CITATIONS
2
READS
23,718
2 authors, including:
Eva Yuniritha
Ministry of Health, Indonesia 5PUBLICATIONS 16CITATIONS
SEE PROFILE
All content following this page was uploaded by Eva Yuniritha on 13 November 2021.
The user has requested enhancement of the downloaded file.
DAFTAR ISI
DAFTAR ISI ... 1
DAFTAR GAMBAR ... 4
BAB I METABOLISME UMUM ... 5
A. PENGANTAR ... 5
B. PENCERNAAN ... 5
C. PENYERAPAN ... 6
D. Metabolisme Energi ... 7
BAB II METABOLISME NUTRIEN ... 9
A. METABOLISME MAKRO NUTRIEN ... 9
1. Metabolisme Karbohidrat ... 9
a. Prinsip ... 9
b. Proses Pada Metabolisme Karbohidrat ... 9
1) Glikolisis ... 9
2) Glikogenesis ... 12
3) Glikogenolisis ... 13
4) Pembentukan Asetil- KoA ... 14
5) Siklus Asam Sitrat ... 14
6) Glukoneogenesis ... 16
7) Pengaturan Glukoneogenesis ... 18
8) Siklus Cori ... 18
c. Gangguan Metabolisme Karbohidrat pada Kesehatan ... 19
2. Metabolisme Protein ... 20
a. Prinsip ... 43
b. Proses Pada Metabolisme Protein ... 43
1) Transpor Asam Amino ke dalam Sel ... 43
2) Proses Reaksi Transaminasi ... 44
3) Pengeluaran Nitrogen Asam Amino Sebagai Amonia ... 44
4) Siklus Urea ... 45
5) Fungsi Siklus Urea Selama Puasa ... 46
6) Biosintesis Asam Amino ... 47
7) Katabolisme Asam Amino ... 49
8) Asam Amino yang Berperan dalam Siklus Krebs ... 50
c. Energi yang dihasilkan ... 55
d. Gangguan Metabolisme Protein pada Kesehatan ... 55
3. Metabolisme Lemak ... 57
B. METABOLISME MIKRO NUTRIEN (VITAMIN DAN MINERAL) ... 80
I. Vitamin ... 20
a. Vitamin Larut dalam Lemak ... 21
1) Vitamin A ... 21
2) Vitamin D ... 23
3) Vitamin E (Tokoferol) ... 26
4) Vitamin K ... 28
b. Vitamin Larut dalam Air ... 30
1) Vitamin B Kompleks ... 31
2) Vitamin C (Asam askorbat) ... 41
II. Mineral ... 80
a. Makro Mineral ... 81
1) Calsium (Ca) ... 81
2) Fosfor (P) ... 83
3) Natrium (Na) ... 84
4) Kalium (K) ... 84
5) Chlor (Cl) ... 85
6) Magnesium (Mg) ... 86
7) Sulfur (S) ... 86
a. Mikro Mineral ... 87
1) Fe ( Fe ) ... 87
2) Iodium (I) ... 97
3) Zink (Zn) ... 99
4) Kobalt (CO) ... 104
5) Mangan (Mn) ... 104
6) Tembaga (Cu) ... 105
7) Molibdenum (Mo) ... 106
8) Selenium (Se) ... 107
9) Kromium (Cr) ... 108
10) Vanadium (V) ... 109
11) Flourin (F) ... 109
12) Silikon (Si) ... 110
13) Timah (Pb) ... 111
14) Nikel (Ni) ... 111
15) Arsen (As) ... 112
DAFTAR GAMBAR
BAB I
METABOLISME UMUM
A. PENGANTAR
Semua organisme hidup mentransformasi energi yang diambil dari sekelilingnya. Energi ini dibutuhkan untuk sintesis makromolekul yang akan digunakan untuk pertumbuhan dan diferensiasi organisme tersebut. Transformasi ini dicapai melaui reaksi sejumlah besar enzim yang mengkatalisis jaringan kompleks kumpulan reaksi kimia yang dikenal sebagai metabolisme (Ngili, 2009).
Metabolisme merupakan segala reaksi kimia yang terjadi didalam makhluk hidup, mulai dari makhluk bersel satu yang sangat sederhana seperti bakteri, protozoa, jamur, tumbuhan, hewan, sampai kepada manusia yang susunan tumbuhannya sangat kompleks. Didalam proses ini, makhluk hidup mendapat, mengubah, dan memakai senyawa kimia dari sekitarnya untuk mempertahankan kelangsungan hidupnya.
Metabolisme meliputi proses sistesis dan proses penguraian senyawa atau komponen dalam sel hidup. Proses sintesis itu disebut anabolisme dan proses penguraian disebut katabolisme. Semua reaksi metabolisme dikatalisis oleh enzim, termasuk reaksi yang sederhana seperti penguraian asam karbonat menjadi air dan karbondioksida, proses pemasukan dan pengeluaran zat kimia dari dan kedalam selmelalui membran; proses biosintesis protein yang panjang dan rumit ; atau pun proses penguraian bahan makanan dalam sistem pencernaan mulai dari mulut, lambung, dan penyerapan hasil penguraian tersebut melalui dinding usus, serta penyebarannya ke seluruh bagian tubuh yang memerlukannya.Hal lain yang penting dari metabolisme adalah peranannya dalam proses pengawaracunan atau detoksifikasi, yaitu mekanisme reaksi pengubahan zat yang beracun menjadi senyawa tak beracun yang dapat dikeluarkan dari tubuh.
Anabolisme dibedakan dari katabolisme dalam beberapa hal: anabolisme merupakan proses sintesis molekul kimia kecil menjadi molekulyang lebih besar, sedangkan katabolisme adalah sebaliknya, yaitu proses penguraian molekul besar menjadi molekul kecil; anabolisme adalah proses yang membutuhkan energi sedangkan katabolisme melepaskan energi; anabolisme merupakan reaksi reduksi, sedangkan katabolisme adalah reaksi oksidasi; seringkai hasil akhir anabolisme merupakan senyawa pemula untuk proses katabolisme. Sebagian besar proses metabolisme terjadi didalam sel, oleh karena itu mekanisme masuk dan keluarnya zat kimia melalui membran sel mempunyai arti pentingdalam mempertahankan keseimbangan energi dan materi di dalam tubuh (Wirahadikusumah, 1985).
B. PENCERNAAN
Pada umumnya makanan yang masuk ke mulut kita masih berbentuk potongan yang masih mempunyai ukuran relatif besar, misalnya nasi, potongan kentang, potongan daging, telur, potongan sayuran atau buah-buahan. Makanan
ini untuk dapat ditelan, perlu mengalami perubahan bentuk maupun ukurannya, yaitu diubah menjadi potongan dengan ukuran yang lebih kecil. Secara garis besar makanan kita terdiri dari atas karbohidrat, lemak, protein, mineral, vitamin dan air.
Untuk dapat digunakan sebagai sumber energi, pemeliharaan dan pertumbuhan bagi tubuh, berbagai makanan tersebut diubah dahulu menjadi molekul-molekul yang dapat masuk kedalam sel-sel dan mengalami berbagai reaksi kimia yang penting. Pengubahan makanan dari sejak awal hingga menjadi berbentuk molekular yang siap untuk diserap melalui dinding usus disebut pencernaan makanan dan proses ini berlangsung dalam sistem pencernaan makanan yang terdiri atas beberapa organ tubuh yaitu mulut, lambung dan usus dengan bantuan pankreas dan empedu (Poedjiadi, 1994).
