KEBERGANTUNGAN MANGGIS (Garcinia mangostana L.) TERHADAP
CENDAWAN MIKORIZA ARBUSKULAR DAN PUPUK FOSFAT
DI LAPANG
(Mycorrhizal dependency of Mangosteen under field to Applicated Phosphate Fertilizer)
Auzar Syarif
ABSTRACT
The field experiment was conducted to evaluate the mycorrhizal dependency of mangosteen to applicated phosphate fertilizer from Januari 2000 until April 2001 in the experimental field of Andalas University Padang. The experiment was to evaluate two arbuscular mycorrhizal fungi species (Glomus etunicatum and Glomus manihotis, and control) and four dosages of phosphate fertilizer (0, 150, 300, and 450 mg plant-1 month-1). The experimental
design were Randomized Blocks Design in factorial pattern with three replications. The results of the experiments showed that (1) the maximum infection levels of Glomus etunicatum, Glomus manihotis, and control on roots of 16 months mangosteen were 68.53, 64.34, and 57.99 percent at the optimum dosages of phosphate fertilizer 70.26, 288.18, and 265.49 mg plant-1 month-1 (2)
total dry weight of mangosteen inoculated with Glomus etunicatum, Glomus manihotis, and controlwere 9.13, 8.58, and 7.64 g plant-1 at the optimum dosages of 243.02,
421.99, and 863.82 mg plant-1 month-1 phosphorus
fertilizer respectively (3) Mycorrhizal dependency of mangosteen under field was highest at the dosage on 150 mg P plant-1 month-1 for Glomus etunicatum and 450 mg P
plant-1 month-1 for Glomus manihotis.
Key words: mangosteen, mycorrhizal, phosphate
PENDAHULUAN
Kebergantungan tanaman terhadap CMA tinggi pada tanaman yang mempunyai sistem perakaran terbatas. Pada tanaman seperti itu, CMA efektif, memperluas daerah jelajah akar dan meningkat-kan pertumbuhan akar serta membebasmeningkat-kan hara terikat menjadi tersedia bagi tanaman. Hasil pe-nelitian Syarif (2002) menunjukkan bahwa keber-gantungan tanaman manggis pada stadia bibit
terhadap inokulasi berbagai jenis CMA (Glomus
etunicatum dan Glomus manihotis) tergolong ren-dah. Kenyataan seperti itu dapat terjadi karena tingkat kebergantungan manggis pada stadia bibit terhadap CMA masih rendah karena selama pe-riode perkecambahan dan pertumbuhan semai, kebergantungan tanaman akan hara lebih dominan ditentukan oleh penyediaan makanan cadangan yang terdapat dalam biji.
Perbedaan kebergantungan tanaman terhadap CMA sebagai akibat dari perbedaan kemampuan CMA meningkatkan serapan hara yang diperlu-kan untuk kebutuhan tanaman. Keberadaan CMA tidak hanya untuk membantu atau memfasilitasi meningkatkan serapan hara, tetapi keberadaannya juga dapat meningkatkan panjang akar tanaman. Menurut Setiadi (2000), respons tanaman terha-dap CMA ditentukan oleh tingkat kebergantung-an tkebergantung-anamkebergantung-an terhadap CMA (mycorrhizal depen-dency) dan tingkat kebergantungan itu ditentukan pula oleh jenis CMA. Konsep kebergantungan tanaman akan CMA merupakan tingkat relatif di-mana tanaman bergantung pada keberadaan CMA untuk mencapai pertumbuhan maksimum pada tingkat kesuburan tanah tertentu (Khalil et al., 1994). Tanaman yang mempunyai tingkat ke-bergantungan tinggi terhadap keberadaan CMA biasanya akan menunjukkan respons pertumbuh-an ypertumbuh-ang lebih baik, tetapi sebaliknya tpertumbuh-anampertumbuh-an tidak tumbuh dengan baik jika berasosiasi dengan CMA (Setiadi, 2000). Pada sisi lain, tanaman te-tap dapat tumbuh dengan baik jika tidak beraso-siasi dengan CMA, tetapi tidak tumbuh dengan
baik jika berasosiasi dengan CMA (Smith et al.,
1999). Selain ditentukan oleh ketersediaan P ta-nah dan atau pupuk P, kebergantungan terhadap
CMA juga ditentukan oleh jenis CMA (Khalil et
al., 1994). Kebergantungan tanaman terhadap CMA tinggi pada tanah yang mengandung P tersedia rendah, namun pupuk P tetap diperlukan karena selain tanaman membutuhkan P yang cu-kup bagi pertumbuhannya, CMA juga memerlu-kan P bagi keberlangsungan hidupnya.
