KERAGAMAN GEN HORMON PERTUMBUHAN PADA
AYAM LOKAL INDONESIA DAN PERSILANGANNYA
RIA PUTRI RAHMADANI
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN
FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR
BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Keragaman Gen Hormon Pertumbuhan pada Ayam Lokal Indonesia dan Persilangannya adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Mei 2014 Ria Putri Rahmadani NIM D14100010
ABSTRAK
RIA PUTRI RAHMADANI. Keragaman Gen Hormon Pertumbuhan pada Ayam Lokal Indonesia dan Persilangannya. Dibimbing oleh CECE SUMANTRI dan SRI DARWATI.
Ayam lokal memiliki potensi genetik yang bernilai ekonomis tinggi seperti produksi daging dan telur, kemampuan bertahan terhadap iklim tropis yang lebih panas serta daya tahan terhadap penyakit. Gen hormon pertumbuhan pada ayam (chicken growth hormone atau cGH) memiliki peran yang penting dalam mengatur pertumbuhan dan metabolisme sehingga keragaman dari gen cGH berpotensi memiliki pengaruh terhadap sifat ekonomisnya. Penelitian ini bertujuan untuk mengidentifikasi keragaman gen cGH pada beberapa populasi ayam lokal Indonesia dan persilangannya serta ayam ras pedaging dan ras petelur. Keragaman gen cGH intron 4 dianalisa pada 229 sampel darah ayam yang terdiri dari ayam kampung, pelung, sentul, ras pedaging, ras petelur, merawang, ras pedaging x kampung, kampung x ras pedaging, sentul x kampung dan pelung x sentul. Primer khusus didesain untuk mengamplifikasi sekuen DNA target. Sekuen yang teramplifikasi dipotong menggunakan enzim restriksi MspI dan menghasilkan 2 alel yaitu A dan B dengan frekuensi 0.88-0.96 dan 0.04-0.12 kecuali pada ayam ras petelur hanya ditemukan 1 alel yaitu A dengan frekuensi 1.00. Hasil penelitian menunjukkan bahwa gen cGH|MspI pada ayam kampung, pelung, sentul, ras pedaging, merawang, ras pedaging x kampung, kampung x ras pedaging, sentul x kampung dan pelung x sentul bersifat polimorfik, sedangkan ayam ras petelur bersifat monomorfik. Populasi ayam lokal Indonesia dan persilangannya yang dianalisa pada penelitian ini memiliki keragaman genetik yang rendah namun tetap berada pada kesetimbangan Hardy-Weinberg.
Kata kunci: ayam lokal, cGH, keragaman, MspI, PCR-RFLP
ABSTRACT
RIA PUTRI RAHMADANI. Polymorphism of Growth Hormone Gene in Indonesia Local Chiken and The Crossbred. Supervised by CECE SUMANTRI and SRI DARWATI.
Indonesia local chickens have genetic potency with high ecomonic value such as meat and egg production, adapted in tropic and strong immunity. The chicken growth hormone (cGH) gene has a crucial role in controlling growth and metabolism, leading to potential correlations between cGH gene polymorphism and economic traits. This study aimed to identify the cGH gene polymorphism and find out the variation of gene at several local chickens, the crossbred, commercial meat type and layer. Polymorphism in intron 4 of cGH gene was studied in 229 chicken blood samples consisted of kampung, pelung, sentul, commercial meat type, layer, merawang, commercial meat type x kampung, kampung x commercial meat type, sentul x kampung and pelung x sentul chickens. A specific primer set was used to amplify a fragment of growth hormone locus
using PCR. PCR products were digested with MspI restriction endonucleases. The amplified fragment digested with MspI enzyme revealed 2 alleles A and B with the frequency of 0.88-0.96 and 0.04-0.12, except in layer chickens only had 1 allele A with the frequency of 1.00. The results showed that cGH gene in MspI locus at kampung, pelung, sentul, commercial meat type, merawang, commercial meat type x kampung, kampung x commercial meat type, sentul x kampung and pelung x sentul chickens were polymorphics, except the layer chickens was monomorphics. The population of Indonesia local chicken and the crossbred that was studied in this research had low heterozygosity value but it was still in the Hardy-Weinberg equilibrium.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Peternakan
pada
Departemen Ilmu Produksi dan Teknologi Peternakan
RIA PUTRI RAHMADANI
KERAGAMAN GEN HORMON PERTUMBUHAN PADA
AYAM LOKAL INDONESIA DAN PERSILANGANNYA
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN
FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR
BOGOR
Judul Skripsi : Keragaman Gen Hormon Pertumbuhan pada Ayam Lokal Indonesia dan Persilangannya
Nama : Ria Putri Rahmadani NIM : D14100010
Disetujui oleh
Pembimbing I
Prof Dr Ir Cece Sumantri, MAgrSc
Pembimbing II Dr Ir Sri Darwati, MSi
Diketahui oleh
Ketua Departemen Prof Dr Ir Muladno, MSA
PRAKATA
Puji dan syukur kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga skripsi ini berhasil diselesaikan. Tema yang dipilih pada penelitian yang dilaksanakan sejak bulan Desember 2013 hingga Februari 2014 ini ialah ayam lokal Indonesia, dengan judul Keragaman Gen Hormon Pertumbuhan pada Ayam Lokal Indonesia dan Persilangannya.
Terima kasih penulis ucapkan kepada Prof Dr Ir Cece Sumantri, MAgrSc dan Dr Ir Sri Darwati, MSi selaku dosen pembimbing. Di samping itu, penghargaan penulis sampaikan kepada Pipih Suningsih Effendi, Amd SPt, Eryk Andreas, SPt MSi, Shelvi, SSi, Dadang dan Ilyas dari Laboratorium Pemuliaan dan Genetika Ternak, yang telah membantu selama penelitian berlangsung. Ungkapan terima kasih juga disampaikan kepada mama (Sri Maharani), papa (Syaiful Firmadi), oma (Warnius), opa (Murice), adik-adik (Irfan dan Adit) serta seluruh keluarga atas segala doa dan kasih sayangnya. Terima kasih kepada teman-teman IPTP 47 khususnya sahabat terbaik Jafar, Laras, Anita, Ica, Ishfi, Fender, Angga, Nenik, Puspita, Nisa, Dita dan Lita atas segala bantuan dan dukungannya. Terima kasih juga kepada Direktorat Pendidikan Tinggi (DIKTI) atas beasiswa Bidik Misi yang telah diperoleh selama 8 semester. Semoga hasil penelitian ini bermanfaat.