Gambar 1. Organ Pencernaan Pada Manusia Sumber. Poedjiadi, Dasar-dasar Biokimia 1994, hlm. 234
Polisakarida atau karbohidrat yang merupakan salah satu komponen bahan makanan, diubah menjadi monosakrida, protein diubah menjadi asam amino, sedangkan lemak diubah menjadi asam lemak dan gliserol. Dengan demikian, pencernaan pada hakekatnya mempunyai peranan menghasilkan molekul yang cukup kecil untuk dapat diserap oleh tubuh melalui dinding usus halus, untuk selanjutnya dibawa oleh aliran darah untuk disebarkan ke seluruh bagian tubuh yang memerlukan (Wirahadikusumah, 1985).
C. PENYERAPAN
Penyerapan atau absorpsi adalah suatu proses masuknya zat makanan ke dalam darah dan limpa melalui dinding usus halus. Usus halus merupakan tempat
utama terjadinya proses penyerapan. Mekanisme penyerapan usus halus adalah khas, sebagian besar molekul dapat diserap dan sebagian lain lagi tidak. Molekul yang masuk kedalam tubuh, yang diserap melalui dinding usus halus, selanjunya mengalami salah satu dari dua macam reaksi metabolisme : reaksi katabolisme dan anabolisme. Reaksi anabolisme membutuhkan energi disebut reaksi endergonik, sedangkan katabolisme menghasilkan energi disebut reaksi eksorgonik. Zat-zat gizi seperti karbohidrat diserap dalam bentuk monosakarida, protein dalam bentuk asam amino, sedangkan lemak pada waktu penyerapan mngalami proses sintesis ulang menjadi lemak yang kemudian masuk kedalam aliran darah dan limpa (Wirahadikusumah, 1985).
Selama penyerapan zat-zat yang memasuki tubuh berlangsung dengan dua langkah; pertama, memasuki mukosa kemudian darah atau limfa; kedua, dikontrol sesuai dengan yang dibutuhkan oleh tubuh. Sel-sel yang melapisi saluran pencernaan mengelupas dan diganti dalam 3-5 hari. Zat-zat yang terperangkap dalam sel-sel tersebut dan tidak diserap ke dalam tubuh akan hilang bersama sel- sel yang mengelupas.
D. Metabolisme Energi
Metabolisme energi berasal dari hasil fotosintesis tumbuhan yang yang ditangkap dalam ikatan kimia molekul-molekul karbohidrat, protein, lemak dan alkohol. Hasil akhir metabolisme zat gizi : CO2, HO2. ATP. Energi yang digunakan sel dalam bentuk ATP.
1. Prinsip metabolisme
Dari total pembakaran berbagai bahan makanan karbohidrat dan lemak secara sempurna dioksidasi menjadi CO2 dan air, dan semua energi dilepas dalam bentuk panas. Dalam tubuh, proses oksidasi persis sama yaitu 1 molekul glukosa dan 6 molekul oksigen dikonversi menjadi 6 molekul CO2 dan 6 molekul air (H2O); pada trigliserida (merupakan 95%-98% lemak makanan) di konversi menjadi CO2 dan H2O (dengan jumlah molekulnya masing-masing spesifik).
2. Sumber metabolisme
Sumber metabolisme karbohidrat berasal dari serelia dan umbi-umbian, sumber metabolisme protein berasal dari daging, kacang-kacangan ikan dan sumber metabolisme lemak berasal dari bahan makanan tumbuhan dan hewan termasuk telur dan susu.
3. Kegunaan/ manfaat
Metabolisme energi kimia dalam sel sebagai biosintesis, kontraksi dan gerakan, transpor aktif, dan transfer bahan genetik.
4. Energi yang dihasilkan adalah ATP (Adenosin Tri Fosfat).
5. Energi berasal dari makanan dan energi yang diektrak dari makanan digunakanan untuk memberi energi gugus fosfat agar dapat membentuk ATP.
6. Anabolisme dan katabolisme a. Anabolisme
Anabolisme adalah proses reaksi molekul zat gizi menjadi bagian dari molekul komplek yang diperlukan oleh tubuh. Reaksi ini disebut endegronic karena memerlukan energi agar proses berlangsung.
b. Katabolisme
Katabolisme adalah reaksi oksidasi (pembakaran zat gizi) pembebasan energi yang memenuhi kebutuhan tubuh dengan segera. Atau reaksi pembebasan energi exergonic yang berlangsung tanpa membutuhkan energi (Murray, dkk, 2003).
7. Interaksi
Di dalam tubuh manusia terjadi metabolisme karbohidrat, protein dan lemak. Untuk menghasilkan energi yang berasal dari katabolisme karbohidrat, protein maupun lemak. Titik temu dari berbagai jalur metabolisme ini berguna untuk saling menggantikan bahan bakar di dalam sel. Hasil katabolisme karbohidrat, protein dan lemak juga bermanfaat untuk menghasilkan senyawa-senyawa lain yaitu dapat membentuk ATP, hormon, komponen hemoglobin ataupun komponen sel lainnya. Lemak (asam heksonoat) lebih banyak mengandung hydrogen terkait dan merupakan senyawa karbon yang paling banyak tereduksi, sedangkan karbohidrat (glukosa) dan protein (asam glutamat) banyak mengandung oksigen dan lebih sedikit hydrogen terikat adalah senyawa yang lebih troksidasi. Senyawa karbon yang tereduksi lebih banyak menyimpan energi dan apabila ada pembakaran sempurna akan membebaskan energi lebih banyak karena adanya pembebasan elektron yang lebih banyak. Jumlah elektron yang dibebaskan menunjukkan jumlah energi yang dihasilkan.
Perlu Anda ketahui pada jalur katabolisme yang berbeda glukosa dan asam glutamat dapat menghasilkan jumlah ATP yang sama yaitu 36 ATP.
Sedangkan katabolisme asam heksanoat dengan jumlah karbon yang sama dengan glukosa (6 karbon) menghasilkan 44 ATP, sehingga jumlah energi yang dihasilkan pada lemak lebih besar dibandingkan dengan yang dihasilkan pada karbohidrat dan protein. Sedangkan jumlah energi yang dihasilkan protein setara dengan jumlah yang dihasilkan karbohidrat dalam berat yang sama (Campbell, dkk, 2003).
BAB II
METABOLISME ZAT GIZI
A. METABOLISME ZAT GIZI MAKRO
1. Metabolisme Karbohidrat a. Prinsip
Glukosa merupakan pusat dari semua metabolisme. Glukosa adalah bahan bakar universal bagi sel manusia dan merupakan sumber karbon untuk sintesis sebagian besar senyawa lainnya. Semua jenis sel manusia menggunakan glukosa untuk memperoleh energi. Gula lain dalam makanan (terutama fruktosa dan galaktosa) diubah menjadi glukosa atau zat antara dalam metabolisme glukosa.
b. Proses Pada Metabolisme Karbohidrat 1) Glikolisis
Proses glikolisis ialah proses awal dari metabolisme gugus gula hasil pemecahan karbohidrat di dalam sel. Proses glikolisis bertujuan untuk menghasilkan piruvat dalam keadaan aerob ataupun laktat dalam keadaan anaerob sehingga dapat terbentuk energi. Glikolisis terjadi di dalam sitoplasma sel/sitosol. Pada keadaan aerob, 1 molekul glukosa yang melalui proses glikolisis dapat menghasilkan 8 ATP sedangkan dalam keadaan anaerob jumlah ATP yang dihasilkan lebih sedikit yaitu 2 ATP. Di eritrosit, proses glikolisis selalu terjadi dalam keadaan anaerob karena ketiadaan mitokondria. Hal ini menyebabkan hasil akhirnya selalu berupa laktat (Harjasasmita, 2003 dan Murray, dkk, 2009).
