Effect of the biocontrol yeast
Pichia anomala
on interactions
between
Penicillium roqueforti
,
Penicillium carneum
, and
Penicillium paneum
in moist grain under restricted air supply
Marianne E. Boysen, Stina Bjo¨rneholm
1, Johan Schnu¨rer *
Department of Microbiology,Swedish Uni6ersity of Agricultural Sciences(SLU),P.O.Box7025,SE-750 07Uppsala,Sweden
Received 29 July 1999; accepted 18 February 2000
Abstract
Penicillium roqueforti was recently reclassified into the three species P. roqueforti, Penicillium carneum, and
Penicillium paneum based on differences in ribosomal DNA sequences and secondary metabolites, e.g. mycotoxins. This is the first report on interaction between these closely related mould species under stress conditions. The yeast
Pichia anomala(J121) inhibits growth ofP.roquefortiin grain stored in malfunctioning airtight storage systems. The ability ofP.anomalato inhibit all three species of theP.roquefortigroup was examined in separate experiments as well as the competition between the three mould species when co-cultured with or without the yeast in non-sterile wheat grain (aw0.95) under restricted air supply. Mould growth was analysed by dilution plating after 14 days and
the individual colonies identified by random amplified polymorphic DNA (RAPD) fingerprinting. When co-culturing theP.roquefortigroup in wheat withoutP.anomalaall three species were able to grow to the same extent. Also, when co-culturing all species of theP.roquefortigroup together withP.anomala, the growth response of the three species was very similar. At yeast levels of 104CFU g−1grain a pronounced inhibition was observed and at 105CFU g−1
grain a fungicidal effect was detected, indicating a potentiated effect ofP.anomalawhen co-culturing the three mould species. © 2000 Elsevier Science B.V. All rights reserved.
Keywords: Fungal interaction; biocontrol; mould; Penicillium roquefortigroup;Pichia anomala; Random amplified polymorphic DNA (RAPD)
www.elsevier.com/locate/postharvbio
1. Introduction
In recent years, there has been an increased interest in biological control of both field and postharvest growth of fungi (Paster et al., 1993; Ho¨keberg et al., 1997; Petersson and Schnu¨rer, 1998). In a temperate climate Penicillium species are the most important fungi in stored grain and,
* Corresponding author. Tel.: +46-18-673215; fax: + 46-18-673392.
E-mail address:[email protected] (J. Schnu¨rer) 1Present address: Arla R&D, Torsgatan 14, S-105 46 Stock-holm, Sweden.
especially under airtight conditions, Penicillium roqueforti will be the dominating species (Lacey and Magan, 1991).
Based on secondary metabolite profiles, and genetic and morphological studies (Boysen et al., 1996), P. roqueforti was recently divided into the three species P. roqueforti, Penicillium carneum, and Penicillium paneum (hereafter collectively re-ferred to as theP.roquefortigroup).P.roqueforti is able to grow at low oxygen/high carbon dioxide concentrations (Pitt and Hocking, 1997) and as recently shown by Petersson and Schnu¨rer (1998) all three species are able to grow to the same extent on grain with restricted air supply. Since the three species all grow well on media contain-ing up to 0.5% acetic acid (Boysen et al., 1996) they could be expected to compete in similar environments. Little is known about the outcome of interactions between closely related moulds when exposed to stress caused by physical factors (e.g. low oxygen, low water availability, pH) or biocontrol agents. However, studies on field and postharvest moulds of grain have shown that inter- and intra-genera competition varies depend-ing on carbon and nitrogen source, temperature, and water activity (aw). Generally, in vitro and in
vivo interactions between different genera show dominance of Fusarium species, especially at higher aw (0.99 – 0.96), while e.g. Penicillium
spe-cies dominate at lower aw (0.95 – 0.93) (Magan
and Lacey, 1985; Marı´n et al., 1997, 1998a; Reddy et al., 1998). Magan and Lacey (1985) saw that intra-genus interactions of Penicillium species on wheat grain were highly dependent on tempera-ture and aw.
We have previously shown that the yeastPichia anomala J121 is an effective biocontrol organism ofP.roquefortiin airtight stored grain (Petersson and Schnu¨rer, 1995, 1998). In the present study we examined the effect of P. anomala on P. roqueforti,P. carneum, andP.paneum inoculated in dual cultures in wheat grain. Furthermore, we carried out a preliminary test of the competition between the three species in mixed cultures under limited air supply, by co-inoculation of all three mould species in moist grain with or without yeast amendment. Mould growth was quantified by di-lution plating of homogenised grain samples.
