e-ISSN 2528-6870
KERAGAMAN ENZIM EKSTRASELULER DIHASILKAN OLEH JAMUR ENDOFIT ASAL Centella asiatica (L.) Urban
Diversity of Extracellular Enzymes Produced by Endophytic Fungus Originated from
Centella asiatica (L.) Urban
DWI N.SUSILOWATI1*, ALFI DWI SETIYANI2,NANI RADIASTUTI2,INDAH SOFIANA3, YADI SURYADI1 1Balai Besar Penelitian dan Pengembangan Bioteknologi dan
Sumberdaya Genetik Pertanian, Jl. Tentara Pelajar No. 3A Cimanggu Bogor 16111 2 Departemen Biologi, Fakultas Sains dan Teknologi
Universitas Islam Negeri Syarif Hidayatullah Jakarta, Jl. Ir. H. Djuanda No. 90, Ciputat, Banten, Indonesia 3Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam
Universitas Negeri Jakarta, Kampus A, Jl. Rawamangun Muka, Jakarta Timur 13220, Indonesia *Email: [email protected]
Diterima: 03-04-2020 ; Direvisi: 04-07-2020 ; Disetujui: 05-08-2020
ABSTRAK
Tanaman Pegagan (Centella asiatica) adalah tanaman obat yang dikenal bersimbiosis dengan berbagai jenis jamur endofit. Jamur endofit dipelajari secara ekstensif sebagai sumber senyawa bioaktif baru, termasuk enzim ekstraseluler. Enzim asparaginase, amilase, selulase, pektinase, protease, glukanase, dan lakase digunakan dalam industri. Penelitian ini bertujuan untuk mengkarakterisasi beberapa produksi enzim dari 40 jamur endofit dari C. asiatica. Penelitian dilakukan di Laboratorium Mikrobiologi, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian Bogor dan Laboratorium Mikrobiologi, PLT UIN Syarif Hidayatullah Jakarta pada bulan Februari hingga April 2019. Skrining enzim asparaginase, amilase, selulase, pektinase, protease, glukanase, dan lakase dilakukan pada medium Potato Dextrose Agar yang diperkaya dengan substrat tertentu. Hasil penelitian menunjukkan bahwa jumlah dan jenis enzim yang dihasilkan oleh jamur bervariasi. Phanerochaete chrysosporium MB02, Fusarium falciforme MB07, Trichaptum sp. MB11, Fusarium keratoplasticum MB12, Penicillium capsulatum MB15, Phoma multirostrata MB16, Fusarium oxysporum MB17, dan Mycochaetophora gentianae MB21 menghasilkan jumlah enzim tertinggi (6 jenis enzim). Berdasarkan enzim yang diproduksi (nilai indeks), Colletotrichum tabaci MB14 menghasilkan asparaginase tertinggi (indeks 2,65), Fusarium keratoplasticum MB12, Colletotrichum tabaci MB14, dan Phoma multirostrata MB16 untuk amilase (indeks 2,00); Peroneutypa scoparia MM10 untuk selulase (indeks 4.10); Colletotrichum karstii MM02 untuk pektinase (indeks 4.12); C. tabaci MB14 untuk protease (indeks 4.37); Acrocalymma vagum MB04 untuk glukanase (indeks 1,68); dan Fusarium solani MM03 untuk lakase (indeks 0,22). Colletotrichum tabaci MB14 merupakan isolat yang unggul penghasil 3 jenis enzim tertinggi (asparaginase, amilase, dan protease). Perlu dilakukan penelitian lebih lanjut untuk menganalisis secara kuantitatif produksi enzim ekstraseluler yang dihasilkan dan prospeknya untuk keperluan industri.
Kata kunci: Enzim ekstraseluler, pegagan, produksi in vitro
ABSTRACT
Asiatic Pennyworth (Centella asiatica) is a medicinal plant known to be symbiotic with various types of endophytic fungi. There are
extensively studied as a source of new bioactive compounds, including extracellular enzymes. This study aimed to characterize enzymes produced by 40 endophytic fungi from C. asiatica. This research was conducted at the Microbiology Laboratory, Indonesian Center for Agricultural Biotechnology and Genetic Resources Research and Development Bogor and the Microbiology Laboratory, PLT UIN Syarif Hidayatullah Jakarta in February to April 2019. Seven enzymes screened were asparaginase, amylase, cellulase, pectinase, protease, glucanase, and laccase on Potato Dextrose Agar enriched with a specific substrate. The results showed that the number and type of enzymes produced by the fungi varied. Phanerochaete chrysosporium MB02, Fusarium falciforme MB07, Trichaptum sp.MB11, Fusariumkeratoplasticum MB12, Penicillium capsulatum MB15, Phomamultirostrata MB16, Fusarium oxysporum MB17, and Mycochaetophora gentianae MB21 produced the highest enzyme number, i.e., six types of enzymes. Colletotrichum tabaci MB14 produced the highest index value for asparaginase (index 2.65), Fusarium keratoplasticum MB12, Colletotrichum tabaci MB14, and Phoma multirostrata MB16 for amylase (index 2.00); Peroneutypa scoparia MM10 for cellulase (index 4.10); Colletotrichum karstii MM02 for pectinase (index4.12); C. tabaci MB14 for protease (index 4.37); Acrocalymma vagum MB04 for glucanase (index 1.68); and Fusarium solani MM03 for laccase (index 0.22). Colletotrichum tabaci MB14 was superior because it produced the highest of 3 enzymes (asparaginase, amylase, and protease). Further study is required to find optimal conditions for each enzyme production for industrial purposes.
Keywords: Asiatic Pennyworth, extracellular enzyme, in vitro
production
PENDAHULUAN
Pegagan (Centella asiatica (L.) Urb.) banyak dimanfaatkan sebagai bahan baku obat tradisional. Tanaman pegagan sebagaimana tanaman lainnya diketahui bersimbiosis dengan jamur endofit. Jamur endofit adalah mikroorganisme yang hidup di dalam jaringan tanaman dengan cara membentuk koloni tanpa
menimbulkan gejala dan kerusakan yang nyata pada tanaman inang (Strobel dan Daisy 2003).
Banyak manfaat dari jamur endofit, seperti sebagai penghasil antibiotik, enzim, zat antimikroba, dan hormon pertumbuhan tanaman. Beberapa enzim ekstraseluler yang dihasilkan oleh jamur endofit telah diketahui, seperti amilase, selulase, glukanase, pektinase, lakase, protease, dan asparaginase (Choi et al. 2005, Theantana et al. 2009, Sunitha et al. 2013). Enzim-enzim yang dihasilkan oleh jamur endofit, juga memiliki peran yang penting dalam proses biodegradasi dan hidrolisis yang menjadi mekanisme penting terhadap infeksi patogen dan untuk memperoleh kebutuhan nutrisinya dari tanaman inang (Sunitha et al. 2013).
Kandungan senyawa utama dari pegagan yang berkhasiat sebagai obat adalah asiatikosida. Ghulamahdi et al. (2007) menyatakan bahwa tanaman pegagan aksesi Malaysia memiliki kandungan asiatikosida tinggi (0,80%). Wahyuno et al. (2010) menyatakan bahwa tanaman pegagan aksesi Bengkulu memiliki tingkat ketahanan yang paling baik terhadap bercak daun (Septoria centellae). Penelitian mengenai potensi jamur endofit dari tanaman pegagan aksesi Bengkulu dan Malaysia sebagai penghasil enzim ekstraseluler belum pernah dilakukan sebelumnya. Oleh karena itu diperlukan penelitian mengenai potensi jamur endofit tanaman pegagan aksesi Bengkulu dan Malaysia sebagai penghasil enzim ekstraseluler.
