BAB II
TINJAUAN PUSTAKA
2.1. Mioma uteri
Mioma uteri merupakan tumor pelvis yang paling sering pada
wanita. Tumor jinak ini berasal dari miometrium uterus dan secara
histopatologi ditandai dengan sel-sel otot polos seperti kumparan yang
membentuk nodul dengan batas yang tegas. Mioma uteri mempunyai
onset puncak pada dekade ketiga dan keempat kehidupan dan
menyebabkan gejala pada 20-25% wanita usia reproduktif.20 Prevalensi mioma uteri pada wanita kulit adalah sebesar 9% dan pada wanita
Afrika-Amerika 16%. Tetapi hanya sepertiga wanita yang didiagnosa pada saat
operasi yang memang sebelumnya sudah didiagnosa sebagai mioma
uteri, yang menunjukkan bahwa mioma tersebut tidak terdeteksi
sebelumnya atau tidak adanya gejala yang dialami oleh pasien.21 Insidensi kumulatif mioma uteri pada usia 50 tahun adalah 70% pada wanita kulit
putih dan 80% pada wanita Afrika-Amerika.20 Angka kejadian mioma uteri di Amerika Serikat sebesar 8 orang per 1000 wanita tiap tahunnya.22 Sedangkan di Indonesia kasus mioma uteri ditemukan sebesar 2,39% -
11,70% dari semua penderita ginekologi yang dirawat.4
Studi pertama dari patologi tumor ini dilakukan pada tahun 1793,
dan miomektomi abdominal pertama dilaporkan pada tahun 1838. Dengan
kemajuan dalam pembedahan dan anastesia, pada awal tahun 1900-an
Insidensi mioma uteri meningkat dengan bertambahnya usia. Pada
usia 25 – 30 tahun insidensi mioma uteri hanya 0.31 per 1000 wanita,
tetapi pada usia 45 – 50 tahun insidensinya meningkat 20 kali menjadi 6.2
per 1000 wanita. Kesempatan untuk terdiagnosa mioma uteri meningkat
sejalan dengan usia sampai usia 50 tahun, kemudian setelah itu menurun
dengan tajam. Selain usia Obesitas juga meningkatkan resiko mioma uteri
sebesar 18% setiap peningkatan berat badan 10 kg dan terdapat
peningkatan resiko mioma sebanyak 2.3 pada wanita dengan indeks
massa tubuh diatas kuartil.21
Risiko terjadinya mioma uteri menurun dengan peningkatan paritas
dan peningkatan usia saat kehamilan aterm. Nulli paritas merupakan
faktor resiko untuk terjadinya mioma uteri dan dengan adanya kehamilan,
akan mengurangi waktu paparan terhadap unopposed estrogen. Data
menunjukkan bahwa penurunan resiko berkisar dari 20 sampai 50% pada
wanita yang melahirkan minimal 1 kali.7 Wanita dengan 2 kali hamil cukup bulan mempunyai resiko setengah kali menjadi mioma. Merokok
menurunkan resiko (dengan menurunkan kadar estrogen), dan obesitas
meningkatkan resiko (dengan meningkatkan kadar estrogen). Walaupun
resiko mioma yang lebih rendah berhubungan dengan faktor yang
menurunkan kadar estrogen, termasuk kurus, merokok, dan latihan,
pemakaian kontrasepsi oral tidak berhubungan dengan peningkatan
resiko mioma uteri.22 Terdapat anggapan sedikit peningkatan resiko mioma berhubungan dengan usia menars yang dini (7-9 tahun). Siklus
dialami miometrium selama usia reproduktif, yang menyebabkan
peningkatan resiko terjadinya mutasi gen yang mengontrol proliferasi
miometrium.3
Diperkirakan sekitar lebih dari 40% saudara tingkat pertama dari
wanita yang menderita mioma akan menderita mioma uteri juga dalam
kehidupannya.23 Hal ini mungkin tanpa gejala, dan jumlah serta lokasinya sulit diprediksi. Sementara mioma umum terjadi pada semua ras,
tampaknya wanita kulit hitam memiliki insidensi yang sedikit lebih tinggi
daripada etnis lain. Di Amerika, wanita kulit hitam mempunyai resiko 3-9
kali lebih tinggi menderita mioma uteri.4,23,24 Mioma uteri ini menimbulkan masalah besar dalam kesehatan dan terapi yang paling efektif belum
ditemukan, karena sedikit sekali informasi mengenai etiologi mioma uteri
