www.elsevier.comrlocaterapplanim
Postingestive feedback from starch influences the
ingestive behaviour of sheep consuming wheat
straw
Juan J. Villalba
), Frederick D. Provenza
Department of Rangeland Resources, Utah State UniÕersity, Logan, UT 84322-5230, USA
Accepted 18 August 1999
Abstract
Plant species present a complex array of biochemicals to herbivores that in conjunction with a plant’s physical structure influence intake. Our objective was to determine the role of
postinges-Ž .
tive feedback from macronutrients on the ingestion of a low-quality forage by sheep OÕis aries .
We compared the ingestive behaviour of two groups of lambs conditioned with intraruminal
Ž . Ž .
infusions of starch 100 grlambrday; Group 1 or water 250 mlrlambrday; Group 2 while they
Ž .
grazed wheat straw held erect in wooden frames; 30 cm height; Trials 1, 2 and 3 or consumed
Ž .
wheat straw from food boxes 1–2 cm particle size; Trial 4 . Lambs were conditioned as
Ž .
individuals during Trials 1 and 4, in pairs during Trial 2, and as two groups Groups 1 and 2 during Trial 3. Ingestive behaviour – intake, intake rate, number of bites, bite size, bite rate — was assessed during individual tests that lasted 5 minrlamb. After conditioning and testing, all
Ž .
animals had restricted access 80% of their daily energy requirements to a basal diet of alfalfa
Ž .
pellets. No differences between groups were detected during initial tests before infusing starch or
Ž . Ž
during Trial 1 P)0.05 . Lambs conditioned with starch in Trial 2 consumed more straw 8 vs. 2
. Ž . Ž .
g; P-0.05 at higher rates 2.3 vs. 1.0 grmin; P-0.1 , and took more 13 vs. 5; P-0.1 and
Ž .
larger 0.59 vs. 0.31 grbite; P-0.05 bites than lambs conditioned with water. This pattern was
Ž .
maintained during Trial 3: Lambs infused with starch showed higher intake 24 vs. 5 g; P-0.01 ,
Ž . Ž . Ž
intake rate 5.4 vs. 1.4 grmin; P-0.001 , bites 29 vs. 10; P-0.01 , bite rate 6.7 vs. 2.6
. Ž .
bitesrmin; P-0.01 and bite size 0.86 vs. 0.54 grbite; P-0.05 than lambs infused with
Ž .
water. In Trial 4, lambs conditioned with starch consumed more straw 21 vs. 9 g; P-0.1 , at
Ž .
higher rates 5.4 vs. 2.6 grmin; P-0.1 , than lambs conditioned with water. Thus, the
)Corresponding author. Tel.:q1-435-797-2539; fax:q1-435-797-3796; e-mail: [email protected] 0168-1591r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved.
Ž .
postingestive effects of energy played an important role in modulating rates of food intake, and our findings suggest postingestive feedback from macronutrients is a fundamental factor influenc-ing the influenc-ingestive behaviour of herbivores.q2000 Elsevier Science B.V. All rights reserved.
Keywords: Sheep; Intake rate; Bite rate; Starch; Feeding behavior; Foraging behaviour
1. Introduction
The process of gathering food is an essential affair for living organisms. For herbivores, behaviours directed toward finding and harvesting plants are of central
Ž .
importance Shipley et al., 1994; Ungar, 1996 . Rate of food intake emerges as a fundamental variable in foraging ecology since it determines the time herbivores spend gathering food to obtain required nutrients, and in an indirect manner the time they have
Ž .
to invest in other activities such as mating or avoiding predators Shipley et al., 1994 . Intake rate over short periods of active grazing is determined by bite weight and bite rate ŽAllden and Whittaker, 1970 . Traditionally, these variables have been regarded as a.
Ž
function of animal morphology, behaviour and sward structure Hodgson, 1981; Black .
and Kenney, 1984; Laca et al., 1994; Wilman et al., 1996 . For instance, intake per bite is influenced primarily by the sensitivity of bite depth to variation in sward height ŽHodgson et al., 1999 , and bite rate is a function of the time spent per bite on searching. Žlocomotion, recognition and decision and handling gathering herbage into the mouth,. Ž
. Ž .
severing the herbage, ingestive mastication and swallowing food Laca et al., 1994 . Besides their physical characteristics, plants present a complex array of biochemicals to herbivores. Recent research has shown that postingestive effects mediated by nutrients
Ž .
and toxins strongly influence food preference Provenza, 1995a,b, 1996 . Lambs can acquire preferences and increase consumption of low-quality forages such as wheat straw when straw is associated with intraruminal infusions of energy and nitrogen that
Ž
represent as little as 2.5% of their daily requirements Villalba and Provenza, 1996, .