DNA was extracted from individual colonies to allow identification from the dilution plates by combining the PCR-based molecular technique random amplified polymorphic DNA (RAPD) fingerprinting and macro-morphological characterisation.
2. Materials and methods
2.1. Fungal isolates and cereal grain
P.anomalaJ121 originating from wheat grain is kept in the fungal collection at the Department of Microbiology, Swedish University of Agricultural Sciences (SLU), Sweden. P. roqueforti IBT18 689 and IBT6754 (type strain) originating from rye bread and blue cheese, respectively; P. carneum IBT14 042 and IBT6884 (type strain) both origi-nating from rye bread; and P. paneumIBT13 929 and IBT12 407 (type strain) originating from bak-ers’ yeast and rye bread, respectively, were all gifts from J.C. Frisvad, Department of Biotechnology (IBT), Technical University of Denmark. Isolates chosen are well-defined culture collection isolates including the type-strains of all three mould species.
Winter wheat (cv. Kosack) of seed quality (without any seed dressing), harvested 1996, was purchased from the Lantma¨nnen Cooperatives, Uppsala, Sweden.
2.2. Inocula
Yeast and mould spore suspensions were pre-pared from yeast malt sucrose broth and on malt extract agar (MEA), respectively (Petersson and Schnu¨rer, 1998).
2.3. Inhibition of P.roqueforti IBT18 689, P. carneum IBT14 042, or P. paneum IBT13 929 in moist wheat by the yeast P. anomala J121
Moistened grain (aw 0.95) was inoculated as
grain. The yeast P. anomala was similarly inocu-lated to achieve 101, 102, 103, 104, 105, or 106 cells
g−1grain. The grain was incubated in ‘mini-silos’
(rubber-sealed tubes containing 17-g portions) with simulated air leakage at 25°C in the dark (Petersson and Schnu¨rer, 1998). This water activ-ity and temperature favour growth of P. roque-forti (Petersson and Schnu¨rer, 1995).
On days 0 and 14, three tubes from each treat-ment were opened, and the contents were diluted with peptone water (2 g l−1
phytone peptone, BBL, USA) with 0.015% Tween 80. Yeast growth was quantified on MEAC plates (MEA supple-mented with 100 ppm chloramphenicol, Sigma Chemical Co., USA) and to avoid growth inhibi-tion by antagonistic yeast on agar plates intended for mould CFU counting, MEACC (MEAC sup-plemented with 10 ppm cycloheximide, Sigma Chemical Co., USA) was used (Bjo¨rnberg and Schnu¨rer, 1993).
2.4. Competition between P.roqueforti, P. carneum, and P. paneum when co-inoculated in moist wheat and inhibition of the mixed culture by P.anomala
Competition between related mould species on natural substrate was tested by co-inoculating the P.roquefortigroup in grain. Grain was moistened (aw 0.95) and inoculated as previously described
(Petersson and Schnu¨rer, 1998). Two combina-tions of isolates were tested: (A) the three type strains (IBT6754, IBT6884, and IBT12 407), or (B) three non-type strains (IBT18 689, IBT14 042, and IBT13 929). Inoculation with spore suspen-sions was made to about 103 spores of each
species g−1grain. The procedure for inoculation,
incubation, and analysis of mould growth was as described above.
The effect of P. anomala on the combined P. roquefortigroup was tested with mould combina-tions as described above (A and B). Moulds were inoculated to about 103spores of each species g−1
grain. P. anomala was inoculated to achieve 103,
104, or 105 cells g−1 grain. Mould growth was
analysed as described above.
The number of mould colony forming units (CFU) g−1 grain was recorded for each of the
species in the mixture after identifying the individ-ual colonies as described below.
2.5. Identification of co-inoculated strains of P. roqueforti, P. carneum, and P. paneum by random amplified polymorphic DNA fingerprinting
Colonies picked at random (up to 100 per experiment) were identified by random amplified polymorphic DNA (RAPD).
The colony top of agar plugs of 5 mm diameter were excised from each colony and DNA ex-tracted using either of two methods: (1) Heating fungal spores and mycelium in NaOH/SDS for 30 min at 105°C as described by Olsson et al. (1999). Samples were diluted about 100 times in sterile H2O and DNA stored at −20°C. (2) Agar plugs
were subjected to DNA extraction using the Fast DNA Prep Kit H (BIO 101, USA). DNA was diluted five times and stored at −20°C. The second method was mostly used as it was faster and gave more consistent results.