Bogor, Mei 2014 Ria Putri Rahmadani
DAFTAR ISI
DAFTAR TABEL viii
DAFTAR GAMBAR viii
DAFTAR LAMPIRAN viii
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 1
Ruang Lingkup Penelitian 2
METODE 2
Waktu dan Tempat Penelitian 2
Materi 2
Prosedur 3
Analisis Data 4
HASIL DAN PEMBAHASAN 5
Amplifikasi Gen Growth Hormone (GH|MspI) 5
Frekuensi Alel dan Frekuensi Genotip GH|MspI 8
Heterozigositas 9
Kesetimbangan Hardy-Weinberg 10
SIMPULAN DAN SARAN 10
DAFTAR PUSTAKA 11
LAMPIRAN 12
DAFTAR TABEL
1 Frekuensi alel dan frekuensi genotip gen cGH pada ayam lokal Indonesia 8 2 Nilai heterozigositas harapan dan nilai heterozigositas pengamatan 9 3 Hasil analisa keseimbangan Hardy-Weinberg dengan uji chi-kuadrat 9
DAFTAR GAMBAR
1 Visualisasi hasil amplifikasi gen cGH ayam lokal Indonesia dengan
panjang 367 pb pada gel agarose 1.5% 5
2 Posisi penempelan primer dan titik potong enzim MspI pada sekuen gen cGH berdasarkan Gen Bank (nomor akses: D10484.1) 6 3 Visualisasi hasil PCR-RFLP gen cGH|MspI ayam lokal Indonesia pada
gel agarose 2% dengan genotip AA (367 pb), BB (259 pb, 108 pb) dan
PENDAHULUAN
Latar Belakang
Ayam lokal memiliki potensi genetik yang bernilai ekonomis tinggi seperti produksi daging dan telur dengan segmentasi pasar yang khusus, kemampuan bertahan terhadap iklim tropis yang lebih panas serta sistem imun yang lebih baik. Pemanfaatan ayam lokal di Indonesia sampai saat ini masih terfokus pada taraf budidaya sehingga ayam lebih banyak digunakan sebagai final stock penghasil daging dan telur. Belum banyak upaya yang serius dan kontinyu untuk memanfaatkan ayam lokal sebagai bahan baku genetik guna membentuk ayam unggul. Penelitian yang intensif terhadap ayam lokal melalui program pemuliaan sifat-sifat ekonomis disertai dengan seleksi yang terarah akan mampu meningkatkan mutu genetiknya.
Identifikasi keragaman suatu gen dilakukan untuk memperoleh marka genetik yang dapat mempermudah seleksi secara molekuler dan salah satu metodenya adalah PCR-RFLP. PCR (polymerase chain reaction) merupakan reaksi in vitro untuk menggandakan jumlah molekul DNA menggunakan enzim dan oligonukleotida sebagai primer pada mesin thermal cycler (Muladno 2010). Green (1998) menyatakan bahwa RFLP merupakan fragmen DNA hasil pemotongan enzim endonuklease untuk berbagai individu. Marker DNA RFLP memiliki tingkat polimorfisme yang tinggi dan digunakan untuk mengidentifikasi gen-gen yang mengkode sifat-sifat penting, salah satunya gen hormon pertumbuhan.
Gen hormon pertumbuhan pada ayam atau dikenal dengan chicken growth hormone (cGH) merupakan salah satu gen penting yang dapat mempengaruhi performa ayam karena fungsinya yang penting dalam pertumbuhan dan metabolisme. Gen cGH terletak pada kromosom lq4 dan memiliki panjang 3 901 pb yang terdiri dari 5 ekson dan 4 intron (Tanaka et al. 1992). Nilai ekonomis dari gen cGH berdasarkan analisa Kuhnlein et al. (1997) pada 12 bangsa ayam White Leghorn non inbreed menggunakan PCR-RFLP dengan 3 situs MspI dan 1 situs SacI dilaporkan terdapat alel yang berlokasi pada intron 1, 3 dan 4 yang berpengaruh terhadap produksi telur serta resisten terhadap penyakit marek dan avian leukosis. Makhsous et al. (2013) menemukan 1 lokus SacI pada gen cGH intron 4 dalam populasi ayam lokal Iran yang berasosiasi dengan produksi telur dan laju bertelur. Biangxue et al. (2003) juga menemukan 1 lokus MspI pada gen cGH intron 4 dalam populasi ayam lokal China yang berasosiasi dengan kecepatan pertumbuhan otot dada dan lemak abdominal. Oleh karena itu, gen cGH dapat digunakan sebagai gen penanda genetik pada proses seleksi untuk meningkatkan performa ayam.
Tujuan Penelitian
Penelitian ini bertujuan mengidentifikasi keragaman gen cGH intron 4 yang terletak pada kromosom lq4 pada ayam lokal Indonesia dan persilangannya serta ayam ras pedaging dan ras petelur sebagai kontrol dengan metode PCR-RFLP.
2
Ruang Lingkup Penelitian
Identifikasi keragaman gen cGH ayam lokal Indonesia dan persilangannya serta ayam ras pedaging dan ras petelur pada intron 4 menggunakan metode PCR-RFLP dengan enzim restriksi MspI.
METODE
Waktu dan Tempat Penelitian
Penelitian dilakukan di Laboratorium Genetika Molekuler Ternak, Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor. Penelitian dilaksanakan selama 3 bulan dari Desember 2013 sampai Februari 2014.
Materi Sampel
Sampel yang digunakan sebagai sumber DNA adalah darah ayam sebanyak 229 sampel yang berasal dari 10 jenis ayam yaitu ayam kampung (29 sampel), pelung (5 sampel), sentul (32 sampel), ras pedaging (11 sampel), ras petelur (14 sampel), merawang (23 sampel), ras pedaging x kampung (12 sampel), kampung x ras pedaging (33 sampel), sentul x kampung (50 sampel) dan pelung x sentul (20 sampel). Ayam merupakan koleksi dari Laboratorium Pemuliaan dan Genetika Ternak, Fakultas Peternakan, Institut Pertanian Bogor.
Bahan dan Alat
Bahan yang dibutuhkan untuk pengambilan sampel darah yaitu EDTA, alkohol 70% dan kapas. Alat yang dibutuhkan yaitu tabung ependorf 1.5 mL, spuit 3 mL dan spidol permanen.
Bahan yang dibutuhkan untuk ekstraksi DNA yaitu sampel darah, NaCl 0.2%, SDS (sodium dodesil sulfat) 10%, proteinase 5 mg mL-1, 1 x STE (sodium tris EDTA), fenol, CIAA (klorofom iso amil alkohol), NaCl 5M, ETOH (etanol alkohol) absolut, ETOH 70% dan TE (tris elusion) 80%. Alat yang dibutuhkan yaitu tabung ependorf 1.5 mL, 1 set pipet mikro beserta tipnya, vortex, centrifuge, inkubator dan freezer.
Bahan yang dibutuhkan untuk amplifikasi DNA yaitu sampel DNA, primer forward 5’-GCCTGGGAGCAAACAAACCC-3’ dan reverse 5’-CCATGACACT TCAGCTGCAGC-3’, DW (air destilasi) dan GoTaq® Green Master Mix. Alat yang dibutuhkan yaitu 1 set pipet mikro beserta tipnya dan mesin thermal cycler. Bahan yang dibutuhkan untuk pemotongan DNA yaitu DNA hasil PCR, DW, buffer tango dan enzim restriksi MspI. Alat yang dibutuhkan yaitu 1 set pipet mikro beserta tipnya dan inkubator.
Bahan yang dibutuhkan untuk elektroforesis yaitu agarose, 0.5 x TBE (tris borat EDTA), EtBr (etidium bromid) dan DNA pengukur yang berukuran 100 pb. Alat yang dibutuhkan yaitu timbangan, pipet mikro beserta tipnya, gelas piala,
3 microwave, hotplate, tray pencetak gel, tank elektroforesis (mupid), power supply 100 V dan UV transilluminator dilengkapi kamera polaroid.
Prosedur Pengambilan Sampel Darah
Ayam dikondisikan di lokasi pengambilan darah. Bagian sayap kiri ayam di sekitar vena axillaris dibersihkan dari bulu dan kotoran menggunakan alkohol 70% kemudian darah diambil menggunakan spuit.
Darah yang telah diambil dimasukkan ke dalam tabung ependorf 1.5 mL yang telah diisi dengan EDTA. Darah dihomogenkan segera agar tidak beku. Sampel darah kemudian disimpan dalam refrigerator dengan suhu 4 ºC.