BAHAN DAN METODE
*) Staf pengajar Fakultas Pertanian Universitas Andalas Padang
kontrol) dan empat level dosis pupuk fosfat (0, 150, 300, dan 450 mg tanaman-1 bulan-1). Percobaan ini merupakan percobaan lapang yang dirancang menurut rancangan acak kelompok berpola faktorial. Pada percobaan ini dikaji 12 kombinasi perlakuan yang diulang 3 kali, sehing-ga terdapat 36 unit perlakuan.
Penanaman bibit manggis ke lapang dilaku-kan pada saat bibit berumur 8 bulan di pembibit-an dpembibit-an bibit tersebut telah diinokulasi dengpembibit-an
CMA jenis Glomus etunicatum dan Glomus
manihotis pada stadia persemaian dengan tingkat infeksinya pada umur 8 bulan adalah 70 %. Pada percobaan ini diberi pupuk buatan N, P, dan K setiap bulan. Pupuk N dan K diberikan dengan dosis yaitu 250 mg tanaman-1 bulan-1, sedangkan pupuk P disesuaikan dengan perlakuan, yaitu 0, 150, 300, dan 450 mg tanaman-1 bulan-1 (0, 1, 2, 3 g pot-1 SP36).
Variabel respons yang diamati adalah infeksi CMA, kandungan P tanaman, kebergantungan terhadap CMA serta bobot kering total bibit manggis. Kebergantungan terhadap CMA dihi-tung berdasarkan formulasi Khalil et al. (1994).
HASIL DAN PEMBAHASAN
1. Tingkat infeksi CMA
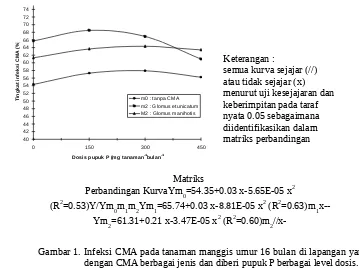
Tingkat infeksi CMA ditentukan oleh efek interaksi antara inokulasi CMA dengan pupuk P. Hubungan tingkat infeksi CMA dengan dosis pupuk P pada tanaman manggis umur 16 bulan di lapangan menunjukkan pola hubungan kuadratik (Gambar1). Pada semua dosis pupuk P, tingkat
infeksi Glomus etunicatum dan Glomus
manihotis selalu lebih tinggi daripada tanpa CMA. Pada pemberian pupuk P dosis rendah, tingkat infeksi Glomus etunicatum selalu lebih
tinggi daripada Glomus manihotis dan sebaliknya
pada tanaman yang diberi pupuk P dosis tinggi. Dari data itu dapat dikatakan bahwa Glomus etunicatum dan Glomus manihotis merupakan CMA yang cocok hidup pada lingkungan rizosfer
tanaman manggis. Perbedaannya, Glomus
etunicatum lebih cocok hidup pada tanaman yang
diberi pupuk P dosis rendah, sedangkan Glomus
manihotis pada tanaman yang diberi pupuk P dosis tinggi. Tanaman manggis yang tidak diino-kulasi dengan CMA pada stadium bibit ternyata terinfeksi juga oleh CMA indigen dan CMA itu lebih cocok hidup pada lingkungan yang diberi pupuk P dosis tinggi.
