozoa yang digunakan untuk memfertilisasi (Tabel 1). Tabel 1 Tingkat perkembangan embrio tahap pembelahan pada kultur hari ke-2
setelah fertilisasi
Sapi sumber permatozoa Jumlah oosit matur yang
difertilisasi
n
Jumlah embrio yang membelah hari ke-2
n (%±SB)
Sapi ongole 341 86 (25.22 ± 3.90)a
Sapi bali 302 119 (39.40 ± 7.43)b
6
Tingkat pembelahan embrio hari ke-2 dari oosit yang difertilisasi semen sapi bali lebih baik (p<0.05) dari embrio yang difertilisasi semen sapi ongole (Tabel 1). Pembelahan embrio hasil fertilisasi in vitro dipengaruhi oleh media yang digunakan, maturasi oosit, dan kemampuan spermatozoa dalam memfertilisasi (Syaiful et al. 2010). Perlakuan oosit, semen, dan kultur embrio antara keduanya sama saja, sehingga tingkat keberhasilan fertilisasi dan perkembangan embrio hanya dipengaruhi oleh faktor yang dibawa spermatozoa. Faktor yang dibawa spermatozoa meliputi motilitas, daya hidup, dan faktor genetik yang memengaruhi fertilisasi dan pembelahan embrio.
Adanya perbedaan terhadap tingkat fertilitas dan pembentukan embrio tahap pembelahan antara keduanya karena faktor fertilitas pejantan sumber spermatozoa. Sapi bali merupakan sapi asli Indonesia yang memiliki tingkat fertilitas dan produktivitas yang tinggi (Purwantara et al. 2012). Tingginya tingkat fertilitas sapi bali karena sapi bali memiliki tingkat adaptasi yang baik (Handiwirawan dan Subandriyo 2004), bahkan tingkat fertilitas spermatozoa sapi bali yang dikoleksi dari kauda epididimis pun masih lebih tinggi dibanding sapi lain (Labetubun dan Siwa 2011). Tingkat fertilitas spermatozoa sapi bali tinggi karena tingkat abnormalitas spermatozoa sapi bali hanya 9.38%, tidak mencapai 18-20%. Tingkat abnormalitas sendiri berbanding terbalik dengan tingkat fertilitas (Barth dan Oko 1989).
Mekanisme spermatozoa memengaruhi tingkat keberhasilan perkembangan embrio tahap pembelahan dengan cara mempercepat renovasi kromatin saat pembentukan pronukleus jantan. Pronukleus jantan sangat penting, karena akan bersatu dengan pronukleus betina (synggami) dan terjadilah penyatuan materi DNA menjadi diploid, dan segera terjadilah sintesis DNA sebagai fase sebelum pembelahan mitosis (Boediono et al. 1997; Ward et al. 2001).
Rendahnya tingkat fertilitas spermatozoa sapi ongole secara in vitro diduga spermatozoa mengalami kegagalan kondensasi dalam sitoplasma oosit. Kegagalan kondensasi spermatozoa dalam sitoplasma oosit menyebabkan gagalnya pembentukan pronukleus jantan (Boediono et al. 2000). Kegagalan kondensasi bisa disebabkan kurang matangnya oosit yang akan difertilisasi. Salah satu syarat keberhasilan fertilisasi in vitro dipengaruhi tingkat maturasi oosit (Syaiful et al. 2010).
Perkembangan Embrio Tahap Blastosis
Kultur embrio pada hari ke-6 hingga hari ke-9 setelah fertilisasi dilakukan pengamatan dan penghitungan expand blastosis yang terbentuk (Gambar 2).
Tabel 2 Tingkat perkembangan embrio tahap blastosis pada kultur hari ke-6, ke-7, ke-8, dan ke-9 setelah fertilisasi
Sapi sumber permatozoa Jumlah embrio membelah hari ke-2 n
Jumah dan persentase blastosis yang terbentuk*
Hari ke-6 Hari ke-7 Hari ke-8 Hari ke-9
n (%±SB) n (%±SB) n (%±SB) n (%±SB)
Sapi ongole 86 3(3.49±8.94)a 11(12.79±8.82)a 3(3.49±6.78)a 2(2.33±3.58)a
Sapi bali 119 15(12.61±9.67)b 22(18.49±7.68)a 11(11.76±5.67)b 8(6.72±4.78)a
Keterangan : Superskrip yang berbeda pada kolom yang sama menunjukan berbeda nyata (p<0.05). * Persentase blastosis diperoleh dari jumlah embrio membelah pada hari ke-2.
