252 (2000) 15–26
www.elsevier.nl / locate / jembe
Settlement in different-sized patches by the gregarious
intertidal barnacle Chamaesipho tasmanica Foster and
Anderson in New South Wales
* Carolyn J. Jeffery
Centre for Research on Ecological Impacts of Coastal Cities and Institute of Marine Ecology, Marine Ecology Laboratories, A11, University of Sydney, Sydney, NSW 2006, Australia
Received 26 November 1999; received in revised form 3 May 2000; accepted 8 May 2000
Abstract
Most models on settlement of open marine invertebrate populations are based on space-limitation. These models, however, do not recognise that free space may not drive the demography of populations when larval numbers are small or when larval supply varies along a gradient in the habitat. They also do not incorporate the effects of larval choice when settling. It has been hypothesised that, in gregarious barnacles, the effects of adult conspecifics, rather than available free space, may play a primary role in settlement. That is, cues from adults along perimeters of patches, rather than space available, may enhance colonisation. This study therefore aimed to distinguish between these separate influences on populations of Chamaesipho tasmanica, a gregarious barnacle characterised by relatively few larvae arriving to settle each year. Patches of 6, 3 and 1.5 cm diameter were cleared within aggregations of barnacles at three heights (Low, Mid, Upper) of Chamaesipho’s distribution at two sites and during 2 years of settlement. Total numbers of settlers in each year were manipulated to determine the separate influences on settlement due to availability of substratum or the effects of conspecific adults. To test for the effects of available free space, numbers of settlers per unit area were analysed. To test for gregarious effects due to the presence of adults, numbers of settlers per unit perimeter were analysed. While available substratum was found not to affect settlement of this barnacle, gregarious settlement in response to adults at perimeters of patches was thought to be confounded by differential larval supply and differential conspecific cues among heights on the shore. Results from this study therefore have important implications for survival of gregarious populations following disturbances, especially in species where larval supply is poor. Crown copyright 2000 Published by Elsevier Science B.V. All rights reserved.
Keywords: Available space; Barnacles; Conspecific adults; Gregarious settlement; Larval supply; Perimeter
*Centre for Marine and Coastal Studies, School of Biological Sciences, University of New South Wales, Sydney, NSW 2052, Australia. Tel.: 161-2-9385-1825; fax:161-2-9662-7995.
E-mail address: [email protected] (C.J. Jeffery)
0022-0981 / 00 / $ – see front matter Crown copyright 2000 Published by Elsevier Science B.V. All rights reserved.
1. Introduction
Larval supply, which is influenced by hydrodynamics, has been shown to determine temporal variability within barnacle populations (Farrell et al., 1991; Roughgarden et al., 1991, 1994; Pineda, 1994a,b), whereas spatial variability has been linked to larval behavioural responses and availability of substratum during settlement (Gaines and Roughgarden, 1985; Bushek, 1988; Minchinton and Scheibling, 1993; Pineda, 1994a). Most models on settlement of open marine populations emphasise free space as the main determinant of demography (Roughgarden et al., 1985, 1988; Roughgarden and Iwasa, 1986; Possingham and Roughgarden, 1990), but they do not recognise that larval behaviour may also influence settlement.
In fact, many previous studies on barnacles have emphasised that amount of free space available determines numbers of larvae that settle. In separate experiments on Semibalanus and Chthamalus spp., Bertness et al. (1992) and Pineda (1994a), respectively, found that patch size determined numbers of barnacles settling; smaller patches had greater settlement. Alternatively, Gaines and Roughgarden (1985) and Minchinton and Scheibling (1993) showed that there was more settlement with increased availability of substratum for S. balanoides and Balanus glandula, respectively. Pineda and Caswell (1997) further suggested that differences in availability of substratum may be responsible for variations in settlement at different times, so that there would be an intensification of settlement when suitable substratum became scarce.
While this may be true for species with large numbers of larvae available and limited free space on the substratum, it is not known whether similar processes operate in species that have abundant free space and relatively few larvae. In fact, other site-specific characteristics, such as local hydrodynamics, may also influence larval supply and determine spatial variability in species where larvae are sparsely distributed and where there is much space available for settlement. Larval behaviour when settling (Raimondi, 1991) may also be important, especially when species are gregarious. Hence, the presence of conspecific adults may define the numbers of gregarious larvae settling and will depend on the degree of exposure to adults at the perimeter of patches (Minchinton, 1997). Abundance of conspecifics may also play a primary role. This is contrary to the previously expressed models of free space where variability in recruitment is said to be driven by the amount of substratum available.
response to more concentrated conspecific cues (Minchinton, 1997). It may be, however, that reduced area of patch will intensify settlement (Pineda and Caswell, 1997) despite other influences.
