J. Biol. Indon. Vol 7, No.2 (2011)
Vol. 7, No. 2 Desember 2011 Akreditasi: No 816/D/08/2009 BOGOR, INDONESIA
JURNAL
BIOLOGI
INDONESIA
ISSN 0854-4425
JURNAL
BIOLOGI
INDONESIA
ISSN 0854-4425
Deforestation in Bukit Barisan Selatan National Park, Sumatra, Indonesia Suyadi
195
Study of Pteridophyte Diversity and Vegetation Analysis in Jatikerep Legonlele and Nyamplung, Karimunjawa Island Central Jawa
Fahreza Saputra & Labibah Qotrunnada
207
Javan Leaf Monkey (Trachypithecus auratus) Movement in a Fragmented Habitat, at Bromo Tengger Semeru National Park, East Java, Indonesia
M.Hari Subarkah, Novianto Bambang Wawandono, Satyawan Pudyatmoko, Subeno , Sandy Nurvianto, & Arif Budiman
213
Impact of Invasive Ant Species in Shaping Ant Community Structure on Small Islands in Indonesia
Akhmad Rizali, Abdul Rahim, Bandung Sahari, Lilik Budi Prasetyo, & Damayanti Buchori
221
Relationship Different Riparian Vegetation Cover with Stream Conditions in Cikapinis Stream, West Jawa
Della Kemalasari & Devi N. Choesin
231
Affect of Canopy Stratum and Methods of Breaking Seed Dormancy on Seedling Growth of Calliandra tetragona Beth. and Acacia tamarindifolia (L.) Willd. Indriani Ekasari
243
Shoot Tip Culture of Nepenthes albomarginata Lobb ex Lindl. In Vitro Lazarus Agus Sukamto, Mujiono, Djukri, & Victoria Henuhili
251
Variasi Gen Mitokondria Cytochrome b pada Dua Jenis Burung Kakatua Putih (Cacatua alba dan C. moluccensis)
Dwi Astuti 263
JURNAL
BIOLOGI
INDONESIA
ISSN 0854-4425
J. Biol. Indon. Vol 7, No. 2 (2011)
Jurnal Biologi Indonesia diterbitkan oleh Perhimpunan Biologi Indonesia.
Jurnal ini memuat hasil penelitian ataupun kajian yang berkaitan dengan masalah biologi yang diterbitkan secara berkala dua kali setahun (Juni dan Desember).
Editor Pengelola Dr. Ibnu Maryanto Dr. I Made Sudiana Deby Arifiani, S.P., M.Sc
Dr. Izu Andry Fijridiyanto
Dewan Editor Ilmiah Dr. Abinawanto, F MIPA UI Dr. Achmad Farajalah, FMIPA IPB
Dr. Ambariyanto, F. Perikanan dan Kelautan UNDIP Dr. Aswin Usup F. Pertanian Universitas Palangkaraya Dr. Didik Widiyatmoko, PK Tumbuhan, Kebun Raya Cibodas-LIPI
Dr. Dwi Nugroho Wibowo, F. Biologi UNSOED Dr. Parikesit, F. MIPA UNPAD
Prof. Dr. Mohd.Tajuddin Abdullah, Universiti Malaysia Sarawak Malaysia Assoc. Prof. Monica Suleiman, Universiti Malaysia Sabah, Malaysia
Dr. Srihadi Agungpriyono, PAVet(K), F. Kedokteran Hewan IPB Y. Surjadi MSc, Pusat Penelitian ICABIOGRAD
Drs. Suharjono, Pusat Penelitian Biologi-LIPI Dr. Tri Widianto, Pusat Penelitian Limnologi-LIPI
Dr. Witjaksono Pusat Penelitian Biologi-LIPI Alamat Redaksi
Sekretariat
d/a Pusat Penelitian Biologi - LIPI
Jl. Ir. H. Juanda No. 18, Bogor 16002 , Telp. (021) 8765056 Fax. (021) 8765068
Email : [email protected]; [email protected] Website : http://biologi.or.id
Jurnal ini telah diakreditasi ulang dengan nilai A berdasarkan SK Kepala LIPI 816/ D/2009 tanggal 28 Agustus 2009.
J. Biol. Indon. Vol 7, No.2 (2011)
JURNAL BIOLOGI
INDONESIA
J. Biol. Indon. Vol 7, No.2 (2011)
KATA PENGANTAR
Jurnal Biologi Indonesia yang diterbitkan oleh PERHIMPUNAN BIOLOGI INDONESIA edisi volume 7 nomer 2 tahun 2011 memuat 17 artikel lengkap, tujuh artikel diantaranya telah dipresentasi pada seminar ATCBC di Bali 2010. Penulis pada edisi ini sangat beragam yaitu dari Balai Penelitian Besar Penelitian Bioteknologi dan Sumber daya Genetik Pertanian Bogor, Balai Tanaman Sayuran Lembang, Balai Besar Penelitian Veteriner Bogor Kementerian Pertanian, BATAN.
Fak. MIPA-Biologi Universitas Indonesia, Fakultas Kehutanan UGM Yogyakarta, Fakultas Kehutanan dan Fakultas Pertanian IPB Bogor, Sekolah Tinggi Hayati dan Departemen Tehnik Kimia ITB Bandung, Fakultas Pertanian Universi-tas Borneo, Tarakan, UniversiUniversi-tas Negeri Yogyakarta, FakulUniversi-tas Sain dan Tehnologi Universitas Islam Hidayatullah Jakarta, Kebun Raya Cibodas LIPI, Puslit Biologi LIPI, Puslit Oseanografi LIPI, PEKA dan Asosiasi Pelestari Curik Bali, Taman Safari Cisarua Bogor. Topik yang dibahas pada edisi ini meliputi bidang Botani, mikrobiologi, zoologi, remote sensing.
J. Biol. Indon. Vol 7, No. 2 (2011)
UCAPAN TERIMA KASIH
Jurnal Biologi Indonesia mengucapkan terima kasih dan penghargaan kepada para pakar yang telah turut sebagai penelaah dalam Volume 7, No 2, Juni 2011: Drs. Roemantyo, Puslit Biologi-LIPI
Dr. Dwi Astuti, Puslit Biologi-LIPI M.Fathi Royani, MA., Puslit Biologi-LIPI Dr. Iwan Saskiawan, Puslit Biologi-LIPI
Drs. Ary Wahyono, Puslit Kemasyarakatan-LIPI Muhamad Irham MSc., Puslit Biologi-LIPI Dr. Enung Fuad, Puslit Bioteknologi-LIPI Drs. Boeadi, Puslit Biologi LIPI (Purna Bakti) Dr. Edi Mirmanto, Puslit Biologi-LIPI
J. Biol. Indon. Vol 7, No.2 (2011)
DAFTAR ISI
Deforestation in Bukit Barisan Selatan National Park, Sumatra, Indonesia Suyadi
195
Study of Pteridophyte Diversity and Vegetation Analysis in Jatikerep Legonlele and Nyamplung, Karimunjawa Island Central Jawa
Fahreza Saputra & Labibah Qotrunnada
207
Javan Leaf Monkey (Trachypithecus auratus) Movement in a Fragmented Habitat, at Bromo Tengger Semeru National Park, East Java, Indonesia
M.Hari Subarkah, Novianto Bambang Wawandono, Satyawan Pudyatmoko, Subeno , Sandy Nurvianto, & Arif Budiman
213
Impact of Invasive Ant Species in Shaping Ant Community Structure on Small Islands in Indonesia
Akhmad Rizali, Abdul Rahim, Bandung Sahari, Lilik Budi Prasetyo, & Damayanti Buchori
221
Relationship Different Riparian Vegetation Cover with Stream Conditions in Cikapinis Stream, West Jawa
Della Kemalasari & Devi N. Choesin
231
Affect of Canopy Stratum and Methods of Breaking Seed Dormancy on Seedling Growth of Calliandra tetragona Beth. and Acacia tamarindifolia (L.) Willd.
