Halaman: 43-48
2000 Jurusan Biologi FMIPA UNS Surakarta
Polimorfisme DNA Gen Hormon Pertumbuhan dan Sifat Produksi
pada Sapi Komposit
SUTARNO
Jurusan Biologi FMIPA UNS Surakarta
ABSTRAK
The objective of the study was to know the association between genetic variation at growth hormone loci and production traits of Composite breed of beef cattle. The composite breed comprising approximately 1/4 Brahman, Charolais and Friesian, and 1/8 Angus and Hereford. PCR-RFLPs were applied for the detection of DNA polymorphism at locus I and II of growth hormone gene, and the result indicated that polymorphisms were found at both loci using AluI and MspI enzymes. Sequence analysis indicated that the polymorphism at AluI restriction site was due to substitution of Leucine/Valine at position 127, while MspI polymorphic site was caused by transition of C to T at position +837. Association between genotype variations and quantitative production traits (birth weight, daily gain, milk production and calving rate) was analysed using two mode Anova implemented in a program of JMP and PEST. The analysis indicated that the MspI genotypes at locus II of the gene was significantly correlated birth weight, but not for locus I detected using AluI.
Key words: DNA polymorphism, growth hormone, production traits, beef cattle
PENDAHULUAN
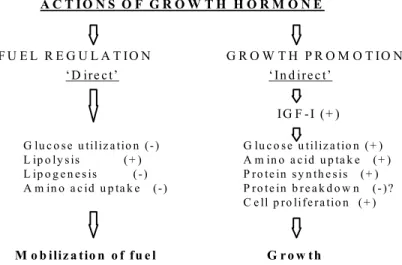
Hormon pertumbuhan merupakan hormon alami yang disekresikan oleh glandula pituitoria anterior semua mammalia. Struktur hormon ini berbeda-beda antara spesies satu dengan yang lain dan pada umumnya memiliki pengaruh spesifik yaitu penga-turan pertumbuhan. Sintesis dan sekresi hormon ini dikendalikan somatostatin yang menghambat sekresi dan growth hormone-releasing factor yang menstimuli sintesis maupun sekresi (Kopchick & Cioffi, 1991). Mekanisme kerja secara langsung hormon ini terjadi melalui mobilisasi nutrien, termasuk penghambatan penggunaan glukosa, lipogenesis dan masukan asam amino, sedangkan kerja secara tidak langsung melalui perantaraan insulin like growth factors (IGFI). Mekanisme secara tidak langsung merupakan akibat dari peningkatan glukosa dan masukan asam amino, sintesis protein, proliferasi sel dan kemungkinan besar penghambatan pemecahan protein (Ballard et al., 1993) (Gambar 1).
Gen Hormon Pertumbuhan pada Sapi
Gen hormon pertumbuhan pada sapi telah diklon dan disekuen secara keseluruhan (Gordon et al., 1983; Woychik et al., 1982). Panjang gen hormon pertumbuhan ini 1793bp, terdiri dari 5 ekson (daerah pengkode) yang dipisahkan oleh 4
intron (intervening sequence) yang terdiri dari 248 bp (intron A), 227 bp (intron B), 227 bp (intron C) dan 274 bp (intron D) (Gordon et al., 1983). Lokus gen hormon pertumbuhan pada sapi telah ditemukan terletak pada kromosom 19 dengan lokasi 19q26-qtr (Hediger et al., 1990).
Pada sapi perah, Lucy et al. (1993) melaporkan adanya variasi situs restriksi AluI pada ekson V karena substitusi asam amino pada posisi 127. Variasi ini mempengaruhi produksi susu pada sapi perah jenis Holstein (Lucy et al., 1993). Pada percobaannya, sapi-sapi dengan allel homozigot untuk leusin 127 (L) memilki superioritas dalam produksi susu, meskipun hasil penemuan ini tidak cocok dengan penelitian menggunakan rekombinan somatotropin sapi, dimana allel rekombinan valin 127 (V) lebih efektif meningkatkan produksi susu ketika diinjeksikan pada sapi perah (Eppard et al., 1992). Penelitian lebih jauh oleh Schlee et al. (1994a) menunjukkan bahwa genotipe AluI tidak mempengaruhi produksi susu, tetapi secara signifikan mempengaruhi sifat produksi daging, dimana genotip heterozigot LV bersifat superior baik pada pencapaian karkas maupun kualitas daging pada sapi jenis Simmental.