itu sendiri.1
2.2 Patogenesis
Mioma uteri merupakan neoplasma jinak yang muncul dari otot
polos uterus. Penyebab pasti mioma uteri sampai saat ini masih belum
ditemukan. Meyer dan De Snoo mengajukan teori Cell Nest atau teori
genitoblas. Menurut Meyer asal mioma uteri adalah sel imatur, bukan dari
sel-sel otot yang matur. Mioma uteri dipercaya berasal dari mutasi somatik
pada sel miometrium, hasil dari kegagalan proses pertumbuhan. Beberapa
penelitian Glucose-6-phospate dehydrogenase menunjukkan bahwa mioma uteri berasal dari monoklonal. Tumor tumbuh sebagai klon
abnormal secara genetik muncul dari sel progenitor tunggal (tempat asal
Perbedaan kecepatan pertumbuhan dapat menunjukkan
perbedaan sitogenetik yang muncul pada masing-masing tumor. Mioma
uteri multipel dalam satu uterus tidak berkaitan secara klonal satu dengan
yang lainnya, masing–masing mioma tumbuh secara individual. Kehadiran
mioma uteri multipel (dimana memiliki tingkat kekambuhan yang lebih
tinggi dibandingkan yang tunggal) dianggap merupakan predisposisi
genetik terhadap pembentukan mioma uteri. Namun, warisan mioma uteri
dalam keluarga masih belum diteliti dengan baik. Tidak pasti apakah
mioma uteri tumbuh secara individu atau berasal dari mioma yang lain.22 Asal dari mioma uteri belum sepenuhnya dipahami, studi
sitogenetik telah menghasilkan beberapa petunjuk tentang bagaimana dan
mengapa mioma ini berkembang. Tiap tumor berkembang dari sel otot
tunggal, yaitu progenitor miosit, dengan demikian tiap mioma adalah
monoklonal. Analisis sitogenetik telah menunjukkan bahwa mioma
mempunyai kelainan kromosom multipel. Semakin besar mioma, semakin
banyak akan terdeteksi kelainan kariotip. Menariknya, kelainan kromosom
dari mioma mempunyai perubahan kluster yang luar biasa. Duapuluh
persen kelainan melibatkan translokasi antara kromosom 12 dan 14. Tujuh
belas persen melibatkan delesi kromosm 7. Duabelas persen melibatkan
delesi kromosom 12. Regio yang terkena pada kromosom 12 juga
abnormal pada banyak jenis tumor solid lainnya.28
Salah satu teori yang diajukan sabagai penyebab mioma uteri
adalah adanya peningkatan kadar estrogen dan progesteron yang
kemungkinan mutasi somatik. Hal lain yang menyokong adalah adanya
temuan peningkatan kadar RE dan RP yang bermakna pada jaringan
mioma daripada miometrium normal.3,12
Jaringan mioma manusia mengandung jumlah sel stem yang lebih
sedikit daripada miometrium normal. Namun, sel stem yang berasal dari
jaringan mioma, dengan mutasi MED 12 (suatu mediator yang mengkode
gen), yang menunjukkan bahwa minimal diawali dengan satu genetic hit
yang merubah sel stem miometrium, dan interaksi selanjutnya dengan
jaringan miometrium disekitrarnya untuk membentuk mioma.2
Model eksperimen in vivo menunjukkan bahwa pertumbuhan
mioma manusia bergantung pada estrogen dan progesteron yang
memerlukan adanya sel stem somatik multipoten. Dibandingkan dengan
populasi sel mioma atau dengan sel miometrium normal, sel stem mioma
mengekspresikan kadar RE dan RP yang rendah. Pertumbuhan sel stem
mioma memerlukan adanya sel-sel miometrium dengan kadar RE dan RP
dan ligannya yang lebih tinggi, yang menunjukkan bahwa kerja hormon
streroid pada sel stem mioma diperantarai oleh sel-sel miometrium dalam
pengaruh parakrin. Kemungkinan bahwa interaksi parakrin ini dengan
sel-sel sekitarnya mendukung kemampuan sel-self-renewal dari sel-sel stem mioma.2 Jaringan miometrium normal dan mioma mengandung pool sel
dengan kapasitas untuk self-renewal, yang disebut dengan sel stem.