Proses glikolisis terjadi melalui tahapan-tahapan tertentu dan membutuhkan peran enzim tertentu. Tahapan-tahapan tersebut adalah:
1. Tahap pertama proses glikolisis adalah pengubahan glukosa menjadi glukosa-6 fospat dengan reaksi fosforilasi dibantu dengan enzim heksokinase.
Gambar 2. Pengubahan glukosa menjadi glukosa- 6 fospat Sumber. Poedjiadi, Dasar-dasar Biokimia 1994, hlm. 249
2. Tahap kedua ialah isomerisasi, yaitu pengubahan glukosa-6 fospat menjadi fruktosa-6-fosfat, dengan enzim fosfoglukoisomerase. Enzim ini tidak memerlukan kofaktor.
Gambar 3. Pengubahan glukosa- 6 fosfat menjadi fruktosa-6-fosfat Sumber. Poedjiadi, Dasar-dasar Biokimia 1994, hlm. 249 3. Tahap ketiga ialah fruktosa -6-fosfat diubah menjadi fruktosa-1,6-bisfosfat
oleh enzim fosfofruktokinase dibantu oleh ion Mg++ sebagai kofaktor.
Gambar 4. Pengubahan fruktosa-6-fosfat menjadi fruktos1,6-disfosfat Sumber. Poedjiadi, Dasar-dasar Biokimia 1994, hlm. 250
4. Tahap keempat dalam rangkaian reaksi glikolisis adalah penguraian molekul fruktosa-1,6-disfosfat membentuk dua molekul triosa fosfat, yaitu dihidroksi aseton fosfat dan D-gliseral-dehida-3-fosfat. Dalam tahap ini enzim aldolase yang menjadi katalis (Poedjiadi, Anna, 1994).
Gambar 5. Penguraian molekul fruktosa-1,6-difosfat Sumber. Poedjiadi, Dasar-dasar Biokimia 1994, hlm. 251
5. Tahap kelima ialah Gliseraldehid 3-P diubah menjadi 1,3 bisfosfogliserat (gliseraldehid 3-P Dehidrogenase). Proses ini memerlukan koenzim NAD+ yang akan bereaksi dengan phospat inorganik menjadi NADH dan melepas
ion hidrogen. Proses ini akan menghasilkan 3 ATP melalui rantai pernapasan. Proses ini dapat dihambat oleh iodoasetat.
Gambar 6. Pengubahan gliseraldehid 3-P menjadi 1,3 bisfosfogliserat Sumber. Poedjiadi, Dasar-dasar Biokimia 1994, hlm. 252
6. Tahap keenam ialah 1,3 bisfosfogliserat diubah menjadi 3 fosfogliserat (fosfogliserat kinase), dengan bantuan ion magnesium, proses ini akan menghasilkan 1 ATP pada tingkat substrat.
7. Tahap ketujuh ialah 3 fosfogliserat diubah menjadi 2 fosfogliserat (mutase)
Gambar 7. Pengubahan 1,3 bisfosfat menjadi 2 fosfogliserat Sumber. Poedjiadi, Dasar-dasar Biokimia 1994, hlm. 253 8. Tahap kedelapan ialah 2 fosfogliserat diubah menjadi fosfoenol piruvat
(enolase), memerlukan ion magnesium dan akan dihambat oleh flourida.
Gambar 8. Pengubahan 2 fosofogliserat menjadi fosfoenol piruvat Sumber. Poedjiadi, Dasar-dasar Biokimia 1994, hlm. 254
9. Tahap kesembilan ialah fosfoenol piruvat diubah menjadi (enol) piruvat (piruvat kinase), proses ini memerlukan ion magnesium dan ADP. Gugus phospat dari phospo enol piruvat akan diambil untuk bergabung dengan ADP membentuk 1 molekul ATP.
10. Tahap kesepuluh, reaksi yang menggunakan enzim laktat dehidrogenase ini adalah reaksi tahap akhir glikolisis, yaitu pembentukan asam laktat
dengan cara reduksi asam piruvat. Dalam reaksi ini digunakan NADH sebagai koenzim.
Gambar 9. Pembentukan asam laktat
Sumber. Poedjiadi, Dasar-dasar Biokimia 1994, hlm. 255
2) Glikogenesis
Tahap pertama metabolisme karbohidrat adalah pemecahan glukosa (glikolisis) menjadi piruvat. Selanjutnya, piruvat dioksidasi menjadi asetil-KoA.
Akhirnya asetil-KoA masuk ke dalam rangkaian siklus asam sitrat untuk dikatabolisir menjadi energi. Proses diatas terjadi jika kita membutuhkan energi, misalnya untuk berpikir, mencerna makanan, bekerja dan sebagainya.
Jika jumlah glukosa melebihi kebutuhan maka dirangkai menjadi glikogen untuk cadangan makanan melalui proses glikogenesis.
Glikogen merupakan simpanan karbohidrat dalam tubuh dan analog dengan amilum pada tumbuhan. Glikogen terdapat didalam hati (sampai 6%) dan otot jarang melampaui jumlah 1%. Tetapi, karena massa otot jauh lebih besar daripada hati maka besarnya simpanan glikogen di otot jarang bisa mencapai tiga sampai empat kali lebih banyak. Glikogen otot adalah heksosa untuk proses glikolisis di dalam otot itu sendiri. Sedangkan, glikogen hati adalah simpanan sumber heksosa untuk dikirim keluar guna mempertahankan kadar glukosa darah, khususnya di antara waktu makan. Setelah 12-18 jam puasa, hampir semua simpanan glikogen hati terkuras. Tetapi glikogen otot hanya terkuras setelah seseorang melakukan olahraga yang berat dan lama.
Rangkaian proses terjadinya glikogenesis sebagai berikut :
1. Glukosa mengalami fosfolirasi menjadi glukosa 6-fosfat (reaksi yang lazim terjadi juga pada lintasan glikolisis). Di otot reaksi ini dikatalisir oleh heksokinase sedangkan di hati oleh glukokinase.
2. Glukosa 6-fosfat diubah menjadi glukosa 1-fosfat dalam reaksi dengan bantuan katalisator enzim fosfoglukomutase. Enzim itu sendiri akan mengalami fosforilasi dan gugus fosfo akan mengambil bagian di dalam reaksi reversible yang intermesiatnya adalah glukosa 1,6-bifosfat.
Enz-P + Glukosa 6-fosfat ↔ Enz + Glukosa 1,6-bifosfat ↔ Enz-P + Glukosa 1-fosfat 3. Selanjutnya glukosa 1,6-fosfat bereaksi dengan uridin trifosfat (UTP)
untuk membentuk uridin difosfat glukosa (UDPGIc). Reaksi ini dikatalisir oleh enzim UDPGIc pirofosforilase.
UTP + Glukosa 1-fosfat ↔ UDPGIc + PPi
4. Hidrolisis pirofosfat inorganik berikutnya oleh enzim pirofosfatase inorgnik akan menarik reaksi kearah kanan persamaan reaksi.