Ekstraksi DNA
Metode ekstraksi DNA berdasarkan Sambrook et al. (1989) yaitu tabung ependorf 1.5 mL yang akan digunakan diberi label terlebih dahulu. Sebanyak 20 µL sampel darah dimasukkan ke dalam tabung ependorf 1.5 mL, kemudian ditambahkan 1 000 µL NaCl 0.2%, dihomogenkan, didiamkan selama 5 menit lalu disentrifugasi pada kecepatan 8 000 rpm selama 5 menit pada suhu ruang. Bagian supernatan yang terbentuk dibuang.
Sebanyak 40 µL SDS 10%, 10 µL proteinase 5 mg mL-1 dan 1 x STE ditambahkan sampai 400 µL kemudian dikocok pelan dalam inkubator suhu 55 ºC selama 2 jam. Sebanyak 400 µL fenol, 400 µL CIAA dan 40 µL NaCl 5 M ditambahkan lalu dikocok pada suhu ruang selama 1 jam kemudian disentrifugasi pada kecepatan 12 000 rpm selama 5 menit. Bagian DNA (bening) dipindahkan ke dalam tabung ependorf baru menggunakan pipet sebanyak 400 µL. Sebanyak 800 µL ETOH absolut dan 40 µL NaCl 5M ditambahkan ke dalam tabung lalu dilakukan freezing over night.
Sampel yang telah didiamkan dalam freezer kemudian didiamkan pada suhu ruang lalu disentrifugasi pada kecepatan 12 000 rpm selama 5 menit. Bagian supernatan dibuang lalu ditambahkan 800 µL ETOH 70% dan disentrifugasi pada kecepatan 12 000 rpm selama 5 menit kemudian bagian supernatan dibuang kembali. Sampel didiamkan di ruang terbuka sampai alkohol kering lalu sebanyak 100 µL TE 80% ditambahkan kemudian DNA dapat disimpan dalam freezer sampai akan digunakan.
Polymerase Chain Reaction-Restriction Fragment Length Polymorphism (PCR-RFLP)
Primer diencerkan menggunakan TE primer sesuai komposisi yag ditentukan. Sebanyak 25 µL forward dicampur degan 25 µL reverse lalu ditambahkan 50 µL DW dalam tabung ependorf 1.5 mL kemudian disentrifugasi. Cocktail larutan PCR dibuat dengan campuran primer, GoTaq® Green Master Mix dan DW sesuai dengan perhitungan jumlah sampel yang akan diamplifikasi lalu cocktail dihomogenkan dan disentrifugasi. Sampel dimasukkan ke dalam tabung 0.2 mL sebanyak 2 µL lalu ditambahkan cocktail sebanyak 14 µL kemudian dimasukkan ke dalam mesin thermal cycler dengan suhu penempelan primer 60 ºC.
4
Cocktail larutan untuk pemotongan dibuat dengan campuran DW, buffer tango dan enzim restriksi MspI sesuai dengan perhitungan jumlah sampel yang akan dipotong lalu dihomogenkan dan disentrifugasi. Sampel sebanyak 5 µL dimasukkan ke dalam tabung 0.5 mL lalu ditambahkan cocktail sebanyak 2 µL lalu disentrifugasi dan diinkubasi selama 16 jam pada suhu 37 ºC.
Elektroforesis
Hasil ekstraksi, amplifikasi dan pemotongan dielektroforesis dengan persentase gel agarose yang berbeda. Sampel hasil ekstraksi dielektroforesis dengan gel agarose 1% yang terdiri dari 0.2 g agarose, 20 mL TBE dan 1.8 µL EtBr. Hasil amplifikasi dielektroforesis dengan gel agarose 1.5% yang terdiri dari 0.3 g agarose, 20 mL TBE dan 1.8 µL EtBr.
Hasil pemotongan dielektroforesis dengan gel agarose 2% yang terdiri dari 0.4 g agarose, 20 mL TBE dan 1.8 µL EtBr. Gel ditunggu hingga mengeras selama 10 menit. Sebanyak 5 µL sampel yang akan dielektroforesis dimasukkan ke dalam sumur sel. Sumur yang pertama diisi DNA pengukur yang berukuran 100 pb. Elektroforesis dilakukan menggunakan mupid dengan arus 100 V selama 30 menit lalu divisualisasi menggunakan UV transilluminator.
Analisis Data Frekuensi Genotip dan Frekuensi Alel
Frekuensi genotip (xii) dapat diketahui melalui perbandingan jumlah genotip tertentu pada setiap sampel (Nei dan Kumar 2000) dengan rumus sebagai berikut:
xii = nNii
Frekuensi alel (xi) adalah rasio relatif suatu alel terhadap keseluruhan alel pada suatu lokus dalam populasi, dengan rumus sebagai berikut:
xi =2nii + 2N∑nij
Keterangan:
xii = frekuensi genotip ke- ii
xi = frekuensi alel ke- i
nii = jumlah individu bergenotip ii
nij = jumlah individu bergenotip ij
N = jumlah sampel
Heterozigositas
Keragaman genetik diketahui melalui perkiraan heterozigositas pengamatan yang diperoleh dengan rumus Weir (1996) sebagai berikut:
Ho = ∑ nij
N i≠j
Keterangan:
Ho = heterozigositas pengamatan
nij = jumlah individu heterozigot
N = jumlah individu yang diamati
Heterozigositas harapan (He) berdasarkan frekuensi alel (Nei dan Kumar 2000) dan dihitung menggunakan rumus sebagai berikut:
5
He = 1− ∑q xi2
i=1
Keterangan:
He = nilai heterozigositas harapan
xi² = frekuensi alel ke- i
q = jumlah alel
Keseimbangan Hardy-Weinberg
Keragaman Hardy-Weinberg dapat diketahui dengan menggunakan chi-kuadrat (Hartl dan Clark 1997):
X2 =∑(O−E)2 E
Keterangan:
X2 = uji chi-kuadrat
O = jumlah genotip pengamatan E = jumlah genotip harapan
HASIL DAN PEMBAHASAN
Amplifikasi Gen Growth Hormone (GH|MspI)
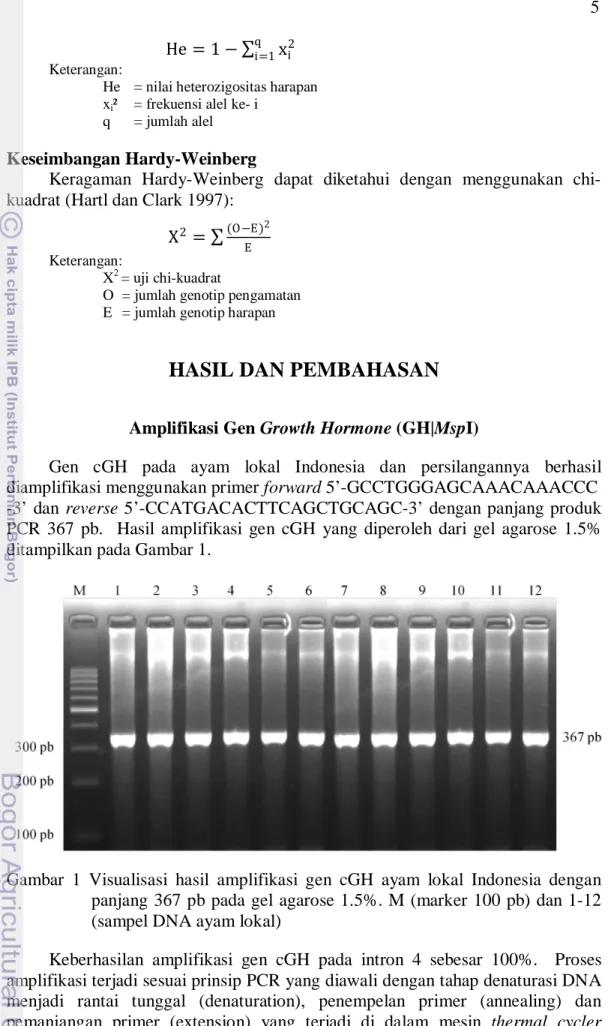
Gen cGH pada ayam lokal Indonesia dan persilangannya berhasil diamplifikasi menggunakan primer forward 5’-GCCTGGGAGCAAACAAACCC -3’ dan reverse 5’-CCATGACACTTCAGCTGCAGC-3’ dengan panjang produk PCR 367 pb. Hasil amplifikasi gen cGH yang diperoleh dari gel agarose 1.5% ditampilkan pada Gambar 1.
