IDENTIFIKASI BATANG BAWAH JERUK JC (JAPANCHE CITROEN) ZIGOTIK DAN NUCELLAR BERDASARKAN MORFOLOGI DAN
ANALISIS ISOZIM
(Identification of JC Citrus Rootstock of Zygotic and Nucellar Based on It's Morphology and Isozynae Analysis)
1 2 3
Hardiyanto , Mila , dan S. Lestari 1
PUSAT PENELITIAN DAN PENGEMBANGAN HORTIKULTURA
2
MAHASISWA FAKULTAS PERTANIAN, UNIVERSITAS BRAWIJAYA
3
FAKULTAS PERTANIAN, UNIVERSITAS BRAWIJAYA
ABSTRAK
Penggunaan batang bawah jeruk JC (Japanche citroen) yang “True to Type” sangat penting artinya dalam mensukseskan keberhasilan pengembangan agribisnis jeruk di Indonesia. Oleh karena itu identifikasi bibit jeruk JC sangat diperlukan. Tujuan penelitian adalah untuk memperoleh informasi mengenai ciri-ciri morfologi dan pola pita serta penyebarannya pada bibit jeruk JC hasil perkembangan embrio zigotik dan nucellar. Penelitian dilakukan di Kebun Percobaan Punten, Balai Penelitian Tanaman Jeruk dan Buah Subtropika dan Laboratorium Biologi Molekuler, Fakultas MIPA, Universitas Brawijaya Malang. Bibit jeruk JC umur 2 dan 4 bulan setelah transplanting digunakan pada penelitian ini. Sedangkan pada analisa isozim menggunakan enzim esterase (EST) dan aspartate aminoptransferase (AAT). Bibit jeruk JC nucellar memiliki ciri morfologi yang berbeda dengan bibit hasil perkembangan zigotik terutama pada tinggi tanaman, panjang ruas, percabangan, duri, bentuk daun, dan warna daun. Bibit jeruk JC zigotik mempunyai variasi morfologi yang dapat dikelompokkan menjadi 8 tipe. Berdasarkan analisa isozim, pola pita yang terekspresikan pada bibit jeruk JC zigotik dan nucellar terdapat perbedaan. Jumlah pita pada enzim EST dan AAT pada umur 4 bulan setelah transplanting masing-masing adalah 8 dan 6 pita. Pada enzim EST, penyebaran pita pada bibit jeruk JC nucellar terlihat di tiga lokasi, yaitu bagian atas, tengah, dan bawah. Sedangkan pada bibit jeruk JC zigotik terlihat di dua lokasi yaitu bagian atas dan tengah. Pola pita terbaik ditampilkan oleh enzim AAT baik pada umur 2 dan 4 bulan maupun sample daun muda dan daun tua. Identifkasi bibit jeruk JC berdasarkan morfologi belum dapat dijadikan dasar dalam pengelompokkan bibit zigotik dan nucellar. Penelitian identifikasi bibit jeruk JC melalui penanda molekuler lainnya perlu dicoba dengan beberapa primer dalam upaya menghasilkan pola pita yang jelas yang nantinya dapat digunakan sebagai pegangan dalam menentukan bibit jeruk JC hasil perkembangan embrio zigotik dan nucellar.
Kata kunci : Aspartate aminoptransferase, enzim, esterase, embrio, isozim, jeruk,
ABSTRACT
The use of stock variety JC (Japanche citroen) which is true to type has important meaning to reach success in agribusiness citrus development in Indonesia, therefore the identification of JC seedling are needed. The research was to find the information about morphological characteristics and the band pattern and its spread in JC seedlings which were resulted from zygotic and nucellar embryo. The research was conducted at Punten Field Experiment of Balitjestro and Molecular Biology Laboratory of Mathematics and Natural Science Faculty of Brawijaya University. The 2 and 4 months old JC seedling, enzyme esterase (EST) and aspartate aminoptransferase (AAT) were used for this research. The nucellar seedlings have morphological characteristics that different from the zygotic on plant height, node length, branches, thorn, leaf color and shape. The zygotic seedlings have morphological variation which can be grouped into 8 types. Based on isozyme analysis, the band pattern of zygotic and nucellar seedlings were different. The number of bands in EST and AAT of the 4 months old seedlings were 8 and 6 respectively. In EST, the spread of the bands of nucellar seedlings were observed in 3 locations: top, middle, and bottom while on the zygotic seedlings the bands were observed in 2 locations: top and bottom. The best band pattern showed by AAT enzyme either on 2 or 4 months old seedlings, and either on young or old leaf samples. Morphological characteristics couldn't use for the identifications of JC seedlings yet. The other research for identifications of JC seedling using another molecular probe and primer need to be conducted to obtain an obvious band pattern as a guide to differ zygotic and nucellar seedlings.
Keywords : Aspartate aminoptransferase, citrus, enzyme, esterase, embryo, isozyme,
Japanche Citroen, nucellar, zygotic.
