www.elsevier.comrlocaterapplanim
Intake and selection for white clover by grazing
lambs in response to gastrointestinal parasitism
G.P. Cosgrove
), J.H. Niezen
AgResearch, Grasslands Research Centre, PB 11008, Palmerston North, New Zealand
Accepted 6 July 1999
Abstract
Lambs indoors seek to compensate the metabolic protein deficiency caused by intestinal parasites by selecting a diet with a higher concentration of protein. A study was conducted to determine if lambs grazing at pasture would enhance their nutrient intake in response to parasitism, by selecting more white clover from a mixed grass–clover pasture. The trial used a split-plot design with two grass–clover mixtures as main plots, two levels of parasite infection as
Ž
the split-plot, and two replications. Lambs with a low level of infection suppressive drenched .
with anthelmintic at 2-weekly intervals to restrict parasite infection; SD or with a moderate level Ž
of infection drenched whenever group mean faecal egg count reached a trigger level 1000 eggs
. Ž .
per g fresh faeces; TD grazed together on either perennial ryegrass Lolium perenne L. and
Ž . Ž . Ž .
white clover Trifolium repens L. RWC , or on browntop Agrostis capillaris L. and white
Ž . Ž .
clover BWC . Plots were continuously stocked with trial lambs from early-summer January to
Ž .
early-winter June . Additional put-and-take lambs were used to maintain sward surface height at 5 cm. A sub-group of 12 lambs, comprised of 4 SD and 8 TD, similar in mean liveweight to their cohorts, was used to determine individual herbage intake, diet composition and diet in-vivo digestibility over a 10-day period in autumn. This period was timed to coincide with the rise towards the trigger level in the egg count of TD lambs. Compared with lambs grazing BWC, those
Ž y1 .
grazing RWC had higher liveweight gain 280 vs. 156 g day ; Ps0.06 and higher herbage
Ž y1 y1 .
intake 1.57 vs. 1.17 kg DM hd day ; Ps0.003 . The proportion of clover in the diet was
Ž .
higher for TD than for SD lambs on BWC 31% vs. 24% , but not on RWC, where the diet
Ž . Ž
contained a higher proportion of white clover 51% for both levels of parasitism species=drench
.
interaction, Ps0.02 . For each treatment, the proportion of white clover in the diet was higher
Ž .
than in the sward. The higher proportion of clover 30% vs. 20% of green DM and lower bulk
Ž y1 y1
density of the RWC swards compared with BWC 180 vs. 310 kg DM ha cm of pasture
)Corresponding author.
0168-1591r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved.
Ž .
( ) G.P. CosgroÕe, J.H. NiezenrApplied Animal BehaÕiour Science 66 2000 71–85
72
.
canopy height assisted RWC lambs in selecting a higher proportion of clover. However, with this higher proportion of clover in their diet, parasitism did not stimulate selection. It is suggested that in the complex foraging environment outdoors, grazing lambs can detect metabolic signals associated with parasitism and make a behavioural response to mitigate the harmful nutritional
effects.q2000 Elsevier Science B.V. All rights reserved.
Keywords: Diet selection; Grazing; White clover; Herbage intake; Gastrointestinal parasites; Lambs
1. Introduction
Gastro-intestinal parasitism disrupts protein metabolism and depresses feed intake ŽPoppi et al., 1986; Poppi et al., 1990 . Recent studies with healthy lambs offered feed. choices indoors shows that they can select a diet that meets their protein requirements
Ž
for fast growth and avoid excessive protein intake Hou et al., 1991; Kyriazakis and .
Oldham, 1993 . Further, in response to intestinal parasitism, they will select a diet with
Ž .
higher protein content Kyriazakis et al., 1994 . These results from studies indoors suggest a nutritional basis for food choices. However, the environment for foraging is more complex outdoors, yet links between parasitism and nutrition have considerable importance for both experimentation and management. The ability of grazing animals to detect a metabolic protein imbalance resulting from parasitism, and to respond to it by changing diet selection among pasture species, has not been tested.
Ž Sheep alter grazing behaviour in response to faecal contamination of pasture
Hutch-.
ings et al., 1998 . In that study, parasitised sheep grazed more selectively from artificial swards of ryegrass than non-parasitised sheep by taking smaller bites at a slower rate interpreted by those authors as an indication of parasite avoidance behaviour. This suggests that parasitised sheep outdoors are capable of adjusting grazing behaviour, although, in this case, not to improve nutrient intake. However, together these and the above quoted studies suggest that sheep might be capable of exploiting differences between plant species to improve their nutrient intake. For lambs grazing mixed grass–clover pasture, there is an opportunity to alter dietary protein in response to parasitism by selecting for plant species with a higher protein content, or species that
Ž .