Indriani Ekasari
243
Shoot Tip Culture of Nepenthes albomarginata Lobb ex Lindl. In Vitro Lazarus Agus Sukamto, Mujiono, Djukri, & Victoria Henuhili
251
Variasi Gen Mitokondria Cytochrome b pada Dua Jenis Burung Kakatua Putih (Cacatua alba dan C. moluccensis)
Dwi Astuti
263
Kajian Pendahuluan: Perpindahan Gen dari Tanaman Kentang Transgenik Katahdin RB ke Tanaman Kentang Non Transgenik
A. Dinar Ambarwati, M. Herman, Agus Purwito , Eri Sofiari,& Hajrial Aswidinoor
277
Virus Influenza Novel H1N1 Babi di Indonesia
NLP Indi Dharmayanti, Atik Ratnawati, & Dyah Ayu Hewajuli
289
Karakterisasi Produk Biosolubilisasi Lignit oleh Kapang Indigenus dari Tanah Pertambangan Batubara di Sumatera Selatan
Irawan Sugoro, Sandra Hermanto,D. Sasongko,D. Indriani & P. Aditiawati
J. Biol. Indon. Vol 7, No. 2 (2011)
Potensi Virus Avian Influenza H5NI Isolat A/Ck/West Java/Pwt-Wij/2006 Sebagai Vaksin R. Indriani, NLP I Dharmayanti, R.M.A Adjid, & Darminto
Variasi dan kekerabatan genetik pada dua jenis baru belimbing (Averrhoa leucopetala Rugayah et Sunarti sp nov dan A. dolichorpa Rugayah et Sunarti sp nov., Oxalidaceae)
berdasarkan profil Random Amplified Polymorphic DNA Kusumadewi Sri Yulita
Pengaruh Dinamika Faktor Lingkungan Terhadap Sebaran Horisontal dan Vertikal Katak Hellen Kurniati
Merekonstruksi Habitat Curik Bali Leucopsar rothschildi Stresemann, 1912 di Bali Bagian Barat
Mas Noerdjito, Roemantyo &Tony Sumampau
Struktur dan Komposisi Vegetasi Hutan Semusim Habitat Curik Bali (Leucopsar rothschildi Stresemann, 1912) di Kawasan Labuan Lalang, Taman Nasional Bali Barat
Roemantyo
Sumbangan Ilmu Etnobotani dalam Memfasilitasi Hubungan Manusia dengan Tumbuhan dan Lingkungannya
263
Jurnal Biologi Indonesia 7 (2): 263-276 (2011)
Variasi Gen Mitokondria Cytochrome b pada Dua Jenis Burung Kakatua
Putih (Cacatua alba dan C. moluccensis)
Dwi Astuti
Bidang Zoologi, Pusat Penelitian Biologi-LIPI, Cibinong Science Centre, Jl. Raya Jakarta-Bogor, Cibinong-Indonesia. E-mail: [email protected]
ABSTRACT
Variation in The Mitochondrial Cytochrome b Gene in The Two White Cockatoo Species (Cacatua alba and C. molucensis). DNA sequence variation in the 791-bp of mitochondrial cytocrome b gene in the two white cockatoo species (C. alba and C. moluccensis) were analyzed in this study. Two pairs of internal primers used to amplify two fragments of cytochrome b from 30 individuals cockatoo. The results show that there were genetic variations among individuals of C. alba and C. moluccensis. Twenthy eight haplotypes occured in 30 individuals studied; 14 haplotypes (Hca1-Hca14) in 16 individuals of C. alba, and 14 haplotypes (Hcm1-Hcm14)in 14 individuals of C. moluccensis. Hca5 was dominant and owned by 3 individuals (H37, KBS62, 28, BBP88). Within C. alba there were 18 variable sites, 0.00701 of nucleotide diversity (Pi), 0.975 ± 0.035 of haplotype diversity (Hd), and 0.005 ± 0.002 of mean genetic distance. Whitin C. moluccensis there were 18 variable sites, 0.00830 of Pi, and 0.9999 ± 0.028 of Hd, and 0,010 ± 0.002 (0.001-0.010) mean genetic distance. Divergence between C.
alba and C. moluccensis was 0.064 ± 0.088 %. Neigbor-joining (NJ) analysis showed two main
clusters consisted of : C. alba and C. moluccensis separately, and indicated that although there were some variations in the DNA sequences, but the individuals within a species remain clustered in the same cluster.
Key words: genetic variation, mitochndrial cytochrome b, cockatoo bird, Cacatua alba, Cacatua moluccensis
PENDAHULUAN
Di dunia terdapat 11 jenis kakatua putih yg termasuk dalam marga Cacatua.
Enam jenis diantaranya terdapat dan endemik di kepulauan Indonesia, dan C. alba dan C. moluccensis merupakan kakatua yang endemik di kepulauan Maluku. Menurut Forshaw (1989) secara umum burung kakatua memiliki jambul di kepalanya dan paruh besar yang melengkung dan kuat. C. moluccensis
memiliki bulu tubuh dengan warna pink,
panjang tubuh 52 cm. C. alba bulu tubuhnya putih dengan panjang tubuh 46 cm.
Saat ini populasi kedua jenis kakatua tersebut di alam cenderung mengalami penurunan, sehingga tercatat dalam dan menjadi prioritas dalam World Conservation Union, IUCN 2006, dan termasuk ke dalam to appendics CITES; dimana C. moluccensis di dalam appendics I dan C. alba di dalam appendics II.
264
Dwi Astuti
Untuk tujuan conservasi dari kedua jenis kakatua ini, maka berbagai penelitian dan kegiatan lain dilakukan baik secara
ex-situ maupun in-situ. Semenjak
penangkaran diketahui berperan penting di dalam mekanisme konservasi satwa langka, maka upaya koservasi kedua jenis kakatua ini dilakukan di beberapa Taman Margasatwa dan penangkaran. Dari perspektif konservasi, harus diprioritas-kan pada kuantifikasi dan karakterisasi tingkat diversitas genetiknya bagi semua satwa terancam punah, karena ini dapat menuntun kita dalam mengevaluasi efek genetik dari perubahan populasi pada masa mendatang dan dapat dijadikan sebagai petunjuk dalam pengelolaan konservasi yang bertujuan untuk menjaga tingkat diversitas saat sekarang (Roques & Negro 2005). Satwa terancam punah memiliki ciri dengan tingkat variasi genetik yang rendah, khususnya bila dibanding-kan dengan jenis kerabat yang tidak terancam punah (Ardern & Lambert 1997; Spielman et al. 2004). Jadi diperlukan pengetahuan tentang variasi ditingkat molekuler dengan menggunakan analisis penanda molekuler.
Penanda molekuler telah berguna untuk menjawab berbagai pertanyaan menyangkut upaya konservasi dan biologi populasi pada berbagai jenis burung , misal pada S. albifrons (Whittier et al.
2006). DNA mitokondria adalah penanda genetik yang sangat penting digunakan dalam mempelajari evolusi, kekerabatan, dan variasi genetik pada berbagai taxa hewan (Kocher et al. 1989, Ingman et al. 2000) dan juga sering digunakan untuk mengevaluasi diversitas genetik pada berbagai jenis burung (Gill et al. 2005; Martens et al. 2006). .
Cytochrome b adalah salah satu dari
gen pada mitokondria yang mengkode protein dan diketahui sebagai penanda DNA dalam mengungkap sejarah evolusi hewan (Kocher et al. 1989; Montgelard
et al. 1997; Prusak et al. 2004) bahkan
untuk mempelajari variasi genetik pada tingkatan jenis maupun marga pada mamalia, burung, reptile, ikan dan hewan vertebrata lainnya. Pada ikan marga
Cynolebia, cytochrome b menunjukkan
tingkat divergensi sekuen yang tinggi (28 %) antara jenis-jenis ikan tersebut (Garci
et al. 2000), namun pada Macruronus
menujukkan kurangnya diferensiasi genetik (Olavarria et al. 2006).
Pada penelitian ini dilakukan analisis pada sekuen DNA dari mitokondria cytochrome-b pada dua jenis burung kakatua; Cacatua alba dan C.
moluccensis untuk mengetahui variasi
genetik diantara individu dalam satu jenis
(intraspecies) maupun beda jenis
(interspecies).