PENDAHULUAN
Perbenihan jeruk merupakan salah satu faktor yang penting bagi pengembangan agribisnis jeruk di Indonesia. Penyediaan benih jeruk yang berkualitas akan menghasilkan buah jeruk yang berkualitas pula. Oleh sebab itu penggunaan teknologi perbenihan jeruk yang tepat diperlukan dalam upaya meningkatkan produktivitas dan kualitas jeruk di Indonesia, salah satunya adalah melalui seleksi (screening) benih jeruk pada tahap perbenihan. Batang bawah jeruk memiliki peranan yang besar dalam menghasilkan bibit jeruk yang berkualitas, sebab umumnya perbanyakan jeruk dilakukan secara vegetatif, yaitu dengan penempelan atau penyambungan mata tunas (scion) pada batang bawah (rootstock). Beberapa batang bawah yang sering digunakan antara lain japanche citroen (Citrus limonia Osbeck), sour orange (Citrus aurantium L.), rough lemon (Citrus jambhiri Lush.), citranges (Poncirus trifoliata x Citrus sinensis), Cleopatra mandarin (Citrus reshni Hort. Ex Tan), Trifoliate orange (Poncirus trifoliata (L.) Raf.), dan Citrumelos (Poncirus trifoliata x
Citrus paradisi) (Ashari et al., 1989). Pemilihan batang bawah dilakukan berdasarkan kondisi lingkungan setempat, misalnya untuk daerah kering dipilih batang bawah yang tahan terhadap cekaman kekeringan. Japanche citroen atau lebih dikenal dengan nama JC merupakan salah satu batang bawah yang banyak digunakan oleh penangkar benih di Indonesia.
Keseragaman genotipe batang bawah merupakan faktor yang penting bagi budidaya jeruk terutama untuk menjaga produktivitas dan kualitas buah yang baik. Salah satu masalah utama pada perbenihan jeruk adalah adanya sifat poliembrioni pada benih jeruk, yaitu benih mempunyai embrio lebih dari satu. Embrio yang dihasilkan dapat berupa embrio zigotik dan nucellar (Ballve et al., 1996; Rodriguez et al., 2004). Embrio zigotik berasal dari peleburan pollen dan ovum, sedangkan embrio nucellar merupakan hasil perkembangan dari sel nucellus tanaman induk. Benih yang berasal dari embrio zigotik tersebut sebagai benih zigotik, demikian pula untuk benih yang berasal dari embrio nucellar disebut sebagai benih nucellar. Embrio nucellar secara genetik sama dengan induk, dan berkembang di dalam kantung embrio (embryo sac) bersama dengan embrio zigotik. Karena memiliki kemiripan sifat dengan induknya, embrio nucellar disebut juga sebagai “true-to-type”, sedangkan embrio zigotik disebut sebagai “off-types” karena berbeda dengan induknya (Moore dan Castle, 1988; Koltunow et al., 1996; Tusa et al., 2002).
Pada saat perbenihan, benih zigotik harus dibuang untuk menjamin kemurnian varietas. Meskipun demikian, penangkar benih jeruk sering mengalami kesulitan dalam membedakan bibit nucellar dan zigotik. Prosentase tanaman nucellar berkisar dari 0% untuk spesies zigotik seperti Citrus maxima sampai 100% untuk spesies nucellar seperti Dancy mandarin. Sebagian besar jeruk mempunyai heterosigositas yang tinggi, oleh sebab itu menghasilkan keturunan zigotik yang beragam. Umumnya kecenderungan untuk menghasilkan embrio nucellar dibandingkan embrio zigotik berbanding 1 : 1, namun dapat terjadi penyimpangan, tergantung dari masing-masing spesies (Davies dan Albrigo, 1991).
Untuk membedakan zigotik dan nucellar umumnya dilakukan oleh pekerja yang berpengalaman dengan mengandalkan perbedaan morfologi antara keduanya. Namun demikian, cara pembedaan ini perlu dievaluasi lebih lanjut mengingat sifat morfologi sangat dipengaruhi oleh lingkungan, sehingga benih JC nucellar hasil seleksi seringkali masih tercampur dengan benih JC zigotik. Oleh sebab itu diperlukan suatu metode seleksi yang lebih pasti, antara lain dengan menggunakan analisis isozim. Analisis isozim dan penanda molekular lainnya seperti RAPD banyak digunakan sebagai penanda genetik untuk mengidentifikasi varietas termasuk membedakan benih nucellar dan zigotik pada jeruk (Torres et al., 1978; Ballve et al., 1997; Anderson et al., 1991). Ashari et al. (1989) telah
menggunakan teknik isozim untuk membedakan zigotik dan nucellar pada beberapa jenis batang jeruk, yaitu rough lemon, trifoliate orange, sweet orange, troyer citrange, dan Cleopatra mandarin dengan menggunakan isozim aspartate aminotransferase (AAT), isocitrate dehy drogenase (IDH), phospho glucoisomerase (PGI), malate dehydrogenase (MDH), dan malic enzyme (ME). Hasil penelitian menunjukkan bahwa benih zigotik dapat dibedakan secara isozim dari benih nucellar pada empat kultivar batang bawah jeruk poliembrionik tersebut. Selain itu juga disebutkan bahwa tidak ada korelasi antara sifat morfologi dengan asal benih.
Analisis isozim digunakan di dalam penelitian dengan cara membandingkan pola pita dari genotipe yang berbeda, sehingga diketahui persamaan dan perbedaannya (Acquaah, 1992). Struktur isozim dikendalikan oleh gen sehingga dapat menampakkan perbedaan yang tegas antara benih zigotik dan nucellar. Analisis isozim dilakukan dengan menggunakan teknik elektroforesis, yaitu suatu teknik pemisahan makro molekul dengan menggunakan arus listrik. Elektroforesis banyak digunakan untuk mendeteksi variasi genetik. Elektroforesis menghasilkan pola pita yang dicatat pada gel. Pola pita tersebut kemudian dibandingkan antar varietas yang berbeda dan dianalisa kemiripan dan perbedaannya.
Adapun penelitian ini bertujuan untuk memperoleh informasi mengenai ciri-ciri morfologi dan pola pita hasil analisis isozim serta penyebarannya pada benih jeruk JC hasil perkembangan embrio zigotik dan nucellar.