Polimorfisme pada gen hormon pertumbuhan yang teridentifikasi dengan enzim restriksi TaqI berhubungan dengan pertumbuhan sapi-sapi Korea (Choi et al., 1997), dan sapi jenis Brahman
G lu c o s e u tiliz a tio n (+ ) A m in o a c id u p ta k e (+ ) P ro te in s y n th e s is (+ ) P ro te in b r e a k d o w n (- )? C e ll p ro life r a tio n (+ ) A C T I O N S O F G R O W T H H O R M O N E F U E L R E G U L A T IO N
‘D ire c t’ ‘In d ire c t’
G R O W T H P R O M O T IO N IG F -I (+ ) G lu c o s e u tiliz a tio n (- ) L ip o ly s is (+ ) L ip o g e n e s is ( -) A m in o a c id u p ta k e (-) M o b iliz a tio n o f fu e l G r o w th
Gambar 1. Skema menunjukkan pengaruh hormon pertumbuhan pada mobilisasi bahan untuk energi dan pertumbuhan (Ballard, 1993).
(Rocha et al., 1991). Polimorfisme pada intron ketiga yang terdeteksi dengan RFLP menggunakan MspI akibat adanya insersi atau transisi pada posisi 837/838 gen hormon pertumbuhan dilaporkan berhubungan dengan lemak susu, dimana frekwensi D-MspI (-) secara signifikan lebih tinggi pada individu-individu jenis Red Dannish yang terseleksi untuk lemak susu yang tinggi daripada yang lemak susunya rendah (Hoj et al., 1993a).
BAHAN DAN METODE
Sapi Percobaan
Penelitian ini menggunakan sapi Komposit sebanyak 176 (terseleksi) dan 51 (kontrol) di pusat penelitian sapi Wokalup, Australia Barat. Sapi Komposit tersebut merupakan hasil perkawinan silang yang komposisinya terdiri dari 1/4 Brahman, Charolais dan Friesian, serta 1/8 Angus dan Hereford. Pengambilan Sampel Darah
Sampel darah diambil secara venepuncture, menggunakan venoject. Darah yang diperoleh dimasukkan dalam 50 ml tabung reaksi yang berisi 2,5 ml EDTA 200 mM sebagai antikoagulan. Sebanyak 10 ml dari darah ini diambil dan disimpan pada suhu –70oC untuk referensi di kemudian hari, sedangkan sisanya digunakan langsung dalam penelitian ini untuk diekstrak sel darah putihnya.
Pemisahan Sel Darah Putih dari Sampel Darah Sel darah putih diekstrak dengan teknik Buffy coat. Total darah dimasukkan ke dalam tabung sentrifus dan disentrifugasi pada kecepatan 1500g selama 15-20 menit. Buffy coat yang diperoleh
diambil dengan pipet, dipindahkan ke dalam 20 ml tabung sentrifus, dipenuhi dengan larutan buffer TE1 dan disentrifugasi lagi pada kecepatan 2000g selama 10-15 menit. Pelet yang diperoleh diresus-pensikan dalam 1 ml bufer TE2 dan dipindahkan ke dalam tabung penyimpan (Nunc) dan disimpan pada suhu –80oC sampai saat dibutuhkan.
Ekstraksi DNA
DNA diekstraksi dari sel darah putih dengan menggunakan Wizard Genomic Purification System (Promega, Madison USA) sesuai dengan petunjuk dari perusahaan.
PCR-RFLP
DNA yang diperoleh langsung digunakan untuk reaksi PCR dengan mesin PCR (thermocycler Omnigene). Reaksi ini bertujuan mengamplifikasi fragmen DNA yang diinginkan, yaitu lokus 1 dan 2 dari gen hormon pertumbuhan. Reaksi dilakukan dalam suatu volume campuran sebanyak 50µL yang berisi 200µM dari masing-masing dNTPs, 2 mM MgCl2, 10x bufer dan 1,5 unit Taq DNA polymerase dalam 0,6 ml tabung PCR.