At Cape Banks, Botany Bay, New South Wales, Australia, contrary to previously documented cases, few larvae of the small gregarious barnacle Chamaesipho tasmanica arrive and settle relative to the space available (Jeffery, 1997). The barnacles often form continuous sheets of joined individuals or occur in isolated groups of joined or separate individuals on mid-littoral areas. In the more wave-exposed areas at Cape Banks, a patchy distribution is prevalent whereas, in sheltered areas, cleared spaces may be scattered throughout aggregations of barnacles. Despite the amount of space differing from place to place, there were consistent patterns in numbers of cyprids arriving from 1990 to 1993. Although the numbers of cyprids varied among places, they were correlated with the distributions of juveniles on the substratum (Jeffery, 1997; Jeffery and Underwood, 2000). Obviously, the suggestions of Pineda and Caswell (1997) that consistent differences in rate of settlement might be explained by the proportion of the substratum available, cannot be substantiated by Chamaesipho. Other processes must also be operating.
For example, because Chamaesipho settle gregariously in response to the presence of adults (Otaiza, 1989; Jeffery, 1997), if different-sized patches were cleared within aggregations of barnacles, it was hypothesised that the degree of exposure to adults along perimeters of patches, rather than the amount of space available, would influence settlement of cyprids. It was therefore predicted that in plots of 6, 3 or 1.5 cm diameter cleared daily of newly-settled barnacles, there would be proportionally greatest settlement in the smallest plots with the greatest relative exposure to adults along perimeters. This study therefore tested the hypotheses that increased availability of substratum and / or exposure to conspecific adults would influence settlement of Chamaesipho.
If free space contributed to numbers settling, it would be expected that there would be no differences in numbers settling per unit area in patches of different sizes. Alter-natively, if adults around the perimeter of a patch influence settlement, there should be no differences in settlement per unit length of perimeter around patches of different sizes.
2. Materials and methods
2.1. Study area
(Short and Trenaman, 1992), which contribute to conditions of variable exposure. Tides are semi-diurnal and range over 2 m.
2.2. Influence of substrate availability and conspecific adults on gregarious settlement
In July 1991 and 1992, 6, 3 and 1.5 cm diameter patches (n53) were cleared on the substratum within aggregations of barnacles at Sites 3 and 4 at each Low, Mid and Upper height of Chamaesipho’s distribution. From July to December each year, in 1991 and 1992, settlers were removed daily from each different-sized patch for approximately 6 days during each new and full moon. Very few, if any, barnacles settled during the intervening period (Jeffery, 1997). Daily removal was necessary to negate possible confounding by gregariousness as neighbouring settlers aged (Jeffery, 1997), and to ensure that only free space and conspecific adults on perimeters could act as cues for settlement. Numbers of barnacles settling in these patches were recorded until December of each year of the experiment.
Total numbers of settlers in the patches in each experiment in 1991 and 1992 were compared. Because different-sized patches were used in this investigation it was necessary to separate the influences of the availability of substratum and numbers of adults along perimeters of patches. Numbers of larvae settling on different-sized patches were manipulated in two ways. To test the hypothesis that amount of available free space will influence settlement, the total numbers of settlers recorded each year on plots where larvae were removed daily were divided by the appropriate areas (28.28, 7.07 and
2
1.77 cm ). To test the hypothesis that larval choice will influence settlement near conspecific adults, the numbers of barnacles settling were divided by the appropriate perimeters (18.85, 9.43 and 4.71 cm).
3. Results
3.1. Total numbers of settlers
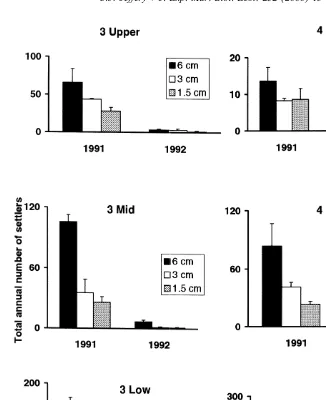
Where settlers were removed from patches from July to December 1991 and 1992, total numbers of settlers were always greater in 6 cm patches than in 3 cm patches, which had more than 1.5 cm patches (Fig. 1). In 1991 there were generally more settlers at Site 3 than Site 4. More settlers were always recorded at Low heights and there was a decrease in numbers of settlers as patch size decreased (6 cm.3 cm.1.5 cm; Fig. 1; Table 1a).