1997a . Thus, it is conceivable that animals consume foods at rates that are also a function of the foods’ postingestive actions.
The majority of grazing studies have concentrated on the impact of a plant’s structure on intake rate, with very little reference to the potential importance of a plant’s
Ž
biochemical composition as a factor controlling intake rate Illius and Hodgson, 1996; .
Hodgson et al., 1999 . This has occurred even though heterogeneity in the nutritional
Ž .
quality of plant communities is frequent rather than peculiar Provenza et al., 1998 . The influence of nutrients on rates of food consumption has been overlooked primarily
Ž
because research on physical plant characteristics height, bulk density, structural .
strength has controlled for plant quality and because experiments were conducted under conditions where nutritional and toxicological differences among plants were not
Ž .
pronounced O’Reagain et al., 1996 .
2. Materials and methods
We conducted four trials at the Green Canyon Ecology Center, located at Utah State University in Logan. The objective of these trials was to determine if lambs modify their rate of straw consumption when they received intraruminal infusions of starch. As the trials progressed, we increased the degree of exposure to straw and we modified the temporal association between food ingestion and postingestive feedback in order to enhance the association between straw and starch. Herbivores constantly manifest
Ž .
dynamic and adaptive interactions with their environment Provenza et al., 1998 . We Ž
show the emergence of some of these interactions e.g., ingestive behaviour–postinges-.
tive feedback in response to the evolving experimental conditions of the study. The same animals and straw–starch or straw–water associations were maintained
Ž
throughout the study. During each trial, lambs Finn–Polypay–Suffolk crossbreds of .
both sexes had free access to salt blocks and fresh water. Alfalfa pellets were the basal diet. Lambs were approximately 3 months of age at the beginning of the study.
2.1. Before conditioning
Ž .
Eighteen lambs 25 kg BW were penned as a group in a 14 by 10 m plot. In order to familiarize the animals with the upcoming experimental procedures, every day at 0800 h
Ž .
pairs of lambs were selected at random and transferred to a testing pen 3 by 4.5 m . Lambs were then offered hand-clipped plants of alfalfa held erect with clamps between
Ž w x w x w x.
the major faces of the two pieces of lumber 13.5 cm H=120 cm W=3.5 cm D that formed a wooden frame.
Ž .
Lambs were exposed in pairs during 2 consecutive days 5 minrpairrday and
Ž .
individually 5 minrlambrday for the ensuing 3 days. After individual exposure, all lambs were consuming alfalfa from the wooden frames. Alfalfa plants were then
Ž .
replaced by wheat straw 100 g; 30 cm height placed along the central 30 cm of the wooden frame. During 4 consecutive days, pairs of lambs selected at random were exposed to straw in the testing pen for a period of 5 minrpairrday.
After exposure to straw, lambs were penned as a group and received an amount of
Ž .
alfalfa pellets 1270 grlambrday , which supplied about 80% of their digestible energy
Ž .
requirements NRC, 1985 .
2.2. Initial indiÕidual tests
The day after exposure to straw ended, lambs were tested individually between 0730 and 1000 h. Each lamb in the testing pen was offered a previously weighed amount of
Ž .
straw about 100 g secured in the wooden frame. We recorded the time each lamb spent eating straw during 5 min. Lambs were observed individually from a distance of approximately 3 m while they foraged. We excluded bouts of inactivity from calcula-tions of active feeding time. The total number of bites was recorded with a hand-counter while observing the lambs. We calculated intake by the difference in straw weight immediately before and after each lamb was tested. We used these data to calculate
Ž . Ž . Ž .
2.3. Trial 1
2.3.1. IndiÕidual tests
After the initial determination of straw intake, lambs were sorted in decreasing order by initial rate of straw intake and sets of two lambs were randomly assigned to two
Ž .
groups Groups 1 and 2; nine lambsrgroup . Thus, differences between groups due to initial recordings of intake rate were balanced.
Ž
Each lamb was offered a previously weighed amount of straw about 100 g; 30 cm .
height — secured in a wooden frame — during a 5-min period. Immediately after being tested for straw consumption, lambs in Group 1 were given intraruminal infusions of a starch suspension by oral intubation. The starch suspension was prepared by mixing 100 g of starch with 250 ml tap water at room temperature. Lambs in Group 2 received infusions of 250 ml tap water after consuming straw. Lambs from Groups 1 and 2 were tested in an alternate and consecutive sequence. The order in which individual lambs were tested was reversed each day. Tests were conducted between 0730 and 1000 h daily during 3 consecutive days. We calculated intake, intake rate, number of bites, bite rate and bite size during 5-min periods, as described previously. After the daily test, lambs were penned as a group and at noon they received alfalfa pellets, as described previously.