7 Tingkat perkembangan embrio tahap blastosis dari oosit yang difertilisasi semen sapi bali lebih baik (p<0.05) dari embrio tahap blastosis yang difertilisasi semen sapi ongole (Tabel 2). Perlakuan kultur embrio antara embrio yang berasal dari oosit yang difertilisasi semen sapi bali dan ongole sama saja, sehingga perkembangan embrio mencapai blastosis hanya dipengaruhi oleh spermatozoa yang digunakan untuk memfertilisasi. Perbedaan kecepetan dan jumlah pembentukan embrio tahap blastosis antara antara kedua blastosis tersebut terjadi pada kultur hari ke-6 dan ke-8 setelah fertilisasi. Menurut Comizzoli et al. (2000) kecepatan perkembangan embrio mencapai tahap blastosis dipengaruhi faktor pejantan, sedangkan faktor maternal atau yang dibawa induk tidak begitu berpengaruh. Mekanisme bangsa pejantan mempercepat perkembangan embrio mencapai tahap blastosis melalui faktor genetik dalam kromosom yang dibawa spermatozoa (Sumantri et al. 1997). Spermatozoa sapi bali yang digunakan untuk memfertilisasi memengaruhi proses renovasi kromatin, pembentukan pronukleus jantan, sintesis DNA, dan pembelahan mitosis pada embrio yang pada akhirnya berkorelasi dengan jumlah embrio yang membelah mencapai blastosis. Mekanisme bangsa pejantan dalam mempercepat perkembangan embrio mencapai blastosis dengan cara memengaruhi waktu dan durasi replikasi DNA sebelum membelah secara mitosis, sehingga siklus pembelahan mitosis lebih cepat terjadi (Comizzoli et al. 2000).
Puncak pembentukan embrio tahap blastosis dari oosit baik yang difertilisasi semen sapi bali dan ongole terjadi pada kultur hari ke-7 setelah fertilisasi. Puncak pembentukan blastosis tersebut sesuai dengan penelitian Ward et al. (2001) yaitu puncak perkembangan embrio tahap blastosis terjadi pada kultur hari ke-7. Blastosis yang dihasilkan merupakan blastosis hybrid, karena ovarium sebagai sumber oosit diambil secara acak tanpa memperhatikan bangsa sapi sumber ovariumnya. Berdasarkan hal tersebut, kecepatan dan puncak pembentukan blastosis yang difertilisasi semen sapi bali dan ongole sudah sesuai dengan penelitian Boediono et al. (2003) yaitu kecepatan pembelahan embrio mencapai blastosis hybrid terjadi pada kultur hari ke-7 setelah fertilisasi. Kecepatan pembelahan embrio membentuk blastosis ada kaitannya dengan kromosom seks blastosis. Blastosis yang cepat terbentuk diduga merupakan blastosis dengan kromosom seks XY (jantan). Menurut Boediono et al. (1998), blastosis dengan kromosom seks XY (jantan) hasil sexing spermatozoa lebih cepat berkembang menjadi blastosis dibandingkan blastosis dengan kromosom seks XX (betina). Total Embrio Tahap Blastosis yang Terbentuk
Total embrio tahap blastosis didapatkan dari total expand blastosis yang terbentuk pada kultur hari ke-6 hingga hari ke-9 (Tabel 3).
Tabel 3 Total embrio tahap blastosis yang terbentuk pada kultur hari ke-6, ke-7, ke-8, dan ke-9 setelah fertilisasi
Sapi sumber spermatozoa
Jumlah embrio yang membelah hari ke-2
N
Jumlah total blastosis yang terbentuk
n (%±SB)
Sapi ongole 86 19(22.09±4.84)a
Sapi bali 119 56(47.05±16.59)b
8
Total embrio tahap blastosis yang terbentuk dari oosit yang difertilisasi semen sapi bali lebih baik (p<0.05) dan lebih tinggi dari pada blastosis yang berasal dari oosit yang difertilisasi semen sapi ongole (Table 3). Tinginya jumlah blastosis pada sapi bali yang terbentuk dipengaruhi oleh kecepatan perkembangan embrio mencapai blastosis. Embrio hasil fertilisasi spermatozoa sapi bali lebih cepat membelah mencapai blastosis dibandingkan yang difertilisasi spermatozoa sapi ongole (Tabel 2), sehingga jumlah total embrio tahap blastosis yang terbentuk pun lebih tinggi. Korelasi antara kecepatan pembelahan dan jumlah blastosis yang terbentuk sesuai dengan penelitian Lonergan et al. (1999) yaitu jumlah total blastosis yang terbentuk lebih tinggi dari embrio yang cepat membelah dibandingkan dari embrio yang lambat membelah.