5. Atom C1 pada glukosa yang diaktifkan oleh UDPGIc membentuk ikatan glikosidik dengan atom C4 pada residu glukosa terminal glikogen, sehingga membebaskan uridin difosfat. Reaksi ini dikatalisir oleh enzim glikogen sintase. Molekul glikogen yang sudah ada sebelumya (disebut glikogen primer) harus ada untuk memulai reaksi ini. Glikogen primer selanjutnya dapat terbentuk pada primer protein yang dikenal sebagai glikogenin.
UDPGIc + (C6)n → UDP + (C6) n+1
Residu glukosa yang lebih lanjut melekat pada posisi 1 → 4 untuk membentuk rantai pendek yang diaktifkan oleh glikogen sintase. Pada otot rangka glikogenin tetap melekat pada pusat molekul glikogen, sedangkan di hati terdapat jumlah molekul glikogen yang melebihi jumlah molekul glikogenin.
6. Setelah rantai dari glikogen primer diperpanjang dengan penambahan glukosa tersebut hingga mencapai minimal 11 residu glukosa, maka enzim pembentuk cabang memindahkan bagian dari rantai 1 → 4 (panjang minimal 6 residu glukosa) pada rantai yang berdekatan untuk rangkaian 1
→ 6 sehingga membuat titik cabang pada molekul tersebut. Cabang- cabang ini akan tumbuh dengan penambahan lebih lanjut 1 → glukosil dan pembentukan cabang selanjutnya. Setelah jumlah residu terminal yang non reduktif bertambah, jumlah total reaktif dalam molekul akan meningkat sehingga akan mempercepat glikogenesis maupun glikogenolisis.
3) Glikogenolisis
Jika glukosa dari diet tidak dapat mencukupi kebutuhan, maka glikogen harus dipecah untuk mendapatkan glukosa sebagai sumber energi. Proses ini digunakan glikogenolisis. Glikogenolisis seakan-akan kebalikan dari glikogenesis, akan tetapi sebenarnya tidak demikian. Untuk memutuskan ikatan glukosa satu demi satu dari glikogen diperlukan enzim fosforilase.
Enzim ini spesifik untuk proses fosforolisis rangkaian 1→ 4 glikogen untuk menghasilkan glukosa 1-fosfat. Residu glukosil terminal pada rantai paling luar molekul glikogen dibuang secara berurutan sampai kurang lebih ada 4 buah residu glukosa yang tersisa pada sisi cabang 1→ 6.
Glukan transferase dibutuhkan sebagai katalisator pemindahan unit trisakarida dari satu cabang ke cabang lainnya sehingga membuat titik cabang 1→ 6 terpanjang. Hidrolisis ikatan 1→ 6 memerlukan kerja enzim pemutus cabang (debranching enzyme) yang spesifik. Dengan pemutusan cabang tersebut, maka kerja enzim fosforilasi selanjutnya dapat berlangsung.
4) Pembentukan Asetil- KoA
Piruvat yang telah terbentuk sebagai hasil proses glikolisis dapat masuk ke dalam mitokondria untuk mengalami oksidasi menjadi molekul asetil koA. 1 molekul glukosa akan menghasilkan 2 molekul piruvat yang memiliki 3 atom karbon. Piruvat akan diubah menjadi asetil koA yang memiliki 2 atom karbon.
Dalam eritrosit, setelah mengalami glikolisis maka piruvat akan diubah menjadi laktat (Murray, dkk, 2009).
Gambar 10. Pembentukan asetil-KoA Sumber. Biology Concepts dan Connection, 2008
Piruvat dehidrogenase ialah enzim yang berperan dalam proses ini.
Konsentrasi dari piruvat dehidrogenase meningkat pada saat makan dan saat piruvat banyak terbentuk. Sebaliknya kondisi kelaparan serta konsentrasi asetil koA yang meningkat akan menghambat kerja dari piruvat dehidrogenase.
Selain itu kinase spesifik juga berperan dalam proses oksidasi piruvat.
Fosforilasi kinase dapat menghambat aktivitas enzim ini, sedangkan defosforilasi kinase dapat mempercepat kerja enzim ini. Enzim ini memerlukan koenzim NAD+ yang melalui rantai pernapasan akan berubah menjadi NADH dan menghasilkan 3 ATP.
Proses reaksi memerlukan 5 vitamin dalam bentuk koenzim, yaitu vitamin asam lipoat, vitamin B1, B2, B5 dan vitamin asam pantotenat. Sedangkan hambatan pada enzim piruvat dehidrogenase dapat menyebabkan laktat asidosis. Kondisi ini dapat terjadi pada keracunan ion merkuri dan pada penderita diabetes melitus (Harjasasmita, 2003). Jumlah ATP yang dihasilkan pada proses ini ialah sebesar 6 ATP.
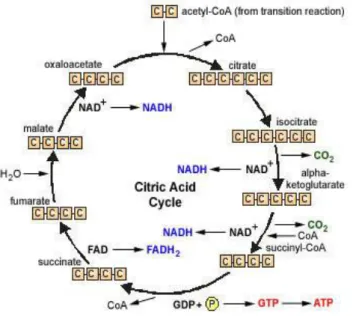
5) Siklus Asam Sitrat
Siklus asam sitrat adalah serangkaian reksi kimia dalam sel yaitu pada mitokondria yang berlangsung secara berurutan dan berulang, bertujuan mengubah asam piruvat menjadi CO2, H2O dan sejumlah energi. Proses ini adalah proses oksidasi dengan menggunakan oksigen atau aerob. Siklus asam sitrat ini disebut juga siklus kreb, menggunakan nama Hans Krebsseorang
ahli biokimia yang berjasa banyak dalam penelitian metabolisme karbohidrat.
Siklus ini merupakan siklus dimana terjadi penggabungan antara molekul asetil koA dengan oksaloasetat hingga terbentuk asam trikarboksilat yaitu asam sitrat. Asam sitrat akan mengalami beberapa reaksi untuk akhirnya kembali membentuk oksaloasetat.
Gambar 11. Siklus asam sitrat
Sumber. Poedjiadi, Dasar-dasar Biokimia 1994, hlm. 270 Tahapan proses yang terjadi adalah sebagai berikut:
1. Asetil koA + oksaloasetat + H2O sitrat + koASH (enzim sitrat sintase) 2. Sitrat isositrat (enzim akonitase). Kerja enzim dapat dihambat oleh fluoroasetat. Hal ini dikarenakan fluoroasetat dapat berkondensasi dengan oksaloasetat membentuk fluorositrat yang menghambat kerja enzim akonitase.
3. Isositrat + NAD+ α – ketoglutarat + CO2 + NADH + H+ (enzim isositrat dehidrogenase). Proses ini melalui rantai pernapasan akan menghasilkan 3 ATP.
4. α – ketoglutarat + NAD+ + koASH Suksinil ko-A + CO2 + NADH H+ (enzim α – ketoglutarat dehidrogenase). Proses ini juga menghasilkan 3 ATP. Kerja enzim dapat dihambat oleh arsenat.
5. Suksinil KoA + GDP +Pi Suksinat + GTP + koASH (enzim suksinat tiokinase). Melalui tingkat substrat maka GTP dapat menyumbang 1 gugus phospat ke ADP untuk menghasilkan ATP.3
6. Suksinat + FAD Fumarat + FADH2 (enzim suksinat dehidrogenase).
Kerja enzim dapat dihambat malonat yang sifat inhibisinya ialah kompetitif. Jumlah ATP yang dihasilkan melalui proses ini ialah 2 ATP.