2.4. Trial 2
Individual lambs were reluctant to eat straw alone in the testing pen, which reduced the active feeding time and their exposure to straw. Consequently, during Trial 2 we attempted to increase exposure to straw — and thus enhance the association of straw with starch — by conditioning animals in pairs.
2.4.1. Conditioning — pairs
Four pairs of lambs from Group 1 and four pairs of lambs from Group 2 were formed at random. Each pair of lambs was transferred to the testing pen and offered wheat straw Ž100 g; 30 cm height secured in a wooden frame for 5 min. Pairs of lambs from Groups. 1 and 2 were tested in an alternate and consecutive order. The order of testing pairs of lambs was reversed each day. After testing, lambs in the pair received intraruminal
Ž . Ž .
infusions of starch 100 grlambrday; Group 1 or water 250 mlrlambrday; Group 2 . Lambs were penned as a group and at noon they received alfalfa pellets, as described for Trial 1.
During 4 consecutive days of conditioning, we calculated intake, intake rate, number of bites, bite rate and bite size for the 5 min period, as described for Trial 1. Pairs of lambs were the experimental units.
2.4.2. IndiÕidual tests
After conditioning in pairs, all lambs were tested as individuals and received
Ž . Ž .
2.5. Trial 3
During Trial 3 we attempted to enhance the association between straw and starch. First, we increased the exposure to straw by increasing the amount of time lambs were allowed to consume straw. We also allowed lambs to consume straw while they were with their peers.
We also modified the temporal ordering between straw offer and starch infusion by first infusing starch and then offering straw. This order may facilitate conditioning
Ž .
because the postingestive effects of starch i.e., the effects of absorbed calories occur
Ž .
during rather than after straw ingestion Boakes and Lubart, 1988 .
2.5.1. Conditioning — groups
At 0730 h, on the first day of conditioning lambs from Group 1 received intraruminal
Ž .
infusions of starch 100 grlambrday by oral intubation. Lambs were then transferred to the testing pen as a group and offered straw held erect in wooden frames. Condition-ing lasted 40 minrday and every 2 min we recorded the number of animals that were consuming straw. After conditioning Group 1, lambs from Group 2 received
intrarumi-Ž .
nal infusions of water 250 mlrlambrday and were offered straw in the same fashion described for Group 1. The order in which groups were conditioned was reversed each day. Four days of conditioning formed one conditioning period and two conditioning periods were performed. After the daily tests, lambs were penned as a group and at noon they received alfalfa pellets, as described for Trial 1.
The experimental area in the testing pen was prepared daily before each group of
Ž .
lambs was conditioned by placing about 2.5 kg of straw 30 cm height in three wooden frames of the same dimensions and characteristics described before.
2.5.2. IndiÕidual tests
After each 4-day conditioning period, all lambs were tested individually, as described
Ž . Ž
for Trial 1. Lambs received intraruminal infusions of starch Group 1 or water Group .
2 immediately before being offered straw secured in a wooden frame. Testing was performed during 1 day and we calculated intake, intake rate, number of bites, bite rate and bite size during a 5-min period, as described for Trial 1.
2.5.3. IndiÕidual tests without infusions
Lambs were tested as described before but without starch or water administrations.
2.6. Trial 4.
2.6.1. Conditioning
Ž .
The day after Trial 3 ended, lambs 33 kg BW were penned individually in adjacent Ž pens of 1.6=2.2 m. Lambs from Group 1 received intraruminal infusions of starch 100
. Ž
grlambrday and lambs from Group 2 received intraruminal infusions of water 250 .
mlrlambrday . Immediately after the infusions, all lambs were offered 100 g of
Ž .
Ž . during 9 consecutive days. At 1200 all lambs received alfalfa pellets 1400 grlambrday
Ž that provided approximately 80% of their daily digestible energy requirements NRC,
. 1985 .
2.6.2. IndiÕidual tests
After conditioning, and immediately before being tested for straw consumption,
Ž .
lambs in Group 1 were given intraruminal infusions of a starch 100 grlambrday and
Ž .
lambs in Group 2 received infusions of water 250 mlrlambrday . We recorded the time each lamb spent eating straw during 5 min. Lambs were observed individually and bouts of inactivity were excluded from calculations of feeding time. We determined the amount of straw ingested as the difference between the weight of food offered and that
Ž .
remaining after 5 min. These data were used to calculate intake rate grmin . At noon all lambs were offered alfalfa pellets, as described during conditioning.