Penelitian ini difokuskan pada 7 macam enzim ekstraseluler, yaitu asparaginase, amilase, selulase, pektinase, protease, glukanase, dan lakase. Enzim L-asparaginase digunakan untuk mengkatalisis hidrolisis L-asparagin menjadi L-aspartat dan amonia (Theantana et al. 2009). L-asparaginase digunakan pada pengobatan kanker, yaitu untuk menghilangkan L-asparagin dari serum untuk mengendalikan pertumbuhan sel-sel tumor (McCredie et al. 1965; Verma et al. 2007). L-asparaginase yang dihasilkan oleh bakteri Escherichia
coli dan Erwinia carotovora diketahui memiliki efek
samping seperti hipersensitivitas yang mengarah pada reaksi alergi dan anafilaksis (Evans et al. 1982; Keating et al. 1993; Shrivastava et al. 2015). Di samping itu, enzim L-asparaginase digunakan dalam industri makanan untuk mencegah pembentukan senyawa akrilamid pada saat pemrosesan makanan pada suhu tinggi (Mohan et al. 2013). Hal ini sangat penting karena akrilamid merupakan neurotoksin yang bersifat karsinogenik pada manusia (Medeiros et al. 2012). L-asparaginase yang berasal dari jamur endofit belum banyak dilaporkan.
Enzim lakase telah lama menjadi perhatian para peneliti karena mampu mendegradasi berbagai macam senyawa polutan. Enzim lakase memiliki kisaran substrat yang luas, aktivitasnya stabil, dan sedikit faktor penghambat kerjanya sehingga sesuai untuk keperluan industri. Lakase juga dapat mengoksidasi berbagai macam substrat organik dan anorganik, termasuk mono-, di-mono-, polifenolmono-, aminofenolmono-, metoksifenolmono-, dan kompleks logam yang menjadi daya tarik utama bagi banyak aplikasi bioteknologi (Upadhyay et al. 2016). Demikian juga dengan tiga jenis enzim lainnya, seperti selulase, amilase, dan protease yang prospektif dikembangkan untuk keperluan industri dan lainnya.
Selulase merupakan mediator proses degradasi
baggase dan produksi etanol generasi kedua. Penambahan enzim selulase dapat mempercepat dan meningkatkan hasil hidrolisis enzimatik dari biomassa lignoselulosa. Sejumlah jamur penghasil selulase adalah
Botryosphaeria sp. dan Saccharicola sp. (Marques et al.
2018), sedangkan enzim amilase dikenal sebagai salah satu enzim yang aplikasinya luas dalam berbagai industri farmasi, makanan, tekstil, dan deterjen. Diperkirakan sebanyak 30% dari total produksi enzim secara global adalah amilase (de Souza dan e Magalhaes 2010). Selanjutnya, enzim protease dapat memecah protein menjadi konstituen yang lebih kecil dan menempati dua per tiga dari pasar enzim dunia. Protease banyak digunakan untuk keperluan bioremediasi, kosmetik, penghancuran sutera, kultur sel hewan, terapi, diagnosis, farmasi, dan industri makanan (Singh et al. 2016).
Penelitian ini bertujuan untuk mengevaluasi kemampuan dari 40 isolat jamur endofit asal tanaman pegagan dalam menghasilkan enzim-enzim ekstraseluler (asparaginase, amilase, selulase, pektinase, protease, glukanase, dan lakase).
BAHAN DAN METODE
Isolat Jamur Endofit
Sebanyak 40 isolat jamur endofit asal tanaman pegagan aksesi Bengkulu dan Malaysia yang digunakan berasal dari Koleksi Kultur Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian (Biogen Culture Collection). Isolat jamur endofit diremajakan pada media Potato Dextrose Agar (PDA) dan diinkubasi selama 5-7 hari pada inkubator suhu 27 ºC.
Tabel 1. Empat puluh isolat jamur endofit asal pegagan aksesi Bengkulu (MB) dan Malaysia (MM)
Table 1. Forty isolates of endophytic fungi from Asiatic Pennyworth Bengkulu (MB) and Malaysia (MM) accessions
No. No. Kode isolate Isolate code Spesies Species No. No. Kode isolate Isolate code Spesies Species
1 MB 01 Aspergillus austroafricanus 21 MM 02 Colletotrichum karstii
2 MB 02 Phanerochaete chrysosporium 22 MM 03 Fusarium solani
3 MB 03 Fusarium oxysporum 23 MM 04 Fusarium falciforme
4 MB 04 Acrocalymma vagum 24 MM 05 Eutypella sp.
5 MB 05 Perenniporia tephropora 25 MM 06 Trametes sp.
6 MB 06 Jamur Endofit 1 26 MM 07 Phyllosticta capitalensis
7 MB 07 Fusarium falciforme 27 MM 08 Phialemonium
dimorphosporum
8 MB 09 Fusarium oxysporum 28 MM 09 Collectotrichum
siamense
9 MB 10 Fusarium falciforme 29 MM 10 Peroneutypa scoparia
10 MB 11 Trichaptum sp. 30 MM 11 Phomopsis asparagi
11 MB 12 Fusarium keratoplasticum 31 MM 12 Phanerochaete
stereoides
12 MB 14 Colletotrichum tabaci 32 MM 13 Aspergillus oryzae
13 MB 15 Penicillium capsulatum 33 MM 14 Colletotrichum
gigasporum
14 MB 16 Phoma multirostrata 34 MM 16 Talaromyces pinophilus
15 MB 17 Fusarium oxysporum 35 MM 18 Colletotrichum tabaci
16 MB 18 Colletotrichum tabaci 36 MM 19 Chaetomium globosum
17 MB 19 Mycochaetophora gentiance 37 MM 20 Fusarium stiatum
18 MB 20 Ceratobasidium cornigerum 38 MM 21 Perenniporia corticola
19 MB 21 Mycochaetophora gentianae 39 MM 22 Fussarium falciforme
20 MM 01 Ceratobasidium sp. 40 MM 23 Colletotrichum tabaci
Uji Kualitatif Aktivitas Enzim Ekstraselular
Metode pengujian enzim ekstraseluler, yaitu amilase, selulase, pektinase, protease, glukanase, dan lakase, mengikuti (Sunitha et al. 2013), sedangkan asparaginase menurut Theantana et al. (2009) dan glukanase menurut Wood and Weisz (1984).
(a) Asparaginase
Miselia isolat jamur endofit pada medium PDA diambil dengan cork borer steril kemudian diinokulasikan pada medium agar Modified Czapex Dox’s (MCD) yang mengandung glukosa (2,0 g/L), L-asparagin (10,0 g/L), KH2PO4 (1,52 g/L), KCl (0,52 g/L), MgSO4.7H2O (0,52 g/L), CuNO3.H2O (0,001 g/L), ZnSO4.7H2O (0,001 g/L), dan FeSO4.7H2O (0,001 g/L), serta phenol red
(konsentrasi akhir 0,009%) sebagai indikator. Sebagai kontrol adalah medium MCD agar tanpa L-asparagin. Kultur jamur endofit diinkubasi pada suhu 30oC selama 3-7 hari. Isolat yang menghasilkan L-asparaginase ditandai dengan perubahan warna media uji dari kuning menjadi oranye-merah, sedangkan yang bukan penghasil L-asparaginase warna media uji tetap kuning (Theantana et al. 2009).