can be eaten at a faster rate. White clover Trifolium repens L. has protein character-istics that may satisfy the nutrient demands arising from parasitism. It has consistently high nitrogen in its leaves, and while it does not necessarily have a higher nitrogen
Ž .
concentration in the leaves than ryegrass Lolium perenne L. , more of this nitrogen
Ž .
reaches the lower gastro-intestinal tract Ulyatt et al., 1988 . Sheep can eat white clover
Ž .
at a faster rate than ryegrass when offered as monocultures Penning et al., 1991b . This would be an added advantage for increasing protein intake, but selective grazing to obtain a desired component from a mixed-species pasture involves a cost to the animal ŽParsons et al., 1994b . This cost is normally incurred through reduced short-term rate of. intake as an animal searches for desired food items, and results in either lower daily intake, or a need for longer grazing time to achieve the same daily intake. The amount of a preferred food item eaten by sheep is influenced by availability, in both the
Ž .
preferred component in the pasture decreases, the cost of selective grazing increases and Ž
the proportion in the diet is likely to decline see Parsons et al., 1994b; Thornley et al., .
1994 .
The objective of this study was to test the hypothesis that in response to intestinal parasitism, grazing lambs will increase the proportion of white clover in their diet as a means of increasing protein intake, and that the extent of this selection for clover will depend on the grass species available as the dietary alternative.
2. Animals, materials and methods
This study was conducted during autumn 1996 at AgResearch Flock House,
approxi-Ž .
mately 50 km west of Palmerston North latitude 408S , in a moist, temperate environ-ment. The trial design was a 2=2 split-plot, with pasture species as main plots, parasite infection level as the split-plot, and there were 2 replications.
2.1. Pastures
Ž .
In the autumn of 1995 March , two species of grass with differing morphology,
Ž . Ž
browntop Agrostis capillaris L. cv. Muster , and perennial ryegrass cv. Grasslands
. Ž .
Nui were each established with white clover cv. Grasslands Tahora on plots of 0.8 ha, with two replicates for each grass–clover combination. Browntop and ryegrass differ in
Ž .
tiller morphology, with browntop tending to form densely tillered swards Harris, 1973 . They also differ in aspects of larval ecology, with pure swards of browntop having a
Ž .
higher density of larvae Trichostrongylus colubriformis and Ostertagia circumcincta
Ž .
in the upper stratum of the sward, compared with ryegrass Niezen et al., 1998 . The Ž
trial area was situated on a recent alluvial sandy-loam soil Rangitikei series soil type, .
Cowie et al., 1967 . Each plot received a standard annual fertiliser dressing of 150 kg hay1
of sulphur-enriched superphosphate in the spring.
The swards were cut and harvested for silage 1 month before the trial started. One week prior to the start of the trial, the pastures were grazed by ewes with a low
Ž Ž . y1.
nematode burden mean faecal egg count FEC 300 eggs g , at a stocking density of 94 ewes hay1 for 24 h to provide each paddock with a light, even contamination of
Ž
gastro-intestinal nematode eggs. During the trial, a variable stocking rate Mott and .
Lucas, 1952 was used to maintain a sward surface height of 5 cm. Non-trial, put-and-take lambs, similar in weight and parasite status to trial lambs, were added or removed as necessary to maintain uniform sward conditions across treatments.
Ž .
Pasture height was measured twice weekly, using a sward stick Bircham, 1981 , by taking a minimum of 25 measurements per plot. Herbage mass was measured monthly, and hand-pluck samples were taken monthly for separation into botanical components. Additional hand-pluck samples were taken weekly by the zig-zag traverse method ŽTaylor, 1939 to determine larval density on pasture. Third-stage nematode larvae were.
Ž
counted after separation from herbage by a modified Baermann procedure Anderson .
( ) G.P. CosgroÕe, J.H. NiezenrApplied Animal BehaÕiour Science 66 2000 71–85
74
2.2. Animals
Recently-weaned Romney ram lambs were randomly allocated to treatments based on Ž w .
liveweight, drenched with ivermectin Ivomec , Merial and then grazed on the trial
Ž .
swards for 134 days January 31 to June 14 . Lambs were weighed and faeces sampled at 2-weekly intervals. To create the parasite infection treatment for each pasture type, one-third of the lambs were drenched with ivermectin at 2-weekly intervals to suppress
Ž .
parasite infection suppressive drench, SD . Two-thirds were allowed a moderate level of infection, but were drenched when mean FEC reached 1000 eggs gy1 of fresh faeces
Žepg on any treatment, in order to prevent debilitating infection trigger drench, TD .. Ž . These TD-lambs required anthelmintic drenching on February 3, March 17, and May 5. A greater number of lambs were allocated to the TD treatment because earlier experiments had shown greater variability in liveweight gain in parasitised than in non-parasitised lambs. Put-and-take lambs were added or removed in multiples of three to maintain a 2:1 ratio of TD to SD lambs. All anthelmintic treatments were given at 300
y1 Ž .
mg kg liveweight 1.5 times the recommended dose rate . Faecal egg counts were
Ž .
done using a modified McMaster technique Whitlock, 1943 , in which one egg counted represented 50 eggs gy1 fresh faeces.