BAHAN DAN METODE
Penelitian dilaksanakan di Kebun Percobaan Punten, di bawah Balai Penelitian Tanaman Jeruk dan Buah Subtropika, Batu, sedangkan untuk analisis isozim dilakukan di Laboratorium Biologi Molekular Jurusan Biologi Fakultas MIPA Universitas Brawijaya Malang.
Bahan yang digunakan untuk penelitian ini yaitu benih jeruk japanche citroen umur dua bulan dan empat bulan setelah pindah tanam. Untuk analisis isozim digunakan buffer ekstraksi, gel poliakrilamid yang meliputi separating gel 7.5% dan stacking gel 4%, buffer elektrolit, larutan pewarna untuk enzim esterase (EST) dan aspartate aminotransferase (AAT), serta larutan fiksatif.
Penelitian ini menggunakan metode analisis deskriptif. Untuk ekstraksi daun dan elektroforesis menggunakan metode King et al. (1996). Untuk pembuatan gel dan buffer elektrolit menggunakan metode Indriani (2002). Pewarnaan esterase (EST) dan aspartate aminotransferase (AAT) menggunakan metode Weeden dan Wendel (1989).
Benih diseleksi dari 2100 benih japanche citroen umur dua bulan dan empat bulan setelah pindah tanam. Selanjutnya dipisahkan menjadi dua kelompok berdasarkan pengamatan visual yang dilakukan oleh penangkar benih yang telah berpengalaman, yaitu kelompok benih jeruk JC zigotik (Z) dan kelompok benih jeruk JC nucellar (N). Morfologi yang diamati adalah tinggi tanaman, bentuk daun, warna daun, ruas, tata letak daun, dan duri pada batang.
Untuk analisis isozim, daun diambil pada pagi h ari jam 05.00-06.00 WIB dan dibilas dengan aquades. Daun yang sudah dibilas dimasukkan tissue basah kemudian dimasukkan plastik dan dijaga pada suhu 2ºC sampai 4ºC. Daun jeruk seberat 0.3 gram digerus den gan segera setelah pemetikan den gan menggunakan nitrogen cair sampai menjadi bubuk, kemudian ditambah den gan 0.8 mL buffer ek straksi dan dimasukkan dalam tabung eppen dorf. Sampel disimpan pada suhu 0ºC sampai 2ºC selama 30 menit. Sampel kemudian disentrifugasi den gan kecepatan 12000 rpm selama tiga menit. Supernatan hasil sentrifugasi diambil sebanyak 200 µL dan ditambah dengan 400 µL buffer ekstraksi lalu divortex dan disimpan pada suhu -20ºC (King et al., 1996).
Sampel sebanyak 25 µL diisikan ke dalam sumur dan elektroforesis dilak ukan pada suhu 2ºC sampai 4ºC selama lima jam dengan arus listrik sebesar 5 mA (King et al., 1996).
Pewarnaan (staining) untuk Esterase (EST) dilakukan dengan menuangkan larutan ke dalam wadah yang berisi gel yang telah dielektroforesis, kem udian gel dioven pada suhu 38ºC selama 15 sampai 20 menit. Setelah pola pita tampak, gel dicuci dengan air yang mengalir. Setelah bersih, gel difiksasi den gan gly cerol dan aquades (Weeden dan Wendel, 1989). Sedangkan untuk Aspartate aminotransferase (AAT), larutan pewarna dituang ke dalam wadah berisi gel, kemudian dibiarkan pada suhu ruang dalam keadaan gelap selama 30 menit atau sampai pola pita tampak. Selanjutnya gel dicuci dengan air yang mengalir. Setelah bersih, gel difiksasi dengan glycerol dan aquades (Weeden dan Wendel, 1989).
Pola pita isozim yang diperoleh diamati dengan bantuan lampu pengamatan dan digambar di atas kertas. Berdasarkan gambar yang dihasilkan dilakukan pembuatan zimogram.
HASIL DAN PEMBAHASAN
Hasil pengamatan terhadap kedua kelompok benih jeruk JC yaitu zigotik dan nucellar didapatkan beberapa variasi morfologi. Benih JC nucellar umunya mempunyai morfologi yang relatif seragam, sebaliknya benih JC zigotik memiliki perbedaan morfologi dibandingkan dengan benih JC nucellar dan dapat digolongkan menjadi delapan tipe, yaitu Z1 sampai dengan Z8. Masing-masing tipe tersebut memiliki ciri khas yang membedakannya dari benih JC nucellar (Gambar 1).
Berdasarkan Gambar 1 terlihat bahwa perbedaan morfologi dapat dilihat dari tinggi tanaman, panjang ruas (internode), ada tidaknya duri, panjang duri, percabangan, bentuk daun, ujung daun, ukuran wing petiole, warna daun, dan sudut daun. Sedangkan letak daun, pangkal daun, ada tidaknya wing petiole, dan susunan tulang daun tidak ditemukan adanya perbedaan. Tinggi tanaman pada benih JC zigotik bervariasi, yaitu berkisar antara 15 cm sampai dengan 31 cm, sedangkan untuk benih JC nucellar tinggi.
Gambar 1. Ciri Morfologi Benih Jeruk JC Nucellar dan Zigotik.
(Morphology Characters of JC Nucellar and Zigotics Seedling)
Benih JC Nucellar Benih JC Zigotik 1 Benih JC Zigotik 2 Benih JC Zigotik 3
Tanaman relatif seragam dan dapat mencapai 39 cm. Demikian pula untuk panjang ruas (internode), benih zigotik memiliki panjang ruas berkisar antara 1 cm sampai dengan 1.5 cm, sedangkan pada benih nucellar panjang ruas dapat mencapai 2.5 cm. Sedangkan letak daun pada benih zigotik dan nucellar menunjukkan kesamaan, semua daun tersebar dan terletak berseling.