Dosis pupuk P (mg tanaman-1bulan-1
T
Perbandingan KurvaYm0=54.35+0.03 x-5.65E-05 x2
(R2=0.53)Y/Ym
semua kurva sejajar (//) atau tidak sejajar (x) menurut uji kesejajaran dan keberimpitan pada taraf nyata 0.05 sebagaimana diidentifikasikan dalam matriks perbandingan
Tingkat infeksi maksimum Glomus etunicatum, Glomus manihotis, dan tanpa CMA adalah 68.29, 64.19, dan 58.33% yang masing-masing diperoleh dengan pupuk P optimum secara berurutan 170,26, 288.18, dan 265.49 mg tanaman-1. Hasil itu terjadi karena tingkat infeksi CMA pada akar tanaman ditentukan oleh pupuk P. Penurunan dosis pupuk P meningkatkan tingkat infeksi CMA pada beberapa akar tanaman (Kurle dan Pfleger, 1996). Franken dan Gnadinger (1994) menemukan bahwa tingkat in-feksi CMA pada tanaman yang tidak diberi pu-puk P lebih tinggi daripada tanaman yang diberi pupuk P. Peneliti lain menemukan pula bahwa
Glomus manihotis memberikan toleransi yang tinggi terhadap P tinggi dan CMA tersebut masih dapat menginfeksi akar tanaman dengan kemam-puan yang juga tinggi, walaupun P tersedia di dalam tanah cukup tinggi (Simarmata, 1995).
Ayako et al. (1999) menemukan bahwa tingkat
infeksi Glomus etunicatum lebih tinggi pada akar
kedelai yang diberi pupuk P daripada yang tidak diberi pupuk P.
2. Kandungan P
Kandungan fosfor tanaman manggis ditentu-kan oleh efek interaksi antara CMA dan pupuk P
(Tabel 1 dan 2). Pada semua dosis pupuk P, inokulasi CMA meningkatkan kandungan fosfor pupus dan fosfor akar tanaman manggis, kecuali kandungan fosfor pupus tidak meningkat jika
diberi pupuk P 450 mg tanaman-1. Pada tanaman
yang diinokulasi dengan Glomus etunicatum,
pe-ningkatan kandungan fosfor pupus tertinggi telah
dicapai dengan pemupukan P 150 mg tanaman-1,
sedangkan dengan Glomus manihotis baru
terca-pai dengan pemupukan 300 mg tanaman-1. Secara
umum data itu dapat disimpulkan bahwa kan-dungan fosfor pupus dan akar tanaman manggis dengan Glomus manihotis tinggi pada tanaman yang diberi pupuk P dosis tinggi, sedangkan
dengan Glomus etunicatum tinggi pada tanaman
yang diberi pupuk P dosis rendah.
Pada tanah yang mengandung P rendah di-perlukan pupuk P yang cukup untuk meningkat-kan efektivitas CMA dalam meningkatmeningkat-kan serap-an hara tserap-anamserap-an. Perbedaserap-an itu tampaknya dapat
dipahami karena Glomus manihotis memberikan
toleransi yang tinggi terhadap P tinggi dan CMA tersebut masih dapat menginfeksi akar tanaman dengan kemampuan yang juga tinggi, walaupun hara P tersedia di dalam tanah cukup tinggi
(Simarmata, 1995). Sebaliknya Glomus
etunicatum efektif menginfeksi akar jika tanaman diberi pupuk P dosis rendah (Ayako et al. (1999).
Tabel 1. Kandungan fosfor pupus tanaman manggis umur 16 bulan di lapangan yang diinokulasi dengan CMA pada stadium bibit dan diberi pupuk P
Jenis CMA (M)
Pupuk P (P) (mg tanaman-1 )
0 150 300 450
mg kg-1
Tanpa CMA 0.12 c(c) 0.13 c(b) 0.13 b(b) 0.14 a(a)
G. etunicatum 0.14 a(c) 0.16 a(a) 0.15 a(b) 0.14 a(c) G. manihotis 0.13 b(c) 0.14 b(b) 0.15 a(a) 0.14 a (b)
Keterangan: Angka-angka yang ditandai dengan huruf yang sama (tanpa tanda kurung arah vertikal, dengan tanda kurung arah horizontal) tidak berbeda menurut uji BNT a = 0.05
Tabel 2. Kandungan fosfor akar tanaman manggis umur 16 bulan di lapangan yang diinokulasi dengan CMA pada stadium bibit dan diberi pupuk P
Jenis CMA (M)
Pupuk P (P) (mg tanaman-1 )
0 150 300 450
mg kg-1