Dua fragmen gen hormon pertumbuhan diamplifikasi dengan PCR. Lokus 1 gen hormon pertumbuhan (GH-L1) yang terdiri dari 223 bp meliputi ekson IV dan V, serta lokus 2 gen hormon pertumbuhan (GH-L2) yang terdiri dari 329bp meliputi ekson III dan IV, diamplifikasi dengan PCR menggunakan primer berturut-turut GH1/GH2 dan GH3/GH4. Primer GH1/GH2 digunakan untuk mengamplifikasi sebagian fragmen intron D dan ekson V, sedangkan GH3/GH4 untuk mengam-plifikasi fragmen antara intron III dan intron IV.
Primer yang digunakan untuk mengamplifikasi fragmen ini adalah:
GH1: 5’-GCTGCTCCTGAGGGCCCTTCG-3’ GH2: 5’-GCGGCGGCACTTCATGACCCT-3’ GH5: 5’-CCCACGGGCAAGAATGAGGC-3’ GH6: 5’-TGAGGAACTGCAGGGGCCCA-3’
Semua reaksi amplifikasi dilakukan dalam kondisi yang sama. Dyna wax (Finnzyme Oy) selalu ditambahkan untuk menghasilkan suatu “hot start” PCR, sehingga diperoleh hasil yang spesifik. Semua reaksi amplifikasi dilakukan dalam volume 50µL campuran reaksi yang terdiri dari: 200 ng DNA template, 0.15 µM dari masing-masing oligonukleotida primer, 200 µM dari masing-masing dNTPs, 2 mM MgCl2, 10x bufer dan 1,5 unit Taq DNA polymerase dalam 0,6 ml tabung effendorf. Kontrol negatif (tanpa DNA template) selalu disertakan dalam semua reaksi dan semuanya tidak menghasilkan suatu produk.
Kondisi reaksi amplifikasi PCR untuk gen hormon pertumbuhan adalah sebagai berikut: reaksi denaturasi awal pada suhu 94oC selama 5 menit, diikuti dengan 30 siklus amplifikasi yang masing-masing siklus terdiri dari: denaturasi pada 94oC selama 45 detik, annealing pada 60oC selama 45 detik dan extension pada 72oC selama 1 menit; diikuti dengan satu tahap polimerasi final pada 72oC selama 5 menit.
Hasil amplifikasi PCR langsung digunakan dalam reaksi digesti menggunakan enzim AluI untuk mengidentifikasi situs polimorfisme AluI, sedangkan lokus 2 menggunakan enzim MspI.
Aliquot yang terdiri dari 7 µL (hasil amplifikasi gen hormon pertumbuhan) dimasukkan ke dalam tabung effendorf steril. Master mix yang terdiri dari campuran enzim, bufer dan air ditambahkan ke dalam tabung yang berisi sampel DNA dan diinkubasi sesuai dengan petunjuk dari produsen enzim tersebut. Hasil digesti dielektroforesis pada bak elektroforesis horizontal dengan menggunakan gel yang terbuat dari 1-2% agarose dalam bufer TAE. Elektroforesis dilakukan selama 90 menit
pada 55 volt. Hal ini sangat tergantung konsentrasi gel dan voltase. Agar hasil elektroforesis dapat divisualisasi, disertakan ethidium bromida pada saat pembuatan gel dengan konsentrasi final 0,12
µg/ml. Setelah selesai elektroforesis, DNA divisualisasi di bawah sinar ultra violet dalam ruang gelap dan difoto menggunakan film Polaroid ukuran 57 dengan filter merah.
Analisis Data
Hubungan antara variasi genotipe pada lokus I dan II gen hormon pertumbuhan terhadap sifat kuantitatif (berat lahir, pencapaian berat harian, produksi susu dan sifat reproduksi) dianalisis menggunakan dua model ANOVA yaitu model I menggunakan program JMP (SAS, 1989) dan model II menggunakan program PEST (Groeneveld, 1990; Groeneveld & Kovac, 1990). Model I: Yijklmn = + Bi + Sj + Lk + Dl + Ym + Gn + eijklmn
Model II: Yijklmno = + Bi + Sj + Lk + Dl + Ym + Gn + Aijklmno + eijklmno
Dimana µ = nilai mean terkecil, Bi = jenis sapi, Sj = jenis kelamin, Lk = kelompok (terseleksi atau kontrol), Dl = umur setiap individu, Ym = tahun lahir, Gn = genotipe, Aijklmno = pengambilan hewan secara random, ditentukan berdasarkan hubungan matriks, dan eijklmno = tingkat kesalahan.