Tingginya jumlah blastosis pada sapi bali karena faktor genetik yang dibawa spermatozoa Sali bali mudah beradaptasi dengan lingkungan kultur embrio in vitro. Hal tersebut sesuai dengan Sumantri et al. (1997) dan Gandolfi (2004), yaitu faktor pejantan meningkatkan kecepatan dan keberhasilan pembentukan blastosis dengan mekanisme memperbaiki interaksi antara faktor genetik pada kromosom yang dibawa bangsa pejantan dengan lingkungannya baik secara in vitro maupun in vivo. Faktor genetik yang dibawa spermatozoa meningatkan kecepatan pembelahan embrio, jumlah embrio, dan kualitas embrio yang terbentuk (Nedambale et al. 2006).
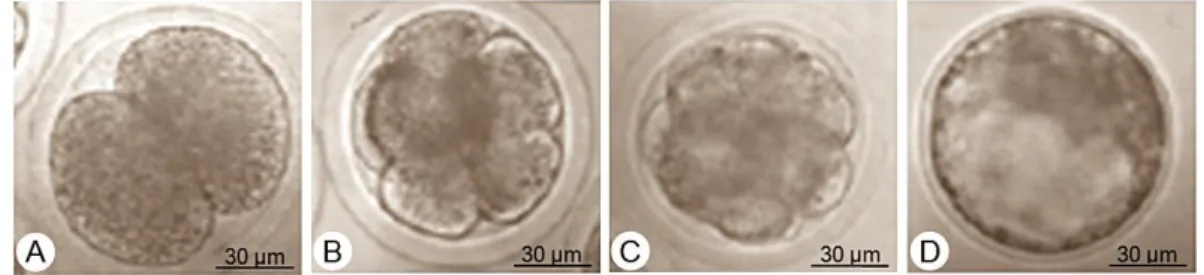
Berikut embrio yang membelah pada kultur hari ke-2 setelah fertilisasi, dan embrio tahap blastosis yang terbentuk pada kultur hari ke-6 hingga ke-9 setelah fertilisasi (Gambar 2).
Gambar 2 Perkembangan embrio hasil fertilisasi in vitro: A) Embrio tahap pembelahan 2 sel; B) Pembelahan >4 sel; C) Embrio tahap morula; D) Embrio tahap expand blastosis. (Dokumentasi Balai Embrio Ternak).
SIMPULAN DAN SARAN
SimpulanKemampuan spermatozoa sapi bali lebih baik dibandingkan spermatozoa sapi ongole dalam melakukan fertilisasi, mendukung perkembangan embrio tahap pembelahan dan blastosis secara in vitro.
Saran
Produksi embrio secara in vitro yang memanfaatkan oosit dari ovarium rumah potong hewan lebih baik difertilisasi menggunakan semen sapi bali dibandingkan semen sapi ongole.
9
DAFTAR PUSTAKA
Barth AD, Oko RJ. 1989. Abnormal Morphology of Bovine Spermatozoa. Iowa (US): State University Pr.
Boediono A, Rusiyantono Y, Mohamad K, Djuwita I, Harliatien. 2000. Developmental competence of caprine oocyte after in vitro maturation, fertilization and culture. Media Veteriner. 7 (4): 11-17.
Boediono A, Tappa B. Saili T, Teliehere MR. 1998. Pengendalian jenis kelamin anak melalui sexing spermatozoa untuk reproduksi ternak. Warta biotek. 12: 1-5.
Boediono A, Ooe M, Murakami M, Saha S. 1997. The effect of sperm-oocyte incubation time on in vitro embryo development using sperm from a tetraparental chimeric bull. J Anim Reprod Sci. 48(2): 187-195.
Boediono A, Suzuki T, Godke R. 2003. Comparison of hybrid and purebred in vitro-derived cattle embryos during in vitro culture. J Anim Reprod Sci. 78:1-11.
Comizzoli, Marquant-Le, Guienne, Heyman, Renard. 2000. Onset the first S-phase is determined by a parental effect during the G1-S-phase in bovine zigotes. J Biol Reprod. 62: 1677-1684.