3.2. Availability of substratum
Fig. 1. Mean total annual numbers of barnacles settling in patches (n53) of different size (6, 3 and 1.5 cm) at Sites 3 and 4 on Low, Mid and Upper heights of Chamaesipho’s distribution in December, 1991 and 1992. Note different scale on y-axis among sites and heights.
Table 1
Analysis of total annual numbers of barnacles settling in 6, 3 and 1.5 cm patches on which settlers were removed daily (n53) at Sites 3 and 4 on Low, Mid and Upper heights of Chamaesipho’s distribution. Experiments were renewed each year and data are totals in December in 1991 and 1992: (a) total numbers of settlers in different-sized patches; (b) total numbers of settlers / unit area of patch (availability of substratum);
a (c) total numbers of settlers / unit perimeter of patch (adults at perimeters)
Source df MS F P
(a) Total numbers of settlers (analysis ofvariance, 0.5
data transformed (x11) ; Cochran’s test, P.0.05)
Time: T 1 631.96 188.65 *
Site: S 1 9.83 7.42 **
Height: H 2 115.80 15.99 ns
Patch size: P 2 61.05 118.79 **
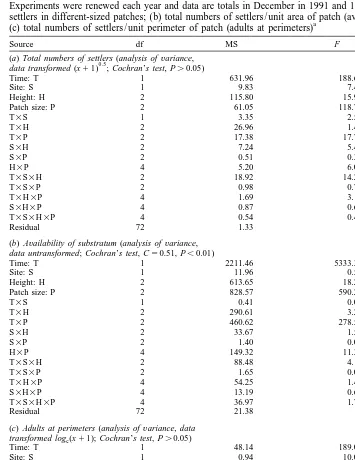
(b) Availability of substratum (analysis ofvariance, data untransformed; Cochran’s test, C50.51, P,0.01)
Time: T 1 2211.46 5333.31 **
Site: S 1 11.96 0.56 ns
Height: H 2 613.65 18.23 ns
Patch size: P 2 828.57 590.25 **
T3S 1 0.41 0.02 ns
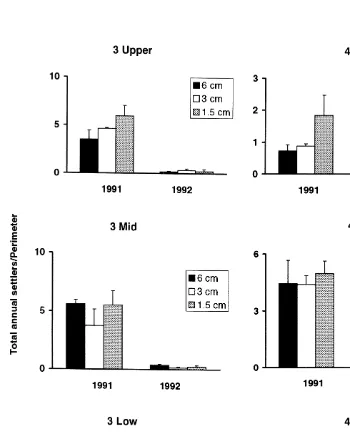
(c) Adults at perimeters (analysis ofvariance, data transformed log (xe 11); Cochran’s test, P.0.05)
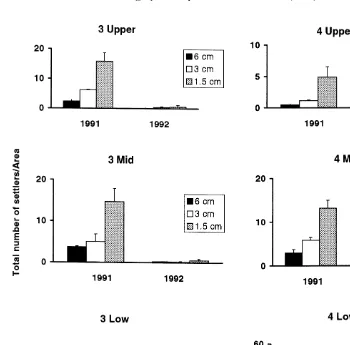
Fig. 2. Mean total annual numbers of settlers / area of patch (n53) of different size (6, 3 and 1.5 cm) at Sites 3 and 4 on Low, Mid and Upper heights of Chamaesipho’s distribution in December, 1991 and 1992. Note different scale on y-axis among sites and heights.
increased availability of free space does not control numbers of settling Chamaesipho cyprids and that larval choice is operating.
3.3. Influence of conspecific adults at perimeters on gregarious settlement
similar numbers of barnacles settled in the larger patches (1.5 cm.3 cm56 cm; Fig. 3; Table 1c). These results suggest that, while bordering adults provide cues for settlement, other processes are also operating.
