Directory UMM :Data Elmu:jurnal:T:Tree Physiology:Vol16.1996:
Bebas
8
0
0
Teks penuh
Gambar
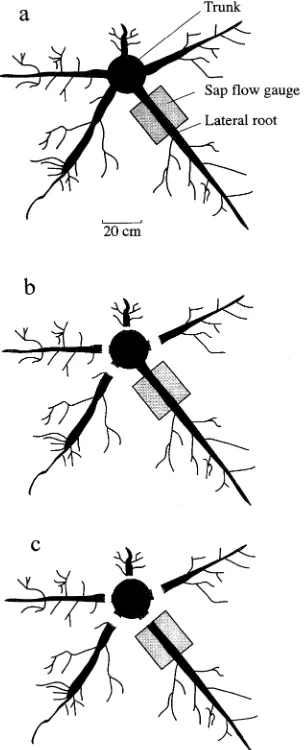
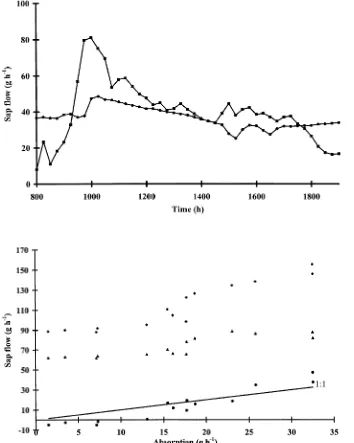
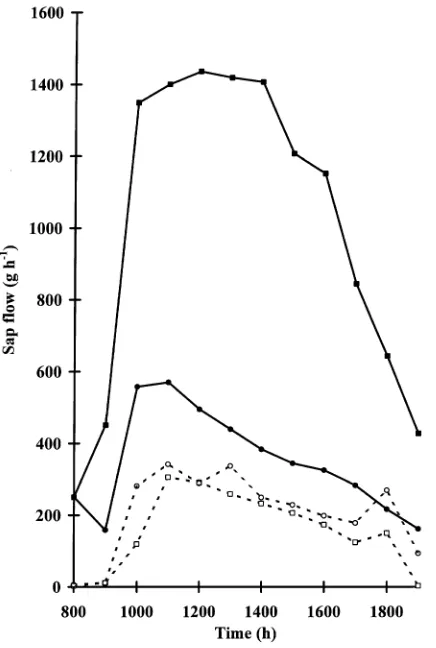
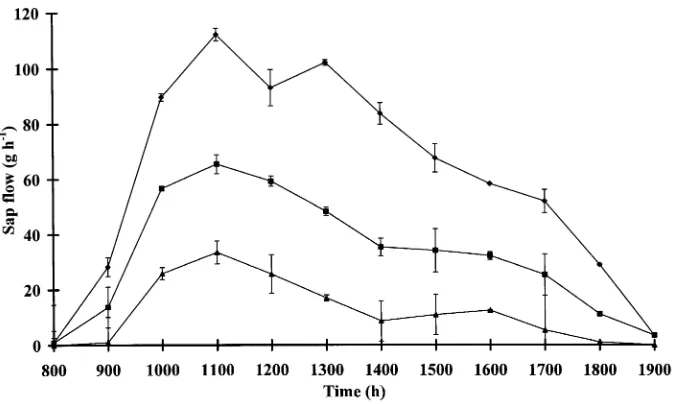
Dokumen terkait
Estimates of evapotranspiration (ET) from riparian vegetation along a 122 m reach of the San Pedro River using both a water balance approach and by scaling up sap flow measurements
Although we lack the data to fully validate these results, two comparisons show the promise of the piezometric weighing lysimeter method for estimat- ing the net moisture balance,
The station method for student laboratories is ideal for teaching large groups of students when a limited amount of equipment is available for the laboratory exercises. With
W e describe a novel student course in membrane physiology in which students record their own nasal potential difference, i.e., the transepithelial potential difference of