HASIL
Polimorfisme pada Situs Restriksi
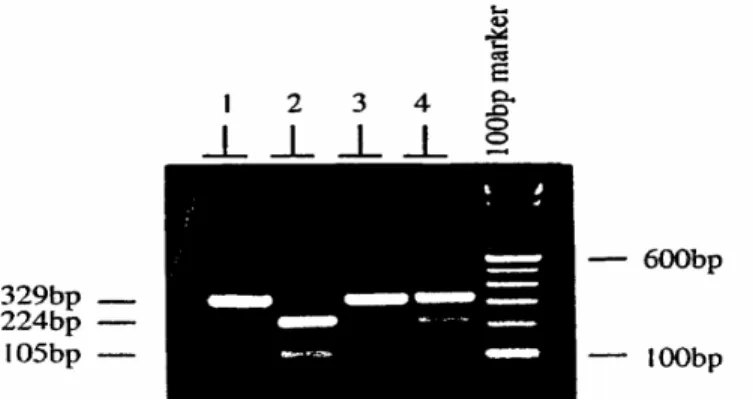
Analisis RFLP terhadap fragmen yang diamplifikasi dengan PCR menunjukkan adanya polimorfisme baik pada lokus I maupun lokus II gen hormon pertumbuhan dengan menggunakan enzim restriksi AluI dan MspI. Contoh fotograf dari gel agarose hasil digesti menggunakan enzim MspI ditunjukkan pada Gambar 2.
Gambar 2. Fotograf dari gel agarose memperlihatkan adanya polimorfisme DNA pada lokus 2 gen hormon pertumbuhan yang dideteksi dengan teknik PCR-RFLP menggunakan enzim MspI. Lane (baris) 1 = Uncut (tanpa pemotongan), 2 = MspI (++), 3 = MspI (--), 4 = MspI (+-).
Tabel 1. Perbandingan diversitas genetik antara kelompok terseleksi dan kontrol pada sapi jenis Komposit. NS = Tidak signifikan.
Jenis Kelompok N Heterozigositas
teramati (observed)
Rerata allel per lokus Heterozigositas diharapkan (expected) t P Komposit Terseleksi 158 .388 + .301 2.000 + .000 .448 + .046 0.289 NS Kontrol 47 .327 + .214 2.000 + .000 .360 + .094
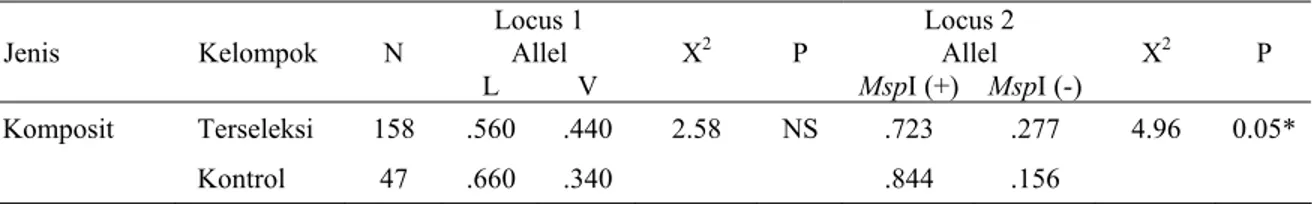
Tabel 2. Perbandingan frekwensi allel antara kelompok terseleksi dan kontrol pada sapi pedaging jenis Komposit. NS = Tidak signifikan, *= signifikan pada 0,05.