To characterise the parasite species being ingested, lambs that had been reared
Ž .
indoors free of nematode infection tracer lambs were grazed on the swards for 3 weeks, and then housed for a further 3 weeks to allow ingested larvae to mature. The lambs were then slaughtered and the abomasa and intestine removed to determine worm burden. Digesta was removed from these organs by washing, rinsed through a 60 m sieve, and 5% aliquots were taken from the washings for adult worm identification and counting, and for counting of fourth-stage larvae. The following genera were identified;
Ž
Haemonchus, Ostertagia, and Trichostrongylus in the abomasum mean 6600 nema-y1.
todes lamb and Trichostrongylus, Nematodirus, and Cooperia in the small intestine Žmean 11300 nematodes lamby1.. There were no significant differences between lambs from ryegrass or from browntop in the number of each species or in the total number of nematodes.
2.3. Diet selection and intake
Sub-groups of 12 lambs grazing each replicate of the ryegrass and the browntop plots were selected to represent the mean liveweight for all lambs on the plot. These sub-groups were comprised of 4 SD, and 8 TD lambs. These lambs received an
Ž .
intra-ruminal controlled-release capsule Captec NZ containing synthetic C32 and C36 Ž .
alkanes, by oral administration on April 12 day 0 . Faecal samples were taken from each dosed lamb, by rectal grab sampling, on days 10, 12, 14, 17, and 19 after dosing. These samples were freeze-dried, ground to pass a 1 mm sieve, and bulked on an equal dry weight basis across days. This resulted in a single faecal sample for each lamb which was then analysed for concentration of individual alkanes ranging in chain length
Ž .
Ž . gas chromatography, as were the faecal samples. The ratios of even chain synthetic
Ž .
and odd-chain alkanes naturally occurring in the plant cuticular wax in the faeces and herbage were used to derive estimates for intake, proportion of grass and clover in the diet and in-vivo digestibility. Intake was calculated using C32 and C33 alkanes, and in vivo digestibility using C35 and C36. Diet composition was estimated using a
least-Ž .
squares optimisation procedure EatWhat, Dove and Moore, 1995 . Diet composition, intake and digestibility estimates are mean values for the 10-day period over which the faecal samples were collected. A further sub-sample of the ryegrass and browntop herbage was analysed for total nitrogen concentration.
Liveweight gain was measured over the 2-week period that encompassed the faecal sampling period, in order to relate these estimates to animal performance. At the start of
Ž .
the intake measurement period lambs on browntop Agrostis capillaris L. and white
Ž . Ž .
clover BWC were heavier than those on perennial ryegrass Lolium perenne L. and
Ž . Ž . Ž .
white clover Trifolium repens L. RWC 37.6 kg vs. 34.2 kg, Ps0.002 , but SD
Ž .
lambs did not differ significantly from TD 36.8 kg vs. 35.0 kg, Ps0.19 . At the end of the measurement period, the difference in liveweight between BWC and RWC had
Ž .
diminished 39.8 kg vs. 38.2 kg, Ps0.07 . Daily intake was analysed on the basis of per animal, per unit of liveweight, and per unit of metabolic liveweight. Statistically similar comparisons were obtained in each case, and results are presented only for intake per animal.
2.4. Statistical analysis
For animal-related variables, group means for each treatment were compared by
Ž . Ž .
analysis of variance ANOVA , using a split-plot model 7 df in which grass species Ž . formed the main-plot and drench regime the split-plot. Grass species effect 1 df was
Ž . Ž .
tested against replicate x grass species effect 2 df and the drench 1 df and
Ž . Ž .
drench=species effect 1 df was tested against residual error 2 df . Main-plot means for pasture variables and larval density, and the trigger-drenched group mean faecal egg
Ž . Ž .
count after log10 transformation were compared by one-way ANOVA 3 df with grass Ž .
species 1 df tested against residual error.
3. Results
3.1. LiÕeweight gain and herbage intake
Ž . Ž .
Liveweight gain was higher Ps0.06 on RWC than on BWC Table 1 , but did not differ between suppressive or trigger-drenched lambs. Daily feed intake was higher ŽPs0.003 for lambs grazing RWC than for those grazing BWC Table 1 . The effect. Ž . of anthelmintic regime on intake was not significant, however there was a tendency for lambs receiving the suppressive drench to eat more than trigger-drenched lambs when
Ž
on RWC but a reverse trend on BWC grass species=anthelmintic drench interaction, .