(b) Amilase
Aktivitas enzim amilase diamati berdasarkan pertumbuhan jamur pada medium agar Glucose Yeast
extract Pepton (GYP) yang mengandung glukosa (1,0
g/L), ekstrak yeast (0,1 g/L), peptone (0,5 g/L), agar (16 g/L) serta soluble starch pH 6,0 (0,2%). Kultur diinkubasi pada suhu 30oC selama 5-7 hari kemudian dituangkan
larutan 1% iodine dalam 2% larutan potassium iodide. Uji amilase positif ditandai dengan perubahan warna media uji dari coklat tua menjadi lebih pudar, sedangkan uji amilase negatif media tetap berwarna coklat tua (Sunitha et al. 2013).
(c) Selulase
Isolat jamur endofit ditumbuhkan pada medium agar GYP yang diperkaya dengan 0,5%
Carboxy-methylcellulose (CMC). Setelah diinkubasikan selama 5-7
hari pada suhu 30oC, larutan 0,2% congo red dituangkan ke dalam medium dan dibiarkan selama 15-20 menit, lalu dibilas dengan larutan 1 M NaCl. Adanya zona bening di sekitar koloni jamur menunjukkan aktivitas selulase positif (Sunitha et al. 2013).
(d) Pektinase
Produksi enzim pektinase dari isolat jamur endofit diamati pada medium pektin agar yang mengandung pektin (5 g/L), ekstrak yeast (1 g/L), dan agar (15 g/L). Kultur jamur endofit diinkubasi pada suhu 30oC selama 3-7 hari, lalu ke dalam cawan petri uji dituangkan larutan 1%
hexadecyl trimethyl ammonium bromide. Zona bening di
sekitar koloni jamur menunjukkan aktivitas pektinase (Sunitha et al. 2013).
(e) Protease
Aktivitas enzim protease diamati pada medium agar GYP yang diperkaya dengan 0,4% gelatin (pH 6,0) dengan menambahkan 5 mL larutan 8% gelatin steril ke dalam 100 mL medium GYP. Penambahan gelatin bertujuan sebagai substrat untuk enzim protease. Isolat jamur endofit diinokulasikan pada medium GYP kemudian diinkubasikan selama 5-7 hari pada suhu 30oC. Selanjutnya ditambahkan larutan ammonium sulfat jenuh ke dalam kultur. Aktivitas proteolitik ditandai dengan terbentuknya zona bening dan endapan di sekitar koloni jamur (Sunitha et al. 2013).
(f) Glukanase
Aktivitas enzim glukanase diukur menggunakan metode dari Wood dan Weisz (1984)menggunakan medium yang mengandung 0,2% glukan padat. Medium glukan padat dibuat dengan terlebih dahulu menyiapkan pelet glukan, yaitu tepung oat (10 g) dilarutkan di dalam akuades (100 mL), kemudian pH disesuaikan menjadi 10 dengan Na2CO3 20% sambil diaduk dengan magnetik
stirer. Setelah itu, larutan glukan dipanaskan pada suhu
45oC selama 30 menit sambil terus diaduk, kemudian disentrifugasi pada kecepatan 5000 rpm pada suhu 4oC
selama 15 menit. Supernatan diambil dan pHnya dijadikan 4,5 dengan menambahkan HCl 1 N sedikit demi sedikit, dan disentrifugasi kembali pada kecepatan 5000 rpm selama 20 menit. Supernatan diambil dan tambahkan etanol 95% secara perlahan sambil diaduk dengan
magnetic stirer dan sentrifugasi kembali dengan
kecepatan 500 rpm pada suhu 4oC selama 10 menit. Endapan diambil, tambahkan larutan Buffer Fosfat Salin (PBS) sebanyak 15-20 mL, dan diaduk hingga larut, kemudian disimpan semalam pada suhu 4oC. Pelet glukan yang dihasilkan siap untuk digunakan.
Medium glukan terdiri atas pelet glukan (1 mL), Na2HPO4 (0,065 g), KH2PO4 (0,15 g), NaCl (0,25 g), (NH4)2SO4 (0,05 g), MgSO4.7H2O (0,012 g), CaCl2 (0,005 g), pepton (0,125 g), yeast ekstrak (0,05 g), dan agar (2 g) dalam akuades 100 mL. Kultur jamur endofit ditumbuhkan pada media glukan padat inkubasikan selama 3-7 hari. Setelah koloni jamur endofit tumbuh, dituangkan larutan 0,2% congo red ke dalam kultur, lalu dibilas dengan larutan 1 M NaCl selama 15-20 menit. Terbentuknya zona bening di sekitar koloni jamur menunjukkan adanya aktivitas glukanase yang dihasilkan oleh jamur yang diuji.
(g) Lakase
Aktifitas enzim lakase diamati berdasarkan pertumbuhan jamur endofit pada media GYP yang diperkaya dengan 1-napthol (0,05 g). Warna biru yang terbentuk di sekitar koloni jamur menunjukkan aktivitas lakase (Sunitha et al. 2013).
Indeks Enzim Ekstraseluler
Pengamatan aktivitas enzim ekstraseluler di atas dilakukan terhadap terbentuknya zona hidrolisis, dan dinyatakan dalam indeks enzim ekstraseluler. Indeks enzim diperoleh dengan membandingkan diameter zona hidrolisis dengan diameter koloni jamur endofit pada media uji.
HASIL DAN PEMBAHASAN
Keragaman Produksi Enzim Ekstraseluler
Sebagian besar jamur endofit asal pegagan aksesi Bengkulu dan Malaysia menghasilkan enzim ekstraseluler. Jumlah dan jenis enzim ekstraseluler yang dihasilkan oleh 40 isolat jamur endofit bervariasi (Gambar 1). Jamur endofit asal pegagan dari Bengkulu menghasilkan lebih banyak enzim ekstraseluler dibandingkan dengan endofit asal pegagan dari Malaysia. Empat belas dari 19 isolat jamur endofit asal pegagan dari Bengkulu (73,7%) dan 8 dari 21 jamur
endofit asal pegagan dari Malaysia (38,1%) menghasilkan 4-7 jenis enzim ekstraseluler.
Gambar 1. Jumlah jamur endofit pegagan aksesi Bengkulu (MB) dan Malaysia (MM) yang menghasilkan enzim ekstraseluler
Figure 1. Amount of endophytic fungi from Asiatic Pennyworth Bengkulu (MB) and Malaysia (MM) accession producing a number of extracellular enzymes
Berdasarkan kemampuan menghasilkan
sejumlah enzim ekstraseluler maka isolat jamur endofit asal pegagan dapat dikelompokkan menjadi 8 grup. Ada 8 isolat jamur endofit asal pegagan dari Bengkulu yang menghasilkan 6 jenis enzim sekaligus, yaitu jamur
Phanerochaete chrysosporium isolat MB02, Fusarium falciforme MB07, Trichaptum sp.MB11, Fusarium keratoplasticum MB12, Penicillium capsulatum MB15, Phoma multirostrata MB16, Fusarium oxysporum
MB17, dan Mycochaetophora gentianae MB21. Hanya 6 isolat jamur endofit asal pegagan aksesi Malaysia yang mampu menghasilkan 6-7 jenis enzim, yaitu
Colletotrichum karstii isolat MM02, Fusarium solaniMM03, Fusarium falciforme MM04, Eutypella
sp.MM05, Aspergillus oryzae MM13, dan Fusarium
falciforme MM22. Hasil penelitian juga menunjukkan
bahwa jamur genus Fusarium paling dominan menghasilkan beberapa jenis enzim ekstraseluler. Jumlah enzim yang dihasilkan dalam penelitian ini sejalan dengan penelitian (Sunitha et al. 2013), bahwa jamur endofit Fusicoccum spp. dan Isaria spp. yang diisolasi dari tanaman obat mampu menghasilkan 5 jenis enzim ekstraseluler.