Hasil elektroforesis menunjukkan bahwa pola pita esterase hanya tampak pada benih JC umur empat bulan, sedangkan pada umur dua bulan pola pita tidak tampak sama sekali. Pada benih JC umur empat bulan pola pita tampak dengan jelas, baik pada sampel daun muda maupun sampel daun tua. Total jumlah pita isozim esterase yang terbentuk pada benih JC umur empat bulan dari sampel daun muda adalah delapan pita. Pita isozim tersebut tersebar di tiga lokasi gel, yaitu di bagian atas, tengah, dan bagian bawah gel (Gambar 2). Pita isozim esterase membentuk delapan tipe pola pita dengan jumlah pita yang bervariasi pada masing-masing tipe (Gambar 3).
Gambar 3. 8 Macam Pola Pita Isozim EST Pada Benih JC 4 Bulan dari Sampel Daun Muda.
(8 Band Pattern of EST Isozyme on Young Leaves Sample of 4 Months JC Seedling)
Gambar 2. Isozim EST Pada Benih JC 4 Bulan dari Sampel Daun Muda.
(EST Isozyme on Young Leaves Sample of 4 Months JC Seedling) Keterangan: Z = Zigote, N = Nuselar
Jumlah pita terbanyak dihasilkan oleh benih JC nucellar N1 dan N2 yaitu sebanyak enam pita, sedangkan untuk benih JC zigotik berkisar antara dua sampai lima pita. Pita tebal hanya tampak pada Z1, N1, N2, dan N3, sedangkan Z2, Z3, Z4, Z5, Z6, Z7, dan Z8 hanya menampakkan pita tipis. Berdasarkan pola pita yang terbentuk, Z2, Z3, dan Z4 memiliki pola pita yang sama, sedangkan Z1, Z5, Z6, Z7, dan Z8 masing-masing memiliki pola pita yang berbeda. Kesamaan pola pita juga ditunjukkan oleh N1 dan N2, namun N3 menunjukkan pola pita yang berbeda dari N1 dan N2, perbedaan tersebut ditunjukkan oleh pita pada bagian atas gel. Variasi tampak pada pita 1, pita 2, pita3, pita 4, pita 6, pita 7, dan pita 8, sedangkan pada pita 5 menunjukkan adanya kesamaan. Pita 5 terdapat pada semua sampel dengan intensitas ketebalan yang sama. Pita 1 hanya dimiliki oleh N1 dan N2, dengan intensitas ketebalan yang tinggi. Demikian pula dengan pita 7 dan pita 8 hanya ditunjukkan oleh benih jeruk JC nucellar.
Hasil elektroforesis untuk isozim esterase pada benih JC umur empat bulan dari sampel daun tua menunjukkan total jumlah pita yang terbentuk sebanyak sembilan pita. Pita-pita tersebut tersebar di tiga lokasi, yaitu pada bagian atas, tengah, dan bagian bawah gel (Gambar 4). Pita isozim esterase tersebut membentuk pola yang dapat dikelompokkan menjadi sembilan tipe (Gambar 5.).
Gambar 5. Pola Pita Isozim EST Pada Benih JC 4 Bulan dari Sampel Daun Muda.
(Band Pattern of EST Isozyme on Mature Leaves Sample of 4 Months JC Seedling)
Gambar 4. Isozim EST Pada Benih JC 4 Bulan dari Sampel Daun Tua.
(EST Isozyme on Mature Leaves Sample of 4 Months JC Seedling) Keterangan: Z = Zigote, N = Nuselar
Jumlah pita terbanyak ditunjukkan oleh Z5, N1, dan N2 yaitu sebanyak enam pita, sedangkan jumlah pita yang paling sedikit ditunjukkan oleh N3 yaitu sebanyak dua pita. Penyebaran pita pada Z3, Z4, Z6, Z7 dan N3 terjadi pada dua lokasi, yaitu di bagian atas dan tengah gel. Demikian pula pada Z2 penyebaran tampak di dua lokasi, yaitu di bagian tengah dan bawah gel. Penyebaran pita pada Z1, Z5, Z8, N1, dan N2 terjadi di tiga lokasi, yaitu di bagian atas, ten gah, dan bagian bawah gel. Berdasarkan pola pita yang terbentuk, terdapat kesamaan pola pita yang ditunjukkan oleh Z3 dan Z7 serta oleh N1 dan N2, sedangkan untuk Z1, Z2, Z4, Z5, Z6, Z8, dan N3 masing-masing memiliki pola pita yang berbeda. Perbedaan pola pita tersebut terdapat pada semua lokasi pita. Dari sembilan pita yang dihasilkan, terdapat variasi pada pita 1, pita 2, pita 3, pita 4, pita 5, pita 7, pita 8 serta pita 9, sedangkan pita 6 menunjukkan adanya kesamaan. Pita 6 terdapat pada semua sampel dengan intensitas ketebalan yang sama. Pita 1 hanya terdapat pada N1, N2 dan N3. Pita 1 pada N1 dan N2 terbentuk tipis, sebaliknya pada N3 pita 1 terbentuk tebal. Pita 6 dimiliki oleh semua sampel dan tampak tebal, sedangkan pita 7 hanya tampak tipis pada Z2, Z3, Z5, Z6, dan Z7. Pita 8 dan pita 9 yang terdapat pada bagian bawah gel hanya terbentuk tipis. Pita 8 hanya terdapat pada Z8, sedangkan pita 9 terdapat pada Z1, Z2, Z5, N1, dan N2.