2.6.3. IndiÕidual tests without infusions
Lambs were tested as described before but without starch or water administrations.
2.7. Statistical analyses
Straw intake, intake rate, number of bites, bite rate, bite size and active feeding time Ž
were analyzed as a split-plot design with lambs nested within groups. Group 1: starch .
or 2: no starch was the between-lamb factor. Pairs of lambs were the between-subject
Ž .
factor when lambs were conditioned as a pair Conditioning; Trial 2 . When tests were
Ž .
repeated on different days — Trial 1: 3 days; Trial 2: 4 days Conditioning — Pairs ;
Ž . Ž .
Trial 3: 2 days Individual Tests after Periods 1 and 2 ; Trial 4: 9 days Conditioning — day was the repeated measure in the analysis. Means were compared using the LSD test.
3. Results
3.1. Initial indiÕidual tests
There were no differences in straw intake between groups of lambs before applying
Ž . Ž .
the treatments starch, water P)0.05 . All the measurements recorded during testing
Ž .
were low and variable Table 1 .
3.2. Trial 1
During Trial 1, lambs’ ability to ingest wheat straw was not affected by infusing
Ž .
starch into the rumen P)0.05 . Intake values and active feeding times were still low
Ž .
Table 1
Straw intake, rate of straw intake, number of bites, bite rate and active feeding time by two groups of lambs that received or not intraruminal infusions of starch associated with straw ingestion
Straw Rate of No. of Bite Intake Active
intake straw intake bites rate per bite feeding time
Ž .g Žgrmin. Žbitesrmin. Žgrbite. Žmin.
Pairs of means within a column lacking a common superscript differ P-0.1 . c
G1: Group to be conditioned with infusions of starch during Trials 1 to 4. Group 2: Group to be conditioned with infusions of water during Trials 1 to 4.
d
3.3. Trial 2
3.3.1. Conditioning — pairs
In Trial 2, we increased the exposure to straw by conditioning animals in pairs. Lambs also had further experience with the postingestive consequences of starch and this treatment began to affect straw consumption. Pairs of lambs that received
intrarumi-Ž .
nal infusions of starch had higher intakes, intake rates P-0.05 and larger bite sizes ŽP-0.001 than lambs that had infusions of water Table 1 . The number of bites and. Ž .
Ž .
bite rate also increased, but the differences were not significant P)0.05; Table 1 .
3.3.2. IndiÕidual tests
Ž . Ž .
Lambs given starch ate more P-0.05 straw, at a faster rate P-0.1 , than lambs
Ž . Ž .
given water Table 1 . Lambs supplemented with starch also took more bites P-0.1
Ž . Ž .
of larger sizes P-0.05 than lambs infused with water Table 1 .
3.4. Trial 3
3.4.1. Conditioning — groups
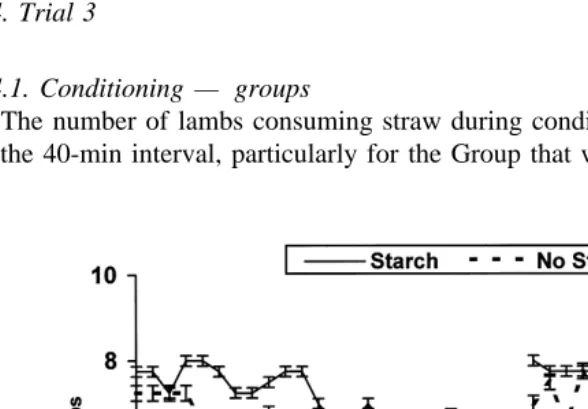
The number of lambs consuming straw during conditioning declined toward the end of the 40-min interval, particularly for the Group that was not infused with starch. The
Ž .
Fig. 1. Number of lambs ingesting straw secured in wooden frames; 30 cm height during two conditioning
Ž .
periods of 4 days each. Lambs were conditioned as two groups Groups 1 and 2 during 40 minrday. The number of lambs consuming straw was recorded every 2 min. Group 1 received intraruminal infusions of
Ž . Ž .
number of lambs consuming straw in the 2-min scans was in general higher for the
Ž .
group infused with starch than for the group infused with water Fig. 1 .
3.4.2. IndiÕidual tests
The effects of infusing starch became stronger in Trial 3 as lambs had further experience ingesting straw and receiving starch infusions. Lambs conditioned with
Ž . Ž .
starch consumed more straw P-0.01 and took more bites P-0.01 than lambs
Ž .
conditioned with water Table 1 . Moreover, after the second period of conditioning ŽPeriod 2 , lambs infused with starch increased straw intake from 21 to 27 g; P. Ž -0.05.