Gambar 1 Visualisasi hasil amplifikasi gen cGH ayam lokal Indonesia dengan panjang 367 pb pada gel agarose 1.5%. M (marker 100 pb) dan 1-12 (sampel DNA ayam lokal)
Keberhasilan amplifikasi gen cGH pada intron 4 sebesar 100%. Proses amplifikasi terjadi sesuai prinsip PCR yang diawali dengan tahap denaturasi DNA menjadi rantai tunggal (denaturation), penempelan primer (annealing) dan pemanjangan primer (extension) yang terjadi di dalam mesin thermal cycler
6
(Muladno 2010). Umumnya sebelum proses PCR dimulai, dilakukan pre denaturasi yang pada penelitian ini dilakukan pada suhu 95 ºC selama 5 menit. Tujuan pre denaturasi adalah untuk meyakinkan bahwa molekul DNA target yang ingin dilipatgandakan jumlahnya benar-benar terdenaturasi (Sulandari dan Zein 2003).
Proses selanjutnya yaitu denaturasi DNA pada suhu 95 ºC selama 10 detik. Denaturasi adalah proses pembelahan untai ganda DNA menjadi untai tunggal. Suhu umum untuk denaturasi DNA berkisar 94-95 ºC selama 30 detik sesuai dengan banyaknya nukleotida G/C. Apabila DNA target mengandung banyak nukleotida G/C maka suhu denaturasi dapat ditingkatkan (Sulandari dan Zein 2003). Proses berikutnya adalah penempelan primer pada suhu yang lebih rendah yaitu 60 ºC selama 20 detik. Suhu penempelan primer pada penelitian ini berbeda dengan Makhsous et al. (2013) yaitu pada suhu 62 ºC untuk primer forward dan 54 ºC untuk primer reverse selama 30 detik. Hal ini dikarenakan produk PCR pada penelitian ini lebih pendek dibandingkan produk PCR pada penelitian Makhsous et al. (2013) yaitu 367 pb. Suhu penempelan primer ditentukan oleh panjang primer yang digunakan, semakin panjang primer maka semakin tinggi suhu penempelan primernya (Sulandari dan Zein 2003). Posisi penempelan primer pada sekuen gen cGH berdasarkan Gen Bank (nomor akses: D10484.1) ditampilkan pada Gambar 2.
1 gcctgggagc aaacaaaccc
51 gtggtgcccc atccctggag gtcccaaggc catggatgga gccctgggca
tccgtcctga cattttcgat cagaaaagtt
101 gtttgagctg gtggggggc cccagcccca tggccagggg gtgagacggg
151 gggggctctg agatcccttc caacacaacc atgctgtgat tccacagccc
201 tgcaatcctt gaaggtccct tccaacccaa cggtgccacg attccatggt
251 ttctgtgatc cttaaggtcc cttccaaccc aaccatgcca tcattccatg
301 gttccatgat ctcagaggcc ccttcccacc caaccacgcc gtgatt
351
ccat gacacttcag ctgcagc
forward : 5’-gcctgggagcaaacaaaccc-3’ reverse : 5’-ccatgacacttcagctgcagc-3’
Gambar 2 Posisi penempelan primer dan titik potong enzim MspI pada sekuen gen cGH berdasarkan Gen Bank (nomor akeses: D10484.1)
Primer yang telah menempel pada target kemudian mengalami ekstensi atau pemanjangan pada suhu 72 ºC selama 30 detik. Pemanjangan terjadi karena primer dengan bantuan enzim DNA polimerase membentuk untaian DNA yang sesuai dengan runutan DNA yang terbelah dengan kecepatan penyusunan nukleotida antara 35-100 nukleotida per detik (Sulandari dan Zein 2003). Proses akhir yaitu pos ekstensi dengan suhu 72 ºC selama 5 menit untuk memastikan bahwa semua hasil PCR berbentuk untai ganda. Ketiga tahapan utama yang terdiri dari denaturasi, penempelan dan pemanjangan primer merupakan 1 siklus termal. Jumlah siklus termal yang dilakukan pada penelitian ini adalah 35 siklus sehingga jumlah fragmen yang diamplifikasi adalah 235.
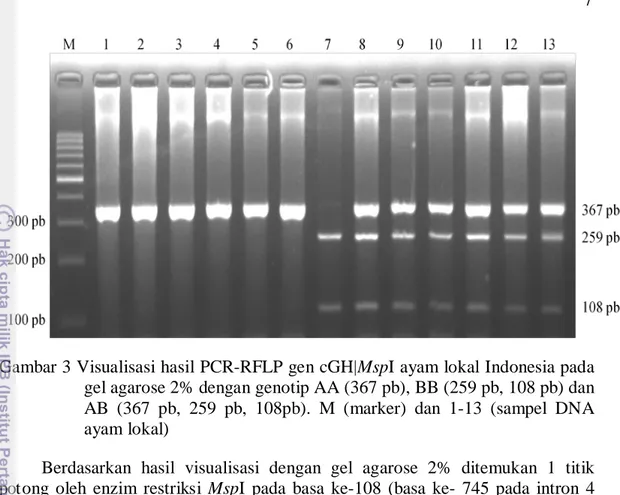
Pemotongan fragmen DNA selanjutnya dilakukan menggunakan enzim MspI dengan situs potong c|cgg. Suhu inkubasi yang digunakan adalah 37 ºC selama 16 jam. Hasil visualiasi PCR-RFLP pada gen cGH yang terlihat pada gel agarose 2% ditampilkan pada Gambar 3.
7
Gambar 3 Visualisasi hasil PCR-RFLP gen cGH|MspI ayam lokal Indonesia pada gel agarose 2% dengan genotip AA (367 pb), BB (259 pb, 108 pb) dan AB (367 pb, 259 pb, 108pb). M (marker) dan 1-13 (sampel DNA ayam lokal)
Berdasarkan hasil visualisasi dengan gel agarose 2% ditemukan 1 titik potong oleh enzim restriksi MspI pada basa ke-108 (basa ke- 745 pada intron 4 atau basa ke- 3 197 pada sekuen lengkap gen cGH berdasarkan Tanaka et al. (1992). Hal ini sesuai dengan penelitian yang dilakukan oleh Makhsous et al. (2013) pada ayam petelur Iran yaitu terjadi pemotongan oleh enzim restriksi MspI pada basa ke- 3 093, basa ke- 3 189, basa ke- 3 197 dan basa ke- 3 293 dari sekuen lengkap gen cGH. Titik potong enzim MspI yang ditemukan pada penelitian ini berada pada basa ke- 3 197 dengan produk PCR berada pada basa ke- 3 091 sampai basa ke- 3 452 dari sekuen lengkap gen cGH.