Asparaginase dan Amilase
Berdasarkan kemampuan menghasilkan enzim asparaginase (asparaginolitik indeks) maka 22 dari 40 isolat jamur endofit asal pegagan dari Bengkulu dan Malaysia (55%) bervariasi kemampuannya (Gambar 2a
dan 2b). Colletotrichum tabaci isolat MB14 memiliki indeks asparaginolitik tertinggi, yaitu sebesar 2,65 (Gambar 3a), diikuti oleh Penicillium capsulatum MB15 dengan indeks asparaginolitik sebesar 2,43. Hal ini sejalan dengan penelitian Theantana et al. (2009). yang menemukan bahwa isolat jamur endofit Fusarium sp.,
Colletotrichum sp., Penicillium sp., Eupenicillium sp.,
dan Talaromyces sp. dari tanaman obat di Thailand mampu memproduksi enzim asparaginase. Jamur-jamur lainnya yang dapat menghasilkan L-asparaginase adalah
Aspergillus sp., A. terreus, A. flavus, dan Emericella nidulans (Lapmak et al. 2010).
Jamur endofit yang dapat menghasilkan enzim L-asparaginase dideteksi melalui pembentukan zona berwarna merah muda pada media uji MCD agar yang mengandung L-asparagin sebagai substrat. Terbentuknya zona merah muda ini sebagai akibat reaksi hidrolisis L-asparagin menjadi L-aspartat dan ammonia. Ammonia yang terbentuk menyebabkan pH media meningkat sehingga mengubah warna indikator phenol
red dari kuning (suasana asam) menjadi merah muda
(suasana basa).
Penemuan sumber-sumber enzim L-asparaginase asal jamur endofit ini membuka jalan pengembangan enzim L-asparaginase untuk industri farmasi. Hal ini disebabkan efek samping L-asparaginase asal jamur lebih rendah daripada L-asparaginase asal bakteri (Sarquis et al. 2004). Enzim L-asparaginase asal bakteri
Escherichia coli dan Erwinia meskipun ampuh
digunakan sebagai agen kemoterapi, namun chrysanthemi menimbulkan efek samping seperti muntah, leukopenia, demam, ruam kulit, mual, embolisis thrombo, kesulitan bernafas, penurunan berat badan, penurunan tekanan darah, berkeringat, imunosupresi, kehilangan kesadaran, pankreatitis akut, dan kejang neurologis (Rossi et al. 2004; Ramya et al. 2012). Asparagin merupakan nutrisi yang diperlukan baik oleh sel-sel normal maupun sel kanker. Asparaginase mampu mendegradasi asparagin menjadi asam aspartat dan ammonia. Asparagin dalam bentuk L-asparagin merupakan asam amino non esensial yang dibutuhkan oleh sel tumbuhan untuk sintesis protein. Konsentrasi asparagin yang sedikit hanya memengaruhi viabilitas sel abnormal, karena sel-sel ini memerlukan asparagin pada jumlah yang tinggi secara abnormal (Mitchell et al. 1994). Pada sel-sel normal menghasilkan enzim asparagin sintetase yang dapat mendegradasi asparagin dari asam aspartat, sedangkan pada sel-sel 0 2 4 6 8 10 0 1 2 3 4 5 6 7 ∑ iso la t ja m u r e n d of it
∑ Enzim yang Dihasilkan
MB MM
kanker dan tumor enzim asparagin sintetase berada pada jumlah yang sedikit (Nakamura et al. 1999).
Gambar 2a. Indeks enzim ektraseluler jamur endofit asal pegagan aksesi Bengkulu.
Figure 2a. Extracellular enzyme index of endophytic fungi from Asiatic pennyworth Bengkulu accessions
Gambar 2b. Indeks enzim ektraseluler jamur endofit asal pegagan aksesi Malaysia.
Figure 2b. Extracellular enzyme index of endophytic fungi from Asiatic pennyworthMalaysia accessions.
2 ,0 6 2 ,1 1 2 ,1 9 1 ,8 2 1 ,6 1 1,9 6 1 ,8 8 2 ,6 5 2 ,4 3 1 1 ,4 6 1,59 1,6 5 1 ,5 8 1 1 ,5 2 2 1 2 1 0 0,5 1 1,5 2 2,5 3 In d e ks e n zi m e xt ra sel u le r Kode isolat Asparaginase Amilase 0 2 ,1 1 1 ,4 3 1 ,4 1 ,3 3 1 ,2 1 ,4 3 1 1 ,2 9 1 ,1 1 1 1 1 1 ,6 4 1 1 ,2 7 1 1 1 1 1 1 ,3 1 1 0 0,5 1 1,5 2 2,5 In d e ks En zi m e xt ra sel u le r Kode isolat Asparaginase Amilase
(a) (b)
Gambar 3a. Hasil uji positif aktivitas L-asparaginase yang dihasilkan oleh jamur endofit
Colletotrichum tabaci MB14, (3b) aktivitas
enzim amilase dari jamur endofit F.
keratoplasticum MB12.
Figure 3a). Positive reaction of L-asparaginase produced by endofitic fungus Colletotrichum tabaci MB14, (3b) amylase production by F. keratoplasticum MB12.
Hasil pengujian aktivitas amilase dari isolat jamur endofit asal pegagan diketahui sebanyak 22 isolat dari 40 isolat yang diuji (55,00%) dapat menghasilkan enzim amilase (Gambar 2a dan 2b). Isolat Fusarium
keratoplasticum MB12 (Gambar 3b), Colletotrichum tabaci MB14, dan Phoma multirostrata MB16 memiliki
nilai indeks amilolitik masing-masing sebesar 2,00. Sejumlah isolat jamur yang juga diketahui positif menghasilkan enzim amilase dengan aktivitas rendah hingga sedang, antara lain Fusarium sp., Colletotrichum sp., Aspergillus sp., dan Biosporus sp. (Amirita et al. 2012). Enzim amilase yang dihasilkan oleh jamur bersifat lebih stabil daripada enzim yang dihasilkan oleh bakteri (Duochuan et al. 1997).
Enzim amilase, lipase, dan protease biasanya disekresikan oleh jamur untuk mendegradasi komponen membran plasma tanaman kemudian digunakan sebagai sumber nutrisi (Liao et al. 2012). Enzim-enzim tersebut bertindak memfasilitasi penetrasi hifa, melepaskan sumber karbon atau memodifikasi sinyal kimia yang diproduksi oleh tanaman inang (Huang 2001; ten Have
et al. 2002). Enzim amilase akan menghidrolisis substrat
pati menjadi senyawa karbohidrat yang lebih sederhana seperti maltosa dan glukosa. Selanjutnya maltosa, glukosa, dan lipid maupun protein hasil degradasi lipase dan protease menjadi sumber nutrisi selama kolonisasi fungi.
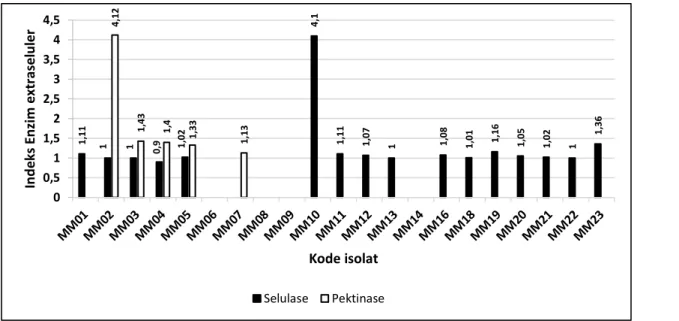
Selulase dan Pektinase
Dari 40 isolat jamur endofit yang diuji, 70% diantaranya (28 isolat) menunjukkan hasil positif sebagai penghasil selulase (Gambar 4a dan 4b). Indeks selulolitik tertinggi (4,10) diperoleh dari isolat jamur endofit Peroneutypa scoparia MM10 (Gambar 5a) dan diikuti oleh Penicillium capsulatum MB15 dengan indeks selulolitik sebesar 1,37. Penelitian sebelumnya
melaporkan bahwa jamur Coletotrichum
gloeosporioides, Aspergillus versicolor, dan
Cladosporium cladosporioides pada jamur endofit yang
ada pada tujuh biji-bijian yang mengandung minyak merupakan produser enzim selulase yang tinggi (Venkatesagowda et al. 2012). Enzim selulase dikelompokkan berdasarkan aktivitas spesifiknya terhadap substrat yaitu endoglukanase, selobiohidrolase, dan eksoglukohidrolase. Ketiga enzim tersebut bekerja sama dalam mengurai selulosa.