Ž . Ž
and number of bites from 24 to 35; P-0.05 compared to Period 1 Group by day .
interaction, P-0.05 .
Ž .
Lambs given starch ate more straw per unit of time P-0.001 , maintained a more
Ž . Ž .
rapid rate of biting P-0.01 , and took larger bites P-0.05 than lambs given water. Ž
Thus, active feeding time was the only variable not affected by the treatments P)0.05; .
Table 1 .
3.4.3. IndiÕidual tests without infusions
Ž .
Lambs given starch continued to display higher intakes P-0.001 , intake rates ŽP-0.001 , number of bites P. Ž -0.001 and bite rates P. Ž -0.01 than lambs given.
Ž .
water, even after starch was no longer administered Table 1 . Bite size and active
Ž .
feeding time did not differ between groups P)0.05; Table 1 .
Ž .
Fig. 2. Intake of straw 1–2 cm particle size by two groups of lambs during 9 days of conditioning. Lambs in Group 1 received intraruminal infusions of starch and lambs in Group 2 intraruminal infusions of water
Ž .
3.5. Trial 4
3.5.1. Conditioning
Groups did not differ in straw intake during the first 3 days of conditioning ŽP)0.05; Fig. 2 . After day 3, intake of straw was higher for lambs infused with starch. ŽGroup 1 than for lambs infused with water Group 2. Ž . ŽGroup by day interaction,
.
P-0.05; Fig. 2 . Average intake of straw across days was 44 g for lambs in Group 1
Ž .
and 27 g for lambs in Group 2 P)0.05; S.E.M.s10 .
3.5.2. IndiÕidual tests
Lambs conditioned with starch ate more straw at higher rates than lambs conditioned
Ž .
with water P-0.10; Table 1 .
3.5.3. IndiÕidual tests without infusions
Ž . Ž .
Lambs treated with starch ate more straw P-0.10 , at faster rates P-0.15 , and Ž
spent more time feeding during the 5-min tests than lambs treated with water P-0.10; .
Table 1 , even when starch infusions were suspended.
4. Discussion
4.1. PostingestiÕe feedback and ingestiÕe behaÕiour
It has been proposed that the potential rate at which a food can be eaten is largely
Ž .
determined by the food’s physical characteristics Kenney and Black, 1984 . However, under most conditions, food structure and quality are confounded because foods with high nutrient content are typically easy to ingest and highly digestible, whereas plants with low nutrient content are usually difficult to ingest and poorly digestible.
Thus, we sought to isolate the effects of a plant’s structure from its postingestive effects. We hypothesized that rates of food intake are not only determined by the physical structure of a forage but also by its postingestive actions. We found that groups of lambs exposed to the same forage — wheat straw — for the same periods of time showed quite different patterns of ingestive behaviour, dependent on the postingestive consequences during straw ingestion. Lambs that received intraruminal infusions of
Ž .
starch increased intake, intake rate, bite rate and intake per bite Table 1 . Lambs manifested this behaviour even when starch was no longer supplied during testing ŽTable 1 . Lambs conditioned with starch continue to prefer flavoured straw Villalba. Ž
.
and Provenza, 1997a , even when starch is no longer administered, which suggests that the postingestive effects of energy increase the hedonic value or liking and make preferences — and ingestive behaviour in this study — more resistant to extinction.
Most studies of grazing have focused on the influences of food structure on intake
Ž .
rate Hodgson et al., 1999 , and they have manipulated sward structure rather than sward nutritional quality. The consensus is that other factors — nutritional and toxicological
Ž .
— being equal, intake rate is largely determined by bite mass Ungar et al., 1991 , Ž
. Ž .
per unit canopy volume Black and Kenney, 1984; Laca et al., 1992 . For instance, sward height has been proposed to affect bite depth and consequently intake per bite ŽGong et al., 1996; Hodgson et al., 1999 ..
Ž
In contrast, our study shows that when physical structure is held constant e.g., same .
forage, same height and density — and nutritional effects are unequal — intake rate, bite rate and bite size are variable and largely influenced by the postingestive conse-quences of food ingestion. Rates of straw intake were low and variable when no starch was infused in the lambs’ stomach, which supports previous work suggesting that
Ž .
animals forage at variable rates, often lower than their potential Owen-Smith, 1993 . Low rates of intake are more likely to occur when foods supply low amounts of macronutrients and other foods of higher quality are available for consumption.