Hasil genotyping menunjukkan bahwa terjadi mutasi pada basa ke- 109. Mutasi yang terjadi adalah mutasi transisi yaitu perubahan basa antar basa purin atau antar basa pirimidin (Hardjosubroto 2001). Mutasi yang ditemukan pada penelitian ini adalah perubahan basa pirimidin yaitu timin (T) menjadi sitosin (C).
Penelitian ini menemukan 3 macam genotip yaitu AA, BB dan AB, dengan 2 alel yaitu A dan B. Alel A jika tidak terpotong dan alel B jika ada 1 titik potong pada basa ke- 108 sehingga muncul 2 pita pada basa ke- 108 dan basa ke- 259. Genotip AA diidentifikasi jika tidak ada pemotongan oleh enzim MspI sehingga hanya muncul 1 pita hasil produk PCR dengan panjang 367 pb. Genotip BB jika ada 1 titik potong pada basa ke- 108 sehingga akan muncul 2 pita yang berada pada basa ke- 259 dan basa ke- 108. Genotip AB ditandai dengan munculnya 3 pita pada basa ke- 367, basa ke- 259 dan basa ke- 108. Ayam dengan genotip AA dan BB disebut homozigot. Hal ini menunjukkan bahwa kedua tetuanya mewariskan alel yang sama yaitu kedua tetua mewariskan alel A pada individu yang bergenotip AA, demikian halnya untuk individu yang bergenotip BB memperoleh alel B dari kedua tetuanya. Individu yang bergenotip AB disebut heterozigot. Hal ini menunjukkan bahwa kedua tetuanya mewariskan alel yang berbeda yaitu alel A dan B (Noor 2010).
8
Frekuensi Alel dan Frekuensi Genotip GH|MspI
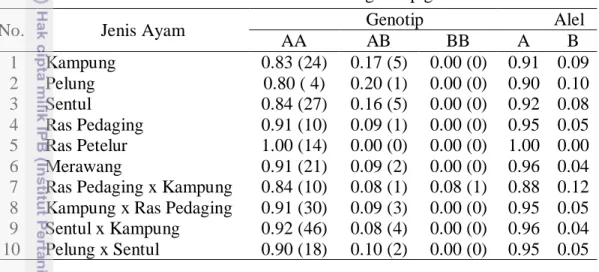
Berdasarkan hasil genotyping diketahui bahwa sampel yang dianalisa yaitu ayam kampung, pelung, sentul, ras pedaging, merawang, ras pedaging x kampung, kampung x ras pedaging, sentul x kampung dan pelung x sentul memiliki alel polimorfik, sedangkan ayam ras petelur memiliki alel monomorfik. Berikut hasil perhitungan frekuensi genotip dan frekuensi alel dari semua sampel ayam disajikan pada Tabel 1.
Tabel 1 Frekuensi alel dan frekuensi genotip gen cGH
No. Jenis Ayam Genotip Alel
AA AB BB A B 1 Kampung 0.83 (24) 0.17 (5) 0.00 (0) 0.91 0.09 2 Pelung 0.80 ( 4) 0.20 (1) 0.00 (0) 0.90 0.10 3 Sentul 0.84 (27) 0.16 (5) 0.00 (0) 0.92 0.08 4 Ras Pedaging 0.91 (10) 0.09 (1) 0.00 (0) 0.95 0.05 5 Ras Petelur 1.00 (14) 0.00 (0) 0.00 (0) 1.00 0.00 6 Merawang 0.91 (21) 0.09 (2) 0.00 (0) 0.96 0.04 7 Ras Pedaging x Kampung 0.84 (10) 0.08 (1) 0.08 (1) 0.88 0.12 8 Kampung x Ras Pedaging 0.91 (30) 0.09 (3) 0.00 (0) 0.95 0.05 9 Sentul x Kampung 0.92 (46) 0.08 (4) 0.00 (0) 0.96 0.04 10 Pelung x Sentul 0.90 (18) 0.10 (2) 0.00 (0) 0.95 0.05 Hasil analisa menunjukkan bahwa semua populasi ayam memiliki 2 genotip yaitu AA dan AB kecuali ayam ras petelur yang hanya memiliki 1 genotip yaitu AA dan ayam ras pedaging x kampung yang memiliki 3 genotip yaitu AA, AB dan BB. Munculnya genotip BB pada 1 individu ayam ras pedaging x kampung dapat terjadi karena individu tersebut memperoleh alel yang berbeda dari kedua tetuanya (Noor 2010).
Frekuensi genotip merupakan proporsi suatu genotip relatif terhadap semua genotip yang ada dalam suatu populasi (Noor 2010). Hardjosubroto (2001) menjelaskan bahwa frekuensi genotip adalah peluang munculnya genotip tersebut dalam populasi. Secara keseluruhan, frekuensi genotip AA pada semua populasi ayam lebih besar daripada frekuensi genotip AB. Hasil analisa ini sejalan dengan hasil analisa Makhsous et al. (2013) bahwa frekuensi genotip AA pada ayam lokal Iran tipe petelur juga lebih besar dibandingkan genotip lainnya yaitu 0.338. Merujuk pada pengertian frekuensi genotip menurut Hardjosubroto (2001), maka peluang munculnya genotip AA pada ayam lokal Indonesia dan persilangannya yang dianalisa pada penelitian ini lebih besar dibandingkan genotip AB.
Suatu alel bersifat polimorfik apabila memiliki frekuensi alel sama dengan atau kurang dari 0.99 (Nei 1987). Berdasarkan hasil genotyping diketahui bahwa nilai frekuensi alel A pada semua populasi (kecuali ayam ras petelur) berkisar antara 0.88-0.96, sedangkan nilai frekuensi alel B berkisar antara 0.04-0.12. Hal ini menunjukkan bahwa gen cGH pada lokus MspI bersifat polimorfik dan dapat dipergunakan sebagai marka seleksi pertumbuhan dan produksi, kecuali pada ayam ras petelur bersifat monomorfik karena nilai frekuensi alelnya mencapai 1.00. Penelitian yang dilakukan oleh Makhsous et al. (2013) terhadap ayam lokal Iran juga menunjukkan bahwa frekuensi alel A lebih besar dibandingkan alel B
9 dan C yaitu sebesar 0.599. Hasil penelitian Nie et al. (2002) pada 20 populasi ayam lokal China menunjukkan bahwa frekuensi alel A lebih besar dibandingkan alel B, C, D dan E yaitu bekisar antara 0.455-0.948. Penelitian lain yang dilakukan oleh Bingxue et al. (2003) menemukan bahwa terdapat perubahan basa timin (T) menjadi sitosin (C) pada gen cGH|MspI intron 4 dari populasi ayam persilangan ras pedaging dan silky. Mutasi ini berasosiasi dengan pertumbuhan otot dada dan lemak abdominal. Individu yang bergenotip BB (mutasi) memiliki laju pertumbuhan dan bobot otot dada yang lebih tinggi (6.64%; 105.78 g) dibandingkan individu bergenotip AB dan AA, sedangkan individu bergenotip AB memiliki laju pertumbuhan dan bobot otot dada yang lebih tinggi (6.59%; 103.25 g) dibandingkan individu bergenotip AA (6.22%; 93.06 g). Kecepatan pertumbuhan otot dada berkorelasi negatif dengan kecepatan pertumbuhan lemak abdominal. Individu yang bergenotip BB memiliki kecepatan pertumbuhan lemak abdominal terendah (1.98%) diikuti oleh genotip AB (2.66%) dan genotip AA (3.21%).