7. Fumarat + H2O Malat (enzim fumarase).
8. Malat + NAD+ Oksaloasetat + NADH + H+ (enzim malat dehidrogenase). Jumlah ATP yang dihasilkan melalui proses ini ialah sebesar 3 ATP.
Regulasi terutama dari siklus asam sitrat adalah konsentrasi produk.
Semakin tinggi konsentrasi produk, maka enzim untuk mensintesisnya semakin dihambat.2
Hasil dari siklus asam sitrat adalah 24 ATP, yang terdiri dari:
1. 3 NADH : 9 ATP 2. 1 FADH2 : 2 ATP 3. 1 GTP : 1 ATP
Karena ada 2 molekul asetil koA, maka jumlah energi menjadi 12 x 2 ATP
= 24 ATP.
Dari ketiga proses diatas total energi yang dihasilkan dalam oksidasi satu molekul glukosa ialah sebesar 38 ATP (glikolisis 8 ATP, pembentukan Asetil KoA 6 ATP dan siklus asam sitrat 24 ATP).
6) Glukoneogenesis
Glukoneogenesis, proses sintesis glukosa dari prekursor bukan karbohidrat, terjadi terutama di hati pada keadaan puasa. Glukoneogenesis berlangsung melalui suatu jalur yaitu (Marsk, dkk 2012):
1. Perubahan Piruvat Menjadi Fosfoenolpiruvat
Piruvat mengalami karboksilasi oleh piruvat karboksilase untuk membentuk oksaloasetat. Enzim ini, yang memerlukan biotin, adalah katalisator reaksi anaplerotik pada siklus asam trikarboksilat. Pada glukoneogenesis, reaksi ini melengkapi lagi oksaloasetat yang digunakan untuk sintesis glukosa.
CO2 yang ditambahkan ke piruvat untuk membentuk oksaloasetat dibebaskan oleh fosfoenolpiruvat karboksikinase (PEPCK) dan dihasilkan fosoenolpiruvat. Untuk reaksi ini, GTP merupakan sumber energi serta sumber gugus fosfat fosfoenolpiruvat. Enzim-enzim yang mengkatalisis kedua langkah ini terletak di dua kompartemen subsel yang berbeda. Piruvat karboksilase dijumpai di mitokondria. Pada berbagai spesies, fosfoenolpiruvat karboksikinase terletak di sitosol atau mitokondria, atau tersebar di kedua kompartemen ini. Pada manusia, enzim ini tersebar hampir sama banyak di masing-masing kompartemen.
Oksaloasetat, yang dihasilkan dari piruvat oleh piruvat karboksilase atau dari asam amino yang membentuk antara pada siklus asam trikarboksilat, tidak mudah menembus mitokondria. Oksaloasetat mengalami dekarboksilasi menjadi fosfoenolpiruvat oleh fosfoenolpiruvat karboksikinase mitokondria, atau diubah menjadi malat atau aspartat. Perubahan oksaloasetat menjadi malat memerlukan NADH. Fosfoenolpiruvat, malat, aspartat dapat dipindahkan ke dalam sitosol.
Setelah menembus membran mitokondria dan masuk ke dalam sitosol, malat dan aspartat diubah kembali menjadi oksaloasetat. Perubahan malat menjadi oksaloasetat menghasilkan NADH. NADH diperlukan untuk mereduksi 1,3-bisfosfogliserat menjadi gliseraldehida 3-fosfat selama glukoneogenesis.
Oksaloasetat, yang dihasilkan dari malat atau aspartat di siotosol, diubah kembali menjadi fosfoenolpiruvat oleh fosfoenolpiruvat karboksikinase sitosol.
Gambar 12. Perubahan Piruvat Menjadi Fosfoenolpiruvat Sumber. Marks, D.B, Marks, A.L dan Smith, C.M, Biokimia Kedokteran
Dasar, 2012
2. Perubahan Fosfoenolpiruvat Menjadi Fruktosa 1,6-Bifosfat
Langkah glukoneogenesis selanjutnya berlangsung di dalam sitosol.
Fosfoenolpiruvt membalikkan langkah pada glikolisis untuk membentuk gliseraldehida 3-fosfat. Untuk setiap 2 molekul gliseraldehida 3-fosfat yang terbentuk, 1 di ubah menjadi dihidroksiaseton fosfat (DHAP).
Kedua triosa fosfat ini, DHAP dan gliseraldehida 3-fosfat, berkondensasi untuk membentuk fruktosa 1,6-biosfosfat melalui kebalikan dari reaksi aldolase.
3. Perubahan Fruktosa 1,6-biosfosfat menjadi fruktosa 6-fosfat
Enzim fruktosa 1,6-biosfosfatase membebaskan fosfat inorganik dari fruktosa 1,6-biosfosfat untuk membentuk 6-fosfat. Enzim glikolitik, fosfofruktokinase-1, tidak mengkatalisis reaksi ini melainkan suatu reaksi yang melibatkan ATP.
4. Perubahan Glukosa 6-fosfat Menjadi Glukosa
Glukosa 6-fosfatase memutuskan Pi dari glukosa 6-fosfat, dan membebaskan glukosa bebas untuk masuk ke dalam darah. Enzim glikolitik glukokinase, yang mengkatalisis reaksi sebaliknya, memerlukan ATP. Glukosa 6-fosfatase terletak di membran retikulum endoplasma.
Glukosa 6-fosfatase digunakan tidak saja pada glukoneogenesis, tetapi juga untuk menghasilkan glukosa darah daripemecahan glikogen hati.
7) Pengaturan Glukoneogenesis
Karena hati dapat membuat glukosa melalui glukoneogenesis dan menggunakan glukosa melalui glikolisis, maka harus ada suatu sistem pengaturan yang mencegah agar kedua lintasan ini bekerja serentak. Sistem pengatur juga harus menjamin bahwa aktivitas metabolik hati sesuai dengan status gizi tubuh, yaitu pembentukan glukosa selama puasa dan menggunakan puasa pada saat glukosa banyak. Aktivitas glukoneogenesis dan glikolisis diatur secara terkoordinasi dengan cara perubahan jumlah relatif glukagon dan insulin dalam sirkulasi (Colby, 1998).
Bila kadar glukosa dan insulin darah turun, asam lemak dimobilisasi dari cadangan jaringan adiposa dan aktivitas 𝛽 −oksidasi dalam hati meningkat. Hal ini mengakibatkan peningkatan konsentrasi asam lemak dan asetil-KoA dalam hati.
Karena asam amino secara serentak dimobilisasi dari otot, maka juga terjadi peningkatan kadar asam amino, terutama alanin. Asam amino hati diubah menjadi piruvat dan substrat lain glukoneogenesis. Peningkatan kadar asam lemak, alanin, dan asetil-KoA semuanya memegang peranan mengarahkan substrat masuk ke glukoneogenesis dan mencegah penggunaannya oleh siklus asam sitrat. Asetil-KoA secara alosterik mengaktifkan piruvat karboksilase dan menghambat piruvat dehidrogenase. Oleh karena itu, menjamin bahwa piruvat akan diubah menjadi oksaloasetat. Piruvat kinase dihambat oleh asam lemak dan alanin, jadi menghambat pemecahan PEP yang baru terbentuk menjadi piruvat (Colby, 1998).