Diwyanto K. 2002. Pemanfaatan sumber daya lokal dan inovasi teknologi dalam mendukung perkembangan sapi potong di indonesia. Jurnal Pengembangan inovasi pertanian. 1(2): 173-188.
Gandolfi F. 2004. Autocrine, paracrine, and environmental factors influencing embryonic development from zygote to blastocyst. Theriogenology. 41:95-100.
Gaspersz V. 1991. Teknik Analisis dalam Penelitian Percobaan. Bandung (ID): Tarsito Bandung.
Handiwirawan E, Subandriyo. 2004. Potensi dan keragaman sumberdaya genetik sapi bali. Wartazoa. 14(3): 107-115.
Hartati, Mariyono, Wijono DB. 2005. Respons Pertumbuhan Sapi Peranakan Ongole dan Silangan pada Kondisi Pakan Berbasis Low External Input. Lokal penelitian sapi potong dalam seminar nasional teknologi peternakan dan veteriner. Pasuruan (ID).
Im KS, Kim HJ, Chung KM, Kim HS, Park KW.1995. Effects of ovary type, oocyte grade, hormone, sperm concentration and fertilization medium on in vitro maturation, fertilization, and development of bovine follicular oocytes. AJAS. 8: 123-127.
Kaiin EM, Said S, Tappa B. 2008. Kelahiran anak sapi hasil fertilisasi secara in vitro dengan sperma hasil pemisahan. Media Peternakan. 31(1): 22-28. Labetubun J, Siwa IP. 2011. Kualitas spermatozoa kauda epididimis sapi bali
dengan penambahan laktosa atau maltosa yang dipreservasi pada suhu 3– 5oC. Jurnal Veteriner. 12(3): 200-207.
Lequaere AS, Marchandase J, Moreau B, Massip A, Donay I. 2003. Cell cycle at the time of maternal zygotic. J Biol Reprod. 69: 1707-1713.
Lonergan PH, Khatir F, Piumi D, Rieger P, Humblot, Boland MP. 1999. Effect of time interval from insemination to first cleavage on the developmental
10
characteristics, sex ratio and pregnancy rate after transfer of bovine embryos. J Reprod and Fert. 117: 159-167.
Margawati ET. 1999. The effective of growth factor on in vitro embryo development. J Med Vet. 6 (3) : 27 – 34.
Pujo M, Bejar ML, Paramio T. 2004. Depelopmental competence of heifer oocytes selected using briliant crsyl blue (BCB) tes. Theriogenology. 61: 35-44.
Purwantara B, Noor RR, Anderson G, Rodriguez-Martinez H. 2012. Banteng dan bali cattle in Indonesia : status and procests. J Reprod Dom Anim. 47(1): 2-6. Rollinsonn DHL. 1984. Bali Cattle. In: Evolution of Domesticated Animals.
Mason IL, editor. NewYork (US): Longman.
Rosenkrans CF, First NL. 1994. Effects of free amino acids and vitamins on cleavage and development rate of bovine zygotes in vitro. J Anim Sci. 72: 434-437.
Rosenkrans CF, Zeng QG, Mcnamara GT, Schoff PK, First NL. 1993. Development of bovine embryos in vitro as affected by energy substrates. J Biol Reprod. 49: 459-462.
Sagirkaya H, Yagmur M, Nur Z, Soylu MK. 2004. Replacement of fetal calf serum with synthetic serum substitute in the in vitro maturation medium: Effects on maturation, fertilization and subsequent development of cattle oocytes in vitro. Turk. J Vet Anim Sci. 28 : 779-784.
Sambery KY, Ngadiyono N, Sumadi. 2010. Estimasi dinamika populasi dan produktivitas sapi bali di kabupaten kepulauan Yapen, Propinsi Papua. Buletin Peternakan. 34(3): 169-177.
Sirard MA, Blodin P. 1996. Oocyte maturation and in vitro fertilization in cattle. J Anim Reprod Sci. 42 : 417-426.
Sobari I, Trilaksana B, Suatha IK. 2012. Perbedaan aktivitas ovarium sapi bali kanan dan kiri serta morfologi oosit yang dikoleksi menggunakan metode slicing. J Med Vet. 1(1): 1-11.