Tanpa CMA 0.19 c(c) 0.19 c(c) 0.20 b(b) 0.21 b(a)
G. etunicatum 0.21 a(c) 0.23 a (a) 0.22 a(b) 0.22 a(b) G. manihotis 0.20 b(c) 0.21 b(b) 0.22 a(a) 0.22 a(a)
3. Bobot kering total
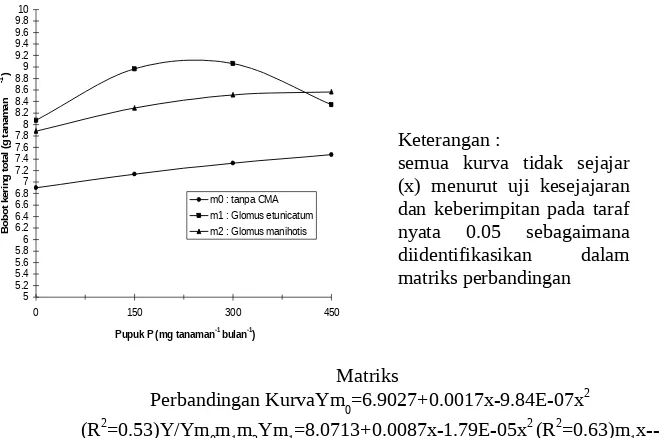
Bobot kering total tanaman manggis umur 16 bulan di lapangan yang diinokulasi dengan CMA berbagai jenis menunjukkan pola hubungan kuadratik dengan pupuk P berbagai level dosis (Gambar 2). Bobot kering totalnya dengan
Glomus etunicatum dan Glomus manihotis me-nunjukkan pola hubungan kuadratik dengan nilai-nilai bobot yang selalu lebih tinggi daripada tan-pa CMA. Bobot kering total maksimum tanaman
manggis dengan Glomus etunicatum, Glomus
manihotis, dan tanpa CMA adalah 9.13, 8.58, dan
7.64 g tanaman-1 yang diperoleh dengan pupuk P
dosis optimum 243.02, 421.99, dan 863.82 mg
tanaman-1. Informasi dosis pupuk P terhadap
ta-naman manggis yang diinokulasi dengan CMA belum ada, namun pupuk P yang diperlukan bagi tanaman manggis dengan inokulasi CMA teruta-ma Glomus etunicatum lebih rendah daripada re-komendasi pupuk P oleh Rukmana (1995), yaitu
400-600 mg tanaman-1.
Efektivitas CMA terhadap bobot kering total itu ternyata sejalan dengan beberapa peneliti lain, CMA dan pupuk P meningkatkan bobot kering
Gmelina arborea, Paraserianthes falcataria dan
Swietenia macrophylla (Mufidah et al., 1999).
Hal itu terjadi karena CMA lebih efektif pada tanaman yang tumbuh di lingkungan bermasalah (Al-Karaki, 1998), seperti P tanah tersedia atau pupuk P rendah (Kurle dan Pfleger, 1996). Lingkungan yang bermasalah itu, terutama yang berhubungan dengan keharaan tanaman yang di-pengaruhi oleh kebergantungan antara CMA dan pupuk P. Dengan demikian dapat dimengerti bah-wa bobot kering total tanaman manggis jelas akan ditentukan pula oleh kebergantungan antara CMA dan pupuk P.
Fakta yang hampir serupa ditemukan oleh peneliti terdahulu bahwa Gigaspora margarita
lebih cocok pada tanaman albisia daripada kaliandra (Suciatmih et al., 1999). Pada albisia,
Glomus manihotis lebih cocok daripada dengan
Glomus fasculatum dan Glomus geosporum
(Suhardi et al., 1999) serta Glomus mosseae
(Mansur et al., 1999). Pada tanaman yang sama,
Scutellospora weresubi lebih cocok daripada
de-ngan Glomus mosseae (Prematuri dan Dodd,
1999). Putri et al. (1999) menemukan pula bahwa
Glomus manihotis Indo-1 lebih cocok daripada
Glomus etunicatum, NPI, Gigaspora roseae
FL-105, dan mycofer pada tanaman mangium,
aka-sia, dan albisia. dan keberimpitan pada taraf nyata 0.05 sebagaimana
diidentifikasikan dalam
matriks perbandingan
Matriks
Perbandingan KurvaYm0=6.9027+0.0017x-9.84E-07x2
(R2=0.53)Y/Ym
0m1m2Ym1=8.0713+0.0087x-1.79E-05x
2 (R2=0.63)m 1
x--Ym2=7.8842+0.0033x-3.91E-06x2 (R2=0.60)m
2
xx-Gambar 2. Bobot kering total tanaman manggis 16 bulan di lapangan yang
4. Kebergantungan terhadap CMA
Tingkat kebergantungan tanaman manggis umur 16 bulan di lapangan ditentukan oleh efek interaksi antara CMA dengan pupuk P (Tabel 3). Tingkat kebergantungan tanaman manggis yang
tidak diberi pupuk P terhadap Glomus etunicatum
lebih rendah daripada tanaman yang diberi pupuk
P 150 mg tanaman-1. Peningkatan pupuk P dari
150 mg tanaman-1 menjadi 450 mg tanaman-1, tingkat kerbergantungan itu menurun, tetapi tetap lebih tinggi daripada tanpa pupuk P. Tingkat
ke-bergantungan tanaman manggis terhadap Glomus
manihotis selalu meningkat sejalan dengan me-ningkatnya dosis pupuk P.