Suatu populasi sel stem bertanggung jawab terhadap proliferasi sel-sel
otot polos miometrium normal (Gambar 1A) yang tampak pada proses
yang matur mengekspresikan kadar RE-α dan RP yang lebih tinggi
daripada sel-sel stem. Dengan demikian kemungkinan proliferasi sel-sel
yang bergantung pada estrogen dan progesteron secara primer di
perantarai oleh RE-α dan RP yang ada pada sel-sel yang matur. Faktor
parakrin seperti ligan WNT, yang dikeluarkan oleh sel-sel matur bekerja
pada sel stem untuk merangsang self-renewal dan proliferasi mereka.
Suatu benturan genetik (genetic hit) seperti mutasi dari MED 12
atau pengaturan ulang kromosom mempengaruhi HMGA2 (suatu protein
yang dapat mempengaruhi proses pada sel seperti difrensiasi, kematian,
pertumbuhan dan proliferasi), yang dapat merubah sel-sel stem
miometrum menjadi sel-sel stem mioma (Gambar 1B). Sel-sel mioma ini
dapat memperbarui diri sendiri dan mulai membelah secara tidak
terkendali sampai berdifrensiasi menjadi sel otot polos mioma. Selama
proses ini sel-sel otot polos mioma memperoleh banyak abnormalitas
epigenetik dan fenotipik. RE-α dan RP terkonsentrasi secara primer pada
sel-sel mioma yang matur dan melewatkan sinyal estrogenik atau
progestogenik pada sel stem melalui mekanisme parakrin. Sel stem
mioma tunggal yang bertransformasi pada akhirnya menjadi mioma yang
jinak dengan batas yang tegas, dimana meluas dalam jaringan
miometrium (Gambar 1C). Pembentukan matriks ekstraselular
berkontribusi terhadap ekspansi tumor.2
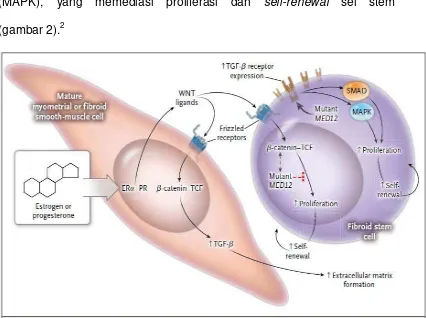
Karena konsentrasi RE-α dan RP sangat tinggi pada sel-sel
miometrium dan sel-sel mioma matur dibandingkankan dengan sel-sel
stem, kemungkinan sinyal estrogen dan progesteron dikirimkan ke sel-sel
stem mioma melalui reseptor hormon pada sel-sel matur dengan cara
parakrin. Estrogen dan progesteron dapat meningkatkan sekresi ligan
WNT, yang bekerja melalui famili frizzeled reseptor yang mengaktivasi
jalur β-catenin-TCF (T-cell Transcription Factor), yang selanjutnya
menginduksi produksi dari TGF-β pada sel-sel matur dan menyebabkan
MED12 yang non-mutan bekerja sebagai modifier fisiologis dari kerja β
-catenin, sedangkan MED12 yang mutan (atau absennya MED12) dapat
menyebabkan kegagalan untuk menyelesaikan fungsi ini. Absennya
MED12 atau adanya bentuk MED12 yang mutan pada sel-sel stem juga
dihubungkan dengan meningkatnya ekspresi reseptor TGF-β, yang
menyebabkan aktivasi dari downstream sinyalnya. Ini kemudian
mengaktivasi protein famili SMAD dan mitogen-activated protein kinase
(MAPK), yang memediasi proliferasi dan self-renewal sel stem
(gambar 2).2