Gambar 4a. Indeks enzim ektraseluler jamur endofit asal pegagan aksesi Bengkulu.
Figure 4a. Extracellular enzyme index of endophytic fungi from Asiatic pennyworth Bengkulu accessions.
1 ,0 6 0 ,9 9 0 ,9 2 1,02 1 0,9 8 1 ,3 7 1 1,0 7 1 ,0 3 1 ,0 7 1 1 ,0 4 1 ,0 6 1 ,0 8 1 ,0 2 1 0,98 1,0 5 5 1 1 ,3 4 0 ,6 8 1 ,1 4 1 ,1 3 1 ,1 1 1,2 9 0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 In d e ks En zi m e xt ra sel u le r Kode isolat Selulase Pektinase
Gambar 4b. Indeks enzim ektraseluler jamur endofit asal pegagan aksesi Malaysia
Figure 4b. Extracellular enzyme index of endophytic fungi from Asiatic pennyworth Malaysia accessions
Terbentuknya zona bening di sekitar pertumbuhan jamur endofit menunjukkan bahwa jamur mampu menghasilkan selulase yang terdeteksi berwarna oranye di sekitar latar berwarna pewarna dari reaksi amilum dengan zat warna congo red. Pembilasan warna
congo red dengan menggunakan NaCl agar congo red
tidak terikat oleh selulosa, sehingga dihasilkan diameter yang ada dalam agar substrat. Selulosa adalah senyawa yang tidak larut di dalam air biasanya selulosa tidak ditemukan dalam keadaan murni melainkan berasosiasi dengan polisakarida lain seperti hemiselulosa atau lignin membentuk kerangka utama dinding sel tumbuhan. Substrat selulosa banyak ditemukan pada dinding sel tumbuhan terutama pada tangkai, batang, dahan, dan semua bagian berkayu dari jaringan tumbuhan (Mosier et al. 2003). Sebanyak 40 isolat yang diuji ternyata 47,5% diantaranya (19 isolat) menghasilkan enzim pektinase (Gambar 4a dan 4b). Colletotrichum karstii MM02 memiliki nilai indeks pektinolitik tertinggi sebesar 4,12 (Gambar 5b).
Enzim pektinase diinduksi dengan adanya substrat berupa pektin pada jamur patogen maupun endofit. Enzim pektinase pada mikroba sangat penting pada proses fitopatologi, simbiosis mikroba tanaman, dan dekomposisi material tanaman yang mati. Degradasi jaringan inang oleh fitopatogen biasanya diawali dengan produksi enzim pektinase yang merupakan enzim utama yang terlibat dalam serangan tanaman (Hoondal et al. 2001). Jika diperoleh jamur endofit yang dapat
mendegradasi substrat pektin, hal ini memiliki indikasi bahwa jamur tersebut merupakan laten patogen (Choi et
al. 2005).
(a) (b)
Gambar 5a. Uji kualitatif produksi selulase isolat jamur endofit asal pegagan pada media GYP + 0,5% Carboxy-methylcellulose (CMC) dari Peroneutypa scoparia MM10, (5b) hasil uji positif dari
Colletotrichum karstii MM02 sebagai
penghasil pektinase.
Figure 5a. The qualitative test of cellulose production from endophytic fungi from Asia pennyworth on GYP + 0.5% Carboxy-methylcellulose (CMC) media of Peroneutypa scoparia MM10, (5b) the positive results of Colletotrichum karstii MM02 as pectinase producer
1 ,1 1 1 1 0,9 1,0 2 4 ,1 1 ,1 1 1 ,0 7 1 1,0 8 1 ,0 1 1 ,1 6 1 ,0 5 1 ,0 2 1 1 ,3 6 4 ,1 2 1 ,4 3 1 ,4 1 ,3 3 1 ,1 3 0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 In d e ks En zi m e xt ra sel u le r Kode isolat Selulase Pektinase
Sieber-Canavesi et al. (1991) menyatakan bahwa kemampuan jamur endofit menghasilkan enzim ekstraseluler selulase dan pektinase secara bersamaan dapat berimplikasi bahwa jamur tersebut mampu melakukan penetrasi sel-sel hidup dan mendekomposisi jaringan mati. Kemampuan jamur endofit menghasilkan enzim-enzim tadi dapat memberikan mekanisme resistensi pada tanaman inangnya terhadap invasi patogen, untuk memperoleh nutrisi dari tanaman inangnya atau sebagai laten patogen (Choi et al. 2005; Saikkonen et al. 1998; Saikkonen et al. 2004).
Berdasarkan Gambar 4a dan 4b diketahui bahwa ada beberapa isolat yang menghasilkan selulase dan pektinase secara bersamaan, diantaranya Phanerochaete
chrysosporium MB02, dan beberapa spesies dari genus Fusarium seperti F. falciforme MB07 dan MB10, F. keratoplasticum MB12, F. oysporum MB17, F. solani
MM03, dan beberapa spesies Colletotrichum (C. tabaci MB18 dan C. karstii MM02). Departemen Energi Pemerintah Amerika yang menyusun draf genom P.
chrysosporium, menunjukkan bahwa P. chrysosporium
memiliki semua gen yang mengkode semua enzim yang diperlukan untuk mendegradasi secara sempurna semua komponen utama dinding sel tanaman yaitu selulosa, hemiselulosa dan lignin (Kersten dan Cullen 2007).
Genus Fusarium tergolong ke dalam pendegradasi selulosa dan hemiselulosa yang sangat kuat, demikian juga dengan genus Colletotrichum (Huang et al. 2015; Velho et al. 2018). Hal ini sejalan dengan laporan (Huang 2001; Kubicek et al. 2014) yang menemukan bahwa enzim selulase, pektinase, cutinase, dan hemiselulase bersinergi untuk mendegradasi kutikula dan dinding sel tanaman inang sebelum melakukan penetrasi lebih lanjut.
Protease, Glukanase, dan Lakase
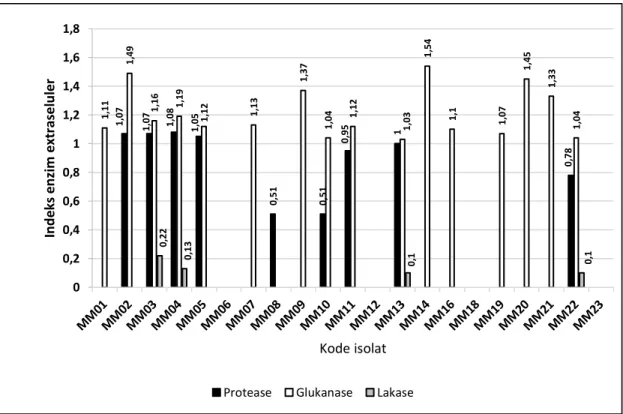
Dari sebanyak 40 isolat jamur endofit yang diuji aktivitas enzim proteasenya, terdapat 23 isolat (57,5 %) yang menunjukkan hasil positif (Gambar 6a dan 6b).