Pengaturan hormonal fosfofruktokinase dan fruktosa 1,6-bisfosfatase diperantarai oleh senyawa fruktosa 2,6-bisfofat. Pembentukan dan pemecahan senyawa pengatur ini dikatakan fruktosa 2,6-bisfosfat sejajar dengan perubahan untuk glukosa dan insulin yaitu konsentrasinya meningkat bila glukosa banyak dan berkurang bila glukosa sedikit. Fruktosa 2,6-bisfosfat secara alosterik mengaktifkan fosfofruktokinase dan menghambat fruktosa 1,6-bisfosfatase. Jadi, bila glukosa banyak, glikolisis aktif dan glukoneogenesis dihambat. Bila kadar glukosa turun, penngkatan glukagon mengakibatkan penurunan konsentrasi fruktosa 2,6-bisfosfat dan penghambatan yang sederajat pada glikolisis dan pengaktifan glukoneogenesis (Colby, 1998).
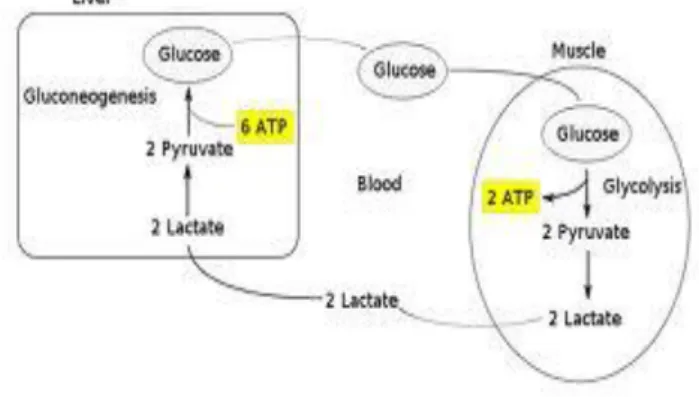
8) Siklus Cori
Lokalisasi enzim-enzim tertentu hanya dalam sel-sel tertentu berarti bahwa beberapa organ tergantung pada yang lain untuk melengkapi metabolisme substrat tertentu. Selama karbohidrat diperhitungkan, hati dan otot rangka menjalankan suatu kerjasama metabolisme tertentu. Otot rangka memperoleh ATP selama berlatih, hampir semuanya dari glikolisis. Sebagai hasilnya, produk akhir laktat memasuki darah. Laktat ini kemudian dihilangkan dari darah oleh hati terutama melalui isozim M4 laktat dehidrogenase yang mengkatalisis konversi cepat laktat menjadi piruvat. Bila hati biasanya dalam keadaan berenergi tinggi, maka mayoritas piruvat ini diubah oleh jalur glukoneogenesis menjadi glukosa 6- fosfat. Senyawa ini bisa dihidrolisis menjadi glukosa dengan enzim glukosa-6- fosfatase yang kemudian dapat memasuki darah, di mana glukosa ditranspor menuju otot rangka. Dalam otot rangka, glukosa diubah menjadi glukosa 6-fosfat dengan enzim heksokinase lalu memasuki glikolisis. Proses ini disebut Siklus Cori,
yang dinamai menurut penemunya yakni Carl dan Gerty Cori yang memenangkan hadiah Nobel pada tahun 1937 (Ngili, 2009).
Gambar 13. Siklus cori
Sumber. Ngili, Y. Biokimia Metabolisme Bionergetika, 2009 c. Gangguan Metabolisme Karbohidrat pada Kesehatan
Diantara berbagai penyakit gangguan metabolisme karbohidrat yang paling banyak diketahui penyebabnya adalah diabetes mellitus, galaktosemia dan penyakit penyimpanan glikogen.
1) Diabetes Mellitus
Gejala pertama yang ditunjukkan oleh penyakit ini adalah hiperglisemia, glukosuria, poliuria, polidipsi dan polipagi, turunanya berat badan, ketonemia, ketonuria, asidosis. Gejala kedua yang diakibatkan oleh penyakit ini dalam waktu yang lama adalah degenerasi dinding pembuluh darah dan pengaruhnya terhadapa berbagai organ tubuh, terutama kemungkinan terjadinya kebutaan. Terdapatnya glukosa di dalam air kemih menunjukkan kadar glukosa yang tinggi di dalam plasma darah (hierglukosemia). Hal ini disebabkan karena pengangkutan glukosa ke dalam sel dihambat, sehingga glukosa tidak dapat dioksidasi (melalui proses glikolisis) atau tidak dapat diubah menjadi glikogen (proses glikogenolisis) akibatnya proses reaksi penghasil energi (glikolisis) akan berkurang sehingga mempengaruhi laju reaksi jalur metabolisme yang memerlukan energi (anabolisme) (Wirahadikusumah, 1985).
Proses pengangkutan glukosa ke dalam sel bergantung pada insulin, suatu hormon yang dikeluarkan oleh kelenjar pankreas. Kekurangan insulin akan menyebabkan terhambatnya proses pengangkutan tersebut. Mekanisme lain dari pengaruh insulin terhadap metabolisme karbohidrat adalah terhalangnya tahap reaksi masuknya asetil – KoA ke dalam daur asam trikarboksilat. Penghalangan ini, sedemikian rupa sehingga asetil- KoA yang juga merupakan hasil katabolisme asam lemak bukannya digunakan sebagai sumber energi oleh daur asam trikarboksilat, tetapi diubah menjadi
asetoasetat, aseton dan β-hidroksibutirat, yang kemudian dikeluarkan dari sel dan diekskresi bersama urin (Wirahadikusumah, 1985).
2) Galaktosemia
Galaktosa sebagai hasil perombakan karbohidrat (melalui laktosa) yang masuk dari usus halus kedalam aliran darah diubah menjadi glukosa 1-fosfat.
Penyakit galaktosemia pada bayi dan anak-anak disebabkan karena tidak terdapat enzim fofogalaktosa uridietransferase. Enzim ini berperan dalam pembentukan glukosa 1-fosfat dari galaktosa. Dalam keadaan ini galaktosa 1- fosfat tetimbun dalam sel darah merah dan jaringan tertentu lainnya. Keadaan ini menyebabkan kerusakan pada hati, otak dan lensa mata. Pada orang dewasa kekurangan enzim ini tidak menyebabkan penyakit galaktosemia karena adanya enzim UDP-galaktosa fosfolirasi (UTP: galaktosa 1-fosfat uridetranferase) yang dapat mengubah galaktosa 1-fosfat menjadi UDP- galaktosa, yang selanjutnya diubah oleh UDP-galaktosa, yang selanjutnya diubah oleh UDP-galaktosa epimerase menjadi UDP-galaktosa (Wirahadikusumah, 1985).