Fingerprinting by RAPD was essentially carried out as described by Boysen et al. (1996) using the universal primers NS2 and NS7, which are spe-cific to the small nuclear ribosomal gene (18S rDNA) (White et al., 1990). Reaction mixtures of 15 ml contained 2 ml of the diluted DNA
extrac-tions, 50 mM Tris – HCl, pH 8.0, 50 mM KCl, 3 mM MgCl2, 0.2 mM of each of the dNTPs, 0.25%
Tween 20, 10% (v/v) DMSO, 0.5 mM of either
primer, and 0.6 U AmpliTaq – DNA polymerase (Perkin Elmer Cetus, USA) and amplification conditions as previously described (Boysen et al., 1996).
2.6. Macro-morphological identification of co-inoculated strains of P. roqueforti, P. carneum, and P. paneum
(IBT6884 and IBT14 042): dark blue-green colour with floccose centre, colony margin arachnoid, no exudate production, P. paneum (IBT12 407 and
IBT13 929): green colour, exudate in colony, colony margin entire. Similar characteristics were observed on MEACC, except that all strains had a smaller colony diameter and that the colonies of P. roqueforti were slightly umbonate with an en-tire colony margin,P.carneumhad only a slightly arachnoid colony margin, and ageing colonies of P. paneum had a white colony margin.
2.7. Statistical analysis
All data were log-transformed prior to analysis with the GLM procedure of the SAS statistics software (SAS Institute Inc., USA). Subsequently, significant differences were calculated based on the least significant differences (LSD). Three repli-cates were used for each treatment and all experi-ments were repeated twice.
3. Results and discussion
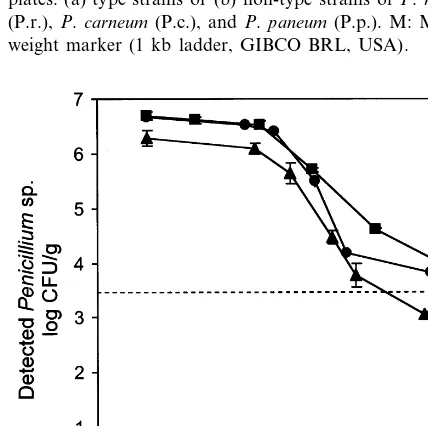
From each of the sets of moulds (A and B) RAPD fingerprinting was carried out on selected colonies to confirm and support the macro-mor-phological identification (Fig. 1). In all cases the identity was confirmed using both NS2 and NS7 as primers. In general, the profiles resembled those previously published for the three species (Boysen et al., 1996), giving e.g. a 0.9 kb band using NS2 and a 2.3 kb band using NS7 for P. roqueforti, and a strong 0.5 kb band using NS2 for P.paneum. Subsequently, all colonies on rele-vant plates were identified based on macro-morphology.
With limited air supply, growth of all three species of the P. roqueforti group in grain was inhibited byP.anomalain a dose-dependent man-ner. When cultured without the addition of P. anomala, mould levels rose from 5×103
CFU g−1
wheat at day 0 to 5×106
CFU g−1
wheat after 14 days (Fig. 2). Increasing the initial inocu-lum level of yeast to 104 cells g−1 wheat had a
markedly inhibitory effect on all three species and at yeast levels of 106 CFU g−1 inhibition was
almost total (Fig. 2). The mould – yeast dual-cul-ture tests carried out in this study thus gave similar results with all three species as previously
Fig. 1. RAPD fingerprinting for identification of single mould colonies. Results of RAPD analysis using either NS2 or NS7 as primer on representative colonies picked from dilution plates. (a) type strains or (b) non-type strains ofP.roqueforti
(P.r.),P.carneum(P.c.), andP.paneum(P.p.). M: Molecular weight marker (1 kb ladder, GIBCO BRL, USA).
Fig. 2. Effect ofP.anomalaas a biocontrol agent on theP.