Colletotrichum tabaci MB14 memiliki nilai indeks
proteolitik yang paling tinggi diantara jenis jamur endofit pegagan lainnya.
Hasil ini sejalan dengan penelitian sebelumnya yang menyatakan bahwa genus Colletotrichum seperti
Colletotrichum gloeosporioides, Colletotrichum carssipes, Colletotrichum falctum, selanjutnya
Curvularia vermiformis, Drechslera hawaiiensis, dan Xylariales merupakan jamur penghasil enzim protease
(Amirita et al. 2012).
Gambar 6a. Indeks enzim ektraseluler jamur endofit asal pegagan aksesi Bengkulu.
Figure 6a. Extracellular enzyme index of endophytic fungi from Asiatic pennyworth Bengkulu accessions.
1 ,0 7 1 ,0 7 1 ,0 8 1 ,0 5 0 ,5 1 0 ,5 1 0 ,9 5 1 0 ,7 8 1 ,1 1 1 ,4 9 1 ,1 6 1 ,1 9 1 ,1 2 1 ,1 3 1 ,3 7 1 ,0 4 1,1 2 1 ,0 3 1 ,5 4 1 ,1 1 ,0 7 1 ,4 5 1 ,3 3 1 ,0 4 0 ,2 2 0 ,1 3 0 ,1 0,1 0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 In d e ks e n zi m e xt ra sel u le r Kode isolat
Gambar 6b. Indeks enzim ektraseluler jamur endofit asal pegagan aksesi Malaysia.
Figure 6b. Extracellular enzyme index of endophytic fungi from Asiatic pennyworth Malaysia accessions
(a) (b) (c)
Gambar 7a. Hasil uji positif produksi protease pada media GYP agar + 0,2% gelatin hasil uji positif dari C.
tabaci MB14, (7b) hasil uji positif dari Acrocalymma vagum MB04 sebagai penghasil glukanase,
dan (7c) hasil uji positif dari Fusarium solani MM03 sebagai penghasil lakase
Figure 7a. The positive results of protease production on GYP agar + 0.2% gelatin of C. tabaci MB14, (7b) the positive results of Acrocalymma vagum MB04 as glucanase producer, and (7c) the positive results of Fusarium solani MM03as laccase producer
Penelitian Sunitha et al. (2013) melaporkan bahwa jamur Fusarium solani Ci24 merupakan produsen protease yang tinggi, selain jamur lainnya seperti Aspergillus sp. Ci1 dan Isaria sp. Ci12. Protease adalah enzim yang menghidrolisis ikatan peptida pada molekul protein yang menghasilkan peptida atau asam amino. Protein terdapat pada hampir seluruh bagian
tumbuhan. Enzim protease ini banyak dipergunakan untuk aplikasi klinis terutama untuk perawatan pasien diabetes. Diperolehnya sejumlah jamur-jamur endofit penghasil protease menjadi peluang besar ditemukannya jenis protease baru yang mungkin bisa menjadi alternatif industri yang khusus.
1 ,0 7 1 ,0 7 1 ,0 8 1 ,0 5 0 ,5 1 0 ,5 1 0 ,9 5 1 0 ,7 8 1 ,1 1 1 ,4 9 1 ,1 6 1 ,1 9 1 ,1 2 1 ,1 3 1 ,3 7 1 ,0 4 1,12 1 ,0 3 1 ,5 4 1 ,1 1 ,0 7 1 ,4 5 1 ,3 3 1 ,0 4 0 ,2 2 0 ,1 3 0 ,1 0,1 0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 In d e ks e n zi m e xt ra sel u le r Kode isolat
Glukanase
Terdapat 34 isolat (85%) yang menunjukkan hasil positif aktivitas enzim glukanase dari 40 isolat yang diujikan (Gambar 6a dan 6b). Indeks glukanolitik paling tinggi didapatkan dari Acrocalymma vagum MB04 sebesar 1,68 (Gambar 7b). Fenomena ini ternyata sejalan dengan hasil penelitian evaluasi jamur endofit pegagan aksesi Bengkulu dan Malaysia ini, karena menunjukkan hasil positif yang cukup tinggi yaitu 85%. Glukanase merupakan salah satu enzim yang dapat menghidrolisis glukan pada dinding sel jamur patogen yang menginfeksi tanaman (Budiarti et al. 2004). Glukan juga banyak terdapat pada dinding sel tumbuhan (Hunter Jr et al. 2002).
Lakase
Sebanyak 40 isolat diuji kemampuan menghasilkan enzim lakase dan diperoleh 5 jenis jamur endofit (15%) yang menunjukkan hasil positif (Gambar 6a dan 6b). Jamur endofit Fusarium oxysporum MB17 dan Fusariumsolani. MM03 merupakan jamur endofit pegagan yang dapat menghasilkan enzim lakase dengan indeks yang tinggi dibandingkan beberapa isolat lainnya dengan nilai indeks tertinggi sebesar 0,22 pada
Fusariumsolani MM03 (Gambar 7c).
Pada penelitian ini dapat dilihat bahwa hanya 6 jamur endofit diantara 40 jamur yang diuji memiliki potensi menghasilkan lakase (Gambar 6a dan 6b). Hal ini sejalan juga dengan hasil penelitian (Sunitha et al. 2013), bahwa sedikit ditemukan jamur endofit maupun jamur asal marin sebagai penghasil enzim lakase. Bahkan Renato et al. (2005) menyatakan bahwa tidak satupun jamur endofit yang berhasil diisolasi dapat menghasilkan enzim lakase. Hal ini kemungkinan besar disebabkan sifat dari jamur endofit sehingga tidak banyak atau bahkan tidak ada jamur endofit yang menghasilkan enzim lakase. Adanya aktivitas enzim lakase tentu saja akan dapat merusak tanaman yang menjadi inangnya.
Hal yang menarik dari penelitian ini adalah sejumlah isolat jamur endofit yang diisolasi dari tanaman pegagan aksesi Bengkulu dan Malaysia baru pertama kali diuji potensinya sebagai penghasil sejumlah enzim ekstraseluler. Berdasarkan hasil penelitian ini tampak bahwa jamur endofit asal pegagan yang berpotensi menonjol sebagai penghasil sejumlah enzim ekstraseluler diantaranya adalah Colletotrichum
tabaci MB14 yang unggul dalam produksi enzim
asparaginase, amilase, dan protease dibandingkan isolat-isolat lainnya. Jamur endofit pegagan menghasilkan sejumlah enzim hidrolitik sebagai mekanisme resistensi
terhadap invasi patogen dan untuk mendapatkan nutrisi dari tanaman inangnya. Informasi mengenai pola-pola penggunaan substrat dan jenis-jenis enzim ekstraseluler yang dihasilkan oleh jamur endofit asal pegagan ini sangat penting untuk mengembangkan peran fungsional dari jamur tersebut (Carroll dan Petrini 1983). Jamur-jamur yang menghasilkan proteinase dan pektinase biasanya tergolong parasit lemah atau laten patogen. Sementara jamur endofit tersebut bersifat mutualistik, terkadang menjadi saprofit biasanya menghasilkan selulase, mannanase, dan xylanase.