3) Penyakit Penyimpanan Glikogen
Penyakit ini terdiri dari berberapa kelainan yang masing-masing merupakan akibat adanya ketidaknormalan dari reaksi ireversibel perubahan glukosa ke glikogen. Penyakit keturunan yang langka ini ditunjukkan dengan tertimbunnya glikogen dalam hati, jantung, atau otot rangka. Keadaan ini dapat mengakibatkan hilangnya daya tahan tubuh terhadap infeksi. Salah satu macam penyakit ini yang disebut penyakit Von Gierke, disebabkan oleh tidak terdapatnya glukosa 6-fosfatase, yaitu enzim yang mengkatalisis perubahan glukosa 6-fosfat ke glukosa. Penyebab lainnya adalah tidak terdapatnya enzim amilo(1->6) gukosidase yang berperan dalam pembentukan atau pemecahan (1->6) glikosida antara satuan glukosa dalam molekul glikogen yang bercabang. Tanpa adanya enzim ini proses perombakan glikogen terbatas hanya sampai molekul dekstrin saja, karena ikatan glikosida ada bagian yang bercabang tidak dapat dipecahkan. Penyakit ini disebut penyakit Cori. Penyakit kelainan glikogen lainnya adalah penyakit Pompe (tidak terdapat enzim 𝛼(1-
>4 ) glukosidase, penyakit Anderson (kurangnya enzim amilo (1->4 ) (1->6) transglukosidase, penyakit McArdle (tidak terdapatnya enzim fosforilase otot), dan penyakit Hers (tidak terdapatnya enzim fosforilase hati) (Wirahadikusumah, 1985).
I. Vitamin
Vitamin adalah suatu senyawa organik yang terdapat di dalam makanan dalam jumlah yang sedikit, dan dibutuhkan dalam jumlah yang besar untuk fungsi metabolisme yang normal. Vitamin dapat larut di dalam air dan lemak. Vitamin yang larut dalam lemak adalah vitamin A, D, E, dan K, dan yang larut dalam air adalah vitamin B dan C (Dorland, 2006).
Vitamin adalah molekul organik yang di dalam tubuh mempunyai fungsi yang sangat berfariasi. Fungsi vitamin dalam metabolisme yang paling utama adalah sebagai kofaktor. Di dalam tubuh diperlukan dalam jumlah sedikit (mikronutrien). Biasannya tidak disintesis di dalam tubuh, jika dapat disintesis jumlahnya tidak mencukupi kebutuhan tubuh, sehingga harus diperoleh dari makanan (Muchtadi, 2009).
Vitamin adalah sekelompok senyawa organik berbobot molekul kecil yang memiliki fungsi vital dalam metabolisme organisme.
Dipandang dari sisi enzimologi (ilmu tentang enzim), vitamin adalah kofaktor dalam reaksi kimia yang dikatalisasi oleh enzim. Istilah
"vitamin" sebenarnya sudah tidak tepat untuk dipakai tetapi akhirnya dipertahankan dalam konteks ilmu kesehatan dan gizi (Saifudin, 2009).
Vitamin bersifat larut lemak dan air. Kelompok vitamin yang bersifat larut lemak yaitu vitamin A, D, E dan K sedangkan vitamin yang larut air yaitu vitamin B dan C.
a. Vitamin Larut dalam Lemak
Vitamin yang larut dalam lemak adalah molekul hidrofobik apolar, yang semua merupakan derivat isoprene. Molekul ini tidak dapat disintesis oleh tubuh dalam jumlah yang memadai sehingga harus dipasok dari makanan. Vitamin larut lemak dapat diserap secara efisien jika terdapat penyerapan lemak yang normal. Begitu diserap, molekul vitamin tersebut harus diangkut di dalam darah seperti halnya lipid apolar yang lain, yaitu dalam lipoprotein atau melekat pada protein pengikat spesifik. Kelompok vitamin yang termasuk larut lemak adalah vitamin A, D, E, dan K. (Muray, dkk 2003).
1) Vitamin A Prinsip
Vitamin A merupakan zat gizi yang penting (esensial) bagi manusia, karena zat gizi ini tidak dapat disintesa oleh tubuh, sehingga harus dipenuhi dari luar. Vitamin A penting untuk kesehatan mata dan mencegah kebutaan dan yang lebih penting lagi vitamin A meningkatkan daya tahan tubuh. Anak-anak yang cukup mendapatkan vitamin A, bila terkena diare, campak atau penyakit infeksi lainnya maka penyakit-penyakit tersebut tidak mudah menjadi parah, sehingga tidak membahayakan jiwa anak. Dengan adanya bukti-bukti yang menunjukkan peranan vitamin A dalam menurunkan angka kematian, maka selain untuk mencegah kebutaan, pentingnya vitamin A saat ini lebih dikaitkan dengan kelangsungan hidup, kesehatan dan pertumbuhan anak.
Vitamin A juga merupakan suatu kristal alkohol berwarna kuning dan larut lemak atau pelarut lemak. Dalam makanan vitamin A biasanya terdapat dalam bentuk ester retinil, yaitu terikat pada asam
lemak rantai panjang. Didalam tubuh, vitamin A berfungsi dalam beberapa bentuk ikatan kimia aktif, yaitu retinol (bentuk alkohol), retinal (aldehida), dan asam retinoat (bentuk asam) (Almatsier, 2002).
Didalam sayur, vitamin A bewujud sebagai provitamin dalam bentuk pigmen 𝛽 – karoten berwarna kuning, yang terdiri atas dua molekul retinal yang bergabung pada ujung aldehid rantai karbonnya.
Meskipun demikian, karena metabolisasi 𝛽 – karoten menjadi vitamin A tidak berlangsung efisien, efektivitas 𝛽 – karoten sebagai sumber vitamin A adalah hanya sekitar seperenam aktivitas retinol berdasaran berat. Senyawa mirip 𝛽 – karoten dikenal sebagai karotenoid. Struktur kimia ketiga bentuk vitamin A dan provitaminnya 𝛽 – karoten (Almatsier, 2002). Dapat dilihat pada gambar 24.
Gambar 23. Struktur kimia tiga bentuk vitamin A dan 𝛽 – karoten Sumber. Winarno, F.G. Kimia Pangan dan Gizi, 2002 hlm 120 Metabolisme Vitamin A
Vitamin A dan 𝛽 - karoten diserap dari usus halus dan sebagian besar disimpam didalam hati. Bentuk karoten dalam tumbuhan selain beta, adalah alpa, Y-karoten serta kriptosantin.
Setelah dilepaskan dari bahan pangan dalam proses pencernaan, senyawa tersebut diserap oleh usus halus dengan bantuan asam empedu (pembentukan micelle). Vitamin A dan karoten diserap oleh usus dari micelle secara difusi pasif, kemudian digabungkan dengan kilomikron dan diserap melalui saluran limpatik, kemudian bergabung dengan saluran darah dan ditransportasikan ke hati. Di hati, vitamin A digabungkan dengan asam palmitat dan disimpan dalam bentuk retinil-palmitat. Bila diperlukan oleh sel-sel tubuh, retinil palmitat diikat oleh protein pengikat retinol (PPR) atau retinol-binding protein (RBP), yang disintesis dalam hati.
Selanjutnya ditransfer keprotein lain, yaitu “transthyretin” untuk diangkut ke sel-sel jaringan (Almatsier, 2002).
Vitamin A yang tidak digunakan oleh sel-sel tubuh diikat oleh protein pengikat retinol selular (celluler retinol binding protein), sebagian diangkut ke hati dan bergabung dengan asam empedu, yang selanjutnya diekskresikan ke usus halus, kemudian dikeluarkan dari tubuh melalui feses. Sebagian lagi diangkut ke ginjal dan diekskresikan melalui urine dalam bentuk asam retinoat (Almatsier, 2002).