BAHAN DAN CARA KERJA Tiga puluh (30) sampel darah dari masing-masing individu burung kakatua (C. alba dan C. moluccensis) dikoleksi dari taman Margasatwa, penangkaran, maupun Taman Burung di Indonesia. Darah dipreservasi dengan etanol absolut di dalam tabung plastik 2 mL dan disimpan dalam almari es pada temperatus 4 ºC di Laboratorium Genetika, Bidang Zoologi-Puslit Biologi_LIPI, Cibinong. DNA total diekstraksi dari masing-masing sampel, dengan menggunakan DNA Extraction Mini Kid Qiagen. Larutan DNA yang didapat di simpan dalam almari es 4 ºC dan untuk selanjutnya digunakan pada
265
Variasi Gen Mitokondria Cytochrome b pada Dua Jenis Burung
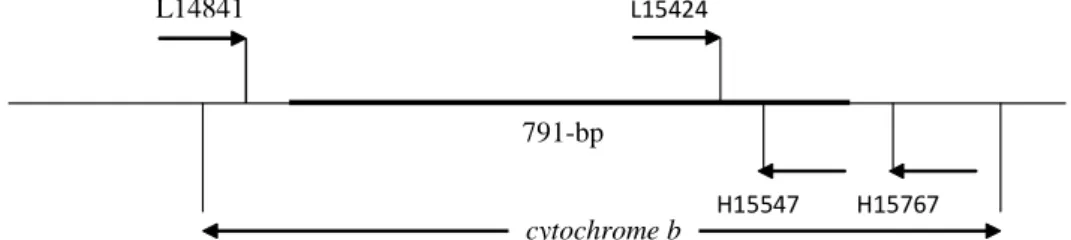
L14841 L15424
H15547 H15767 791-bp
cytochrome b
proses analisis berikutnya. Probociger
aterrimus digunakan sebagai outgroup
dalam mengkonstruksi pohon NJ. Dua fragmen dari gen cytochrome
b diamplifikasi melalui proses PCR dengan menggunakan dua pasang primer internal: L14841 (Kocher et al. 1989) / H 15547 dan L15424/H15767 (Edwards
et al. 1991). Posisi primer-primer ini
digambarkan pada Figure 1. Larutan untuk PCR dibuat dalam volume 20 ìL , dan dalam kondisi 35 siklus dari (denaturasi 94°C - 1 menit; penempelan 52°C -1 min; ekstensi 72°C -2 menit) dan dilengkapi dengan 1 x siklus pada 72°C -10 min. Selanjutny 3 mL dari produk PCR dielektroforesis pada 1 % gel agarose dan direndam dalam larutan ethidium bromide, dan divisualisasi di bawah ultraviolet illuminator. Sisa larutan produk PCR dipurifikasi dengan QIAquick (QIAGEN) dan kemudian disekuen DNA-nya pada dua sisi (forward-reverse) using the ABI Prism™ 3100 automated sequencer. Primer yang digunakan sama dengan primer PCR.
Proses mensejajarkan (aligment) sekuen DNA dari semua individu yang diteliti, dilakukan dengan dengan menggunakan perangkat lunak ProSeq software. Karakter data sekuen DNA
seperti komposisi basa, subtitusi transisi-transversi, jarak genetik, situs bervariasi, dan sekuen variasi dianslisis mengguna-kan perangkat lunak MEGA3 (Kumar et al. 2004) . Diversitas nukleotida dan diversitas haplotipe dianalisis dengan perangkat lunak DnaSP4. Data tersebut juga dikonfirmasi ke asam amino untuk mengetahui apakah fragmen DNA yang teramplifikasi adalah bener-benar target yang diinginkan. Untuk mengkonfirmasi bahwa variasi genetik yang terdapat diantara individu dalam satu spesies tidak mempengaruhi dalam pengelompoka-nnya, maka dikonstruksi pohon neighbor-joining (Saitou & Nei 1987) berdasarkan Kimura’s 2-parameter distance pada MEGA3 dan nilai bootstrap dianalisis dengan 1000 x pengulangan.
HASIL
Pasangan primer L14841/ H 15547 mengamplifikasi fragmen DNA sepanjang sekitar 760-bp, sedangkan L15424-H15767 mengamplifikasi fragmen 370-bp. Pada proses menseja-jarkan data sekuen DNA (aligment), sepanjang sekitar 100-bp data sekuen posisinya overlapping diantara dua set primer tersebut. Disamping itu,
Gambar 1 Posisi dari primer-primer yang digunakan dalam mengamplifikasi gen cytochrome
b. Garis tebal dengan angka 791- bp adalah posisi dan panjang sekuen DNA yang
266
Dwi Astuti
beberapa data sekuen DNA dari beberapa individu kakatua yang diteliti tidak lengkap, sehingga hanya 791-bp yang dianalisis pada penelitian kali ini. Hasil dari proses menterjemahkan data sekuen DNA menjadi asam amino, menunjukkan bahwa fragmen ataupun sekuen DNA yang teramplifikasi, benar-benar merupakan gen target, ini ditunjukkan dengan tidak terdapatnya
stop codon sebagi ciri khas dari gen
pengkode protein.
Komposisi basa timin, sitosin, adenin, dan guanin pada sekuen DNA
cytochrome b sepanjang 791-bp
ditampilkan dalam Tabel 1. Kompo-sisinya meliputi 40.3 % basa purine dan 59, 7 % basa pirimidin. Komposisi guanin paling rendah (14,2 %) dan sitosin tertinggi (34,2 %). Ini juga terlihat pada
cytochrome b baik pada C. alba
maupun C. moluccensis jika kedua spesies ini dianalisa secara terpisah. Substitusi transisi dan transversi
Transisi merupakan substitusi antara dua basa purin (adenin dan guanin) atau
Posisi codon
Komposisi basa
C. alba C. moluccensis (C. alba & C. moluccensis)
T C A G T C A G T C A G
Total 25.8 34.0 26.5 13.7 25.5 34.4 25.7 14.4 25.5 34.2 26.1 14.2
ke-1 23.5 29.2 24.8 22.6 23.3 29.1 24.8 22.8 23.6 29.0 24.8 22.6
ke-2 38.6 28.0 20.0 13.4 38.4 28.0 20.3 13.3 38.5 27.6 20.1 13.7
ke-3 15.3 44.7 34.9 5.1 14.9 46.2 32.0 6.9 14.4 46.1 33.4 6.2
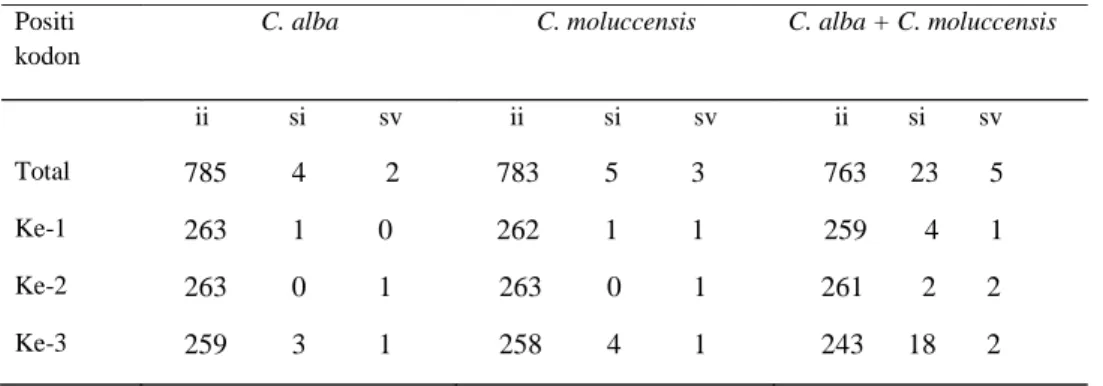
Tabel 1 : Komposisi basa di dalam 791-bp gen cytochrom b pada kakatua yang diteliti antara dua basa pirimidin (timin dan sitosin). Tranversi merupakan substitusi basa antara purin dan pirimidin. Table 2 menampilkan frekuensi substitusi basa yang terjadi pada 30 data sekuen
cytochrome b sepanjang 791-bp,
terdapat 23 substitusi yang bersifat transisi, yang meliputi 11 purin (T <--> C) dan 12 pirimidin (A <--> G). Sedangkan substitusi transversi hanya 5. Jumlah transisi lebih banyak dari transversi, ditunjukkan baik pada C. alba maupun C. moluccensis.