()
G.P.
Cosgro
Õ
e,
J.H.
Niezen
r
Applied
Animal
Beha
Õ
iour
Science
66
2000
71
–
85
76
Table 1
Effect of grass species and anthelmintic drenching regime on the liveweight gain, daily herbage intake, proportion of clover in the diet and in vivo digestibility of the diet of grazing lambs
Item Anthelminitic status Grass species Statistical significance
a b
Ryegrass Browntop S.E.D. S.E.D. P value
Species Drench Species=Drench
y1 y1
Ž .
Liveweight gain g hd day Suppressive 271 186 46.6 72.7 0.06 0.60 0.35
Trigger 290 126
y1 y1
Ž .
Intake kg DM hd day Suppressive 1.65 1.10 0.074 0.086 0.003 0.86 0.11
Trigger 1.49 1.24
Clover proportion Suppressive 0.53 0.24 0.012 0.237 0.18 0.20 0.02
Trigger 0.49 0.31
Ž .
Digestibility % Suppressive 68 60 2.64 7.10 0.48 0.42 0.14
Trigger 65 67
a
S.E.D.sstandard error of difference between means for anthelmintic status within grass species. b
Fig. 1. Mean sward surface height of ryegrass–white clover and browntop–white clover treatments, compared with the target height, during summer and autumn, and for the duration of the intake measurement period
Žindicated by arrows ..
3.2. Proportion of cloÕer in diet
Ž .
The proportion of clover in the diet Table 1 was higher for trigger-drenched than for Ž
suppressive-drenched lambs grazing on BWC grass species=anthelmintic drench .
interaction, Ps0.02 . In contrast, trigger-drenched and suppressive-drenched lambs had a similar proportion of clover in their diet on RWC. For both trigger-drenched and suppressive-drenched lambs, the proportion of clover in the diet was higher on RWC than on BWC. In vivo digestibility was not significantly affected by grass species or anthelmintic treatment, although the lowest recorded value was for the
suppressive-Ž . drenched lambs grazing on BWC Table 1 .
Table 2
Ž .
Herbage mass, total lamb liveweight and herbage allowance for ryegrass-white clover RWC and
browntop-Ž .
white clover BWC swards
Pasture treatment Statistical significance a
RWC BWC S.E.M. P value
y1
Ž .
Herbage mass kg DM ha April 905 1555 162.7 0.11
May 960 1560 88.5 0.05
y1
Ž .
Total lamb liveweight kg LW ha April 386 530 5.76 0.003
May 466 479 11.8 0.50
y1
Ž .
Herbage allowance kg DM kg LW April 2.35 2.94 0.31 0.31
May 2.07 3.26 0.23 0.06
a
( ) G.P. CosgroÕe, J.H. NiezenrApplied Animal BehaÕiour Science 66 2000 71–85
78
3.3. Pasture characterisation
Ž Sward height was slightly less than the target height of 5 cm during the period April
. Ž .
22–May 1 when intake measurements were made Fig. 1 . Total herbage mass was Ž .
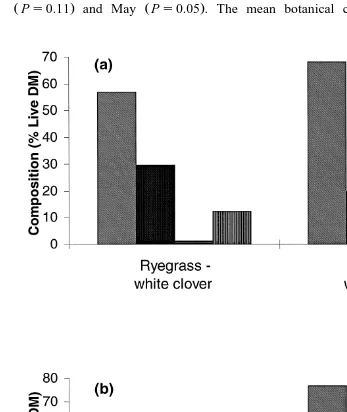
stable through April and May Table 2 , but was lower for RWC than BWC during April ŽPs0.11. and May ŽPs0.05 . The mean botanical composition of the swards,.
Ž . Ž .
Table 3
Ž .
Nematode larval density on pasture swards and faecal egg counts of trigger-dosed lambs natural means
Ž . Ž .
grazing swards of ryegrass-white clover RWC and browntop-white clover BWC
a
Larval density number L3 larvae kg DM April 2260 9870 7897 0.51
May 3800 1110 1184 0.12
y1
Ž .
Faecal egg count eggs g fresh faeces March 3110 4000 3498 0.35
May 840 1190 777 0.23
June 300 260 321 0.65
a
Analysis of variance performed on Log10 transformed data. b
S.E.M.sstandard error of means.
Ž
determined from the pluck samples taken in late March before the intake measurement
. Ž .
period and in early May at the end of the intake measurement period are shown in Ž
Fig. 2. Both swards contained a high proportion of dead matter 74% and 34% for RWC, and BWC, respectively, before intake measurement, and 17% and 19% for RWC and
.