Ž .
Rates of food ingestion are elastic Owen-Smith, 1993 , and influenced by an
Ž .
animal’s physiological state Newman et al., 1994; Villalba and Provenza, 1999a . Fasted animals can compensate for increased nutrient demands by increasing bite rate
Ž .
and bite size Paterson et al., 1998 . Even though larger bites require more mastication, Ž
cattle can reduce the amount of chewing to increase intake rate Greenwood and .
Demment, 1988 . Increases in chewing due to increases in bite mass can also be Ž
compensated by using jaw movements for both cropping and chewing Laca et al., .
1994 .
Besides the general effects of fasting, specific nutritional needs can also modify rates
Ž .
of food intake. Intake rates of an energy-dense food barley are higher in lambs after a
Ž .
meal high in protein than after fasting for 15 h Villalba and Provenza, 1999a . This response is likely due to an increased need to balance the excess nitrogen supplied by the protein-rich meal with rapidly fermentable carbohydrates in barley. In the current study, energy-restricted lambs increased their rate of straw intake when straw was paired with starch. Collectively, these results suggest that rates of food intake are dynamic and depend on the nutrient composition of the food and the animal’s physiological condition. The plasticity of rates of food intake are also manifest through their dependence on an animal’s past history. Foraging skills, acquired through experience, have a significant impact on foraging behavior by increasing the efficiency of food consumption and by
Ž
enhancing rates of forage intake Provenza and Balph, 1988; Flores et al., 1989; .
Ortega-Reyes and Provenza, 1993 .
4.2. PostingestiÕe feedback and food choice
Food preferences also have been regarded as a function of the rate at which foods can be ingested. Other factors being equal, plants that can be eaten quickly are likely to be
Ž .
preferred to tougher species Wilman and Asiedu, 1983; Kenney and Black, 1984 .
Ž . Ž .
Cattle Distel et al., 1995 and sheep Black and Kenney, 1984 select those patches that allow higher intake rates from swards that differ in height and density. Thus, in horizontally heterogeneous pastures of similar quality, animals may be able to assess the
Ž .
reward i.e., postingestive consequences from a patch by sensing the rate at which food Ž
can be obtained from the patch, and modify their preferences accordingly Distel et al., .
1995 . However, when sward quality is variable, nutrient composition can be even more
Ž . Ž
.
Illius et al., 1987 swards patches may be preferred when their nutritional quality is higher. Foods that yield lower intake rates can be preferred by lambs fed unbalanced basal diets when those foods provide ratios of energyrprotein closer to the lambs’
Ž .
nutritional needs Villalba and Provenza, 1999a . Thus, rates of food intake should be considered an emergent property of the interaction between the structure and biochemi-cal composition of food.
A trade-off between diet quality and intake rate has been found for ewes grazing
Ž .
cocksfoot swards. In order to consume higher quality components leaves , ewes must take smaller bites which promotes lower intake rates; less selective ewes take larger
Ž .
bites and have higher intake rates to the detriment of diet quality Prache et al., 1998 . Thus, degree of selectivity can influence rates of food intake. Rates of food intake are then a result, rather than a cause, of food selection. We have shown that lambs ingest wheat straw at rates lower than their potential — when offered straw paired with water, presumably because of a lack of reward during and after straw ingestion. Conversely, lambs increased straw ingestion — bite rate, bite size, number of bites — when they received infusions of starch. If the food preferences of ruminants are controlled by specific associations between a food’s flavour and the food’s postingestive actions ŽProvenza, 1995a,b, 1996; Villalba and Provenza, 1996, 1997a,b , then it is likely that. once a particular food is preferred, postingestive feedback from nutrients may enhance rates of food consumption. In contrast, when preferred items become less accessible, intake per bite will decrease and animals may lower their intake rate.
4.3. PostingestiÕe feedback and palatability
Palatability is classically defined to be a function of the taste, odour and surface
Ž .
characteristics of foods Arnold, 1981 . Within this context, it has been proposed that palatability is probably not an important determinant of intake for better-quality
Ž
roughages, but it may limit intake of poor-quality roughages like straw Greenhalgh and .
Reid, 1971 . This was concluded from studies where sheep were fed straw while straw was put through a fistula in the rumen. Under these conditions, sheep were extremely reluctant to eat straw, but they significantly increased straw intake when grass hay, a
Ž
more nutritious food, was put into the rumen replacing straw Greenhalgh and Reid, .
1971 .