roqueforti group. Results show effects of increasing initial levels ofP.anomalaon growth of species of theP.roqueforti
group when cultured separately in moist wheat stored under restricted air supply for 14 days.P.roquefortiIBT18 689 ( ),
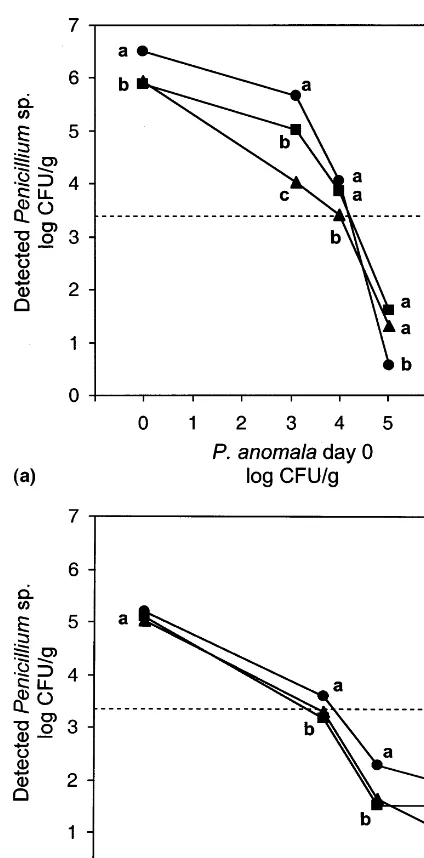
Fig. 3. Interaction of theP.roquefortigroup when exposed to biocontrol. Effects of increasing initial levels ofP.anomalaon growth of mixed cultures of theP.roquefortigroup in moist wheat stored under restricted air supply for 14 days. P.
roqueforti ( ), P. carneum () and P. paneum (). (a) Growth of the type strains (IBT6754, IBT6884, and IBT12 407) in combination. LSD is 0.38 at P=0.05. (b) Growth of the non-type strains (IBT18 689, IBT14 042, and IBT13 929) in combination. LSD is 0.28 atP=0.05. Dotted lines represent mean values of initial inoculated levels of moulds (n=9). Means (n=3) with the same letter for any one set of points are not significantly different (P=0.05) according to the GLM procedure.
shown forP.roquefortionly (Fig. 1a and Fig. 2 in Petersson and Schnu¨rer, 1998).
All three species of theP.roquefortigroup grew to the same extent in mixed cultures without yeast addition. After inoculating about 103 CFU g−1
grain at day 0, all species increased to 105– 106
CFU g−1 grain after 14 days (Fig. 3a and b;
0-level ofP. anomala). No significant change was seen in the relative levels of the three species between day 0 and 14. When adding antagonistic yeast to mixed cultures of theP.roquefortigroup, the three species showed very similar responses (Fig. 3a and b). At initial yeast levels of 104
CFU g−1
grain, a marked degree of inhibition was observed and at 105 yeast cells g−1 grain, mould
levels decreased below inoculation levels after 14 days (Fig. 3a and b), suggesting a potentiating fungicidal effect of the interaction between the moulds. None of thePenicilliumspecies seemed to dominate, although small variations were seen depending on which isolates were used. Also, Petersson and Schnu¨rer (1998) found that three of these isolates, and an additional three isolates, do not differ in their colonisation of grain from a number of barley, rye, and wheat cultivars. To our knowledge, no previous studies have been carried out on the effect on growth/colonisation of interacting closely related fungal species when exposed to a biocontrol agent. However, the ob-served potentiated effect seen in this study sug-gests that in vivo use of biocontrol agents would be favoured by the natural complexity of the microflora in grain.
Magan and Lacey (1985) evaluated interactions between the Penicillium species, P. bre6 icom-pactum, P. 6errucosum, P. hordei, P. piceum, and P. roqueforti on wheat grain at aw between 0.90
and 0.99 and at 15, 25 and 30°C during a 4-week period. At lower aw(0.90 – 0.92) and temperature
(15°C) all species exceptP.piceumcolonised well, and P. roqueforti, in particular, increased from 103CFU g−1to about 107CFU g−1compared to
the others that increased from about 105 to 107
CFU g−1. On the other hand, at higher
aw(0.95 –
differences observed by Magan and Lacey (1985) suggest that in a case like the one described here with three closely related species, competition also could differ depending on environmental factors, allowing one or two of the species to dominate. However, more isolates should be tested in order to evaluate the effects of co-culturing closely re-lated species and especially their growth response to higher doses of biocontrol agents in general.
To our knowledge this is the first report where the molecular technique RAPD has been used for species identification with mixed cultures of fungi. Though tedious, RAPD fingerprinting provides a faster and more reliable way to distinguish colonies from the dilution plates than traditional identification (RAPD results obtainable within 24 h from dilution plates compared to 5 – 7 days culturing on special substrates). This is especially true in cases such as the present study, where fungal morphological characteristics are very sim-ilar and often require complementary tests for exact identification. In a controlled laboratory system, one can learn to distinguish the different species morphologically, but once new/natural strains are entered this might not be the case. Here the RAPD-supported identification provides a reliable alternative to the traditional methods.