Gambar 2.Interaksi antara hormon ovarium , β-Catenin dan jalur TGF-β, dan MED12 pada sel mioma (dikutip dari 2)
2.3 Hormon steroid
Secara umum estrogen, progesteron dan androgen merupakan
hormon ini diproduksi oleh ovarium. Bahan dasar pembentukan hormon–
hormon ini adalah kolesterol dan proses pembentukan hormon–hormon,
disebut juga steroidogenesis ini dibantu oleh beberapa enzim dan protein
regulator. Kemudian hormon steroid ini akan aktif dan bekerja pada organ
target.29
Gambaran yang mencolok dari mioma uteri adalah
ketergantungan mereka pada hormon steroid ovarium, estrogen dan
progesteron. Aktivitas ovarium penting untuk pertumbuhan mioma, dan
kebanyakan mioma menyusut setelah menopause. Peningkatan dan
penurunan yang tajam pada produksi estrogen dan progesteron yang
berhubungan dengan kehamilan yang sangat dini dan periode paska
melahirkan mempunyai pengaruh yang dramatis pada pertumbuhan
mioma. GnRH analog, yang menekan aktivitas ovarium dan mengurangi
kadar estrogen dan progesteron yang bersirkulasi, menyusutkan mioma
dan mengurangi perdarahan dari uterus.10
Umumnya reseptor-reseptor hormon steroid mempengaruhi
transkripsi gen. Reseptor steroid meregulasi transkripsi gen melalui
beberapa mekanisme, tidak semuanya membutuhkan interaksi langsung
dengan DNA. Peran estrogen yang penting adalah memodifikasi aktifitas
hormonnya sendiri dan yang lainnya dengan mempengaruhi konsentrasi
reseptor. Estrogen meningkatkan respon jaringan target untuknya sendiri
dan terhadap progesteron serta androgen dengan meningkatkan
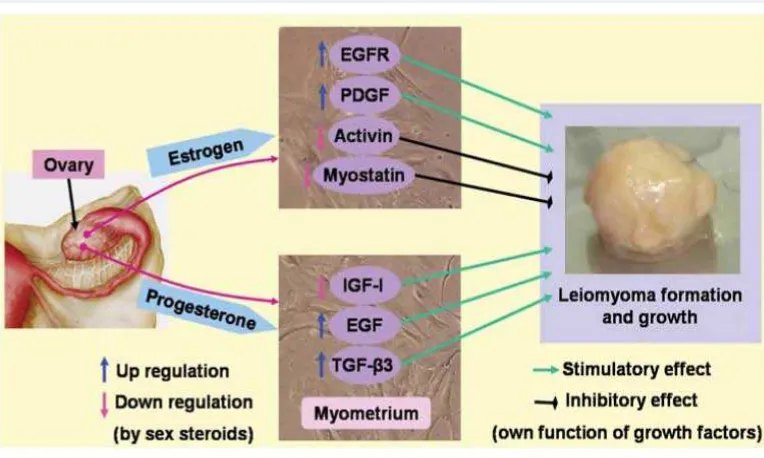
Estrogen dapat meningkatkan pertumbuhan mioma uteri memalui
up-regulation dari ekspresi EGFR dan PDGF dan dengan down-regulating
ekspresi aktivin dan miostatin. Demikian juga dengan progesteron dapat mempengaruhi pertumbuhan mioma uteri dengan up-regulating ekspresi EGF dan TGF-β3. Sebaliknya, progesteron juga dapat menghambat
pertumbuhan mioma dengan down-regulating ekspresi IGF-I. Peranan sebenarnya dari steroid seks, bagaimana mereka berinteraksi dengan
faktor pertumbuhan dan bagaimana mereka mempengaruhi atau mengatur pertumbuhan mioma belum dapat dimengerti. Namun, dengan menggunakan konsep kemampuan seks steroid mempengaruhi
pertumbuhan, beberapa antagonis progesteron / anti progestin, SPRMs
(selective progesterone receptor modulators) dan SERMs (selective
estrogen receptor modulators) telah diajukan mempunyai potensi
terapeutik untuk penanganan mioma.11
Beberapa bukti klinis dan biokimiawi menunjukkan peranan
progesteron dalam patogenesis mioma uteri. Telah ditunjukkan bahwa
progesteron dapat merangsang aktivitas mitosis dan proliferasi mioma.