Sondy, Cecep F, Anita F. 2009. Analisis Permintaan Faktor Produksi Pada Usaha Ternak Sapi Potong Rakyat Dengan Pola Pemeliharaan Intensif. Bandung (ID) : Universitas Padjajaran.
Sumantri C, Boediono A, Oae A, Saha S, Suzuki T. 1997. Fertility of sperm from a tetraparen Ull chimeric bull. J Anim Reprod Sci. 46:35-45.
Susilowati T, Sumitro SB, Djati MS, Ciptadi G, Permono B. 1998. Optimalisasi maturasi oosit metode in vitro dengan kombinasi konsentrasi serum dan hormon pada TCM 199. Nat J. 2(1): 16-23.
Syaiful FL, Zesfin BP, Saladin R, Jaswandi, Hendri. 2010. Efektifitas berbagai ko-kultur sel pada sistem inkubasi CO2 5% untuk meningkatkan produksi embrio sapi secara in vitro. JITV. 15(1): 56-62.
Syaputri Ella. 2013. Inilah penjelasan tentang besaran impor daging sapi: kuota impor daging beku sapi bakalan tahun 2013. Antaranews [Internet]. [diunduh 2014 Jun 3]. Tersedia pada: Antaranews.com/berita/381060/inilah-penjelasan-tentang-besaran-impor-daging-sapi.
Takahashi Y, Hishinuma M, Matsui M, Tanaka H, Kanagawa H. 1996. Development of in vitro matured/fertilized bovine embryos in a chemically defined medium: influence of oxygen concentration in the gas atmosphere. J Vet Med Sci. 58(9): 897-902.
11 Vicanco, Mackie HW. 2001. Embrio transfer in ovine and caprine In : Biotechnology of reproduction. Palma G, editor. Buenos Aires (AGT): Ediciones Inta.
Ward FD, Rizos D, Corridan K, Quinn M, Boland, Lonergan P. 2001. Paternal influence on the time of first embryonic cleavage post insemination and the implications for subsequent bovine embryo development in vitro and fertility in vivo. Mol Reprod and Dev. 60:47-55.
12
RIWAYAT HIDUP
G Andri Hermawan lahir di Majalengka pada tanggal 15 April tahun 1991 merupakan anak ketiga dari pasangan Iwik Wikana dan Munah. Penulis menempuh pendidikan sekolah dasar pada tahun 1998 di SD Negeri Cibulan 2, masuk ke SMP Negeri 1 Lemahsugih tahun 2004, masuk ke SMA Negeri 1 Talaga tahun 2007. Tahun 2009 penulis mengikuti undangan seleksi masuk IPB (USMI) jalur nilai rapot dan diterima di Fakultas Kedokteran Hewan Institut Pertanian Bogor pada tahun 2010.
Selama menjadi mahasiswa, penulis aktif sebagai staf community development di unit kegiatan mahasiswa (UKM) bidang keilmiahan yaitu FORCES periode 2011-2012, anggota organisasi mahasiswa daerah (OMDA) himmpunan mahasiswa majalengka (HIMMAKA) 2010-2013, aktif sebagai ketua divisi kubah himpunan minat profesi ornithologi dan unggas Fakultas Kedokteran Hewan IPB periode 2011-2013, aktif sebagai pengurus paguyuban beasiswa karya salemba empat IPB sebagai kadiv HRD dan aktif sebagai pengurus divisi kesehatan kuda di club berkuda soorchery ridding club IPB dari tahun 2010-2013. Penulis pernah aktif menjadi pelaksana kontes ayam ketawa nasional menteri pertanian pada tahun 2012 sebagai ketua divisi humas pada tahun 2012 dan tahun 2013 sebagai wakil ketua pelaksana.
Selain aktif di UKM penulis mempunyai prestasi di bidang karya ilmiah yaitu dua proposal didanai program kreativitas mahasiswa pada tahun 2011, tiga proposal pada tahun 2012. Penulis pernah mendapatkan penghargaan sebagai presenter favorit dan peraih medali perak bidang pengambian masyarakat di pekan ilmiah nasional (PIMNAS) ke-25 tahun 2012 di Yogyakarta yang dilaksanakan oleh Direktorat Pendidikan Tinggi (DIKTI). Penulis pernah mengikuti kompetisi menunggang kuda serasi atau dressage pada pertandingan internal dan peserta jumping horse di detasemen kavaleri berkuda parongpong Bandung tahun 2011 dan 2012. Penulis penerima beasiswa karya salemba empat (KSE) tahun 2012-2015.