Heterozigositas
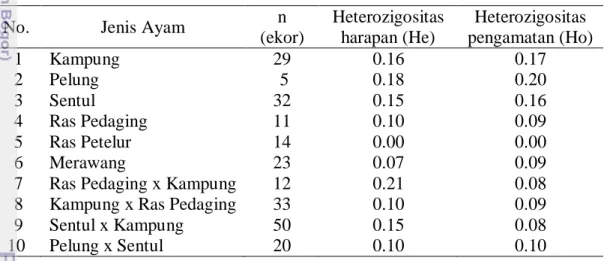
Heterozigositas dapat digunakan untuk mengukur keragaman genetik suatu populasi. Nilai heterozigositas dari populasi ayam yang dianalisa pada penelitian ini disajikan pada Tabel 2.
Tabel 2 Nilai heterozigositas harapan dan nilai heterozigositas pengamatan
No. Jenis Ayam n
(ekor) Heterozigositas harapan (He) Heterozigositas pengamatan (Ho) 1 Kampung 29 0.16 0.17 2 Pelung 5 0.18 0.20 3 Sentul 32 0.15 0.16 4 Ras Pedaging 11 0.10 0.09 5 Ras Petelur 14 0.00 0.00 6 Merawang 23 0.07 0.09
7 Ras Pedaging x Kampung 12 0.21 0.08
8 Kampung x Ras Pedaging 33 0.10 0.09
9 Sentul x Kampung 50 0.15 0.08
10 Pelung x Sentul 20 0.10 0.10
Nilai heterozigositas penting diketahui untuk mendapatkan gambaran keragaman genetik suatu populasi. Nilai heterozigositas dipengaruhi oleh beberapa faktor yaitu jumlah sampel, jumlah alel dan frekuensi alel (Allendorf dan Luikart 2006). Berdasarkan hasil perhitungan yang ditampilkan pada Tabel 2 diketahui bahwa nilai heterozigositas harapan (He) dan heterozigositas pengamatan (Ho) pada semua populasi ayam tidak berbeda. Nilai Ho pada semua populasi ayam (kecuali ayam ras petelur) yang dianalisa pada penelitian ini berada pada kisaran 0.08-0.17, berarti nilai heterozigositas berada di bawah 0.5 dan menggambarkan rendahnya keragaman genetik pada populasi tersebut (Allendorf dan Luikart 2006).
10
Kesetimbangan Hardy-Weinberg
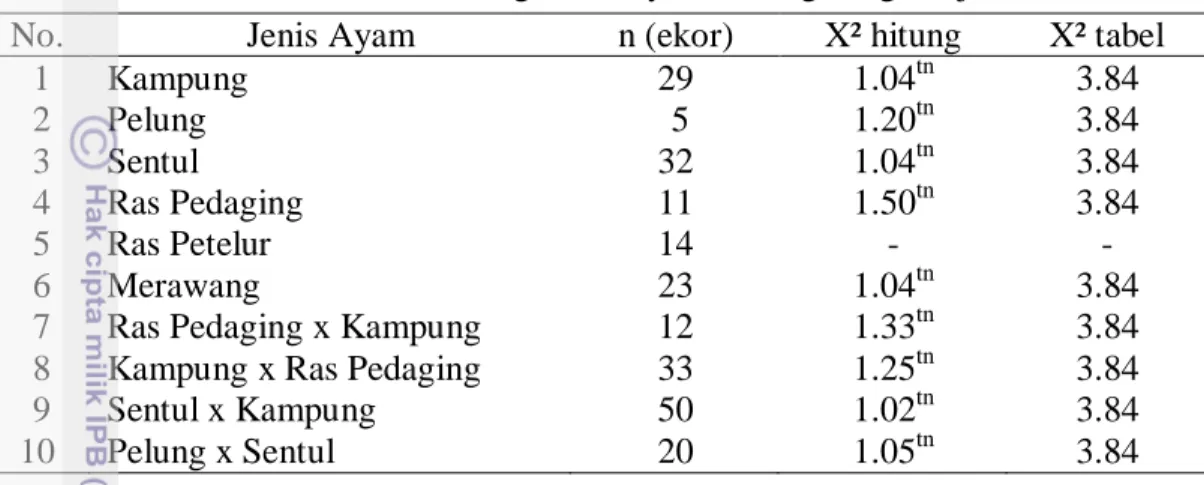
Analisa kesetimbangan Hardy-Weinberg dilakukan menggunakan uji chi-kuadrat. Perhitungan Hardy-Weinberg disajikan pada Tabel 3.
Tabel 3 Hasil analisa kesetimbangan Hardy-Weinberg dengan uji chi-kuadrat
No. Jenis Ayam n (ekor) X² hitung X² tabel
1 Kampung 29 1.04tn 3.84 2 Pelung 5 1.20tn 3.84 3 Sentul 32 1.04tn 3.84 4 Ras Pedaging 11 1.50tn 3.84 5 Ras Petelur 14 - - 6 Merawang 23 1.04tn 3.84
7 Ras Pedaging x Kampung 12 1.33tn 3.84
8 Kampung x Ras Pedaging 33 1.25tn 3.84
9 Sentul x Kampung 50 1.02tn 3.84
10 Pelung x Sentul 20 1.05tn 3.84
Keterangan: tn = tidak nyata (P<0.05)
Hasil pada Tabel 3 menunjukkan bahwa semua populasi ayam (kecuali ayam ras petelur) memiliki nilai X² hitung yang tidak berbeda nyata dengan X² tabel (P<0.05) sehingga populasi berada dalam keadaan setimbang. Nilai kesetimbangan Hardy-Weinberg pada ayam ras petelur tidak terdefinisi karena hanya ditemukan 1 alel sehingga dapat dipastikan bahwa populasi ayam ras petelur tidak setimbang karena sudah terjadi fiksasi dalam populasi tersebut. Beberapa faktor yang dapat mempengaruhi kesetimbangan Hardy-Weinberg yaitu seleksi, migrasi, mutasi dan genetic drift (Noor 2010). Pada penelitian ini terjadi mutasi di tingkat gen namun tidak menyebabkan perubahan pada kesetimbangan populasi, karena individu yang mengalami mutasi sangat sedikit sehingga tidak mempengaruhi kesetimbangan populasi secara keseluruhan. Berdasarkan nilai frekuensi alel pada Tabel 1 diketahui bahwa frekuensi alel B (mutasi) pada semua jenis ayam berada pada kisaran 0.00-0.10 atau kurang dari 10% sehingga tidak merubah kesetimbangan populasi.
SIMPULAN DAN SARAN
Simpulan
Genotip yang ditemukan pada gen cGH|MspI dari populasi ayam pada penelitian ini ada 2 yaitu AA dan AB kecuali pada ayam ras petelur hanya ditemuka n 1 genotip yaitu AA dan pada ayam ras pedaging x kampung ditemukan 3 genotip yaitu AA, AB dan BB. Gen cGH pada ayam kampung, pelung, sentul, ras pedaging, merawang, ras pedaging x kampung, kampung x ras pedaging, sentul x kampung dan pelung x sentul bersifat polimorfik, sedangkan ayam ras petelur bersifat monomorfik. Semua populasi ayam lokal Indonesia dan persilangannya pada penelitian ini memiliki keragaman genetik yang rendah namun tetap berada dalam keadaan setimbang.