Kawaguchi dkk (1989) menemukan peningkatan aktivitas mitosis pada
mioma pada fase sekresi siklus menstruasi yang menunjukkan bahwa
pertumbuhan mioma dipengaruhi oleh kadar progesteron. Tiltman (1985)
melaporkan bahwa pemberian medroksiprogesteron asetat meningkatkan
aktivitas mitosis mioma secara bermakna dibandingkan dengan kelompok
yang tidak diobati. Pengobatan dengan antagonis progesteron RU-486
(mifepriston) telah dilaporkan untuk merangsang regresi mioma dengan
mengurangi immunoreaktivitas RP, yang menunjukkan pengaruh
langsung anti progesteron. Sebaliknya, progestin dapat menghambat
pengecilan mioma yang diinduksi GnRH agonis. Brandon dkk (1993)
menunjukkan peningkatan mRNA RP dan kadar protein pada mioma
bersamaan dengan peningkatan proliferasi yang berhubungan dengan
antigen Ki-67 dibandingkan terhadap miometrium normal, yang
menunjukkan hubungan dari sinyal yang diperantarai progesteron dengan
pertumbuhan mioma. Hasil ini mendukung pandangan bahwa progesteron
memegang peranan penting dalam pertumbuhan mioma uteri.30
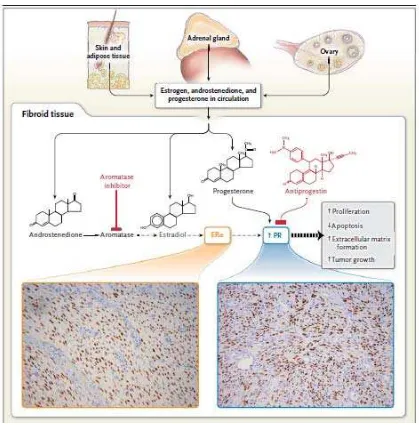
Pada jaringan perifer (kulit dan jaringan lemak) dan ovarium,
aromatase mengkatalisasi pembentukan estrogen, yang mencapai
jaringan mioma melalui sirkulasi. Selain itu, aromatase pada jaringan
mioma mengubah androstenedion yang berasal dari adrenal atau ovarium
estradiol, menginduksi produksi dari RP dengan cara berikatan dengan
RE-α. RP penting sebagai respon dari jaringan mioma terhadap
progesteron yang disekresikan oleh ovarium. Progesteron dan RP sangat
diperlukan terhadap pertumbuhan tumor, meningkatkan proliferasi sel dan
survival dan meningkatkan pembentukan matriks ekstraselular. Pada
ketiadaan progesteron dan RP, estrogen dan RE-α tidak mencukupi untuk
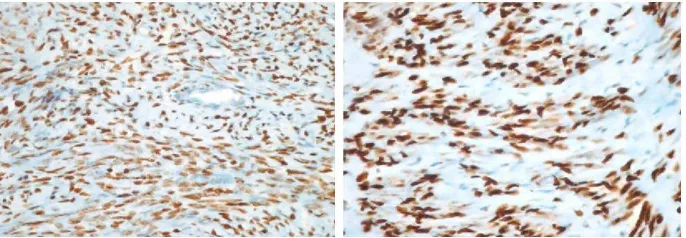
pertumbuhan mioma. Pewarnaan imunuhistokimia pada jaringan mioma
menunjukkan lokalisasi nukleus dari RE-α atau RP pada sel-sel otot polos.
Faktanya bahwa aromatase inhibitor atau antiprogestin yang dapat
mengecilkan ukuran tumor menunjukkan dukungan dari mekanisme ini
dari pertumbuhan mioma (gambar 4).2
2.4 Reseptor Estrogen
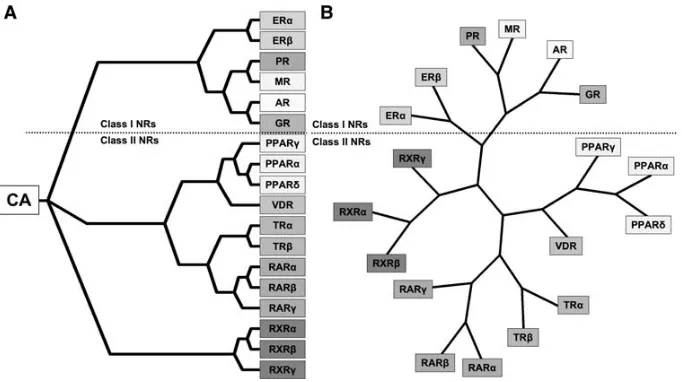
Reseptor estrogen (RE) merupakan anggota dari super famili
reseptor nukleus, dimana kebanyakan berasal dari sumber yang sama
(gambar 5). Super famili ini terdiri dari 18 anggota reseptor, yang dibagi
menjadi reseptor nukleus kelas I dan kelas II. Reseptor nukleus kelas I
termasuk reseptor hormon steroid : reseptor estrogen a dan b (RE a/b),
reseptor progesteron A dan B (RP A/B) reseptor glukokortikoid (RG),
Gambar 4. Efek biologis dari estrogen dan progesteron pada jaringan mioma (dikutip dari 2)
Yang termasuk reseptor nukleus kelas II, reseptor asam retinoat
(RAR a/b/c), reseptor retinoid X (RRX a/b/c), reseptor vitamin D (RVD),
reseptor peroxisome proliferator activated receptor (PPAR a/c/d) dan reseptor thyroid (RT a/b). Semua reseptor dari super famili reseptor
nukleus menghambat faktor transkripsi, dimana menjadi aktif saat
Gambar 5. Super famili reseptor nukleus (dikutip dari 31)
17β - estradiol (estrogen) merupakan ikatan ligan utama pada
RE-α/β. Estrogen disekresikan kedalam pembuluh darah oleh korteks