Though previous results have shown that the inhibition of Penicillium 6errucosum by P. anomala (J121) also reduces the production of ochratoxin A (Petersson et al., 1998), Marı´n et al. (1998b) found that fuminosin production from Fusarium species was maintained despite growth inhibition by competing fungi. It is therefore im-portant in future studies on the interaction of the P. roqueforti group to analyse the effect on their toxin production during biocontrol.
Acknowledgements
We wish to thank Dirk Jan van der Gaag for helping with the statistical analysis and Jan Sten-lid for helpful comments on the manuscript. This study was supported by the Foundation for Strategic Environmental Research (MISTRA) and the Swedish Farmers’ Foundation for Agricul-tural Research (SLF).
References
Bjo¨rnberg, A., Schnu¨rer, J., 1993. Inhibition of the growth of grain storage molds in vitro by the yeastPichia anomala
(Hansen) Kurtzman. Can. J. Microbiol. 39, 623 – 628. Boysen, M., Skouboe, P., Frisvad, J., Rossen, L., 1996.
Re-classification of thePenicillium roquefortigroup into three species on the basis of molecular genetic and biochemical profiles. Microbiology 142, 541 – 549.
Ho¨keberg, M., Gerhardson, B., Johnsson, L., 1997. Biological control of cereal seed borne diseases by seed bacterization with greenhouse selected bacteria. Eur. J. Plant Pathol. 103, 25 – 33.
Lacey, J., Magan, N., 1991. Fungi in cereal grains: their occurrence and water and temperature relationships. In: Chelkowski, J. (Ed.), Cereal Grain. Mycotoxins, Fungi and Quality in Drying and Storage. Elsevier, Amsterdam, pp. 77 – 118.
Magan, N., Lacey, J., 1985. Interactions between field, and storage fungi on the wheat grain. Trans. Br. Mycol. Soc. 85, 29 – 37.
Marı´n, S., Rull, F., Ramos, A.J., Torres, M., Sala, N., San-chis, V., 1997. Interactions on growth betweenFusarium moniliforme,F.proliferatum, andPenicillium implicatumon maize grain depending on water activity and temperature levels. Cereal Res. Commun. 25, 497 – 498.
Marı´n, S., Sanchis, V., Ramos, A.J., Vinas, I., Magan, N., 1998a. Environmental factors, in vitro interactions, and niche overlap betweenFusarium moniliforme,F.prolifera
-tum, andF.graminearum,AspergillusandPenicillium spe-cies from maize grain. Mycol. Res. 102, 831 – 837. Marı´n, S., Sanchis, V., Rull, F., Ramos, A.J., Magan, N.,
1998b. Colonization of maize grain byFusarium monili
-formeand Fusarium proliferatumin the presence of com-peting fungi and their impact on fumonisin production. J. Food Protect. 61, 1489 – 1496.
Olsson, J., Schnu¨rer, J., Pedersen, L.H., Rossen, L., 1999. A rapid and efficient method for DNA extraction from fun-gal spores and mycelium for PCR-based detection. J. Food Mycol. 2, 251 – 260.
Paster, N., Droby, N., Chalutz, E., Menasherov, M., Nitzan, R., Wilson, C.L., 1993. Evaluation of the potential of the yeast Pichia guilliermondii as a biocontrol agent against
Aspergillus fla6usand fungi of stored soya beans. Mycol. Res. 97, 1201 – 1206.
Petersson, S., Schnu¨rer, J., 1995. Biocontrol of mold growth in high-moisture wheat stored under airtight conditions by
Pichia anomala, Pichia guilliermondii, and Saccharomyces cere6isiae. Appl. Environ. Microbiol. 61, 1027 – 1032. Petersson, S., Schnu¨rer, J., 1998.Pichia anomalaas a
biocon-trol agent ofPenicillium roquefortiin high-moisture wheat, rye, barley, and oats stored under airtight conditions. Can. J. Microbiol. 44, 471 – 476.
Petersson, S., Hansen, M.W., Axberg, K., Hult, K., Schnu¨rer, J., 1998. Ochratoxin A accumulation in cultures ofPenicil
Pitt, J.I., Hocking, A.D., 1997. Fungi and Food Spoilage. Chapman and Hall, New York.
Reddy, V.K., Kumari, D.R., Reddy, S.M., 1998. Effect of carbon and nitrogen sources on the interaction of myco-toxigenic fungi and mycotoxin production. J. Food Sci. Technol. 35, 268 – 270.
White, T.J., Bruns, T.D., Lee, S., Taylor, J., 1990. Amplifica-tion and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J. (Eds.), PCR Protocols: A Guide to Meth-ods and Applications. Academic Press, San Diego, CA, pp. 315 – 322.