Pada 30 individu yang dianalisis, substitusi basa terjadi paling banyak pada posisi codon 3 (20 substitusi) diikuti codon pertama (5 substitusi) dan kedua (4 substitusi). Begitu juga jika analisis dilakukan secara terpisah, baik pada C. alba maupun C. moluccensis menunjuk-kan substitusi basa yang juga mengikuti pola yang sama yaitu substitusi paling banyak terjadi pada posis codon ketiga, diikuti posisi pertama dan kedua (Tabel 2).
Jarak genetik antara individu-individu
267
Variasi Gen Mitokondria Cytochrome b pada Dua Jenis Burung
(between species) adalah 0.064 ± 0.088.
Sedangkan jarak genetik di antara individu-individu di dalam satu jenis (within species) kakatua adalah 0.005 ± 0.002 (0.001 – 0.015) pada C. alba, dan 0.010 ± 0.002 (0.001-0.010) pada C.
moluccensis.
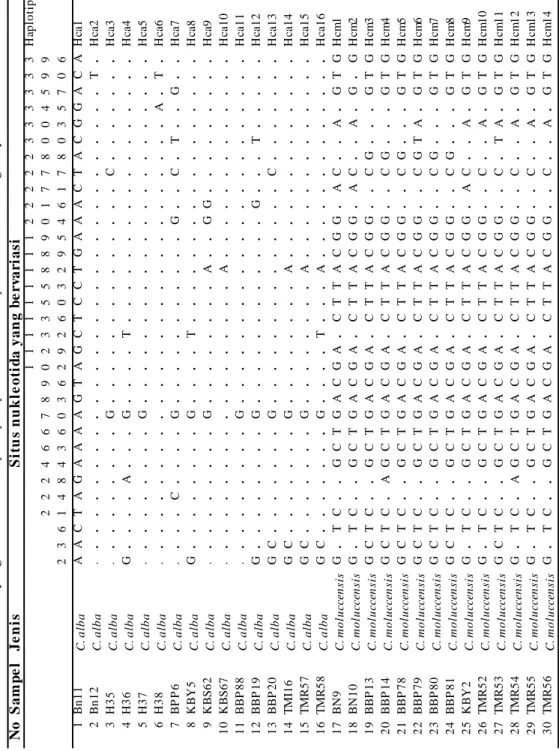
Variasi sekuen DNA, diversitas nukleotida, haplotipe, dan diversitas haplotipe.
Dari semua data sekuen DNA sepanjang 791-bp pada 30 individu kakatua, terjadi substitusi basa pada 63 situs, ini menunjukkan adanya situs yang bervariasi (variable sites) sebesar sekitar 8,8 % . Di antara individu-individu C. alba terdapat 18 (2,42 %) situs yang bervariasi , dan di antara individu-individu
C. moluccensis terdapat 19 (2,44 %)
situs yang bervariasi. Terdapatnya substitusi basa dintara sekuen DNA
cytochrome b ini memperlihatkan adanya
susunan basa yang bervariasi dan juga tipe substitusi basanya, sehingga pada penelitian ini terdeteksi adanya
haplotipe-Positi kodon
C. alba C. moluccensis C. alba + C. moluccensis
ii si sv ii si sv ii si sv Total 785 4 2 783 5 3 763 23 5 Ke-1 263 1 0 262 1 1 259 4 1 Ke-2 263 0 1 263 0 1 261 2 2 Ke-3 259 3 1 258 4 1 243 18 2
Catatan: ii = pasangan identik, si = transisi, sv = transversi.
Table 2 : Rata-rata frekuensi substitusi basa di dalam 791-bp dari cytochrome b pada C. alba dan C. moluccensis
haplotipe dari sekuen DNA gen
cytochrome b, baik dintara maupun
antara jenis kakatua yang diteliti. Jumlah situs yang mengalami subsitusi diantara dua jenis C. alba dan
C. moluccensis relatif sedikit bahkan
kurang dari 10 %. Namun demikian dari sejumlah situs yang bervariasi itu mengahsilkan nilai diversitas nukleotida maupun diversitas haplotipe yang tinggi. Analisis dari seluruh 30 individu (16
C. alba dan 14 C. moluccensis) yang
diteliti secara keseluruhan terdapat 28 haplotipe di dalam 791- bp cytochrome
b. Haplotipe ini meliputi 14 haplotipe (Hca1-Hca14) terdapat pada C. alba, dan 14 haplotipes (Hcm1-Hcm14) pada
C. moluccensis. Tiga individu (H37,
KBS62, dan BBP88) memiliki haplotipe yang sama yaitu Hca5. Ini artinya bahwa haplotipe Hca5 adalah paling dominan dan ketiga individu tersebut memiliki urutan sekuen DNA yang sama atau tidak tejadi substitusi basa diantara ketiganya. Mengejutkan, bahwa di antara individu-individu C. moluceensis, tidak
268
Dwi Astuti
ada satupun individu yang memiliki urutan sekuen DNA yang sama. Sehingga ini memunculkan haplotipe yang berbeda satu dengan lainnya dan tidak ada haplotipe yang dominan pada C.
moluccensis. Situs yang bervariasi,
variasi sekuen DNA, dan gambaran haplotipe disajikan pada Tabel 3.
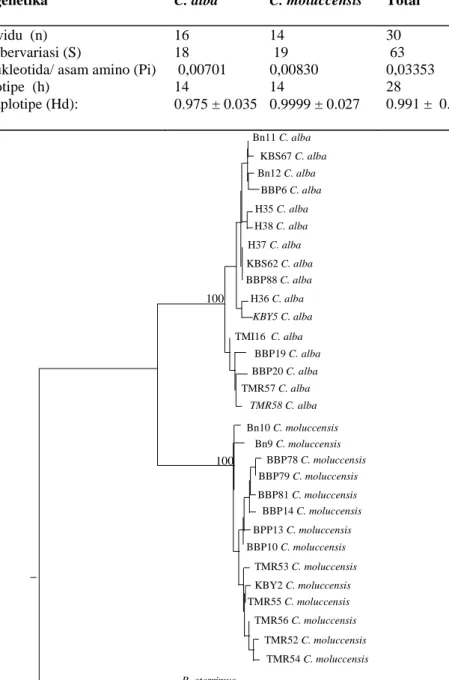
Parameter diversitas meliputi ba-nyaknya variasi, diversitas nukleotida, jumlah dan diversitas haplotipe yang terdapat pada sekuen DNA cytochrome
b pada semua individu (Tabel 4). Tabel 4 menunjukkan, bahwa diversitas nukleotida dan haplotipe dari
C. moluccensis relatif lebih tinggi dari
C. alba.
Analisis Neighbor-joining
Analisis pohon Neighbor-joining (NJ) menghasilkan 2 kelompok yang terpisah secara jelas yaitu: 1) kelompok yang terdiri dari individu-individu C.
alba, dan 2) kelompok individu-individu
C. moluccensis, yang masing-masing
didukung dengan nilai bootstrap 100 %. Kedua jenis dipisahkan dengan divergensi sekuen 6,4 %. Kelompok C.
alba terbagi menjadi 2 kelompok utama,
sedangkan pada C. moluccensis dua individu Bn9 dan Bn10 terpisah dari individu-individu lainnya. Individu-individu
C. moluccensis dengan kode BBP
terpisah dari individu-individu TMR dan KBY, namun pengelelompokan di dalam kelompok C. alba maupun kelompok C.
moluccensis hanya didukung nilai
boostrap kurang dari 50 %. Pohon NJ (Gambar 2) memperlihatkan bahwa individu-individu satu jenis mengelompok pada kelompok yang sama.