Hasil elektroforesis untuk isozim AAT pada benih JC umur dua bulan dan empat bulan menunjukkan pola pita yang jelas. Pada benih JC umur dua bulan, hanya dihasilkan lima pita, sedangkan pada benih JC umur empat bulan dapat dihasilkan enam pita. Penyebaran pita pada benih JC umur dua bulan dan empat bulan menunjukkan kesamaan, yaitu pita hanya terdapat pada bagian atas dan bagian tengah gel. Total jumlah pita yang dihasilkan dari elektroforesis untuk isozim AAT pada benih JC umur dua bulan adalah lima pita. Pita-pita tersebut tersebar di dua lokasi, yaitu di bagian atas dan tengah gel (Gambar 6). Pita isozim AAT pada benih JC umur dua bulan dari sampel daun muda membentuk beberapa pola yang dapat digolongkan ke dalam delapan tipe (Gambar 7).
Gambar 6. Isozim AAT Pada Benih JC 2 Bulan dari Sampel Daun Muda.
(AAT Isozyme on Young Leaves Sample of 2 Months JC Seedling) Keterangan: Z = Zigote, N = Nuselar
Jumlah pita terbanyak terdapat pada Z7 yaitu sebanyak lima pita, sedangkan jumlah pita terendah terdapat pada N1 yaitu sebanyak tiga pita. Variasi jumlah pita hanya ditunjukkan pada bagian atas gel. Z1, Z2, Z3, Z4, Z5, Z6 serta N2 memiliki satu pita pada bagian atas gel, sedangkan Z7 mempunyai dua pita. N1 tidak menunjukkan adanya pita pada bagian atas gel. Pita pada bagian tengah gel sejumlah tiga pita dimiliki oleh seluruh sampel, baik benih zigotik maupun nucellar. Kesamaan pola pita hanya ditunjukkan oleh Z2 dan N2, untuk sampel yang lain masing-masing menunjukkan perbedaan. Perbedaan tersebut terdapat pada jumlah pita, lokasi pita, maupun intensitas ketebalan pita. Variasi pita ditunjukkan oleh semua pita kecuali pita 4 yang terdapat pada semua sampel dengan intensitas ketebalan yang sama. Pita 4 semuanya tampak tebal dan terdapat pada benih JC zigotik dan nucellar.
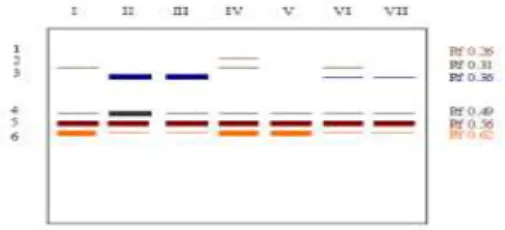
Hasil elektroforesis untuk isozim AAT pada benih JC umur empat bulan dari sampel daun muda menunjukkan total jumlah pita yang terbentuk sebanyak enam pita. Penyebaran pita-pita tersebut terjadi di dua lokasi, yaitu di bagian atas dan tengah gel (Gambar 8). Pita isozim AAT tersebut membentuk pola yang dapat dikelompokkan menjadi tujuh tipe (Gambar 9).
Gambar 7. Pola Pita Isozim AAT Pada Benih JC 2 Bulan dari Sampel Daun Muda.
(Band Pattern of AAT Isozyme on Young Leaves Sample of 2 Months JC Seedling)
Gambar 8. Isozim AAT Pada Benih JC 4 Bulan dari Sampel Daun Muda.
(AAT Isozyme on Young Leaves Sample of 4 Months JC Seedling) Keterangan: Z = Zigote, N = Nuselar
Berdasarkan pita yang terbentuk diketahui bahwa jumlah pita terbanyak dimiliki oleh Z4, Z6, dan Z8, yaitu sebanyak lima pita, disusul kemudian oleh Z1, Z2, Z3, N1, N2, dan N3 sebanyak empat pita, dan terakhir Z5 dan Z7 sebanyak tiga pita. Pita isozim AAT yang terbentuk pada benih JC ini memiliki dua daerah penyebaran, yaitu di bagian atas dan tengah gel, kecuali pada Z5 dan Z7 yang hanya memiliki satu lokasi pita yaitu pada bagian tengah gel. Variasi jumlah pita ditunjukkan pada bagian atas gel. Pada bagian tengah gel terdapat tiga pita dari seluruh sampel, sedangkan pada bagian atas gel terdapat satu sampai dua pita. Kesamaan pola pita juga ditemukan pada semua benih JC nucellar, yaitu N1, N2, dan N3. Elekroforesis isozim AAT pada JC umur empat bulan dari sampel daun muda menghasilkan enam pita yang beragam, kecuali pada pita 5 yang tidak menunjukkan perbedaan. Pita 1, pita 2, dan pita 3 yang terdapat pada bagian atas gel terdapat perbedaan jumlah dan ketebalan pita. Pita 5 terdapat pada semua sampel dan tampak tebal.
Hasil elektroforesis untuk isozim AAT pada benih JC umur empat bulan dari sampel daun tua menunjukkan bahwa total pita isozim yang terbentuk sebanyak enam pita. Pita-pita tersebut tersebar pada dua lokasi penyebaran, yaitu di bagian atas dan bagian bawah gel (Gambar 10). Pita isozim tersebut membentuk pola tertentu yang dapat dikelompokkan menjadi delapan tipe (Gambar 11).
Gambar 9. Pola Pita Isozim AAT Pada Benih JC 4 Bulan dari Sampel Daun Muda.
(Band Pattern of AAT Isozyme on Young Leaves Sample of 4 Months JC Seedling)
Gambar 10. Pita Isozim AAT Pada Benih JC 4 Bulan dari Sampel Daun Tua.