kelenjar adrenal dan gonad serta memegang peranan yang menonjol
dalam memperantarai perkembangan seksual, fungsi reproduksi,
proliferasi dan difrensiasi dari berbagai jaringan melalui RE. Sebagai
contoh, interaksi Estrogen/RE-α bertanggung jawab untuk proliferasi
payudara dan jaringan uterus yang dirangsang estrogen. RE-α pertama
sekali diisolasi pada tahun 1962, dan gen yang berhubungan di klon pada
tahun yang sama dan berlokasi pada lengan panjang dari kromosom 6
(6q24-q27; sekarang 6q25.1). Tiga dekade kemudian pada tahun 1993,
RE-α pertama tikus percobaan diciptakan dan menemukan bahwa
perkembangan mungkin tanpa RE-α. Pada waktu itu, hanya RE-α yang
difikirkan menjadi reseptor yang memperantarai respon pada estrogen,
tetapi pada tahun 1996 telah di klon REβ dan berlokasi pada kromosom
Estrogen berikatan dengan afinitas yang tinggi pada RE, sementara
hasil metabolik estrogen seperti estron dan estriol, berikatan dengan
afinitas yang lebih rendah. Aksi estrogenik dapat dipengaruhi secara
farmakologi oleh anti estrogen dan SERMs. SERMs yang pertama sekali
di coba adalah tamoxifen pada tahun 1970 dan sampai sekarang
tamoxifen menunjukkan pengaruh pada terapi ajuvan dari kanker
payudara dengan RE (+) pada wanita premenopause.31
Uterus merupakan jaringan target yang sensitif terhadap estrogen
memiliki kedua reseptor dalam jumlah yang banyak. Namun reseptor ini
juga ditemukan di jaringan yang lain, contoh RE-α banyak dijumpai juga
pada ginjal, hati dan jantung. RE-β juga dijumpai pada jaringan otak, paru,
saluran pencernaan dan folikel ovarium.22,26
Pada seluruh sel endometrium dan miometrium, ekspresi RE
mencapai maksimum pada fase folikuler akhir. Selama fase luteal awal,
ekspresi RE menurun, diikuti dengan peningkatan pada pertengahan dan
akhir fase luteal. Perubahan ini menggambarkan perubahan siklus
estradiol. Walaupun RE-β dijumpai pada endometrium manusia, namun
kurang menonjol dibandingkan RE-α dan memperlihatkan perubahan yang
minimal selama siklus menstruasi.22
Lingkungan dalam mioma uteri bersifat hiperestrogenik dan
hipersensitif terhadap estrogen. Mioma uteri menciptakan lingkungan
hiperestrogeniknya sendiri, yang diperlukan untuk pertumbuhan dan
pemeliharaan mereka (mioma uteri). Mioma uteri memiliki RE dan RP
normal)18,25,26,27 sehingga mioma uteri mengikat estrogen lebih banyak dan mioma uteri juga sangat sedikit merubah estradiol menjadi estron
lemah. Tidak dijumpai perbedaan RE yang signifikan berdasarkan ukuran
massa mioma uteri.32
Selain itu, teori mengenai kadar aromatase sitokrom 450 yang
lebih tinggi pada mioma dibandingkan dengan miosit normal. Aromatase
sitokrom 450 merupakan kelompok enzim yang terlibat dalam biosintesis
hormon steroid juga aktivasi metabolik karsinogen. Isoform sitokrom yang
spesifik ini mengkatalisasi konversi androgen menjadi estrogen pada
beberapa jaringan. Diduga sel-sel mioma uteri mensintesis estrogen
in-situ.18 Estrogen dapat menyebabkan pembesaran tumor dengan meningkatkan produksi matriks ekstraseluler. 22,25
Bukti akumulatif mendukung konsep bahwa estrogen sangat
berhubungan dengan tumorigenesis dan pertumbuhan mioma. Estrogen
menggunakan efek fisiologinya pada sel-sel target dengan berikatan pada
reseptor nukleus spesifik yaitu RE-α dan RE-β. RE-β dapat dianggap