Locus 1 Locus 2 Allel Allel Jenis Kelompok N L V X2 P MspI (+) MspI (-) X2 P Komposit Terseleksi 158 .560 .440 2.58 NS .723 .277 4.96 0.05* Kontrol 47 .660 .340 .844 .156 Data fenotip
Sifat fenotip yang digunakan dalam analisis ini adalah berat sewaktu lahir, pertumbuhan harian sejak lahir sampai saat disapih, produksi susu dan rata-rata tingkat melahirkan selama 4 tahun (reproduksi) (data tidak ditunjukkan). Perbandingan diversitas genetik antara kelompok terseleksi dan kontrol pada sapi Komposit ditunjukkan pada Tabel 1, sedangkan Tabel 2 memperlihatkan perbandingan frekwensi allel dari kelompok terseleksi dan kontrol pada sapi Komposit. Perbedaan frekwensi allel kedua kelompok ini adalah signifikan pada lokus II.
Pengaruh genotipe AluI
Tabel 3 menunjukkan nilai masing-masing genotipe (AluI) pada gen hormon pertumbuhan lokus I untuk sifat fenotip berat lahir, pertumbuhan harian, produksi susu (selama 3 tahun) dan reproduksi (calving rate selama 4 tahun). Sedangkan Tabel 4 menunjukkan probabilitas perbedaan mean antar genotipe dari Anova model I dan II. Tidak ada perbedaan yang signifikan antara genotipe satu dengan yang lain.
Tabel 3. Mean kuadrat terkecil + S.E. dari berat lahir (BW), pertumbuhan harian (DG), produksi susu (Milk) dan mean calving rate/reproduksi (CR) sapi Komposit dari genotipe AluI yang berbeda.
Sifat LL LV VV
BW 38.29 + 0.69 37.88 + 0.72 37.21 + 1.04 DG 5.44 + 0.57 3.64 + 0.58 4.18 + 0.89 Milk 4.07 + 0.33 3.64 + 0.32 3.99 + 0.53 CR 0.67 + 0.05 0.70 + 0.05 0.62 + 0.08
Tabel 4. Probabilitas perbedaan mean antar genotipe dari Anova model I dan II. BW = birth weight (berat lahir), DG = daily gain (pencapaian pertumbuhan harian), Milk = milk production (produksi susu), CR = calving rate (reproduksi).
Sifat Nilai P Model I Model II BW 0.51 0.61 DG 0.94 0.94 Milk 0.40 0.32 CR 0.56 0.21 Pengaruh MspI
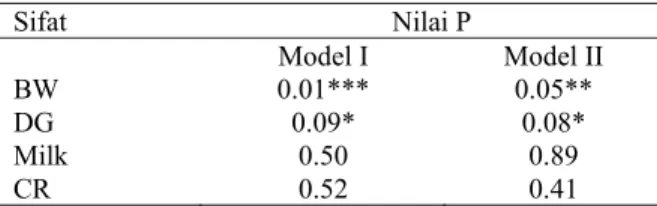
Tabel 5 menunjukkan nilai dari masing-masing genotipe (MspI) pada gen hormon pertumbuhan lokus I untuk sifat fenotip berat lahir, tambahan berat harian (pertumbuhan), produksi susu (selama 3 tahun) dan reproduksi (calving rate selama 4 th). Sedangkan Tabel 6 menunjukkan probabilitas perbedaan mean antar genotipe dari Anova model I dan II. Perbedaan berat lahir adalah signifikan baik pada Anova model I maupun II, sedangkan pertumbuhan harian signifikan pada level 8%. Tabel 5. Mean kuadrat terkecil + S.E. dari berat lahir (BW), pertumbuhan harian (DG), produksi susu (Milk) dan mean calving rate/ reproduksi (CR) sapi Komposit dari genotipe MspI yang berbeda.
Sifat ++ +- --
BW 37.21 + 0.72 38.22 + 0.75 41.27 + 1.41 DG 1.16 + 0.02 1.18 + 0.02 1.24 + 0.04 Milk 3.84 + 0.29 3.89 + 0.44 4.94 + 0.97 CR 0.69 + 0.04 0.68 + 0.07 0.51 + 0.16
Tabel 6. Probabilitas perbedaan mean antar genotipe dari Anova model I dan II. Berat lahir (BW), pertumbuhan harian (DG), produksi susu (Milk) dan mean calving rate/reproduksi (CR), ***, **, * signifikan berturut-turut pada level 0.01, 0.05 dan 0.1.