(AAT Isozyme Band on Mature Leaves Sample of 4 Months JC Seedling) Keterangan: Z = Zigote, N = Nuselar
Jumlah pita terbanyak dimiliki oleh Z6, yaitu sebanyak enam pita, kemudian N3 sebanyak lima pita. Jumlah pita terendah sebanyak empat pita dan terdapat pada Z1, Z2, Z3, Z4, Z5, Z7, Z8, N1, dan N2. Semua pita tersebar di dua lokasi, yaitu bagian atas dan bagian tengah gel. Pada bagian atas gel jumlah pita bervariasi, dari satu sampai tiga pita. Namun, tidak ditemukan adanya variasi ketebalan pita pada lokasi ini. Sebaliknya, pada bagian tengah gel terdapat variasi ketebalan pita, namun tidak ditemukan adanya variasi jumlah pita. Terdapat delapan macam pola pita yang tampak pada gel. Tipe I terdiri dari Z1 dan Z3, yang memiliki empat pita dengan pola yang sama. Z2, Z6, Z8, N1, N2, dan N3 masing-masing memiliki pola pita yang berbeda dan dikelompokkan ke dalam tipe yang berbeda pula. Kesamaan pola pita juga ditunjukkan oleh Z4, Z5, dan Z7 yang termasuk dalam tipe III. Hasil elektroforesis tersebut menunjukkan enam pita yang bervariasi, kecuali pita 5 yang seragam pada semua sampel. Pita 1 sampai dengan pita 3 terdapat pada bagian atas gel dengan letak dengan jumlah yang bervariasi untuk setiap sampel. Pita 1 tampak tipis dan terdapat pada Z1, Z2, Z3, dan Z6, demikian pula untuk pita 2 yang juga tampak tipis dan terdapat pada Z6, Z8, N1, dan N3. Pita 3 tampak tipis dan muncul lebih banyak, yaitu di Z4, Z5, Z6, Z7, N2, dan N3. Pada bagian tengah gel terdapat tiga pita yaitu: pita 4, pita 5, dan pita 6. Ketiga pita tersebut pada seluruh sampel dengan ketebalan yang berbeda. Pita 4 tampak tebal hanya pada Z2 dan Z6, namun untuk pita 6 tebalnya pita hanya ditunjukkan oleh Z8 dan N2. Semua pita tampak tebal dan seragam pada pita 5.
Hasil analisis isozim diketahui bahwa pola pita pada batang bawah jeruk hasil perkembangan embrio zigotik dan nucellar menunjukkan perbedaan. Namun, perbedaan pola pita tersebut tidak hanya terjadi di antara benih JC zigotik dan nucellar saja, melainkan juga di antara benih zigotik maupun di antara benih nucellar. Hal tersebut dapat terjadi karena penggolongan benih JC ke dalam tipe zigotik dan nucellar pada penelitian ini didasarkan atas pengamatan secara visual. Hal ini sesuai dengan pernyataan Mooney dan Killen (1991) yang menyatakan bahwa tidak semua tanaman zigotik dapat dibedakan dari
Gambar 11. Pola Pita Isozim AAT Pada Benih JC 4 Bulan dari Sampel Daun Tua.
tanaman nucellar berdasarkan penampakan visualnya. Pengenalan secara visual menjadi mudah dan efektif apabila tetua jantan dan betina berbeda secara signifikan. Berdasarkan hasil elektroforesis untuk isozim esterase pada benih JC diketahui bahwa pola pita untuk isozim ini hanya tampak pada benih JC umur empat bulan, sedangkan pada benih JC umur dua bulan tidak menunjukkan adanya pita. Hal tersebut diduga karena esterase pada benih JC umur dua bulan belum menunjukkan aktifitas enzimatik. Enzim esterase pada tanaman tergolong nonspesifik (Goodman dan Stuber (1983) dalam Acquaah, 1992), dengan kata lain enzim tersebut tidak mengkatalisis reaksi kimia tertentu di dalam tanaman, sebab esterase memiliki spesifisitas substrat yang luas sehingga pita menjadi sulit untuk dipisahkan (Acquaah, 1992). Hal ini dapat dilihat dari resolusi pita yang rendah pada esterase dibandingkan dengan resolusi pita pada AAT. Pada esterase, pita cenderung untuk menyatu, sehingga pita yang terpisah tegas tidak dapat dihasilkan. Kenampakan pita isozim esterase pada benih JC umur empat bulan ditunjukkan baik oleh sampel daun muda maupun sampel daun tua. Dengan demikian dapat diketahui bahwa aktifitas enzim esterase tersebut terdapat pada jaringan muda maupun jaringan tua, tetapi aktifitasnya tidak dapat ditemukan pada fase pertumbuhan awal. Menurut Pierce dan Brewbaker (1973) dalam Acquaah (1992) hasil elektroforesis mengindikasikan bahwa pola dan intensitas pita isozim spesifik untuk bagian atau jaringan tertentu, kematangan jaringan, dan fase pertumbuhan tanaman. Isozim esterase pada benih batang bawah JC umur empat bulan memperlihatkan pita dengan nilai Rf 0.64, yang ditunjukkan oleh pita 5 pada daun muda dan pita 6 pada daun tua, mengalami peningkatan intensitas ketebalan pada daun tua. Ini menunjukkan adanya peningkatan aktifitas enzimatik pada jaringan yang lebih tua. Perbedaan kematangan jaringan tampaknya dapat menimbulkan intensitas ketebalan pita yang berbeda karena jumlah enzim pada setiap fase pertumbuhan tidak selalu sama. Pada hasil elektroforesis dari daun muda diketahui bahwa pita yang terletak di bagian atas gel sebanyak tiga pita dengan nilai Rf berkisar antara 0.33 sampai 0.49. Pada daun tua, jumlah pita pada bagian atas gel sebanyak empat pita dengan nilai Rf 0.31 sampai 0.47. Pada bagian bawah gel, terdapat dua pita dengan nilai Rf yang berbeda. Pada daun muda, pita terbawah memiliki nilai Rf 0.91, sedangkan pada daun tua pita terbawah memiliki nilai Rf 0.82. Dengan demikian dapat diketahui bahwa secara umum jarak migrasi pita pada daun tua berbeda dibandingkan daun muda. Hal tersebut mengindikasikan bahwa terjadi perbedaan berat molekul yang berpengaruh pada gaya gravitasi pada saat proses elektroforesis (Alberts et al., 1994). Dari hasil elektroforesis untuk isozim esterase pada benih JC diperoleh pola pita yang berbeda di antara benih nucellar. Perbedaan tersebut terjadi pada N3, yang memiliki pola pita yang berbeda dari N1 dan N2. Hal ini menunjukkan bahwa masih terdapat kesalahan pengelompokan zigotik dan nucellar yang didasarkan atas pengamatan secara visual.