4. Discussion
This study tested the hypotheses that increased availability of substratum and / or exposure to conspecific adults will influence settlement in the gregarious barnacle C. tasmanica. While it was initially found that most larvae settled in the largest patches, when total numbers of barnacles settling each year were divided by unit area (total settlers / area of patch) to standardise the effects of differential availability of free space, greatest numbers of settlers were recorded in the smallest patches. Some differences in settlement among heights on the shore for the different-sized patches were also recorded. While there was no difference in settlement among heights in 6 cm patches, in the smaller 1.5 and 3 cm patches most settlement occurred at Low heights where resident adults and larvae settling next to them were most dense. Also, on Upper heights, where barnacles were sparsely distributed, settlement was similar for all different-sized patches. Conversely, on Low and Mid heights, where adults were more dense, most settlement occurred in the smallest patches. In addition, greatest numbers of cyprid larvae arrive at Low heights of Chamaesipho’s range of distribution at Cape Banks than at Mid / Upper heights (Jeffery, 1997; Jeffery and Underwood, 2000). Results therefore suggest that, instead of available substratum, differential larval supply and differential conspecific cues from adults among heights may interact to influence settlement. In fact, free space may not be important because Chamaesipho larvae generally arrive at Cape Banks in small numbers, except for a few pulses of large numbers each year (Jeffery, 1997; Jeffery and Underwood, 2000). The poor larval supply and gregarious behaviour when settling contribute to numbers that settle. That is, Chamaesipho cyprids do not act as passive particles. Larval behaviour, rather than available free space, dominates settle-ment in gregarious barnacles when relatively few larvae arrive. It may therefore be that, on Low heights, the 6 cm patches were too big to have been filled by larvae whenever major pulses occurred. Conversely, because barnacles are more sparsely distributed on Mid and Upper shores, there may have been fewer conspecific cues from adults around perimeters to initiate settlement. Results therefore suggest that large amounts of space within aggregations of barnacles do not enhance settlement in species with few larvae arriving or in gregarious species where conspecific cues play a primary role in initiating settlement.
Results from this study therefore have implications for survival of populations of Chamaesipho because they are gregarious and adults provide cues for settlement: fewer cyprids would settle in large areas of bare space typical of disturbed areas. In fact, from 1990 to 1993 the total number of Chamaesipho cyprids available for settlement declined at Cape Banks (Jeffery, 1997; Jeffery and Underwood, 2000) and, in 1994, very few juveniles were observed on the substratum. There was also an overall decrease in numbers of adults recorded at all sites and heights at Cape Banks from 1989 to 1993 because the numbers of new barnacles that recruited each year were not enough to balance the numbers lost (Jeffery, 1997). It would therefore be expected that, because fewer adults were present, fewer larval barnacles would settle in successive years. Raimondi and Keough (1990), however, have documented larval variability in settle-ment by the Californian barnacle C. anisopoma which has a similar spatial distribution to that of Chamaesipho. They found that while 95% of the population was gregarious and settled close to already established adults, the remaining 5% settled independently. Solitary larvae and adults of Chamaesipho were also observed during this study at Cape Banks. It may be that larval variability at settlement maintains the status quo of Chamaesipho in unfavourable conditions because a small proportion of Chamaesipho shows this non-gregarious behaviour. Independent settling within gregarious populations of barnacles would have a selective advantage if mass mortality of adults occurred. It could ensure survival of populations which would otherwise be exterminated during intense disturbances. Recovery of populations of Chamaesipho after disturbance would still be expected to be slow because most larvae settle gregariously.
In summary, settlement in Chamaesipho was shown to be influenced by conspecifics at the edge of patches, not by the amount of habitat available for settlement. There were, however, some conflicting results which reflected the variation in spatial distributions of larvae and adults. In his study on the intertidal tubeworm Galeolaria caespitosa, Minchinton (1997) also showed that conspecific adults induced settlement of gregarious larvae, but his results were more clearcut. This is because he cleared plots within continuous aggregations of adults, whereas, in this study, although different-sized patches were also cleared within aggregations, adult barnacles were more sparsely distributed on Upper heights of Chamaesipho’s distribution. Also, while larval supply of Galeolaria is unknown (O’Donnell, 1986), larval supply of Chamaesipho is poor with great temporal and spatial variation (Jeffery, 1997; Jeffery and Underwood, 2000). Finally, while space-limited open populations may be driven by differences in availability of suitable substrate (Pineda and Caswell, 1997), results from this study on the gregarious barnacle Chamaesipho implicate larval supply and larval choice. Future models on settlement of other gregarious species should therefore also incorporate the effects of differential larval supply and differential conspecific cues on settlement.