11
Saran
Sekuensing pada setiap tipe genotip perlu dilakukan untuk mengetahui sekuen DNA yang sebenarnya pada sampel DNA ayam yang diteliti. Perlu dilakukan analisa lebih lanjut untuk menemukan marka genetik lain yang memiliki keragaman yang lebih tinggi dan dapat diasosiasikan dengan sifat pertumbuhan pada ayam.
DAFTAR PUSTAKA
Allendorf FW, Luikart G. 2006. Conservation and The Genetics of Populations. Oxford (UK): Blackwell Publishing.
Bingxue Y, Xuemei D, Jing F, Xiaoxiang H, Changxin W, Ning L. 2003. Single nucleotide polymorphism analysis in chicken growth hormone gene and its associations with growth and carcass traits. Chin Sc Bull. 48(15): 1561-1564.
Green EK. 1998. Restriction Fragment Length Polymorphism. Molekular Biology Methods Handbook. New Jersey (US): Human Pr.
Hardjosubroto W. 2001. Genetika Hewan. Yoygyakarta (ID): Universitas Gadjah Mada Pr.
Kuhnlein U, Ni L, Zadworny D, Weigend S, Gavora JS, Fairfull W. 1997. DNA polymorphisms in the chicken growth hormone gene: response to selection for disease resistanse and association with egg production. An Gene. 28(4): 116-123.
Makhsous SG, Mirhoseini SZ, Zamiri MJ, Niazi A. 2013. Polymorphisms of growth hormone gene in a native chicken population : association with egg production. Bull Vet Inst Pulawy. 57: 73-77.
Muladno. 2010. Teknlogi Rekayasa Genetika. Edisi ke-2. Bogor (ID): IPB Pr. Nei M, Kumar S. 2000. Molecular Evolution and Phylogenetics. New York
(US): Oxford University Pr.
Nei M. 1987. Molecular Evolutionary Genetics. New York (US): Colombia University Pr.
Nie Q, Ip SCY, Zhang X, Leung FC, Yang G. 2002. New variation in intron 4 of growth hormone gene in chinese native chickens. J Heredity. 93(4): 277-279.
Noor RR. 2000. Genetika Ternak. Jakarta (ID): Penebar Swadaya.
Sambrook J, Fritsch EF, Medrano JF. 1989. Molecular Cloning: a Laboratory Manual. 2nd ed. New York (US): Cold Spring Harbor Laboratory Pr. Sulandari S, Zein MSA. 2003. Panduan Praktis Laboratorium DNA. Cibinong
(ID): Bidang Zoologi, Pusat Penelitian Biologi-LIPI Pr.
Tanaka M, Hosokaw Y, Watahiki M, Nakashima K. 1992. Structure of the chicken growth hormone-encoding gene and its promoter region. J Gene. 112(2): 235-239.
Weir BS. 1996. Genetic Data Analysis II: Method for Discrete Population Genetic Data. 2nd ed. Sunderland (US): Sinauer Associates Pr.
12
LAMPIRAN
Sekuen Lengkap Gen cGH (Nomor Akses Gen Bank: D10484.1)
LOCUS CHKGHG 3901 bp DNA linear VRT 21-DEC-2007
DEFINITION Gallus gallus gene for growth hormone, complete cds. ACCESSION D10484 D90458
VERSION D10484.1 GI:222822 KEYWORDS .
SOURCE Gallus gallus (chicken)
ORGANIS
Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi; Testudines + Archosauria group; Archosauria; Dinosauria;
Saurischia; Theropoda; Coelurosauria; Aves; Neognathae; Galliformes; Phasianidae; Phasianinae; Gallus.
REFERENCE 1
AUTHORS Tanaka,M., Hosokawa,Y., Watahiki,M. and Nakashima,K.
TITLE Structure of the chicken growth hormone-encoding gene and its promoter region
JOURNAL Gene 112 (2), 235-239 (1992)
PUBMED
REFERENCE 2 (bases 1 to 3901)
AUTHORS Tanaka,M., Hosokawa,Y., Watahiki,M. and Nakashima,K. TITLE Direct Submission
JOURNAL Submitted (02-AUG-1991) Contact:Minoru Tanaka Nippon Veterinary and
Life Science University, Laboratory of Animal Physiology, Faculty of Applied Life Science; 1-7-1, Kyonan-cho, Musashino-shi, Tokyo 180-8602, Japan FEATURES Location/Qualifiers source 1..3901 /organism="Gallus gallus" /mol_type="genomic DNA" /strain="White leghorn" /db_xref="ta /dev_stage="adult"
/note="5 bp upstream of PstI site.; Common name: Chicken"
/number=1 join(545..554,1270..1430,1873..1989,2291..2452,3519..3719) /codon_start=1 /product="growth hormone" /protein_id= /db_xref="GI:222823" /translation="MAPGSWFSPLLIAVVTLGLPQEAAATFPAMPLSNLFANAVLRAQ HLHLLAAETYKEFERTYIPEDQRYTNKNSQAAFCYSETIPAPTGKDDAQQKSDMELLR FSLVLIQSWLTPVQYLSKVFTNNLVFGTSDRVFEKLKDLEEGIQALMRELEDRSPRGP QLLRPTYDKFDIHLRNEDALLKNYGLLSCFKKDLHKVETYLKVMKCRRFGESNCTI" /number=1 /number=2 /number=2 /number=3
13 /number=3 /number=4 /number=4 /number=5 ORIGIN
1 ctgcagtgga tccaagcaag cactgcctgt gaagctcgtg acaacatcca tcctgctcta 61 acccactgaa tgggaaacca tcccacctgc tctaagccag aagtcccctg cactgccctg 121 gcagccctgt taaccgtggg gcagaaaaac caggcaggaa aatcaggtgg attttctacc 181 tgcgtgagaa attcccccac gtaagcacag aacagatttg ggatgggtct ttcaatggtg 241 ataaaacctc tggttgcaat aaacagcaga atatgaagaa aaagttcagc actaatttta 301 tccccaggca aacatcctcc ccaacctttc catctccgta taaatgacta caatgaggta 361 gcaccatggc gaacacatct gcatttatgc aaggagggga tatggagagg tggcagtgat 421 cacgagcacc cccatccatt ttaaacagac cccccagcta tataaggggt gtctcacctg 481 ttatcatcac ctggatgaaa ggaggaaacg ttcaagcaac acctgagcaa ctctcccggc 541 aggaatggct ccaggtactt tgctttatct cagttctaat gggtgttcca atgctgctgc 601 atgctttggg tgatgggata cgatggtggg gtgtgctgtg gtgggctgac acacgcagag 661 ccggctctga actaaaatgt ggcaacttac agatcagtga caaaggatct ccttccctac 721 agtgcaactt caaaccatga gctgactcag gtaaccctga gcctaacctt gacagggggc 781 aggaatgagc tgcaggctcc agggcattcc tccactgaag ttaaacccta ctgagattaa 841 cttttgtaag cggacactca tgtgagctgg atgtcgaggg ttaataacct tcaggcttga 901 cagtgacctc cagatcctac aggtgtgtcc cagagagcca cagcgcaggt aatgcagcca 961 cttctcaccc cagtgaaggc agacagtgcc atggcagcag cacggtgcaa ataggctcag 1021 ctgagctgtt cccagtcctc acccactgct ctccaccctg ttggctcacg cgccaaagag 