KESIMPULAN
Kemampuan dari 40 isolat jamur endofit asal tanaman pegagan dalam menghasilkan enzim-enzim ekstraseluler (asparaginase, amilase, selulase, pektinase, protease, glukanase, dan lakase) beragam; ada yang dapat memghaislkan 1- 6 jenis enzim. Tujuh isolat jamur yang mampu menghasilkan 6 macam enzim adalah Phanerochaete chrysosporium MB02, Fusarium
falciforme MB07, Trichaptum sp. MB11, Fusarium keratoplasticum MB12, Penicillium capsulatum MB15, Phoma multirostrata MB16, Fusarium oxysporum
MB17, dan Mycochaetophora gentianae MB21. Aktivitas enzim tertinggi ditunjukkan oleh isolat
Colletotrichum tabaci MB14 untuk enzim asparaginolitik dengan nilai indeks 2,65. Beberapa isolat lainnya adalah Fusarium keratoplasticum MB12,
Colletotrichum tabaci MB14, dan Phoma multirostrata
MB16 penghasil amilolitik tertinggi (2,00);
Peroneutypa scoparia MM10 untuk selulotik tertinggi
(4,10); Colletotrichum karstii MM02 untuk pektinolitik tertinggi (4,12); C. tabaci MB14 untuk proteolitik tertinggi (4,37); Acrocalymma vagum MB04 untuk glukanolitik tertinggi (168); Fusarium solani MM03 untuk lignolitik tertinggoi (0,22), dan F. oysporum untuk lakase tertinggi (0,21). Colletotrichum tabaci MB14 merupakan isolat yang unggul sebagai penghasil 3 jenis enzim tertinggi, yaitu asparaginase, amilase, dan protease. Perlu dilakukan penelitian lebih lanjut untuk menganalisis secara kuantitatif produksi enzim ekstraseluler yang dihasilkan dan prospeknya untuk keperluan industri.
UCAPAN TERIMA KASIH
Diucapkan terima kasih kepada Dr. Nurliani Bermawie dari Balittro dan Dr Ika Roostika dari BB Biogen atas ijin akses tanaman pegagan aksesi Bengkulu untuk diisolasi kapang endofitnya. Ucapan terima kasih
disampaikan juga kepada Siti Aminah dan Jajang Kosasih atas bantuannya selama pelaksanaan kegiatan penelitian di laboratorium mikrobiologi BB Biogen.
PERNYATAAN KONTRIBUSI
Dwi N. Susilowati berperan sebagai kontributor utama dalam perencanaan, pelaksanaan, dan penulisan naskah. Nani Radiastuti berkontribusi dalam memberikan saran dan masukan selama penyusunan naskah, Alfi Dwi Setiyani berkontribusi melaksanakan kegiatan penelitian di laboratorium mikrobiologi BB Biogen dan laboratorium PLT Terpadu UIN Syarif Hidayatullah, Yadi Suryadi berkontribusi melakukan analisis data, Indah Sofiana berkontribusi melakukan rekapitulasi data awal dan visualisasi data menjadi grafik.
DAFTAR PUSTAKA
Amirita, A., Sindhu, P., Swetha, J., Vasanthi, N.S. & Kannan, K.P. (2012) Enumeration of Endophytic Fungi From Medicinal Plants and Screening of Extracellular Enzymes. World Journal of Science
and Technology. 2 (2), 13–19.
Budiarti, S.W., Widyastuti, S.M. & Margino, S.T. (2004) β-1,3-Glucanase Enzyme Production by Tricoderma reeseiduring Mycoparasitism.
Makalah Seminar Pertemuan Bioteknologi Indonesia, Malang.
Carroll, G. & Petrini, O. (1983) Patterns of Substrate Utilization by some Fungal Endophytes from Coniferous Foliage. Mycologia. 75 (1), 53–63. doi:10.1080/00275514.1983.12021637.
Choi, Y.W., Hodgkiss, I.J. & Hyde, K.D. (2005) Enzyme Production by Endophytes of Brucea javanica. Journal Agricurtural Technol. 1, 22– 66.
Duochuan, L., Yijun, Y., Youliang, P., Chongyao, S., Peijin, Z. & Yicun, H. (1997) Purification and Properties of Thermostable Alpha Amylase from the Thermophilic Fungus Thermomyces lanuginosus. Acta Microbiologica Sinica. 32 (2), 107–114.
Evans, W.E., Tsiatis, A., Rivera, G., Murphy, S.B., Dahl, G. V., Denison, M., Crom, W.R., Barker, L.F. & Mauer, A.M. (1982) Anaphylacfoid Reactions to Escherichia coli and Erwinia asparaginase in Children with Leukemia and Lymphoma. Cancer. 49 (7), 1378–1383.
doi:10.1002/1097- 0142(19820401)49:7<1378::AID-CNCR2820490713>3.0.CO;2-Z.
Ghulamahdi, M., Azis, S.A., Bermawie, N. & Hernaini (2007) Evaluasi Karakter Morfologi, Fisiologi
dan Genetik Pegagan mendukung Standarisasi Mutu Pegagan. Kerjasama Kemitraan Peneitian
Pertanian dengan Perguruan Tinggi (KKPJT). ten Have, A., Tenberge, K.B., Benen, J.A.E., Tudzynski,
P., Visser, J. & van Kan, J.A.L. (2002) The Contribution of Cell Wall Degrading Enzymes to Pathogenesis of Fungal Plant Pathogens. In: Kempken F, editors. Agricultural Applications.
The Mycota (A Comprehensive treatise on fungi as Experimental Systems for Basic and Apllied Research). 11, 1st ed.Berlin (DE):
Springer-Verlag Berlin Heidelberg, 341–358. doi:10.1007/978-3-662-03059-2_17.
Hoondal, G.S.B., Kapoor, M., Mahajan, L. & Hoondal, G.S. (2001) Microbial xylanases and their industrial applications: a review. Applied
microbiology and biotechnology. 56 (3–4), 326–
338. doi:10.1007/s002530100704.
Huang, J.-S. (2001) Plant Pathogenesis And Resistance:
Biochemistry and Physiology of Plant-microbe Interactions. Netherlands: Springer. Chapter 2,
Degradation of Cell Walls by Plant Pathogens, Springer Science & Business Media.
Huang, Y., Busk, P.K. & Lange, L. (2015) Cellulose and Hemicellulose-degrading Enzymes in Fusarium Commune Transcriptome and Functional Characterization of Three Identified Xylanases.
Enzyme and Microbial Technology. 73, 9–19.
doi:10.1016/j.enzmictec.2015.03.001.
Hunter Jr, K.W., Gault, R.A. & Berner, M.D. (2002) Preparation of Microparticulate β-Glucan from Saccharomyces cerevisiae for use in Immune Potentiation. Letters in Applied Microbiology. 35
(4), 267–279.
doi:10.1046/j.1472-765X.2002.01201.x.
Keating, M.J., Holmes, R., Lerner, S. & Ho, D.H. (1993) L-asparaginase and PEG Asparaginase-Past, Present, and Future. Leukemia & Lymphoma. 10
(sup1), 153–157.
doi:10.3109/10428199309149129.
Kersten, P. & Cullen, D. (2007) Review of Extracellular Oxidative System of the Lignin-Degrading Basidiomycetes Phanerochaete chrysosporium.
Fungal Genetics and Biology. 44, 77–87.
Kubicek, C.P., Starr, T.L. & Glass, N.L. (2014) Plant Cell Wall-Degrading Enzymes and their Secretion in Plant-Pathogenic Fungi. Annual
Review of Phytopathology. 52, 427–451.
Lapmak, K., Lumyong, S., Thongkuntha, S., Wongputtisin, P. & Sardsud, U. (2010) L-Asparaginase Production by Biopolaris sp. BR 438 Isolated from Brown Rice in Thailand.
Chiang Mai J Sci. 37, 160–164.