Tabel 3. Kebergantungan tanaman manggis umur 16 bulan di lapangan terhadap CMA yang diinokulasikan pada stadium bibit dan diberi pupuk P
Jenis CMA (M)
Keterangan: Berdasarkan sidik ragam, jenis CMA, pupuk P, dan CMA x P teruji nyata.
Angka-angka yang ditandai dengan huruf yang sama (tanpa tanda kurung arah vertikal, dengan tanda kurung arah horizontal) tidak berbeda menurut uji BNT a = 0.05
Tingkat kebergantungan tanaman manggis terhadap Glomus etunicatum lebih rendah
dari-pada Glomus manihotis jika diberi pupuk P dosis
tinggi (450 mg tanaman-1) dan sebaliknya pada
tanaman yang diberi pupuk P dosis rendah (0,
150, dan 300 mg tanaman-1). Secara umum dari
semua data itu dapat disimpulkan bahwa tingkat kebergantungan tanaman manggis terhadap
Glomus etunicatum tinggi pada tanaman yang di-beri pupuk P dosis rendah, sedangkan
kebergan-tungannya terhadap Glomus manihotis tinggi
pa-da tanaman yang diberi pupuk P dosis tinggi.
KESIMPULAN
(1) Tingkat infeksi maksimum Glomus
etunicatum, Glomus manihotis, dan kontrol pada akar tanaman manggis umur 16 bulan di lapangan adalah 68.29, 64.19, dan 58.33 persen yang diperoleh dengan pupuk P dosis optimum 170.26, 288.18, dan 265.49 mg tanaman-1 bulan-1.
(2) Inokulasi dengan Glomus etunicatum lebih efektif daripada Glomus manihotis, tetapi keduanya lebih efektif daripada kontrol ter-hadap kandungan P dan bobot kering total manggis. Bobot kering total maksimum tanaman manggis yang diinokulasi dengan
Glomus etunicatum, Glomus manihotis, dan kontrol berurut adalah 9.13, 8.58, dan 7.64 g
tanaman-1 yang diperoleh dari pupuk P dosis
optimum 243.02, 421.99, dan 863.82 mg
(3) Tingkat kebergantungan tanaman manggis di
lapang terhadap Glomus etunicatum
terting-gi, yaitu 24.87 % jika diberi 150 mg P ta-naman-1.bulan-1, dan Glomus manihotis
yai-tu 21.72 % jika diberi 450 mg P tanaman-1
bulan-1.
DAFTAR PUSTAKA
Al-Karaki, G.N. 1998. Benefit, cost, and water-use efficiency of arbuscular mycorrhizal durum wheat grown under drought stress. Mycorrhiza 8:41-45.
Ayako, F., P. Katsura, and H. Hiroshi. 1997. Inoculation effect of arbuscular mycorrhizal fungus (AMF) on soybean
(Glycine max) growth and phosphorus uptake under
different fertilized andosol. Papers Presented at the International Coference Mycorrhizas in Sustainable Trop. Agric. and Forest Ecosystem, Bogor, Indonesia, Oct. 26-30, 1997. 5p.
Khalil, S.E., E.L. Thomas, M.A. Tabatabai. 1994. Mycorrhizal dependency and nutrition uptake by improved and unimproved corn and soybean cultivars. Agron. J. 86:949-958.