BWC, respectively, after intake measurement . To more accurately describe the compo-sition available for grazing, sown grass, white clover, unsown grass and other unsown species are expressed as proportions of the live fraction. On this basis, the proportion of white clover was 30% for RWC and 20% for BWC before intake measurement, and
Ž . 14% for both RWC and BWC after intake measurement Fig. 2 .
The nitrogen concentration of ryegrass and browntop was 52 and 53 g kg DMy1,
Ž .
respectively data not shown . No corresponding analysis was conducted for white clover because insufficient dried herbage remained after analysis for alkane concentra-tion.
( )
3.4. LarÕal L3 density on pasture and faecal egg counts
Estimates of larval density showed no consistent pattern with time, and did not differ Ž . significantly between grass species for the estimates made in April and May Table 3 .
Ž .
The faecal egg count for TD lambs declined from March Table 3 . The slightly higher count for BWC compared with RWC lambs in March and May was not significant.
4. Discussion
4.1. Effects of parasitism on selection and intake
( ) G.P. CosgroÕe, J.H. NiezenrApplied Animal BehaÕiour Science 66 2000 71–85
80
lambs on these plots were also higher, consistent with the low liveweight gains particularly for the trigger-drench regime. These lambs selected more clover than did the lambs receiving suppressive-drench, although this improvement in diet quality was insufficient to completely overcome the detrimental effects of parasitism and increase liveweight gain to the rate of their non-parasitised cohorts. Parasitism did not reduce
Ž .
intake, in contrast to indoor trials Poppi et al., 1990; Kyriazakis et al., 1994 . There are two possible explanations for this difference. First, the parasitised lambs received some anthelmintic treatment, and the non-parasitised lambs received some larval challenge, both of which would have reduced the contrast between treatments. The suppressive
Ž
effect of parasites on intake in the calf is rapidly alleviated by treatment Fox et al.,
. Ž
1989 . Secondly, intake recovers as lambs develop immunity Steel et al., 1980, Steel et
. Ž .
al., 1982 , and Kyriazakis et al. 1996 have shown that responses of lambs to parasitism change with time in longer-term studies. Intake and selection were measured when these 8-month-old lambs had been on the trial for 12 weeks, possibly beyond the period when parasites have their most severe effect. Faecal egg counts were lower at the end of the measurement period than they were 4 weeks previously. However, the intake measure-ment period was timed to coincide with the rise towards the trigger level, and egg counts
Ž .
were still similar to the levels reported by Kyriazakis et al. 1994 , in a trial where Ž .
intake was suppressed. Kyriazakis et al. 1998 proposed several hypotheses to explain anorexia in parasite-challenged animals. One is that anorexia leads to increased diet selectivity. While our data does not show anorexia, for reasons discussed above, our results are consistent with this hypothesis of a parasite challenge providing both the incentive and opportunity to enhance nutritional status. Further, selecting a diet with higher nutrient density may minimise re-infection by reducing larval intake per unit of food intake.
The nitrogen concentration in the two species of grass was similar to that normally recorded for clover, although the greater total dry matter intake of lambs grazing RWC gave them a higher level of nitrogen intake. White clover consistently has high nitrogen
Ž . Ž .
concentration 4–6% in its leaves Ulyatt et al., 1988 , and there is evidence that it is
Ž .
internally regulated by the nitrogen fixation symbiosis Chapman and Parsons, 1996 . White clover herbage taken from a nearby site in the same year had 4.8% nitrogen Žunpubl. data , and we consider it is reasonable to assume that grass and clover were. similar in nitrogen concentration in this experiment. For the RWC treatment, parasitised lambs did not increase selection for clover compared with non-parasitised, perhaps because they were already at a high nutritional status with a diet of 50% clover and 50% grass, and a high daily intake. This ratio of grass and clover may be an optimum during autumn, even for healthy ruminants. Cattle offered a choice of adjacent monocultures of ryegrass and white clover also selected a diet containing approximately 50% white
Ž .
clover and 50% ryegrass Cosgrove et al., 1996 , when they could have easily obtained a higher or a lower proportion of clover. They selected a higher proportion of clover Ž67% in an earlier phase of the experiment, conducted in the summer, similar to that.
Ž .
recorded for sheep in summer Parsons et al., 1994a . The intake and liveweight gain of Ž
RWC lambs was high compared with other studies Bown et al., 1991; van Houtert et .