DISKUSI
Komposisi basa pada kedua kakatua ini sesuai dengan cytochrome b pada burung lain (e.g Turdus: Pan et al. 2007), vertebrata lain (Prusak & Grzybowski 2004) termasuk ikan (Abol-Munafi et al.
2007). Ciri khas dari gen pengkode protein termasuk cytochrome b adalah komposisi guanin menempati paling rendah dan sitosin yang tertinggi (Avise1994). Seperti yang dikatakan oleh Brown et al. (1982) bahwa biasanya substituti transisi lebih dominan terjadi dibandingan dengan transversi. Basa purin (A ad G) memperlihatkan terjadinya saturasi pada tingkat divergensi yang rendah dibandingkan pirimidin (T dan C), itu menyebabkan frekuensi A dan G menjadi jauh berbeda dari frekuensi C dan T (Kocher & Carleton 1997). Pola substitusi basa dimana substitusi paling banyak terjadi pada posisi codon ketiga, diikuti posisi pertama dan kedua, ini sesuai dengan pola substitusi pada DNA mitikondria khususnya gen cytochrome
b pada beberapa kelompok burung lainnya misalkan Turdus (Pan et al.
2007).
Hasil analisis terhadap jarak genetik menunjukkan dengan pasti, bahwa antara dua jenis yang berbeda memiliki jarak genetik lebih tinggi dibandingkan dengan di antara individu-individu pada jenis yang sama (intraspecies). Rata-rata jarak genetik antara individu pada C.
moluccensis (0.010 ± 0.002) lebih tinggi
dari C. alba (0.005 ± 0.002), dan jarak di antara kedua jenis adalah 0.064 ± 0.088. Jarak genetik pada Oryx 0.062 (Khan et al. 2008). Dilaporkan Dai et
269
Variasi Gen Mitokondria Cytochrome b pada Dua Jenis Burung
T
able 3 :
Situs nukleotida yang bervariasi dan tipe haplotipe di dalam 791-bp sekuen DNA
dari gen cytochrome b
N o S a m p e l Je n is S it u s n u k le o ti d a yan g b e rv ar ia s i 11111111122222333333H ap lot ipe 22 246678902335588901778004599 23614 843603629260329546178035706 1B n 1 1 C . al ba AAC T A GAAAAGT AGC T C C T GAAAC T A C GGAC AH ca 1 2B n 1 2 C . al ba . .... ... T . Hc a2 3H 3 5 C . al ba . .... .... G ... C ... Hc a3 4H 3 6 C . al ba G .... A ... G .... T ... Hc a4 5H 3 7 C . al ba . .... .... G ... Hc a5 6H 3 8 C . al ba . .... ... A . T . Hc a6 7 B PP6 C . al ba . ... C .... G ... G .. C . T .. G .. Hc a7 8K B Y 5 C . al ba G .... .... G .... T ... Hc a8 9K B S 6 2 C . al ba . .... .... G ... A .. GG ... Hc a9 10 K B S 6 7 C . al ba . .... ... A ... Hc a1 0 1 1 BBP 8 8 C . al ba . .... .... G ... Hc a1 1 1 2 BBP 1 9 C . al ba G .... .... G ... G ... T ... Hc a1 2 1 3 BBP 2 0 C . al ba GC ... .... G ... C ... Hc a1 3 14 T M I1 6 C . al ba GC ... .... G ... A ... Hc a1 4 1 5 T MR5 7 C . al ba GC ... .... G ... A ... Hc a1 5 1 6 T MR5 8 C . al ba GC ... .... G .... T ... A ... Hc a1 6 17 B N 9 C. m o lu ccen si s G . TC .. GC T GAC GA . C TTA C G G . AC .. A . GT GH cm 1 18 B N 10 C. m o lu ccen si s G . TC .. GC T GAC GA . C TTA C G G . AC .. A . G . GH cm 2 1 9 BBP 1 3 C. m o lu ccen si s GC T C .. GC T GAC GA . C TTA C G G .. CG ... GT GH cm 3 2 0 BBP 1 4 C. m o lu ccen si s GC T C . A GC T GAC GA . C TTA C G G .. CG ... GT GH cm 4 2 1 BBP 7 8 C. m o lu ccen si s GC T C .. GC T GAC GA . C TTA C G G .. CG ... GT GH cm 5 2 2 BBP 7 9 C. m o lu ccen si s GC T C .. GC T GAC GA . C TTA C G G .. CG T A . GT GH cm 6 2 3 BBP 8 0 C. m o lu ccen si s GC T C .. GC T GAC GA . C TTA C G G .. CG ... GT GH cm 7 2 4 BBP 8 1 C. m o lu ccen si s GC T C .. GC T GAC GA . C TTA C G G .. CG ... GT GH cm 8 25 K B Y2 C. m o lu ccen si s G . TC .. GC T GAC GA . C TTA C G G . AC .. A . GT GH cm 9 2 6 T MR5 2 C. m o lu ccen si s G . TC .. GC T GAC GA . C TTA C G G .. C .. A . GT GH cm 1 0 2 7 T MR5 3 C. m o lu ccen si s GC T C .. GC T GAC GA . C TTA C G G .. C . TA . GT GH cm 1 1 2 8 T MR5 4 C. m o lu ccen si s G . TC . AGC T GAC GA . C TTA C G G .. C .. A . GT GH cm 1 2 2 9 T MR5 5 C. m o lu ccen si s G . TC .. GC T GAC GA . C TTA C G G .. C .. A . GT GH cm 1 3 3 0 T MR5 6 C. m o lu ccen si s G . TC .. GC T GAC GA . C TTA C G G .. C .. A . GT GH cm 1 4
270 Dwi Astuti T abel 3. Lanjutan N o S a m p e l Je n is S it u s n u k le o ti d a yan g b e rv ar ia s i 44444555555555666666666777777H ap lot ipe 23588334567789012234689233478 62939472841324054562864123596 1B n 1 1 C . al ba C C AAT C T T AAAT C AAC GGT AC GC AGT C T AH ca 1 2B n 1 2 C . al ba ... H ca2 3H 3 5 C . al ba ... H ca3 4H 3 6 C . al ba ... H ca4 5H 3 7 C . al ba ... H ca5 6H 3 8 C . al ba ... H ca6 7 B PP6 C . al ba .. C ... H ca7 8K B Y 5 C . al ba T ... T ... H ca8 9K B S 6 2 C . al ba ... H ca9 10 K B S 6 7 C . al ba ... C ... H ca1 0 1 1 BBP 8 8 C . al ba T ... H ca1 1 1 2 BBP 1 9 C . al ba .. C ... G ... T ... H ca1 2 1 3 BBP 2 0 C . al ba .. C ... G ... C ... H ca1 3 14 T M I1 6 C . al ba ... G ... H ca1 4 1 5 T MR5 7 C . al ba .. C ... G ... H ca1 5 1 6 T MR5 8 C . al ba .. C ... G ... H ca1 6 17 B N 9 C. m o lu ccen si s .. TG C T C . GGG .. CC . AA ... T .. AC T . GH cm 1 18 B N 10 C. m o lu ccen si s .. TG C T C . GGG .. CCT A . C ... G . AC T AGH cm 2 1 9 BBP 1 3 C. m o lu ccen si s . TTG C T C . GGGC . CC . A ... GT AC T AGH cm 3 2 0 BBP 1 4 C. m o lu ccen si s . TTG C T C . GGGC . CCT A A C .. TG . AC T AGH cm 4 2 1 BBP 7 8 C. m o lu ccen si s . T T G C T C AGGGC . CC . AA ... TG . AC T AGH cm 5 2 2 BBP 7 9 C. m o lu ccen si s . TTG C T C . GGGC . CC . AA ... TG . AC T AGH cm 6 2 3 BBP 8 0 C. m o lu ccen si s . TTG C T C . GGGC T C C . AA ... T . T A C T AGH cm 7 2 4 BBP 8 1 C. m o lu ccen si s . CT G C T C . GGGC . CC . AA ... TG . AC T . GH cm 8 25 K B Y2 C. m o lu ccen si s .. TG C T C . GGG .. CC . AA ... TG . AC T . GH cm 9 2 6 T MR5 2 C. m o lu ccen si s .. T G C T C AGGG . TC C . AA ... TG . AC T . GH cm 1 0 2 7 T MR5 3 C. m o lu ccen si s . TTG C T C . GGG .. CC . AA ... TG . AC T . GH cm 1 1 2 8 T MR5 4 C. m o lu ccen si s .. TG C T C . GGG .. CC . AAC .. TG . AC T . GH cm 1 2 2 9 T MR5 5 C. m o lu ccen si s .. TG C T C . GGG .. CC . AA ... TG . AC T . GH cm 1 3 3 0 T MR5 6 C. m o lu ccen si s .. TG C T C . GGG .. CC . AAC .. TG . AC T . GH cm 1 4
271