homolog dengan RE-α pada daerah ikatan DNA dan daerah ligand-binding. Beberapa studi telah menunjukkan bahwa baik mRNA RE-α dan RR-β diekspresikan pada miometrium dan mioma. Sakaguchi dkk (2003)
melaporkan bahwa kadar mRNA RE-α dan RE-β pada miometrium
berubah selama siklus menstruasi, tetapi kadar mRNA RE-α lebih
menonjol daripada mRNA RE-β. Dua orang penulis telah melaporkan
bahwa kadar mRNA RE-α dan RE-β meningkat pada mioma dibandingkan
menstimulasi transkripsi dari gen target, walaupun aktivasi dari RE-β lebih
rendah daripada RE-α.30 2.5 Reseptor Progesteron
Reseptor progesteron (RP) terdapat dalam 2 bentuk yang berbeda
yang disebut dengan RP-A dan RP-B. Fungsi reseptor ini sebagai faktor
transkiripsi yang diaktivasi-ligan, tetapi isoform kedua reseptor
menunjukkan fungsi biologi yang berbeda. RP-B berfungsi sebagai
aktivator transkripsional dari gen yang responsif progesteron, sedangkan
RP-A bertindak sebagai repressor ligan dependen yang poten dari
aktivitas transkripsional B dalam promoter dan konteks sel dimana
RP-A tidak aktif sebagai aktivator transkripsional. Terdapat hubungan yang
kompleks antara jalur sinyal RE dan RP sebagaimana yang ditunjukkan
dengan observasi bahwa estrogen dapat menginduksi ekspresi RP pada
sel-sel miometrium pada monyet, dan mentransformasi miosit hamster,
serta meningkatkan transkripsi dari gen RP-B pada sel-sel kanker
payudara wanita, sementara isoform kedua RP dapat bertindak sebagai
represor ligan dependen yang poten dari aktivitas RE. kemudian,
progesteron men down-regulate transkripsi RP yang distimulasi
estrogen.30
RP-A dan RP-B telah diidentifikasi pada jaringan mioma dan
miometrium normal. Dua peneliti telah menunjukkan bahwa kandungan
RP-A dan RP-B lebih tinggi pada jaringan mioma daripada miometrium
normal dengan kandungan RP-A yang lebih dominan secara bermakna
antara konsentrasi mRNA yang mengkode RP-A dan RP-B pada mioma
dan miometrium normal, yang menunjukkan kontrol post translasi. Selain
itu, GnRH agosis men down-regulate ekspresi imunoreaktif RP, RP-A dan
RP-B, dan kadar mRNA RP di jaringan mioma. Menariknya, Fujimoto dkk
(1998) menemukan over ekspresi relatif dari mRNA RP-B pada
permukaan mioma, yang menunjukkan bahwa ekspresi yang dominan dari
RP-B pada bagian ini menunjukkan fenotip yang diaktivasi untuk
proliferasi progestasional yang berhubungan dengan pertumbuhan
mioma. Namun, masih belum diketahui apakah tingginya RP-A
berhubungan dengan berkurangnya responsiveness progesteron terhadap
sel-sel mioma.30
2.6 Peranan faktor pertumbuhan pada mioma uteri
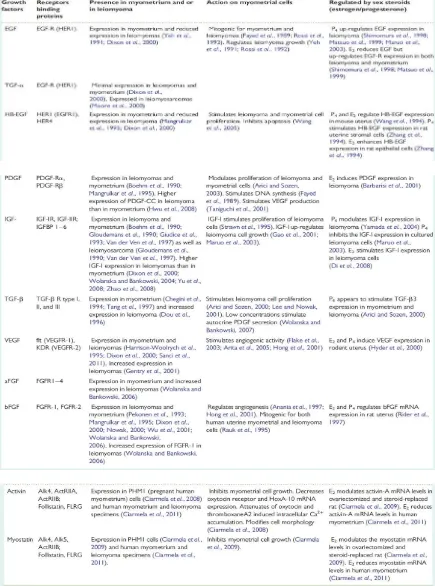
Pada uterus normal, pengaruh estrogen dan progestin pada mioma
diperantarai oleh faktor pertumbuhan. EGF diekspresikan berlebih pada
mioma, reseptor EGF terdapat pada mioma dan pengobatan dengan
GnRH agonis (dan hipogonadisme) menurunkan konsentrasi EGF pada
mioma (tetapi tidak pada miometrium normal). IGF-I dan IGF-II dan
reseptornya berlebihan pada miometrium dan secara aktif dieskpresikan
berlebih pada mioma. Mioma mengekspresikan lebih banyak IGF-II dan
sedikit IGFBP-3 daripada miometrium, suatu situasi yang akan
meningkatkan availabilitas dan aktivitas faktor pertumbuhan pada tumor.