Kurle, J.E, and F. L. Pfleger. 1996. The effects of cultural practices and pesticide on VAM fungi. p.101-131. In: F.L. Pfleger and R.G. Linderman (eds.). Mycorrhizae and Plant Health. The Amer. Phytopath. Soc. St. Paul, MN.
Mansur, I., J.C. Dodd, P. Jeffries, and Y. Setiadi. 1999. Dual inoculation effects of microbial and AMF isolates on early growth and nitrogen fixation of Paraserianthes
falcatara. p. 219-220. In: F.A. Smith et al. (eds.). Proc.
Int. Conf. Mycorrhizae in Sustainable Trop. Agric. and Forest Ecosystem. Bogor, Indonesia, Oct. 27-30, 1997. Mufidah, A., T. Setiana, N. Mayangsari, Y. Setiadi, dan C.
macrophylla King. Kumpulan Abstr. Sem. Nas. Mikoriza I, Bogor, 15-16 Nop. 1999. Hal 5.
Prematuri, R., and J. C. Dodd. 1999. The effect of arbuscular mycorrhizal fungi on Albisia saman and their bioche-mical detection in roots. p. 219-220. In: F.A. Smith et al. (eds.). Proc. Int. Conf. Mycorrhizae in Sustainable Trop. Agric. and Forest Ecosystem. Bogor, Indonesia, Oct. 27-30, 1997.
Putri, E.R., Y. Setiadi, dan Y. Alkatiri. 1999. Uji keefektifitas beberapa isolat mikoriza arbuskula untuk meningkatkan pertumbuhan semai Paraserianthes falcataria (L) Nielsen, Acacia crassicarpa, A.Cunn ExBenth, dan
Acacia mangium Wild pada media tailing. Kumpulan
Abstr. Sem. Nas. Mikoriza I, Bogor, 15-16 Nop. 1999. Hal 11.
Setiadi, Y. 2000. Pengembangan cendawan mikoriza arbuskula sebagai alat biologis untuk merehabilitasi la-han kritis di Indonesia. Makalah “Seminar sehari”, Peranan mikoriza dalam pertanian yang berkelanjutan. Univ. Padjadjaran, Bandung, 28 Sept. 2000,13 hal. Simarmata, T. 1995. Strategi pemanfaatan mikroba tanah
(pupuk biologis) dalam era bioteknologi untuk mening-katkan produktivitas lahan-lahan marginal di Indonesia menuju pertanian yang berwawasan lingkungan. Maka-lah seminar Bioteknologi, Fakultas Pertanian Unpad, Bandung, 21 hal.
Smith, S. E., S.M. Ayling, G. Rosewarne, S. Dickson, D. P. Schachtman, S. J. Barker, R.J. Reid, G. D. Delp, and F. A .Smith. 1999b. Transport of nutrients between Vesicular-arbuscular (VA). mycorrhizal symbiont insight from molecular and physiological studies. p. 25-34. In: F.A. Smith et al. (eds.). Proc. Int. Conf. Mycorrhizae in Sustainable Trop. Agric. and Forest Ecosystem. Bogor, Indonesia, Oct. 27-30, 1997.
Suciatmih, Suliasi, and N. Hidayati. 1999. Application of microsymbiont and organic fertilizer on fast growing legume trees for reclamation of degraded lands. p. 219-220. In: F.A. Smith et al. (eds.). Proc. Int. Conf. Mycorrhizae in Sustainable Trop. Agric. and Forest Ecosystem. Bogor, Indonesia, Oct. 27-30, 1997. Suhardi, M. Naiem, B. Radjagukguk, O. Karyono, and
Widada, W. W. Wiennarni, T. Herawan. 1997. Interaction among progenies/provenance of sengon (Paraserianthes
falcataria), arbuscular mycorrhizal and rhizobial isolates
grown on Ultisol Soils. Papers Presented at the International Coference Mycorrhizas in Sustainable Trop. Agric. and Forest Ecosystem, Bogor, Indonesia, Oct. 26-30, 1997. 13p.
Syarif, A. 2002. Kebergantungan bibit manggis yang diberi pupuk fosfat terhadap cendawan mikoriza arbuskular. Stigma J.10(3):242-246.