Variasi Gen Mitokondria Cytochrome b pada Dua Jenis Burung
Parameter genetika C. alba C. moluccensis Total
Jumlah individu (n) 16 14 30
Jumlah situs bervariasi (S) 18 19 63
Diversitas nukleotida/ asam amino (Pi) 0,00701 0,00830 0,03353
Jumlah haplotipe (h) 14 14 28
Diversitas haplotipe (Hd): 0.975 ± 0.035 0.9999 ± 0.027 0.991 ± 0.012
Table 4. Parameter-parameter diversitas mitokondria DNA gen cytochrome b di dalam 791-bp pada dua jenis kakatua (C. alba dan C. moluccensis)
TMR54 C. moluccensis TMR53 C. moluccensis KBY2 C. moluccensis TMR55 C. moluccensis TMR56 C. moluccensis TMR52 C. moluccensis BBP81 C. moluccensis BBP14 C. moluccensis BPP13 C. moluccensis BBP10 C. moluccensis Bn9 C. moluccensis BBP78 C. moluccensis BBP79 C. moluccensis Bn10 C. moluccensis Bn11 C. alba H36 C. alba BBP88 C. alba KBS62 C. alba H37 C. alba H38 C. alba H35 C. alba BBP6 C. alba Bn12 C. alba KBS67 C. alba KBY5 C. alba TMI16 C. alba BBP20 C. alba TMR57 C. alba TMR58 C. alba BBP19 C. alba P. aterrimus 100 100
Gambar 2. Pohon Neighbor-joining (NJ) dari 30 individu C. alba dan C. moluccensis yang dikonstruksi berdasarkan substitusi transisi dan transversi pada 791 bp sekuen DNA gen cytochrome b. Nilai-nilai bootsrap di bawah 50 % tidak ditampilkan.
272
Dwi Astuti
al. (2010), bahwa divergensi sekuen
DNA cytochrome b di antara
individu-individu pada jenis burung yang sama (within species) berkisar dari 0 (Parus
dichrous) hingga 2,8 % (Aegithalos
concinnus), dan antara dua jenis dalam
satu marga sebesar 8,10 ± 0.007
(Aegithalos) dan 0.077 ± 0.005 (Parus).
Individu-individu dari populasi yang sama dibedakan oleh 1 % divergensi sekuen, sedangkan divergensi sekuen antara lokasi yang berbeda sebesar 0.25% - 8%, dan antara dua jenis 3 %- 21 %. Divergensi intraspesifik secara ekstrim bervariasi dari 0 hingga 1% pada sebagian besar kasus tetapi bisa lebih tinggi pada bebrapa jenis burung (4.1% antara dua individu S. collaris, dan 5.3%-6.3% di antara S. castaneiventris
(Lijtmaer et al. 2004).
Jumlah situs yang bervariasi diantara individu pada jenis yang sama pada kedua kakatua ini tidak lebih dari 10 %, tetapi kedua kakatua ini memiliki nilai diversitas tinggi. Dilaporkan oleh Germin et al. ( 2007) bahwa di antara populasi burung Alectoris chukar
diversitas haplotipenya tinggi (0,85 %) demikian juga dengan diversitas nukleotidanya di dalam populasi 0,17 % - 0, 25 %. Pada burung Taiko yang berstatus terancam punah juga memiliki diversitas haplotipe (0,68 %) dan nukleotida (0,13 %) (Lawrence & Taylor 2008 ), dan pada mamalia kecil
(Lontra felina) yang terancam punah
dengan diversitas haplotipe 0.86 dan diversitas nukleotida 0.0117 (Valqui et al. 2010). Tingginya nilai diversitas dipengaruhi oleh beberapa faktor misalnya karena seleksi terkadang
mendukung retensi keanekaragaman genetik yang sangat tinggi dalam populasi kecil dan terisolasi (Kaeuffer et al. 2007), kemungkinan kenyataan bahwa ada burung yang belum ditemukan berbiak di tempat lain, sehingga dapat meningkat-kan estimasi ukuran populasi. Keaneka-ragaman genetik relatif tinggi pada jenis langka dapat menunjukkan bahwa penurunan jumlah populasi tersebut terjadi relatif baru-baru ini (Moritz 1994). Tingginya keanekaragaman genetik walaupun secara substansial ada penurunan populasi telah ditunujukkan pada satwa berumur panjang dengan kematangan seksual tertunda seperti burung Taiko (Lawrence & Taylor, 2008), Ornate Box Turtle Terrapene ornate (Kuo &Janzen 2004) ; Orang-utan Pongo pygmaeus (Goossens et al. 2005), Copper Redhorse Moxostoma hubbsi (Lippe et al. 2006). Hal ini menunjukkan bahwa ciri-ciri sejarah kehidupan generasi lama dan kematangan sexual yang tertunda dapat menjadi buffer (penyangga) hilangnya variasi genetik, terutama ketika penurunan terjadi baru-baru ini. Teori genetika populasi memprediksi bahwa severe bottlenecks
tidak dapat secara drastis mengurangi keragaman genetik ketika mereka berakhir dengan jumlah populasi yang kecil (Amos & Balmford 2001). Kakatua juga merupakan burung berumur panjang, bisa hidup mencapai umur 80 tahun dan reproduksi lambat dengan dewasa kelamin dicapai paling cepat pada umur 3-4 tahun (Forshaw 1989), sehingga bisa mengurangi tingkat pergeseran genetik tetapi juga memperlambat pemulihan jumlah populasinya. Jadi wajar jika pada
273
Variasi Gen Mitokondria Cytochrome b pada Dua Jenis Burung
kakatua yang di teliti di sini memiliki nilai diversitas genetik yang tinggi, walau populasi di alam mengalami penurunan. Dari hasil ini diharapkan dapat memberikan informasi dasar dalam program pelestarian burung kakatua baik di alam maupun di penangkaran. Seperti dikatakan oleh Iyengar et al. ( 2007) bahwa perspectif yang memiliki nilai global tentang program penangkaran yang lebih efektif, adalah dengan perlunya mengetahui dan merawat diversitas genetik untuk menjaga jenis-jenis terancam punah, dan dimungkinkan dalam mendukung upaya konservasi ini, karena secara nyata dapat berkontribusi di dalam program penangkaran dan strategi reintroduksi untuk konservasi berbagai satwa yang terancam punah (Russello & Amato 2007).
Secara keseluruhan rata-rata jumlah basa yang bervariasi (k) dan nilai diferensiasi pada C. alba lebih rendah dari C. moluccensis. Demikian juga untuk nilai diversitas nukleotida dan diversitas haplotipe. Pada penelitian ini tidak diketahui lokasi maupun populasi individu-individu tersebut, karena sampel darah di koleksi dari individu-individu yang sudah berada di taman Margasatwa maupun tempat penangkaran. Ini kemungkinan individu-individu berasal dari populasi yang berbeda, bahkan mungkin dari pulau-pulau yang berbeda, atau berkaitan dengan isolasi geografiknya (Randi et al. 2006). Seperti dikatakan oleh Forshaw (1989) bahwa
C. moluccensis hanya tersebar di
Ceram, Saparua and Haruku, dan C. alba terdistribusi Obi, Bacan, Halmahera. Dikatakan bahwa
perbe-daan sejarah populasi dan hidup serta ekologi reproduksi hewan dapat berefek pada kecepatan penurunan dan diversitas genetiknya (Kuo & Janzen 2004).