1081 tgtaccgtgc tctgctcagg aaggtgaaac ctaccaaaaa acatgagcac gttaggggaa 1141 aataaaggga cggacccaca tggagcaaca tgtgggcttg tgtggagggc tgcactcaca 1201 ggtggacaca accctgagcc ccaggtgcca ccaaaggacg ggtaacccct cccctctcct 1261 ctgctgtagg ctcgtggttt tctcctctcc tcatcgctgt ggtcacgctg ggactgccgc 1321 aggaagctgc tgccaccttc cctgccatgc ccctctccaa cctgtttgcc aacgctgtgc 1381 tgagggctca gcacctccac ctcctggctg ccgagacata taaagagttc gtaagtgttg 1441 gccatctcct cattagcttg atgcctccag gaccgtgcca cggcttccca ccctgcataa 1501 attcctgcaa gcagagactt ttcagctatc ggtgcctcct gaggaggaga aagggtctta 1561 ataggggagg agaggtgatg accgaccagc ctgtcctccc cacagctctc tgcctcctcc 1621 tgcaaggagt cacctcctcc tgtccatgtg ttctgtgctc acctcaaccc ttcagtgaga 1681 gcaatctcta agaccagtag gtgttgtgcc aacacgtggg ctctgctttc ccacctgcca 1741 gtcctgcaca gggatgcaca tcatgtccca cgtttccttc ttgcaaagag caacctgccg 1801 ggaaagagtg aggaaagagt ccgtgctctt ctcttatcac acgacctgag tgcatttggg 1861 atgtctccac aggaacgcac ctatattccg gaggaccaga ggtacaccaa caaaaactcc 1921 caggctgcgt tttgttactc agaaaccatc ccagctccca cggggaagga tgacgcccag 1981 cagaagtcag taagttgtct cccctgggta aacacagcac tgttttatgg aacagagggt 2041 ctccacgtgg tatcagtccc gagaaggaga aatgccttct tacttttcac accctgcatg 2101 cagaaagaca cgggttgggc agtaaatcat attcccaccc taaataaagt cctaaaaaaa 2161 caggctcgag tctgagtggt ggtgctcagc ttacagagct gcctctgggc tgcttcaggg 2221 agagcagggc atgcagcagc actgcagaac acctcacctg cacagctctg aaatcccttt 2281 gtcatttcag gacatggagc tgcttcggtt ttcactggtt ctcatccagt cctggctcac 2341 ccccgtgcaa tacctaagca aggtgttcac gaacaacttg gtttttggca cctcagacag 2401 agtgtttgag aaactaaagg acctggaaga agggatccaa gccctgatga gggtaggtct 2461 gcatactgat ggaagcctgc gctctgctat ttctcttacc tgacatttgg attaacacag 2521 cacccagacc acgagcagct ggagagtgtt cttcagaaag gtcactctta atgcattggc 2581 acagctgccc agggaggtgg gggggtcacc gtccctggag gcgtcccagg gacgtggaga 2641 tgtggcactg agggacgtgg ttatgggcac ggcgggagtg ggttggggtt gggctttggg 2701 atcttggagg tctttccaac cttaataact ctatgatgaa gttttgcttt gttattttga 2761 atgacgttgc ttattgtcag aatcctttct tggatgtgtt cagcatcacc acagctagag 2821 acccacatct atgccacatg agtctggaca gttgtttatg tcttatagac cctaaataac 2881 tagaaaagcc aggcctggga gcaaacaaac cctccgtcct gacattcgat cagaaaagtt 2941 gtggtgcccc atccctggag gtcccaaggc catggatgga gccctgggca gtttgagccg 3001 gtggggggca cccagcccat ggcagggggt gagacggggg ggctctgaga tcccttccaa 3061 cacaaccgtg ctgtgattcc acagccctgc aatccttgag gtcccttcca acccaacagt 3121 gccacgattc catggttctg tgatccttaa ggtcccttcc aacccaacca tgccatcatt 3181 ccacggttcc atgatctcag aggccccttc caacccaacc acgccgtgat tccatgacac 3241 ttcagctgca tctcaatgtg cagcaccact tccagatctt actgtccact gatacgagca 3301 caggtgctga caaccgatac gcagaccatc tccatccctt ccactgctgc ttacttaccc 3361 caaacacttc cgagccacgc ggggtgagca gtgggtgcag ctcacagctc cacgacggcc 3421 agggctgggt cagagctctg tccactagaa aactggagca aagacagcat cactttgcca 3481 gcagcccctc gctcagccgc agccctctcg tcccacagga gctggaggac cgcagcccgc 3541 ggggcccgca gctcctcaga cccacctacg acaagttcga catccacctg cgcaacgagg 3601 acgccctgct gaagaactac ggcctgctgt cctgcttcaa gaaggatctg cacaaggtgg
14
3661 agacctacct gaaggtgatg aagtgccggc gcttcggaga gagcaactgc accatctgag 3721 gccctgtgcc tgcgccatgg ctgacggccc tgtccccccc cccccccttc ctccccgtca 3781 ccaaaaacac gaggaataaa ccccacagcg ctgagctctg cctgctgtct gctggctggg 3841 gatatagggc gggttcgggg cgggctcagg gccgggcaaa ggggagggag gaggggggcc 3901 c
15
RIWAYAT HIDUP
Penulis bernama lengkap Ria Putri Rahmadani dilahirkan di Bukittinggi pada tanggal 15 Maret 1993 dari pasangan ayah Syaiful Firmadi dan ibu Sri Maharani (almarhumah). Penulis merupakan anak pertama dari 3 bersaudara dengan adik M. Irfan Putra Pratama dan M. Fisqal Aditia. Penulis telah menyelesaikan pendidikan Taman Kanak-Kanak (TK) di TK Raudhatul Atfal Soreang tahun 1998, Sekolah Dasar (SD) di SDN 28 Limo Suku tahun 2004, Sekolah Menengah Pertama (SMP) di SMPN 6 Bukittinggi tahun 2007, Sekolah Menengah Atas (SMA) di SMAN 3 Bandar Lampung tahun 2010 dan langsung diterima di Institut Pertanian Bogor (IPB) melalui jalur Undangan Seleksi Mahasiswa IPB (USMI) pada program studi Ilmu Produksi dan Teknologi Peternakan (IPTP).
Selama menjadi mahasiswa IPB, penulis memperoleh beasiswa Bidik Misi dari Direktorat Pendidikan Tinggi (DIKTI) selama 8 semester. Kegiatan penulis di luar akademik yaitu pernah menjadi staf departemen pengembangan sumber daya mahasiswa (PSDM) di BEM TPB tahun 2010/2011, sekretaris umum paguyuban bidik misi tahun 2010/2011, sekretaris umum DMP Fakultas Peternakan tahun 2011/2012 dan 2012/2013 serta serangkaian kepanitian kegiatan di Fakultas Peternakan IPB. Penulis juga menjadi asisten praktikum pada mata kuliah Genetika Ternak tahun 2014.
Prestasi akademik yang diraih oleh penulis selama kuliah di IPB yaitu mendapatkan IPK 3.64 selama 7 semester, juara II lomba esai peternakan di Fakultas Peternakan tahun 2011, PKM penelitian didanai DIKTI tahun 2013 dengan tema pangan fungsional dari hasil ternak, juara I aerobik Dekan Cup tahun 2012, 2013 dan 2014, publikasi karya tulis pada prosiding internasional di Taiwan 2013 dengan tema peternakan integrasi, seminar dan publikasi karya tulis pada prosiding internasional TRIU 2013 di Jepang dengan tema pangan fungsional dari hasil ternak serta PKM kewirausahaan didanai DIKTI tahun 2014 dengan tema diversifikasi pangan dari hasil ternak.