We also found that straw intake was enhanced when lambs had an increased supply
Ž .
of macronutrients to the rumen starch . However, we differ in the interpretation of the results. Palatability is best understood as a compound phenomenon emerging from the
Ž .
interrelationship between the chemical senses taste, smell and postingestive feedback ŽProvenza, 1995a,b, 1996 . Thus, palatability per se is probably not the main reason for. the low consumption of straw displayed by sheep, but an outcome of the association
Ž .
between organoleptic characteristics of straw e.g., flavour and the low amount of nutrients supplied to the rumen during and after straw ingestion. Decreases in preference for straw due to a lack of nutrient feedback can explain why sheep become increasingly
Ž .
reluctant to eat straw Greenhalgh and Reid, 1971 , and why lambs given water displayed lower intake rates as exposure to straw increased. Conversely, lambs increase
Ž .
when straw ingestion is followed by intraruminal infusions of starch. This increase can be interpreted as an enhanced incentive or drive to eat straw due to its association with a readily available source of energy.
5. Conclusions
The postingestive effects of energy have a significant effect on food intake. Rates of food ingestion are not only influenced by the structure of a forage, but also by the amount of nutrients that the forage supplies. Lack of feedback from the gut during food consumption is one of the main reasons for the low preferences and intake rates displayed for low-quality foods like straw. Thus, we suggest that food quality should be considered in conjunction with food structure to better understand intake rate of grazing herbivores.
Acknowledgements
This research was supported by grants from CSREES and the Utah Agricultural Experimental Station, Utah State University. Approved as journal paper number 7160.
References
Allden, W.G., Whittaker, I.A.McD., 1970. The determinants of herbage intake by grazing sheep: the interrelationships of factors influencing herbage intake and availability. Aust. J. Agric. Res. 21, 755–766.
Ž .
Arnold, G.W. 1981. Grazing behaviour. In: Morley, F.H.W. Ed. , Grazing Animals. Elsevier, Amsterdam, pp. 79–104.
Ž .
Bazely, D.R., 1990. Rules and cues used by sheep foraging in monocultures. In: Hughes, R.N. Ed. , Behavioural Mechanisms of Food Selection. NATO ASI Series, Vol. G20, Springer-Verlag, Berlin, pp. 343–367.
Black, J.L., Kenney, P.A., 1984. Factors affecting diet selection by sheep: II. Height and density of pasture. Aust. J. Agric. Res. 35, 565–578.
Boakes, R.A., Lubart, T., 1988. Enhanced preference for a flavour following reversed flavour–glucose pairing. Q. J. Exp. Psychol. 40B, 49–62.
Distel, R.A., Laca, E.A., Griggs, T.C., Demment, M.W., 1995. Patch selection by cattle: maximization of intake rate in horizontally heterogeneous pastures. Appl. Anim. Behav. Sci. 45, 11–21.
Flores, E.R., Provenza, F.D., Balph, D.F., 1989. Relationship between plant maturity and foraging experience of lambs grazing hycrest crested wheatgrass. Appl. Anim. Behav. Sci. 23, 279–284.
Gong, Y., Hodgson, J., Lambert, M.G., Gordon, I.L., 1996. Short-term ingestive behaviour of sheep and goats grazing grasses and legumes: 2. Quantitative relationships between sets of sward and ingestive behaviour variables. N. Z. J. Agric. Res. 39, 75–82.
Greenhalgh, J.F.D., Reid, G.W., 1971. Relative palatability to sheep of straw, hay and dried grass. Br. J. Nutr. 26, 107–116.
Greenwood, G.B., Demment, M.W., 1988. The effect of fasting on short-term cattle grazing behaviour. Grass Forage Sci. 43, 377–386.
Hodgson, J., Cosgrove, G.P., Woodward, S.J.R., 1999. Research on foraging strategy: progress and priorities. Proceedings of the XVIII International Grassland Congress, in press.
Illius, A.W., Hodgson, J., 1996. Progress in understanding the ecology and management of grazing systems.
Ž .
In: Hodgson, J., Illius, A.W. Eds. , The Ecology and Management of Grazing Systems. Commonwealth Agricultural Bureau International, Wallingford, UK, pp. 429–457.
Illius, A.W., Wood-Gush, D.G.M., Eddison, J.C., 1987. A study of the foraging behaviour of cattle grazing patchy swards. Biol. Behav. 12, 33–44.
Kenney, P.A., Black, J.L., 1984. Factors affecting diet selection by sheep: I. Potential intake rate and acceptability of feed. Aust. J. Agric. Res. 35, 551–563.
Laca, E.A., Ungar, E.D., Seligman, N., Demment, M.W., 1992. Effects of sward height and bulk density on bite dimensions of cattle grazing homogeneous swards. Grass Forage Sci. 47, 91–102.