Nilai divergensi sekuen atau jarak genetik antara individu-individu satu jenis dan antara individu-individu beda jenis kakatua, sangat jelas bedanya. Perbedaan jarak genetik ini berimplikasi pada pengelompokan yang terjadi pada hasil analisis neighbor-joining, dimana individu-individu beda jenis pada penelitian ini berada pada kelompok yang berbeda dan individu-individu sejenis mengelokpok pada kelompok yang sama, seperti ditunjukkan pada pohon NJ. KESIMPULAN
Jumlah situs yang mengalami substitusi basa dan bervariasi pada individu-individu sejenis kurang dari 10 % dari panjang basa cytochrome b yang dianalisis, namun diversitas nukleotida/ asam amino maupun haplotipe pada individu-individu C. alba maupun C.
moluccensis yang diteliti, relatif tinggi.
Analisis neigbor joining memisahkan kelompok antara C. alba dan C.
moluccensis, dengan nilai divergensi
sekuen 6,4 %.
UCAPAN TERIMA KASIH
Terima kasih disampaikan kepada sdr. Agus Kundarmasno dan Alwin Marakamah yang telah membantu dalam proses koleksi sampel darah, staf di Laboratorium Genetika, Bidang Zoologi-Puslit Biologi LIPI atas kerjasamanya. Penelitian ini didukung oleh JICA
274
Dwi Astuti
Konservation Project dan JSPS Core University Project.
DAFTAR PUSTAKA
Abol-Munafi AB, MA. Ambak, P.Ismail, BM. Tam. 2007. Molecular data from the cytochrome b for phylogeny of Channidae (channa sp.) in Malaysia. Biotechnology 6 (1): 22-27.
Amos W, & A. Balmford. 2001. When does conservation genetics matter?
Heredity 87:257– 265
Ardern SL, DM.Lambert 1997. Is the Black Robin in genetic peril? Mol Ecol 6:21–28
Avise JC. 2004. Molecular markers,
natural history, and evolution.
2nd edn. Sinauer Associates, Massa-chusetts
Brown WM, EM. Prager, A. Wang, & AC. Wilson. 1982. Mitochondrial DNA sequences of primates: tempo and mode of evolution. J. Mol.
Evol. 18: 225–239.
Crochet, PA., JD. Lebreton, & F. Bonhomme. 2002. Systematics of large white-headed gulls: patterns of mitochondrial DNA variation in western European taxa. Auk 119: 603-620.
Dai C, K. Chen, R. Zhang, X. Yang, Z. Yin , H. Tian, Z. Zhang, Y. Hu, F. Lei. 2010. Molecular phylogenetic analysis among species of Paridae, Remizidae and Aegithalos based on mtDNA sequences of COI and cyt
b. Chinese Birds 1(2):112–123
Edwards SV, P. Artander, & AC.Wilson. 1991. Mitochondrial resolution of a deep branch in the geneological tree for perching birds. Proceedig Royal Society london. Science 243: 99-107.
Forshaw JM. 1989. Parrots of the World. Tird Edn. Lansdowne Editions, Sydney, Australia.
Garci AG, G. Wlasiuk & EP. Lessa. 2000. High levels of mitochondrial
cytochrome b divergence in annual
killifishes of the genus Cynolebias
(Cyprinodontiformes, Rivulidae).
Zool. J. Linnean Soc.129: 93–110
Gill FB, B. Slkas, FH. Sheldon. 2005. Phylogeny of titmice (Paridae): II. Species relationships based on sequences of the mitochondrial cytochrome-b gene. Auk. 122:121– 143.
Goossens B, L. Chikhi, MF. Jalil, M.Ancrenaz, I. Lackman Ancre-naz, M. Mohamed, P. Andau, & MW. Bruford. 2005. Patterns of genetic diversity and migration in increasingly fragmented and declining Orang-utan (Pongo pygmaeus) populations from Sabah, Malaysia. Mol. Ecol. 14:441–456 Guerrini, M., P. Panayides, P. Hadjigerou,
L.Taglioli, F. Dini,. & F. Barbanera. 2007. Lack of genetic structure of Cypriot Alectoris chukar (Aves, Galliformes) populations as inferred from mtDNA sequencing data. Animal Biodiv. Cons. 30.1: 105–114.
Ingman M, H. Kaessmann, S. Paabo., & U. Gyllensten. 2000. Mitochon-drial genome variation and the
275
Variasi Gen Mitokondria Cytochrome b pada Dua Jenis Burung
origin of modern humans. Nature
408: 708-713.
IUCN (The World Conservation Union). 2006. 2006 IUCN red list of threatened species. http://www. iucnredlist.org, Cited 13 Nov 2006 Iyengar A, T. Gilbert, T. Woodfine, JM. Knowles. 2007. Remnants of ancient genetic diversity preserved within captive groups of scimitar-horned oryx (Oryx dammah). Mol.
Ecol. 16: 2436-2449.
Kaeuffer R, DW. Coltman, JL. Chapuis, D. Pontier, &D. Reale. 2007. Unexpected heterozygosity in an island Mouflon population founded by a single pair of individuals. Proc
Royal. Soc B 274:527–533.
Kocher TD, WK. Thomas, A. Meyer, SV. Edwards, S. Paabo, FX. Villablanca, & AC. Wilson. 1989. Dynamics of mitochondrial DNA evolution in animals: amplification and sequencing with conserved primes. Proc. Nat. Acad. Sci USA.86: 6196–200.
Kocher RD & KL Carleton. 1997. Base substitution in fish mitochondrial DNA: patters and rates. In Molecular systematics of Fish. Kocher, TD and Ca Stepien (Eds.). Academic Press: San Diego. Kuo C-H, & FJ. Janzen. 2004. Genetic
effects of a persistent bottleneck on a natural population of Ornate Box Turtles (Terrapene ornata).
Cons. Gen. 5:425–437
Khan HA, IA. Arif, AA. Al Homaidan, & AH. Al Farhan. 2008. Application of 16S rRNA,
cytochrome b and control region
sequences for understanding the phylogenetic relationships in Oryx
species. Cons.Gen. 9:1293–1301 Kumar S, K. Tamura, & M. Nei. 2004.
MEGA3. Integrated software for molecular evolutionary genetics analysis and sequence alignment.
Briefings in Bioinformatics 5:
150-163
Lijtmaer DA, NMM. Sharp, PL. Tubaro, SC. Lougheed. 2004. Molecular phylogenetics and diversification of the genus Sporophila (Aves: Passeriformes). Mol.Phyl.Evol.
33: 562–579
Lippe C, P. Dumont, L. Bernatchez 2006). High genetic diversity and no inbreeding in the endangered Copper Redhorse, Moxostoma hubbsi (Catostomidae, Pisces): the positive sides of a long generation time. Mol Ecol 15:1769–1780 Martens J, DT. Tietze, & YH. Sun. 2006.
Molecular phylogeny of Parus
(Periparus), a Eurasian radiation
of tits (Aves: Passeriformes: Paridae). Zool. Abh. Mus. Tierkd
Dresden. 55:103–120.
Montgelard C, FM. Catzeflis, E. Douzery. 1997. Phylogenetic relationships of artiodactyls and cetaceans as deduced from comparison of
cytochrome b and 12S rRNA
mitochondrial sequences. Mol. Biol Evol. 14: 550–9.
Moritz, C. 1994 Applications of mitochondrial DNA analysis in conservation: a critical review. Mol
Ecol. 3:401–411
Olavarria C, F. Balbontin, R.Bernal, &C. Baker. 2006. Lack of divergence
276
Dwi Astuti
in the mitochondrial cytochrome b gene between Macruronus
species (Pisces: Merlucciidae) in the Southern Hemisphere. Short communication. New Zealand
J.Mar. Freshwater Res. 40: 299–
304.