Sifat Nilai P Model I Model II BW 0.01*** 0.05** DG 0.09* 0.08* Milk 0.50 0.89 CR 0.52 0.41 PEMBAHASAN
Beberapa gen utama (major) yang mempengaruhi sifat-sifat bernilai ekonomi tinggi, khususnya pada hewan yang digunakan dalam produksi daging, telah diidentifikasi misalnya: double muscling gene (gen mh) pada sapi (Hanset & Michaux, 1985), acid meat gene (gen RN) yang mempengaruhi kualitas daging babi (Le Roy et al., 1990) dan gen Boorola yang mempengaruhi ovulasi dan ukuran tubuh domba (Piper et al., 1985). Namun demikian, kebanyakan sifat yang memiliki nilai ekonomi tinggi pada hewan ternak secara alami dikontrol oleh banyak gen yang masing-masing memilki pengaruh, sehingga untuk mendekteksi kualitas semacam ini dan menjadikannya sebagai marker gen menjadi sulit.
Penelitian-penelitian terdahulu telah dilakukan untuk membuktikan hipotesis yang mengatakan bahwa perbedaan genotipe gen hormon pertumbuhan berhubungan dengan parameter-parameter produksi. Pada hewan-hewan percobaan di laboratorium, Winkelmann et al. (1992; 1990) misalnya, menemukan bahwa RFLPs pada gen hormon pertumbuhan berhubungan secara signifikan dengan berat pada 42-hari.
Pada penelitian ini, tampak bahwa genotipe AluI gen hormon pertumbuhan tidak berpengaruh secara signifikan pada beberapa parameter produksi, meskipun percobaan sebelumnya oleh Eppard et al. (1992) menunjukkan bahwa rekombinan valin 127bST lebih efektif dibanding dengan leusin 127bST dalam meningkatkan produksi susu bila diinjeksikan pada sapi perah. Terjadinya substitusi asam amino ini mempengaruhi konsentrasi sirkulasi hormon pertumbuhan dan insulin-like growth factor I (IGF-I) (Schlee et al., 1994b). Di samping itu, penelitian sebelumnya juga menunjukkan adanya pengaruh yang signifikan terhadap produksi susu pada sapi jenis Holstein (Lucy et al., 1993) dan pertumbuhan serta produksi daging (Schlee et al., 1994a).
Pada Tabel 6 diperlihatkan bahwa meskipun polimorfisme MspI gen hormon pertumbuhan tidak menunjukkan efek yang signifikan baik pada produksi susu maupun fertilitas, polimorfisme MspI ini mempunyai pengaruh yang signifikan pada berat lahir dan rata-rata pertumbuhan harian. Analisis lebih jauh menunjukkan bahwa hanya berat lahir yang secara signifikan (0,02) dipengaruhi oleh polimorfisme MspI (Tabel 7). Hal ini ditunjukkan pula oleh adanya kecenderungan (signifikan) meningkatnya frekwensi allel MspI (-) yang mungkin disebabkan terjadinya proses seleksi untuk meningkatkan pertumbuhan dari jenis ini (Tabel 2).
Tabel 7. Probabilitas perbedaan mean berat lahir (BW) dan rata-rata pertumbuhan (DG) dari analisis dengan menggunakan Anova model I.
Jenis Kelompok Probabilitas
BW DG
Komposit Terseleksi 0.02** 0.55
Kontrol 0.20 0.61
Analisis data di atas menunjukkan bahwa polimorfisme DNA pada gen hormon pertumbuhan mempengaruhi secara signifikan berat lahir pada sapi jenis Komposit, sedangkan polimorfisme DNA yang dideteksi dengan AluI tidak menunjukkan adanya pengaruh yang signifikan pada sifat-sifat produksi, meskipun laporan terdahulu menunjukkan adanya pengaruh polimorfisme AluI terhadap sifat produksi susu. Eppard et al. (1992) menunjukkan bahwa rekombinan valin 127bST lebih efektif dari pada leusin 127bST dalam meningkatkan produksi susu bila diinjeksikan pada sapi perah.