Dilihat dari pola pita di antara nucellar yang hampir sama, maka perbedaan morfologi yang ditunjukkan oleh N3 mungkin sangat kecil, dan tidak berhasil dideteksi secara visual. Namun, untuk tanaman yang digolongkan sebagai zigotik, semuanya menunjukkan perbedaan yang tegas dibandingkan dengan nucellar, sehingga lebih mudah dideteksi secara visual.
Pita isozim AAT tampak jelas pada benih JC umur dua bulan maupun empat bulan. Ini menunjukkan bahwa aktifitas enzimatik AAT terdapat pada setiap fase pertumbuhan, karena enzim AAT mempunyai peran penting dalam siklus krebs. AAT melepaskan nitrogen dari asam amino untuk diubah menjadi asam keto dalam siklus krebs (Goodman dan Stuber (1983) dalam Acquaah, 1992). Siklus krebs sebagai bagian dari fotosintesis terjadi pada setiap fase pertumbuhan tanaman baik pada jaringan muda maupun jaringan tua, oleh sebab itu aktifitas enzim AAT tampak pada benih JC umur dua bulan dan empat bulan, baik dari daun m uda maupun daun tua. Dari pola p ita isozim AAT pada benih JC umur empat bulan dari sampel daun muda dapat diketahui bahwa pita yang dihasilkan sebanyak enam pita, yang tersebar di dua lokasi. Tiga pita dengan Rf 0.26 sampai 0.36 terletak di bagian atas gel, dan tiga pita yang lain dengan Rf 0.49 sampai 0.62 terletak di bagian tengah gel. Pita 5 (Rf 0.56) memiliki intensitas ketebalan yang sama pada semua sampel. Pola pita dari daun tua menunjukkan adanya perbedaan. Pada daun tua, pita yang terletak pada bagian atas gel memiliki nilai Rf 0.11 sampai 0.23, sedangkan pada bagian tengah gel nilai Rf berkisar 0.38 sampai 0.53. Demikian pula untuk pita yang memiliki intensitas ketebalan sama tiap sampel ditunjukkan oleh pita dengan Rf 0.45. Dengan demikian dapat diketahui adanya perbedaan jarak migrasi pada daun muda dan daun tua. Perbedaan tersebut mengindikasikan adanya perbedaan berat molekul yang berpengaruh pada gaya gravitasi pada saat proses elektroforesis (Alberts et al., 1994). Keseragaman pita isozim AAT pada benih JC umur dua bulan dari daun muda ditun jukkan oleh pita 4 (Rf 0.55), sedangkan p ada benih JC umur empat bulan keseragaman pita ditun jukkan oleh pita 5 (Rf 0.56 pada daun muda dan Rf 0.45 pada daun tua). Pada benih JC umur dua bulan, dihasilkan lima pita isozim yang tersebar di dua lokasi, dua pita di bagian atas gel, dan tiga pita di bagian tengah gel. Pada bagian atas gel, pita isozim AAT memiliki nilai Rf 0.29 dan 0.36, sedangkan pada bagian tengah gel, nilai Rf berkisar antara 0.47 sampai 0.60. Dari jarak migrasi pita pada benih JC umur dua bulan dan umur empat bulan dari sampel daun muda diketahui adanya kemiripan. Nilai Rf untuk pita yang seragam pada benih JC umur dua bulan yaitu 0.55 sedangkan pada benih JC umur empat bulan dari sampel daun muda didapatkan nilai Rf 0.56. Demikian pula untuk jarak migrasi terjauh, pada benih JC umur dua bulan nilai Rf untuk pita terjauh (pita 5) yaitu 0.60, sedangkan pada benih JC umur empat bulan dari sampel daun muda didapatkan nilai Rf 0.62. Dengan demikian diketahui bahwa terdapat peningkatan nilai
jarak migrasi yang menunjukkan adanya penambahan berat molekul seiring dengan pertambahan umur tanaman. Pada hasil elektroforesis isozim AAT ditemui adanya perbedaan di antara benih JC nucellar, yang mengindikasikan bahwa terdapat kesalahan pengelompokan benih JC ke dalam tipe zigotik dan nucellar yang didasarkan atas pengamatan secara visual. Dengan demikian dapat diketahui bahwa penggolongan benih ke dalam tipe zigotik dan nucellar tidak dapat dilakukan sepenuhnya dengan hanya berdasarkan penampakan visual tanaman, karena adanya pengaruh lingkungan yang dapat mempengaruhi aktifitas enzimatik di dalam tanaman. Hal tersebut juga dibuktikan oleh Brown (1978) dalam Acquaah (1992) yang melakukan penelitian terhadap aktivitas enzim alcohol dehydrogenase (ADH). Pada tanah yang dilakukan penggenangan, aktivitas enzim ini meningkat sehingga terjadi perubahan pada pola pita isozimnya. Hal ini dikarenakan enzim ADH sangat berperan dalam respirasi anaerob pada tanaman. Hasil analisis isozim terlihat bahwa hasil yang terbaik ditunjukkan oleh isozim AAT, baik pada umur dua bulan maupun empat bulan dari daun muda maupun daun tua. Sedangkan isozim esterase hanya nampak pada benih JC umur empat bulan pada daun muda dan daun tua dengan resolusi pita yang rendah.