Ž y1 with indoor studies offering prepared feeds e.g., 90 vs. 214 g crude protein kg DM
.
used by Kyriazakis et al., 1994 , where the low-protein feed was probably deficient,
Ž .
even for healthy lambs National Research Council, 1985 . A small difference in nitrogen concentration between feed choices would reduce the value of selective grazing for lambs on RWC swards. This was not the case for lambs grazing BWC. Their protein status was lower because of lower intake, and they had a lower proportion of white clover in their diet, compared with RWC lambs. However, at this lower status, parasitised lambs selected more clover, which is consistent with the hypothesis that nutritional factors increase parasitised lambs’ selection for white clover. Lambs may select food based on characteristics of protein other than the total amount in the feed. The amount and rate at which protein reaches the lower gastro-intestinal tract is higher
Ž .
for clover than for grass Ulyatt et al., 1988 . Additional protein supplied to the lower Ž
gastro-intestinal tract either as a continuous infusion via abomasal catheter Bown et al.,
. Ž .
1991 , or as ruminal-protected fish-meal protein in the diet van Houtert et al., 1995 , will reduce the debilitating effects of parasites. Other, non-nutritional characteristics also
Ž .
differ between grass and clover, such as rate of intake Penning et al., 1991b . Healthy sheep offered a choice between grass and clover growing as adjacent monocultures, did not alter their preference for clover in response to differences in the nitrogen
concentra-Ž .
tion of the grass Harvey et al., 1996 . Thus, factors other than just the concentration of nitrogen evidently are involved in preference between grass and clover.
Parasitised and non-parasitised lambs grazed together so that all lambs were subjected to a consistent level of natural, larval challenge, and were offered pasture swards with similar horizontal and vertical structure and composition. It is not possible to maintain the same larval challenge for animals of contrasting parasite status grazing separately,
Ž .
without resorting to artificial techniques e.g., dosing with larvae . There is a possibility of confounding in this experimental design, from interactions between treatment groups. However, while facilitation may affect time spent grazing and therefore daily intake ŽRook and Penning, 1991 , there is no evidence that diet selection is similarly influ-. enced. For the experimental design we used, such an effect on daily intake would reduce, rather than overstate the magnitude of difference between anthelmintic treat-ments.
4.2. Effect of grass species on selection and intake
( ) G.P. CosgroÕe, J.H. NiezenrApplied Animal BehaÕiour Science 66 2000 71–85
82
times higher than the initial proportion in the sward. However, they achieved a higher proportion in their diet on RWC, where the proportion in the sward was higher than on
Ž
BWC. The herbage mass of white clover i.e., the proportion of clover in the sward .
multiplied by the total herbage mass for that sward was similar for both treatments at the start of the experiment, but was lower for RWC than for BWC at the end. This greater depletion of white clover from RWC is consistent with estimates of both the higher proportion of clover in the diet and the higher intake of clover. However, that the lambs ate more clover from RWC than from BWC indicates that proportion, and not the
Ž .
total availability herbage mass , was the critical parameter affecting diet composition for these continuously stocked swards. Under continuous stocking, daily herbage
Ž y1 y1.
allowance kg DM hd day was high, which on mixed-species pasture gave lambs the maximum scope to select their preferred diet.
Ž
Selection is driven in part by preference, but modified by circumstance Hodgson, .
1979 . Our results demonstrate enhanced selection for clover in response to the nutritional consequences of parasitism, when animals are grazing on typical mixed-species pastures. Because of the complexities in this foraging environment, it is not possible to deduce the effect of parasitism on preference per se, which would provide the strongest test of our hypothesis. Although we controlled for overall sward surface
Ž .
height, which has a large influence on daily intake Penning et al., 1991a , selection is influenced by many other factors. The vertical and horizontal abundance of each plant
Ž .
species Parsons et al., 1994b; Edwards et al., 1996b , and the scale of aggregation of
Ž .
the species Edwards et al., 1996a , strongly influence selection. For a mixed grass– clover sward, the nitrogen concentration of the grass is affected by clover abundance ŽSchwinning and Parsons, 1996 and cannot be independently controlled. Moreover,. when animals are grazing a mixed pasture, diet composition is not easily measured. For the alkane technique we used, there are possible errors from inaccurately representing the morphological components of the diet by hand plucking, even for a relatively simple two-species sward. To explore further the links between selection and nutritional state of parasitised animals, techniques where preference per se is clearly demonstrated, such as separate, adjacent monocultures of grass and clover which provide animals free access to
Ž .
either species Parsons et al., 1994a may offer advantages. This approach removes the physical constraints which influence selection by making it possible to control vertical and horizontal abundance, avoids problems associated with heterogeneity and scale of species aggregation, and simplifies the measurement of diet composition. Finally, these results highlight the need to consider the impact of intestinal parasites, both in the animal and on the pasture, in the design, conduct and interpretation of field studies on preference and diet selection, not just those investigating the effects of parasitism.