Kazmer et al. (1986) dan Klemetsdal et al. (1991) menunjukkan adanya pelipatgandaan konsentrasi hormon pertumbuhan pada sapi yang terseleksi untuk penghasil susu yang tinggi dibandingkan dengan yang rendah maupun kontrol. Peningkatan ini mungkin terjadi secara langsung melalui peningkatan ketersediaan nutrien untuk kelenjar susu atau secara tidak langsung dengan meningkatkan aktifitas kelenjar susu melalui peningkatan kadar IGF-I (Bauman & Vernon, 1993; Burton et al., 1994). Pemberian hormon pertumbuhan juga diketahui meningkatkan rata-rata pertumbuhan sapi (Groenewegen et al., 1990), meningkatkan efisiensi pakan serta meningkatkan komposisi karkas, yaitu mengurangi lemak karkas (Peters, 1986) dan protein karkas (Peters, 1986). Efek yang sama diperlihatkan juga pada domba (Pell et al., 1990; Zainur et al., 1989). Pemberian hormon pertumbuhan diperkirakan mempengaruhi reseptor hormon pertumbuhan. Meningkatnya jumlah dan afinitas reseptor hormon pertumbuhan
telah ditunjukkan pada babi (Chung & Etherton, 1986) dengan pGH yang dilabel dengan 125I.
Pengaruh genotipe gen hormon pertumbuhan terhadap sifat produksi mungkin berbeda antara penelitian satu dengan yang lain, karena kompleks-nya proses fisiologi yang melibatkan berbagai mekanisme aksi, dan pengaruh dari setiap gen tunggal dimediasi oleh sifat genetik pada aksis hormon pertumbuhan.
DAFTAR PUSTAKA
Ballard, F. J., Francis, G. L., Walton, P. E., Knowles, S. E., Owens, P. C., et al. 1993. Modification of animal growth with growth hormone and insulin-like growth factors.
Australian Journal of Agricultural Research 44:567-577 Bauman, D. E., Vernon, R. G. 1993. Effects of exogenous
bovine somatotropin on lactation. Annual Review of Nutrition 13:437-461
Burton, J. L., McBride, B. W., Block, E., Glimm, D. R., Kennelly, J. J. 1994. A review of bovine growth hormone.
Canadian Journal of Animal Science 74:167-201
Choi, Y. J., Yim, D. S., Cho, J. S., Cho, B. D., Na, K. J., Balk, M. G. 1997. Analysis of Restriction Fragment Length Polymorphism in the Bovine Growth Hormone Gene Related to Growth Performance and Carcass Quality of Korean Native Cattle. Meat Science 45:405-410
Chung, S. C., Etherton, T. D. 1986. Characterization of porcine growth hormone (pGH) binding to porcine liver microsomes: chronic administration of pGH induces pGH binding. Endocrinology 119:780-786
Eppard, P. J., Bentle, L. A., Violand, B. N., Ganguli, S., Hintz, R. L., et al. 1992. Comparison of the galactopoietic response to pituitary-derived and recombinant-derived variants of bovine growth hormone. Journal of
Endocrinology 132:47-56
Gordon, D. F., Quick, D. P., Erwin, C. R., Donelson, J. E., Maure, R. A. 1983. Nucleotide sequence of the bovine growth hormone chromosomal gene. Molecular and
Cellular Endocrinology 33:81-95
Groeneveld, E. 1990. PEST User Manuals Newstad - Germany: Institute of Animal Husbandary & Animal Behaviour, Federal Agricultural Research Centre.
Groeneveld, E., Kovac, M. 1990. A generalized computing procedure for setting up and solving mixed linear models.
Journal of Dairy Science 73:513-531
Groenewegen, P. P., McBride, B. W., Burton, J. H., Elsasser, T. H. 1990. Bioactivity of milk from bST-treated cows.