KESIMPULAN DAN SARAN
Benih JC nucellar memiliki morfologi yang berbeda dari benih JC zigotik. Perbedaan tersebut terletak pada tinggi tanaman, panjang ruas (internode), ada tidaknya duri, panjang duri, percabangan, bentuk daun, ujung daun, ukuran wing petiole, dan warna daun.
Benih JC nucellar memiliki morfologi yang relatif sama, namun pola pita yang dihasilkan berbeda, kecuali pita yang dihasilkan oleh isozim AAT pada benih JC umur empat bulan dari sampel daun muda yang menghasilkan pola pita yang seragam di antara benih nucellar.
Benih JC zigotik memiliki pola pita yang berbeda dari benih JC nucellar. Perbedaan tersebut terlihat pada pita isozim esterase maupun AAT.
Pengamatan morfologi tidak dapat dijadikan dasar dalam pengelompokan benih JC ke dalam tipe zigotik dan nucellar karena dipengaruhi oleh lingkungan.
Pola pita terbaik dihasilkan oleh isozim AAT pada benih JC umur dua bulan maupun empat bulan, dari sampel daun muda maupun daun tua.
Penelitian perlu dilanjutkan dengan menggunakan metode penanda DNA seperti RAPD, AFLP, ISSR maupun metode lainnya dengan menggunakan banyak primer agar benih jeruk JC zigotik dan nucellar dapat dibedakan secara jelas dan tepat.
DAFTAR PUSTAKA
Acquaah, G. 1992. Practical Protein Electrophoresis for Genetic Research. Dioscorides Press. Portland Oregon. 127 hal.
Alberts, Bruce, D. Bray, J. Lewis, M. Raff, K. Roberts, dan J.D. Watson. 1994. Biologi Molekuler Sel : Mengenal Sel. Edisi Kedua. Diterjemahkan oleh Alex tri Kantjong W. PT Gramedia Jakarta. Anderson, C. M., W.S. Castle, dan G.A. Moore. 1991. Isozymic Identification of Zygotic Seedling in
Swingle Citrumilo (Citrus paradise x Poncirus trifoliata) Nursery and Field Population. J. Amer. Soc. Hort. Sci. 116 (2) : 322-326.
Ashari, S; D. Aspinal, and M. Sedgley. 1989. Discrimination of zygotic and nucellar seedling of five polyembrionic citrus rootstocks by isozeme analysis and seedling morphology. J. Hort.Science 63: 695-703.
Ballve, Rosa M. L., H.P. Medina-Filho, dan R. Bordignon. 1997. Identification of Reciprocal Hybrids in citrus by The Broadness of The Leaf Petiole Wing. Braz. J. Genet. 20 (4) : 697-702.
Davies, F.S. dan L.G. Albrigo. 1998. Citrus. The University Press. Cambridge. 13-51 pp.
Indriani, F.C. 2002. Keragaman Genetik Plasma Nutfah Kenaf (Hibiscus cannabinus L.) dan Beberapa Spesies yang Sekerabat Berdasarkan Analisis Isozim. Thesis. Fakultas Pertanian Universitas Brawijaya.Malang.
King, B.J., L.S. lee, dan P.T. Scott. 1996. Identification of Triploid Citrus by Isozyme Analysis. Euphytica 90 : 223-231.
Koltunow, A.M., T. Hidaka, dan S.P. Robinson. 1996. Polyembryony in Citrus. Accumulation of Seed Storage Proteins in Seed and in Embryos Cultured in vitro. Plant Physiol. 110 (2) : 599-609. Moore, G.A. dan W.S. Castle. 1988. Morphological and Isozymic Analysis of Open Pollinated Citrus
Rootstock Populations. Journal of Heredity 79 : 59-63.
Rodriguez, M.A., A.V. Monte, G.C. Castaneda, and A.G. Velazquez. 2004. Polyembryony and identification of Volkameriana Lemon zygotic and nucellar seedlings using RAPD. Pesq.Agropec.Bras. 39(6): 551-559.
Torres, A.J., R.K. Soost, and U. Diedenhofen. 1978. Leaf isozymes as genetic markers in Citrus. Amer.J.Bot. 65: 869-881.
Tusa, Abbate, Ferrante, Lucretti, dan Scarano. 2002. Identification of Zygotic and Nucellar Seedlings in Citrus Interploid Crosses by Means of Isozymes, Flow Cytometry, and ISSR-PCR. Cellular and Molecular Biology Letters. 7: 703-708.
Weeden, J.F. dan N.F. Weeden. 1989. Visualization and Interpretation of Plant Isozymes. Dalam Isozymes in Plant Biology. Volume 4. Editor D.E. Soltis dan P.S. Soltis. Dioscorides Press. Portland, Oregon.