Laca, E.A., Ungar, E.D., Demment, M.W., 1994. Mechanisms of handling time and intake rate of a large mammalian grazer. Appl. Anim. Behav. Sci. 39, 3–19.
Newman, J.A., Penning, P.D., Parsons, A.J., Harvey, A., Orr, R.J., 1994. Fasting affects intake behaviour and diet preference of grazing sheep. Anim. Behav. 47, 185–193.
NRC, 1985. Nutrient Requirements of Sheep 6th edn. National Academy Press, Washington, DC.
O’Reagain, P.J., Goetsch, B.C., Owen-Smith, R.N., 1996. Effect of species composition and sward structure
Ž .
on the ingestive behaviour of cattle and sheep grazing South African sourveld. J. Agric. Sci. Cambridge 127, 271–280.
Ortega-Reyes, L., Provenza, F.D., 1993. Experience with blackbrush affects ingestion of shrub live oak by goats. J. Anim. Sci. 71, 380–383.
Owen-Smith, R.N., 1993. Evaluating optimal diet models for an African browsing ruminant, the kudu: how constraining are the assumed constraints?. Evol. Ecol. 7, 499–524.
Paterson, D.M., McGilloway, D.A., Cushnahan, A., Mayne, C.S., Laidlaw, A.S., 1998. Effect of duration of fasting period on short-term intake rates of lactating dairy cows. Anim. Sci. 66, 299–305.
Prache, S., Roguet, C., Petit, M., 1998. How degree of selectivity modifies foraging behaviour of dry ewes on reproductive compared to vegetative sward structure. Appl. Anim. Behav. Sci. 57, 91–108.
Provenza, F.D., 1995a. Postingestive feedback as an elementary determinant of food preference and intake in ruminants. J. Range Manage. 48, 2–17.
Provenza, F.D., 1995b. Role of learning in food preferences of ruminants: Greenhalgh and Reid revisited. In:
Ž .
Engelhardt, W.V., Leonhard-Marek, S., Breves, G., Giesecke, D. Eds. , Ruminant Physiology: Digestion, Metabolism, Growth and Reproduction. Proceedings of the VIII International Symposium on Ruminant Physiology. Ferdinand Enke Verlag, Stuttgart, Germany. pp. 233–247.
Provenza, F.D., 1996. Acquired aversions as the basis for varied diets of ruminants foraging on rangelands. J. Anim. Sci. 74, 2010–2020.
Provenza, F.D., Balph, D.F., 1988. The development of dietary choice in livestock on rangelands and its implications for management. J. Anim. Sci. 66, 2356–2368.
Provenza, F.D., Villalba, J.J., Cheney, C.D., Werner, S.J., 1998. Self-organization of foraging behavior: from simplicity to complexity without goals. Nutr. Res. Rev. 11, 199–222.
Shipley, L.A., Gross, J.E., Spalinger, D.E., Hobbs, N.T., Wunder, B.A., 1994. The scaling of intake rate in mammalian herbivores. Am. Nat. 143, 1055–1082.
Ž .
Ungar, E.D., 1996. Ingestive behaviour. In: Hodgson, J., Illius, A.W. Eds. , The Ecology and Management of Grazing Systems. Commonwealth Agricultural Bureau International, Wallingford, UK, pp. 185–218. Ungar, E.D., Genizi, A., Demment, M.W., 1991. Effects of sward height, bulk density and height
heterogene-ity on bite dimensions and the short-term intake rate of cattle. Agron. J. 83, 973–978.
Villalba, J.J., Provenza, F.D., 1996. Preference for flavored wheat straw by lambs conditioned with intraruminal administrations of sodium propionate. J. Anim. Sci. 74, 2362–2368.
Villalba, J.J., Provenza, F.D., 1997a. Preference for wheat straw by lambs conditioned with intraruminal infusions of starch. Br. J. Nutr. 77, 287–297.
Villalba, J.J., Provenza, F.D., 1997b. Preference for flavored foods by lambs conditioned with intraruminal administrations of nitrogen. Br. J. Nutr. 78, 545–561.
Villalba, J.J., Provenza, F.D., 1999b. Nutrient-specific preferences by lambs conditioned with intraruminal infusions of starch, casein, and water. J. Anim. Sci. 77, 378–387.
Wilman, D., Asiedu, F.H.K., 1983. Growth, nutritive value and selection by sheep of sainfoin, red clover,
Ž .
lucerne, and hybrid ryegrass. J. Agric. Sci. Cambridge 100, 115–126.