5. Conclusions
even outdoors where selection is more complex, where the contrast in nutrient concen-tration between dietary alternatives is lower, and where non-equal availability of diet alternatives may force a trade-off between selection for a preferred item and rate of intake of that item. There are practical implications to these results that warrant further investigation. At a time of increasing resistance of nematodes to broad-spectrum anthelmintics, and of increasing consumer resistance to the reliance on chemicals, it is essential to explore opportunities for further experimental testing and application of this novel result. Increasing the proportion of white clover in the pasture, and making it easier for lambs to select their desired proportion of white clover in the diet, are both strategies that might ameliorate the effects of parasitism in the short-term. However, increased selection for white clover depletes its proportion in the sward and is ultimately
Ž counter-productive for the animals gaining a high clover proportion in their diet Parsons
.
et al., 1991 . This indicates the need for different management strategies in the longer-term to allow lambs to enhance dietary clover proportion in response to nutri-tional demands caused by parasitism.
Acknowledgements
We thank Heather Robertson for assistance with collection of animal and pasture data, and staff of the Herbage Laboratory for botanical identification and separation. We acknowledge financial support from Meat NZ and Wools of NZ.
References
Anderson, F.L., Walters, G.T., 1973. Efficacy of the Baermann technique for recovery of Dictyocaulus
ÕiÕiparus larvae from bovine feces. Am. J. Vet. Res. 34, 39–40.
Bircham, J., 1981. Herbage growth and utilisation under continuous stocking management. PhD Thesis, Univ. of Edinburgh.
Bown, M.D., Poppi, D.P., Sykes, A.R., 1991. The effect of post-ruminal infusion of protein or energy on the pathophysiology of Trichostrongylus colubriformis infection and body composition in lambs. Aust. J. Agric. Res. 42, 253–267.
Chapman, D.F., Parsons, A.J., 1996. Management of clover in grazed pastures: expectations, limitations and opportunities. In: White Clover: New Zealand’s Competitive Edge. Agron. Soc. NZ Special Publication No. 11, pp. 55–64.
Cosgrove, G.P., Anderson, C.B., Fletcher, R.H., 1996. Do cattle exhibit a preference for white clover? In: White Clover: New Zealand’s Competitive Edge. Agron. Soc. NZ Special Publication No. 11, pp. 83–86. Cowie, J.D., Fitzgerald, P., Owers, W., 1967. Soils of the Manawatu–Rangitikei Sand Country. New Zealand
Soil Bur. Bull. 29. New Zealand Department of Scientific and Industrial Research, p. 40.
Dove, H., Moore, A.D., 1995. Using a least squares optimization procedure to estimate botanical composition based on the alkanes of plant cuticular wax. Aust. J. Agric. Res. 46, 1535–1544.
Edwards, G.R., Newman, J.A., Parsons, A.J., Krebs, J.R., 1996a. Effects of the scale and spatial distribution of the food resource and animal state on diet selection: an example with sheep. J. Anim. Ecol. 63, 816–826. Edwards, G.R., Newman, J.A., Parsons, A.J., Krebs, J.R., 1996b. Effects of the total, vertical and horizontal
( ) G.P. CosgroÕe, J.H. NiezenrApplied Animal BehaÕiour Science 66 2000 71–85
84
Fox, M.T., Gerrelli, D., Pitt, S.R., Jacobs, D.E., Gill, M., Gale, D.L., 1989. Ostertagia ostertagi infection in the calf.: effects of a trickle challenge on appetite, digestibility, rate of passage of digesta and liveweight gain. Res. Vet. Sci. 47, 294–298.
Harris, W., 1973. Why browntop is bent on creeping. Proc. NZ Grssld. Ass. 35, 101–109.
Harvey, A., Orr, R.J., Parsons, A.J., Penning, P.D., Vincent, J.F.V., 1996. The effects of grass nitrogen status on the preference by sheep grazing ryegrass and white clover. Br. Grssl. Soc. Occas. Symp. 30, 227–229. Hodgson, J., 1979. Nomenclature and definitions in grazing studies. Grass Forage Sci. 34, 11–18.
Hou, X.Z., Emmans, G.C., Anderson, D.H., Illius, A.W., Oldham, J.D., 1991. The effect of different pairs of feeds offered as a choice on food selection by sheep. Proc. Nutr. Soc. 50, 94A.
Hutchings, M.R., Kyriazakis, I., Anderson, D.H., Gordon, I.J., Coop, R.L., 1998. Behavioural strategies used by parasitized and non-parasitized sheep to avoid ingestion of gastro-intestinal nematodes associated with faeces. Anim. Sci. 67, 97–106.
Kyriazakis, I., Anderson, D.H., Oldham, J.D., Coop, R.L., Jackson, F., 1996. Long-term subclinical infection with Trichostrongylus colubriformis: effects on food intake, diet selection and performance of growing lambs. Vet. Parasitol. 61, 297–313.