Acknowledgements
to my supervisor, Tony Underwood, for his helpful comments on drafts of this manuscript, and for his support throughout this study. [AJ]
References
Bertness, M.D., Gaines, S.D., Stephens, F.G., Yund, P.O., 1992. Components of recruitment in populations of the acorn barnacle Semibalanus balanoides (Linnaeus). J. Exp. Mar. Biol. Ecol. 156, 199–215. Bushek, D., 1988. Settlement as a major determinant of intertidal oyster and barnacle distributions along a
horizontal gradient. J. Exp. Mar. Biol. Ecol. 122, 1–18.
Farrell, T.M., Bracher, D., Roughgarden, J., 1991. Cross-shelf transport causes recruitment to intertidal populations in central California. Limnol. Oceanogr. 36, 279–288.
Gaines, S., Roughgarden, J., 1985. Larval settlement rate: a leading determinant of structure in an ecological community of the marine intertidal zone. Proc. Natl. Acad. Sci. 82, 3707–3711.
Jeffery, C.J., 1997. The ecology of the rocky shore intertidal barnacle Chamaesipho tasmanica in New South Wales. Ph.D. thesis, University of Sydney.
Jeffery, C.J., Underwood, A.J., 2000. Consistent spatial patterns of arrival of larvae of the honeycomb barnacle Chamaesipho tasmanica Foster and Anderson in New South Wales. J. Exp. Mar. Biol. Ecol., in press. Le Tourneaux, F., Bourget, E., 1988. Importance of physical and biological settlement cues used at different
spatial scales by the larvae of Semibalanus balanoides. Mar. Biol. 97, 57–66.
Minchinton, T.E., 1997. Life on the edge: conspecific attraction and recruitment of populations to disturbed habitats. Oecologia 111, 45–52.
Minchinton, T.E., Scheibling, R.E., 1993. Free space availability and larval substratum selection as determinants of barnacle population structure in a developing rocky intertidal community. Mar. Ecol. Prog. Ser. 95, 233–244.
O’Donnell, M.A., 1986. The ecology and early life history of Galeolaria caespitosa. Ph.D. thesis, University of Sydney.
Otaiza, R.D., 1989. Arrival and recruitment of intertidal barnacles on rocky shores in central New South Wales, Australia. Ph.D. thesis, University of Sydney.
Pineda, J., 1994a. Spatial and temporal patterns in barnacle settlement rate along a southern California rocky shore. Mar. Ecol. Prog. Ser. 107, 125–138.
Pineda, J., 1994b. Internal tidal bores in the nearshore: warm-water fronts, seaward gravity currents and the onshore transport of neustonic larvae. J. Mar. Res. 52, 427–458.
Pineda, J., Caswell, H., 1997. Dependence of settlement rate on suitable substrate area. Mar. Biol. 129, 541–548.
Possingham, H.P., Roughgarden, J., 1990. Spatial population dynamics of a marine organism with a complex life cycle. Ecology 71, 973–985.
Raimondi, P.T., 1990. Patterns, mechanisms, consequences of variability in settlement and recruitment of an intertidal barnacle. Ecol. Monogr. 60, 283–309.
Raimondi, P.T., 1991. Settlement behavior of Chthamalus anisopoma larvae largely determines the adult distribution. Oecologia 85, 349–360.
Raimondi, P.T., Keough, M.J., 1990. Behavioural variability in marine larvae. Aust. J. Ecol. 15, 427–437. Roughgarden, J., Iwasa, Y., 1986. Dynamics of a metapopulation with space-limited subpopulations. Theor.
Popul. Biol. 29, 235–261.
Roughgarden, J., Iwasa, Y., Baxter, C., 1985. Demographic theory for an open marine population with space-limited recruitment. Ecology 66, 54–67.
Roughgarden, J., Gaines, S., Possingham, H., 1988. Recruitment dynamics in complex life cycles. Science 241, 1460–1466.
Roughgarden, J., Pennington, J.T., Alexander, S., 1994. Dynamics of the rocky intertidal zone with remarks on generalization in ecology. Trans. R. Soc. London B 343, 79–85.
Short, A.D., Trenaman, N.L., 1992. Wave climate of the Sydney region, an energetic and highly variable ocean wave regime. Aust. J. Mar. Freshwater Res. 43, 765–791.