Liao, C.-Y., Chen, M.-Y., Chen, Y.-K., Wang, T.-C., Sheu, Z.-M., Kuo, K.-C., Chang, P.-F.L., Chung, K.-R. & Lee, M.-H. (2012) Characterization of Three Colletotrichum Acutatum Isolates from Capsicum spp. European Journal of Plant
Pathology. 133 (3), 599–608. doi:10.1007/s10658-011-9935-7.
Marques, N.P., de Cassia Pereira, J., Gomes, E., da Silva, R., Araujo, A.R., Ferreira, H., Rodrigues, A., Dussan, K.J. & Bocchini, D.A. (2018) Cellulases and Xylanases Production by Endophytic Fungi by Solid State Fermentation using Lignocellulosic Substrates and Enzymatic Saccharification of Pretreated Sugarcane Bagasse. Industrial Crops and Products. 122, 66–75. doi:10.1016/j.indcrop.2018.05.022. McCredie, J.A., Inch, W.R., Kruuv, J. & Watson, T.A.
(1965) The Rate of Tumor Growth in Animals. Growth 29:331.
Medeiros, V.R., Mestdagh, F. & De Meulenaer, B. (2012) Acrylamide Formation in Fried Potato Products – Present and Future, Acritical Review on Mitigation Strategies. Food Chemistry. 133
(4), 1138–1154.
doi:10.1016/j.foodchem.2011.08.001.
Mitchell, L., Hoogendoorn, H., Giles, A.R., Vegh, P. & Andrew, M. (1994) Increased Endogenous Thrombin Generation in Children with Acute Lymphoblastic Leukemia: Risk of Thrombotic Complications in L-Asparaginase Induced Antithrombin III Deficiency. Blood. 83, 386– 391.
Mohan, K.N.S., Shimray, C.A., Indrani, D. & Manonmani, H.K. (2013) Reduction of Acrylamide Formation in Sweet Bread Withl-Asparaginase Treatment. Food and Bioprocess
Technology. 7 (3), 741–748. doi:10.1007/s11947-013-1108-6.
Mosier, N., Wyman, C., Dale, B., Elander, R., Lee, Y.Y., Holtzapple, M. & Ladisch, M. (2003) Features of Promising Technologies for Pretreatment of Lignocellulosic Biomass. Bioresource Technology. 96 (6), Purdue University, West
Lafayette, USA, 673–686.
Nakamura, C.T., Wilkinson, R. & Woodruff, K. (1999) Pancreatitis and Perotitis Following Therapy with L-asparaginase. International Pediatrics.
14, 25–27.
Ramya, L.N., Doble, M., Rekha, V.P.B. & Pulicherla, K.K. (2012) L-asparaginase as Potent Anti-leukemic Agent and its Significance of Having Reduced Glutaminase Side Activity for the Better Treatment of Acute Lymphoblastic Leukemia. Applied Biochemistry and Biotechnology. 167 (8), 2144–2159. doi:10.1007/s12010-012-9755-z.
Renato, J., Cavallazzi, P., Kasuya, C.M. & Soares, M.A. (2005) Screening of Inducers for Laccase Production by Lentinula edodes in Liquid Medium. Brazilian Journal of Microbiology. 36, 383–387.
Rossi, F., Incorvaia, C. & Mauro, M. (2004) Hypersensitivity Reactions to Antineoplastic Chemotherapeutic Agents. Recenti Prog Med. 95, 476–481.
Saikkonen, K., Faeth, S.H., Helander, M. & Sullivan, T.J. (1998) A Continuum of Interactions with The Host Plants. Annual Review of Ecology and
Systematic. 29, 319–343.
Saikkonen, K., Wäli, P., Helander, M. & Faeth, S.H. (2004) Evolution of Endophyte-Plant Symbioses.
Trends in Plant Sciencelant science. 9 (6), 275–
280. doi:10.1016/j.tplants.2004.04.005.
Sarquis, M.I. de M., Oliveira, E.M.M., Santos, A.S. & Costa, G.L. da (2004) Production of L-asparaginase by Filamentous Fungi. Memorias
do Instituto Oswaldo Cruz. 99 (5), 489–492.
doi:10.1590/S0074-02762004000500005. Shrivastava, A., Khan, A.A., Khurshid, M., Kalam,
M.A., Jain, S.K. & Singhal, P.K. (2015) Recent Developments in L-asparaginase Discovery and its Potential as Anticancer Agent. Critical
Reviews in Oncology/Hematology. 100, 1–10.
doi:10.1016/j.critrevonc.2015.01.002.
Sieber-Canavesi, F., Petrini, O. & Sieber, T.N. (1991) Endophytic Leptostroma Species on Picea abies, Abies alba and Abies balsamea: A Cultural, Biochemical and Numerical Study. Mycologia.
83 (1), 89–96.
doi:10.1080/00275514.1991.12025981.
Singh, R., Kumar, M., Mittal, A. & Mehta, P.K. (2016) Microbial Enzymes: Industrial Progress in 21st
Century. 3 Biotech. 6 (2), 174.
doi:10.1007/s13205-016-0485-8.
de Souza, P.M. & e Magalhaes, P. de O. (2010) Application of Microbial α-amylase in Industry - A Review. Brazilian Journal of Microbiology. 41
(4), 850–861.
Strobel, G. & Daisy, B. (2003) Bioprospecting for Microbial Endophytes and Their Natural Products. Microbiology and Molecular Biology
Reviews. 67 (4), 491–502. doi:10.1128/MMBR.67.4.491-502.20.
Sunitha, V.H., Nirmala Devi, D. & Srinivas, C. (2013) Extracellular Enzimatic Activity of Endophytic Fungal Strains Isolated from Medical Plant.
World Journal of Agricultural Sciences. 9 (1), 1–
9. doi:10.5829/idosi.wjas.2013.9.1.72148. Theantana, T., Hyde, K.D. & Lumyong, S. (2009)
Asparaginase Production by Endophytic Fungi from Thai Medicinal Plants: Cytoxicity Properties. Journal for Biologi Beyond Border. 7 (1), Pharmacogenomics Group, BMERC, 1–8. Upadhyay, P., Shrivastava, R. & Agrawal, P.K. (2016)
Bioprospecting and Biotechnological Applications of Fungal Laccase. 3 Biotech. 6 (1), 15.
Velho, A.C., Mondino, P. & Stadnik, M.J. (2018) Extracellular Enzymes of Colletotrichum fructicola Isolates Associated to Apple Bitter Rot and Glomerella Leaf Spot. Mycology. 9 (2), 145– 154. doi:10.1080/21501203.2018.1464525.
Venkatesagowda, B., Ponugupaty, E., Barbosa, A.M. & Dekker, R.F.H. (2012) Diversity of Plant Oil Seed-Associated Fungi Isolated from Seven Oil-Bearing Seeds and Their Potential for The Production of Lipolytic Enzymes. World Journal
of Microbiology and Biotechnology. 28 (1), 71–
80. doi:10.1007/s11274-011-0793-4.
Verma, N., Kumar, K., Kaur, G. & Anand, S. (2007) L-asparaginase: Apromising Chemotherapeutic Agent. Critical Reviews in Biotechnologyviews
in biotechnology. 27 (1), 45–62. doi:10.1080/07388550601173926.
Wahyuno, D., Amalia, N., Rossiana, N. & Bermawie, N. (2010) Respon Lima Aksesi Pegagan Terhadap Septoria centellae, Penyebab Bercak Daun.
Buletin Penelitian Tanaman Rempah dan Obat.
21 (2), 156–170.
Wood, P.J. & Weisz, J. (1984) Use of Calcuflour in Analysis of Oat Beta-D-Glucan. Cereal Chemistry. 6, 73–75.