Journal of Nutrition 120:514
Hanset, R., Michaux, C. 1985. On the genetic determinism of muscular hypertrophy in the Belgian White and Blue cattle breed. I. Experimental data. Genetics, Selection and Evolution 17:359-368
Hediger, R., Johnson, S. E., Barendse, W., Drinkwater, R. D., Moore, S. S., Hetzel, J. 1990. Assignment of the growth hormone gene locus to 19q26-qter in cattle and to 11q25-qter in sheep by in situ hybridization. Genomics 8:171-174. Hoj, S., Fredholm, M., Larsen, N. J., Nielsen, V. H. 1993a.
Growth hormone gene polymorphism associated with selection for milk fat production in lines of cattle. Animal Genetics 24:91-96
Kazmer, G. W., Barnes, M. A., Akers, R. M., Pearson, R. E. 1986. Effect of genetic selection for milk yield and increased milking frequency on plasma growth hormone and prolactin concentration in Holstein cows. Journal of
Animal Science 63:1220-1227
Klemetsdal, G., Tveit, B., Vingelen, M., Starova, J., Sejrsen, K. 1991. The plasma level of growth hormone in two genetic lines of dairy cattle selected for high and low milk yield. In
42 nd Annual Meeting EAAP. pp. G5b7. Berlin:
Kopchick, J. J., Cioffi, J. A. 1991. Exogenous and endogenous effects of growth hormone in animals. Livestock Production Science 27:61-75
Le Roy, P., Naveau, J., Elsen, J. M., Sellier, P. 1990. Evidence for a new major gene influencing meat quality in pigs.
Genetical Research 55:33-40
Lucy, M. C., Hauser, S. D., Eppard, P. J., Krivi, G. G., Clark, J. H., et al. 1993. Variants of Somatotropin in Cattle - Gene Frequencies in Major Dairy Breeds and Associated Milk Production. Domestic Animal Endocrinology 10:325-333 Pell, J. M., Elcock, C., Harding, R. L., Morrell, D. J.,
Simmonds, A. D., Wallis, M. 1990. Growth, Body composition, hormonal and metabolic status in lambs treated long-term with growth hormone. British Journal of Nutrition 63:431-445
Peters, J. P. 1986. Consequences of accelerated gain and growth hormone administration for lipid metabolism in growing beef steers. Journal of Nutrition 116:2490-2503 Piper, L. R., Bindon, B. M., Davis, G. H. 1985. The single
gene inheritance of the high litter size of the Booroola merino. In Genetics of Reproduction in Sheep, ed. R. B. Land, D. W. Robson. pp. 115-125. London: Butterworths Rocha, J. L., Baker, J. F., Womack, J. E., Sanders, J. O.,
Taylor, J. F. 1991. Associations between RFLPs and quantitative sifat in beef cattle. Journal of Animal Science
69 (suppl 1):201
SAS. 1989. JMP User's Guide, Version 2 of JMP Cary, NC, USA: SAS Institute Inc.
Schlee, P., Graml, R., Rottmann, O., Pirchner, F. 1994a. Influence of Growth-Hormone Genotypes On Breeding Values of Simmental Bulls. Journal of Animal Breeding & Genetics Zeitschrift fur Tierzuchtung und
Zuchtungsbiologie 111:253-256
Schlee, P., Graml, R., Schallenberger, E., Schams, D., Rottmann, O., et al. 1994b. Growth Hormone and Insulin Like Growth Factor I Concentrations in Bulls of Various Growth Hormone Genotypes. Theoretical & Applied Genetics 88:497-500
Winkelmann, D. C., Hodgetts, R. B. 1992. RFLPs for somatotropic genes identify quantitative trait loci for growth in mice. Genetics 131:929-937
Winkelmann, D. C., Querengesser, L. D., Hodgetts, R. B. 1990. Growth hormone restriction fragment length polymorphisms that segregate with 42-day live weight of mice. Genome 33:235-239
Woychik, R. P., Camper, S. A., Lyons, R. H., Horowitz, S., Goodwin, E. C., Rottman, F. M. 1982. Cloning and nucleotide sequencing of the bovine growth hormone gene.
Nucleic Acid Research 10:7197-7210.
Zainur, A. S., Tassel, R., Kellaway, R. C., Dodemaide, W. R. 1989. Recombinant growth hormone in growing lambs: effects on growth, feed utilization, body and carcass characteristics and on wool growth. Australian Journal of Agricultural Research 40:195-206