Kyriazakis, I., Oldham, J.D., Coop, R.L., Jackson, F., 1994. The effect of subclinical intestinal nematode infection on the diet selection of growing sheep. Br. J. Nutr. 72, 665–677.
Kyriazakis, I., Oldham, J.D., 1993. Diet selection in sheep: the ability of growing lambs to select a diet that
Ž .
meets their crude protein nitrogen=6.25 requirements. Br. J. Nutr. 69, 617–629.
Kyriazakis, I., Tolkamp, B.J., Hutchings, M.R., 1998. Towards a functional explanation for the occurrence of anorexia during parasitic infections. Anim. Behav. 56, 265–274.
Mott, G.O., Lucas, H.L., 1952. The design, conduct, and interpretation of grazing trials on cultivated and improved pastures. Proc. 6th Int. Grssld. Congr. pp. 1380–1385.
National Research Council, 1985. Nutrient requirements of sheep. Sixth revised edn.. National Academy Press, Washington, DC.
Niezen, J.H., Charleston, W.A.G., Hodgson, J., Miller, C.M., Waghorn, T.S., Robertson, H.A., 1998. Effect of plant species on the larvae of gastrointestinal nematodes which parasitise sheep. Int. J. Parasitol. 28, 791–803.
Parsons, A.J., Harvey, A., Johnson, I.R., 1991. Plant–animal interactions in a continuously grazed mixture: II. The role of differences in the physiology of plant growth and of selective grazing on the performance and stability of species in a mixture. J. Appl. Ecol. 28, 635–658.
Parsons, A.J., Newman, J.A., Penning, P.D., Harvey, A., Orr, R.J., 1994a. Diet preference of sheep: effects of recent diet, physiological state and species abundance. J. Anim. Ecol. 63, 465–478.
Parsons, A.J., Thornley, J.H.M., Newman, J., Penning, P.D., 1994b. A mechanistic model of some physical determinants of intake rate and diet selection in a two-species temperate grassland sward. Func. Ecol. 8, 187–204.
Penning, P.D., Parsons, A.J., Orr, R.J., Treacher, T.T., 1991a. Intake and behaviour responses by sheep to changes in sward characteristics under continuous stocking. Grass Forage Sci. 46, 15–28.
Penning, P.D., Rook, A.J., Orr, R.J., 1991b. Patterns of ingestive behaviour of sheep continuously stocked on monocultures of ryegrass or white clover. Appl. Anim. Behav. Sci. 31, 237–250.
Poppi, D.P., MacRae, J.C., Brewer, A.C., Coop, R.L., 1986. Nitrogen transaction in the digestive tract of lambs exposed to the intestinal parasite Trichostrongylus colubriformis. Br. J. Nutr. 55, 593–602. Poppi, D.P., Sykes, A.R., Dynes, R.A., 1990. The effect of endoparasitism on host nutrition — the
implications for nutrient manipulation. Proc. NZ Soc. Anim. Prod. 50, 237–243.
Rook, A.J., Penning, P.D., 1991. Synchronisation of eating, ruminating and idling activity by grazing sheep. Appl. Anim. Behav. Sci. 32, 157–166.
Schwinning, S., Parsons, A.J., 1996. Analysis of the coexistence mechanisms for grasses and legumes in grazing systems. J. Ecol. 84, 799–813.
Steel, J.W., Symons, L.E.A., Jones, W.O., 1980. Effect of level of larval intake on the productivity and physiology and metabolic response of lambs infected with Trichostrongylus colubriformis. Aust. J. Agric. Res. 31, 821–828.
Steel, J.W., Jones, W.O., Symons, L.E.A., 1982. Effects of a concurrent infection of with Trichostrongylus
Taylor, E.L., 1939. Technique for the estimation of pasture infestation by strongyloid larvae. Parasitology 31, 473–478.
Thornley, J.H.M., Parsons, A.J., Newman, J.A., Penning, P.D., 1994. A cost–benefit model of grazing intake and diet selection in a two-species temperate grassland sward. Func. Ecol. 8, 5–16.
Ulyatt, M.J., Thomson, D.J., Beever, D.E., Evans, R.T., Haines, M.J., 1988. The digestion of perennial
Ž . Ž .
ryegrass Lolium perenne L. cv. Melle and white clover Trifolium repens L. cv. Blanca by grazing
cattle. Br. J. Nutr. 60, 137–149.
Van Houtert, M.J.F., Barger, I.A., Steel, R.G., Windon, R.G., Emery, D.L., 1995. Effects of dietary protein intake on responses of young sheep to infection with Trichostrongylus colubriformis. Vet. Parasitol. 56, 163–180.