Sel-sel mioma menseksresikan lebih banyak protein yang berhubungan
dengan hormon paratiroid (faktor pertumbuhan lainnya) daripada
mensekresikan prolaktin, dan prolaktin berfungsi pada uterus sebagai
faktor pertumbuhan.22
Salah satu konsekuensi dari perubahan ekspresi faktor
pertumbuhan pada mioma adalah abnormalitas vaskulatur, yang ditandai
dengan pleksus vena yang berdilatasi. Gambaran morfologi ini mungkin
hasil dari regulator vaskular spesifik dari angiogenesis, seperti faktor
pertumbuhan fibroblast dan vascular endothelial growth factor. Perubahan
ini mungkin berkontribusi terhadap perdarahan menstrual yang berat yang
berhubungan dengan mioma submukosa.22
2.7. Imunohistokimia reseptor estrogen dan reseptor progesteron
Imunohistokimia / Immunohistochemistry (IHC) adalah sebuah
metoda pemeriksaan dengan menggunakan prinsip antibodi dengan
spesifikasi yang tinggi untuk menunjukkan lokasi dan keberadaan sebuah
protein dalam jaringan, yang biasanya dilakukan untuk penelitian, dan
tujuan diagnostik atau prognostik.33 Prinsip IHC meliputi langkah : 33
a. Deparafinisasi dan rehidrasi
b. Aktivasi antigen
c. Penghambatan peroksidase endogen
d. Inkubasi antibodi primer
e. Inkubasi antibodi skunder
f. Deteksi antibodi
i. Interpretasi slide.
Penilaian IHC diinterpretasikan berdasarkan gabungan antara
kualitas intensitas ikatan antigen dengan antibodi yang terbentuk di
sitoplasma atau inti sel dengan persentase sel yang terwarnai dalam
lapang pandang. Diantara metode penilaian IHC tersebut adalah : 34
1. H score, merupakan penjumlahan dari persentase sel yang terwarnai lemah, persentase sel yang terwarnai sedang dikalikan
dengan dua, dan persentase sel yang terwarnai kuat dikalikan
dengan tiga. Penilaian ini memberikan skor dari 0 – 300.34,35
2. Allred score, merupakan penjumlahan dari skor persentase sel yang terwarnai (0 = tidak terwarnai, 1 = terwarnai < 1%, 2 = 1-10%,
3 = 10-33%, 4 = 33-67%, 5 = 67-100%) dan skor dari intensitas sel
yang terwarnai (0 = tidak terwarnai, 1 = terwarnai lemah, 2 =
terwarnai sedang, 3 = terwarnai kuat). Penilaian ini memberikan
skor dari 0 – 8.36,37
3. Intensitas warna pada sel, merupakan derajat intesitas sel yang
terwarnai, dengan nilai : negatif (-) jika tidak ada sel yang terwarnai,
(+) jika sel terwarnai lemah, (++) jika sel terwarnai sedang, dan
(+++) jika sel terwarnai kuat.38
2.8 Hipotesis Penelitian
Hipotesis pada penelitian ini adalah :
1. Ekspresi RE pada mioma lebih tinggi daripada miometrium normal.
2.9 Kerangka Teori
2.10 Kerangka Konsep
Variabel tergantung
Variabel bebas
Variabel perancu (tidak diteliti) Ekspresi reseptor estrogen dan
reseptor progesteron pada miometrium