Pan QW, FM. Lei, ZH. Yin, A. Kristin, & P. Kanuch. 2007. Phylogenetic realationships within Turdus species: mitochondrial cytochrome-b gene analysis. Ornis Fennica
84: 1-11.
Prusak B, G. Grzybowski, & G. Zieba. 2004 Taxonomic position of Bison
bison (Linnaeus, 1758) and Bison
bonasus (Linnaeus, 1758) based
on analysis of cytb gene. Anim. Sci
Pap Rep. 22: 27–35.
Prusak B & T. Grzybowski. 2004. Non-random base composition in codons of mitochondrial cytochrome b gene in vertebrates. Acta
Biochimica Polonica 51 (4): 897–
905
Randi, E., C. Tabarroni. & S. Kark 2006. The role of history vs. demography in shaping genetic population structure across an ecotone: chukar partridges (Alectoris chukar) as a case study. Diversity and
Distributions, 12: 714–724.
Rocha-Olivares A, CA. Kimbrell, BJ. Eitner & RD. Vetter. 1999. Evolution of a Mitochondrial
Cytochrome b Gene Sequence in
the Species-Rich Genus Sebastes
(Teleostei, Scorpaenidae) and Its Utility in Testing the Monophyly of the Subgenus Sebastomus. Molecular Phylogenetics and
Evolution 11 (3): 426–440.
Roques S, & JJ. Negro. 2005. MtDNA genetic diversity and population history of a dwindling raptorial bird, the Red Kite (Milvus milvus). Biol
Cons. 126:41–50
Russello MA & G. Amato. 2007. On the horns of a dilemma: molecular approaches refine ex situ
conservation in crisis. Mol. Ecol.
16: 2405-2406.
Spielman D, BW. Brook, & R. Frankham. 2004. Most species are not driven to extinction before genetic factors impact them. Proc.
Natl Acad Sci USA 101:15261–
15264
Saitou N & M. Nei. 1987 The neighbor-joining method: A new method for reconstructing phylogenetic trees.
Mol Biol Evol 4: 406–425
Whittier JB, JR. Leslie. RA.van Den Bussche. 2006. Genetic variation among subspecies of Least Tern (Sterna antillarum): implications for conservation. Waterbirds 29:176– 184.
Memasukkan: Desember 2010 Diterima: Mei 2011
J. Biol. Indon. Vol 7, No.2 (2011)
PANDUAN PENULIS
Naskah dapat ditulis dalam bahasa Indonesia atau bahasa Inggris. Naskah disusun dengan urutan: JUDUL (bahasa Indonesia dan Inggris), NAMA PENULIS (yang disertai dengan alamat Lembaga/ Instansi), ABSTRAK (bahasa Inggris, maksimal 250 kata), KATA KUNCI (maksimal 6 kata), PENDAHULUAN, BAHAN DAN CARA KERJA, HASIL, PEMBAHASAN, UCAPAN TERIMA KASIH (jika diperlukan) dan DAFTAR PUSTAKA.
Naskah diketik dengan spasi ganda pada kertas HVS A4 maksimum 15 halaman termasuk gambar, foto, dan tabel disertai CD. Batas dari tepi kiri 3 cm, kanan, atas, dan bawah masing-masing 2,5 cm dengan program pengolah kata Microsoft Word dan tipe huruf Times New Roman
berukuran 12 point. Setiap halaman diberi nomor halaman secara berurutan. Gambar dalam bentuk grafik/diagram harus asli (bukan fotokopi) dan foto (dicetak di kertas licin atau di scan). Gambar dan Tabel di tulis dan ditempatkan di halam terpisah di akhir naskah. Penulisan simbol α, β, χ, dan lain-lain dimasukkan melalui fasilitas insert, tanpa mengubah jenis huruf. Kata dalam bahasa asing dicetak miring. Naskah dikirimkan ke alamat Redaksi sebanyak 3 eksemplar (2 eksemplar tanpa nama dan lembaga penulis).
Penggunaan nama suatu tumbuhan atau hewan dalam bahasa Indonesia/Daerah harus diikuti nama ilmiahnya (cetak miring) beserta Authornya pada pengungkapan pertama kali.
Daftar pustaka ditulis secara abjad menggunakan sistem nama-tahun. Contoh penulisan pustaka acuan sebagai berikut :
Jurnal :
Hara, T., JR. Zhang, & S. Ueda. 1983. Identification of plasmids linked with polyglutamate production in B. subtilis. J. Gen. Apll. Microbiol. 29: 345-354.
Buku :
Chaplin, MF. & C. Bucke. 1990. Enzyme Technology. Cambridge University Press. Cambridge. Bab dalam Buku :
Gerhart, P. & SW. Drew. 1994. Liquid culture. Dalam : Gerhart, P., R.G.E. Murray, W.A. Wood, & N.R. Krieg (eds.). Methods for General and Molecular Bacteriology. ASM., Washington. 248-277.
Abstrak :
Suryajaya, D. 1982. Perkembangan tanaman polong-polongan utama di Indonesia. Abstrak Pertemuan Ilmiah Mikrobiologi. Jakarta . 15 –18 Oktober 1982. 42.
Prosiding :
Mubarik, NR., A. Suwanto, & MT. Suhartono. 2000. Isolasi dan karakterisasi protease ekstrasellular dari bakteri isolat termofilik ekstrim. Prosiding Seminar nasional Industri Enzim dan Bioteknologi II. Jakarta, 15-16 Februari 2000. 151-158.
Skripsi, Tesis, Disertasi :
Kemala, S. 1987. Pola Pertanian, Industri Perdagangan Kelapa dan Kelapa Sawit di Indonesia.[Disertasi]. Bogor : Institut Pertanian Bogor.
Informasi dari Internet :
Schulze, H. 1999. Detection and Identification of Lories and Pottos in The Wild; Information for surveys/Estimated of population density. http//www.species.net/primates/loris/ lorCp.1.html.
J. Biol. Indon. Vol 7, No. 2 (2011)
Kajian Pendahuluan: Perpindahan Gen dari Tanaman Kentang Transgenik Katahdin RB ke Tanaman Kentang Non Transgenik
A.Dinar Ambarwati, M. Herman, Agus Purwito , Eri Sofiari,& Hajrial Aswidinoor
277
Virus Influenza Novel H1N1 Babi di Indonesia
NLP Indi Dharmayanti, Atik Ratnawati, & Dyah Ayu Hewajuli
289
Karakterisasi Produk Biosolubilisasi Lignit oleh Kapang Indigenus dari Tanah Pertambangan Batubara di Sumatera Selatan
Irawan Sugoro, Sandra Hermanto,D. Sasongko,D. Indriani & P. Aditiawati
299
Potensi Virus Avian Influenza H5NI Isolat A/Ck/West Java/Pwt-Wij/2006 Sebagai Vaksin
R. Indriani, NLP I Dharmayanti, R.M.A Adjid, & Darminto
309
Variasi dan kekerabatan genetik pada dua jenis baru belimbing (Averrhoa leucopetala Rugayah et Sunarti sp nov dan A. dolichorpa Rugayah et Sunarti sp nov., Oxalidaceae) berdasarkan profil Random Amplified Polymorphic DNA Kusumadewi Sri Yulita
321
Pengaruh Dinamika Faktor Lingkungan Terhadap Sebaran Horisontal dan Vertikal Katak Hellen Kurniati
331 Merekonstruksi Habitat Curik Bali Leucopsar rothschildi Stresemann, 1912 di Bali
Bagian Barat
Mas Noerdjito, Roemantyo &Tony Sumampau
341
Struktur dan Komposisi Vegetasi Hutan Semusim Habitat Curik Bali (Leucopsar rothschildi Stresemann, 1912) di Kawasan Labuan Lalang, Taman Nasional Bali Barat
Roemantyo
361
Sumbangan Ilmu Etnobotani dalam Memfasilitasi Hubungan Manusia dengan Tumbuhan dan Lingkungannya
Eko Baroto Walujo