Karoten diserap oleh usus seperti halnya vitamin A, sebagian dikonversi menjadi retinol dan metabolismenya sama seperti diatas. Sebagian kecil karoten disimpan dalam jaringan adiposa dan tidak digunakan oleh tubuh diekskresikan bersama asam empedu melalui feses. Pada diet nabati di lumen usus, oleh enzim beta-karoten 15, 15-deoksigenase, 𝛽-karoten tersebut dipecah menjadi retinal (retinaldehid), yang kemudian direduksi menjadi retinol oleh enzim retinaldehid reduktase. Pada diet hewani, retinol ester dihidrolisis oleh esterase dari pankreas, selanjutnya diabsorsi dalam bentuk retinol, sehingga diperlukan garam empedu (Almatsier, 2001).
Proses diatas sangat terkontrol, sehingga tidak memungkinkan produksi vitamin A dari karoten secara berlebihan.
Tidak seluruh karoten dapat dikonversi menjadi vitamin A, sebagian diserap utuh dan masuk ke dalam sirkulasi, hal ini akan digunakan tubuh sebagai antioksidan (Almatsier, 2001).
Gangguan Metabolisme Vitamin A Bagi Kesehatan
Penyakit yang ditimbulkan akibat kekurangan vitamin A, antara lain rabun senja (night blindness), katarak, infeksi saluran pernapasan, menurunnya daya tahan tubuh, keratinisasi (sel epitel kering), kulit yang tidak sehat, bersisik dan mengelupas.
Hiperviaminosis A terjadi pada anak-anak, kelebihan ditandai dengan kemunculan gejala-gejala, antara lain hilangnya nafsu makan, mual, berat badan menurun, pusing, luka disudut mulut, bibir pecah-pecah, rambut rontok dan nyeri tulang.
2) Vitamin D Prinsip
Vitamin D merupakan prohormon steroid. Vitamin ini diwakili oleh sekelompok senyawa steroid yang terutama terdapat pada hewan, tanaman, dan ragi. Meleluiberbagai perubahan metabolik di dalam tubuh, vitamin D menghasilkan suatu hormon yang dikenal dengan nama kalsitriol yang mempunyai peranan sentral pada metabolisme kalsium dan fosfat (Murray, dkk 2003).
Vitamin D adalah nama genetik dari dua molekul yaitu ergokalsiferol (vitamin D2) dan kolekalsiferol (vitamin D3). Prekusor
vitamin D hadir dalam fraksi sterol dalam jaringan hewan (di bawah kulit) dan tumbuh-tumbuhan berturut-turut dalam bentuk 7- dehidrokolesterol dan ergosterol. Keduanya membutuhakan radiasi sinar ultraviolet untuk mengubah ke dalam bentuk provitamin D3
(kolekalsiferol). Kedua provitamin membutuhkan konversi menjadi bentuk aktifnya melalui penambahan dua gugus hidroksil. Gugus hidroksil pertama ditambahkan di dalam hati pada posisi 25 sehingga membentuk 25-hidroksi-vitamin D. Gugus hidroksil kedua ditambahkan di dalam ginjal sehingga membentuk 1,25-dihidroksi- vitamin D. Provitamin D berasal dari hewan membentuk 1,25
dihidroksikolekalsiferol, dikenal sebagai kalsitriol, sedangkan yang berasal dari tumbuh-tumbuhan membentuk 1,25 dihidroksi ergokalsiferol, dikenal sebagai erkalsitriol (lihat gambar 24). Kedua bentuk vitamin D efektif untuk manusia (Almatsier, 2001).
Gambar 24. Sintesis vitamin D di dalam tubuh
Sumber. Almatsier, S. Prinsip-prinsip Dasar Ilmu Gizi, 2001.hlm 169 Metabolisme Vitamin D
Vitamin D3 (kolekalsiferol) dibentuk di dalam kulit oleh sinar ultraviolet dari 7-dehidrokolesterol. Sinar matahari juga dapat
mengubah provitamin D3 menjadi bahan yang tidak aktif.
Banyaknya provitamin D dan bahan tidak aktif yang dibentuk bergantung pada intensitas radiasi ultraviolet. Faktor lain yang berpengaruh terhadap pembentukan provitamin D adalah pigmentasi, penggunaan alas penahan matahari dan lama waktu penyingkapan terhadap matahari (Almatsier, 2001).
Vitamin D3 di dalam hati diubah menjadi lemak aktif 25- hidroksi kolekalsiferol (25(OH)D3) yang lima kali lebih aktif daripada vitamin D3. Bentuk 25(OH)D3 adalah bentuk vitamin D yang paling banyak di dalam darah dan banyaknya bergantung pada konsumsi dan penyingkapan tubuh terhadap matahari. Bentuk paling aktif adalah kalsitriol atau 1,25-dihidroksi kolekalsiferol (1,25(OH)2D3) yang sepuluh kali lebih aktif dari vitamin D3. Bentuk ini dibuat oleh ginjal, kalsitriol pada usus halus meningkatkan absorpsi kalsium dan fosfor pada tulang meningkatkan mobilisasinya (Almatsier, 2001).
Sintesis kalsitriol diatur oleh taraf kalsium dan fosfor dalam serum. Hormon paratiroid (PTH) yang dikeluarkan bila kalsium dalam serum darah rendah, tampaknya merupakan perantara yang merangsang produksi 1,25-dihidroksi kolekalsiferol oleh ginjal. Jadi taraf konsumsi kalsium yang rendah tercermin pada taraf kalsium serum yang rendah. Hal ini akan mempengaruhi sekresi PTH dan peningkatan sintesis kalsitriol oleh ginjal. Taraf fosfat dari makanan mempunyai pengaruh yang sama, tetapi tidak membutuhkan PTH (Almatsier, 2001). Gambar metabolisme vitamin D dapat dilihat pada gambar 25.
Gambar 25. Metabolisme vitamin D
Sumber. Holick, M.F. Vitamin D: photobiology, metabolism, mechanism of action, and clinical application. In: M.J. Favus (ed.) Primer on the Metabolic Bone Diseases and Disorders of Mineral Metabolism, Third Edition, pp. 74-8l, Lippincott-Raven: Philadelphia, 1996.
Gangguan Metabolisme Vitamin D Bagi Kesehatan
Defisiensi vitamin D menyebabkan rakitis dan osteomalasia.
Penyakit rakitis terjadi pada anak, dan osteomalasia pada orang dewasa, yang tidak terpajan cahaya matahari atau yang tidak mendapat vitamin D dalam jumlah memadai dari makanan. Kedua penyakit ini disebabkan oleh pelunakan tulang yang terjadi akibat kekurangan kalsium dan fosfat. Ikan berlemak, kuning telur, dan hati merupakan sumber vitamin D yang baik. Pemajanan cahaya matahari pada diri seseorang, yang ditentukan oleh ketinggian wilayah, musim, dan sejumlah faktor lain turut mempengaruhi kebergantungan relatif pada sumber makanan untuk memenuhi kebutuhan vitamin D. Asupan vitamin D dalam jumlah berlebihan akan menimbulkan hipervitamninosis yang ditandai oleh kadar kalsium yang tinggi di dalam darah serta kelsifikasi jaringan lunak (Murray, dkk 2003).
3) Vitamin E (Tokoferol) Prinsip
Vitamin E yang larut dalam lemak banyak terdapat dalam buah-buahan, susu, telur, sayur-sayuran, terutama pada biji-bijian.
Sekali diserap dalam tubuh, Tempat penyimpanan utama vitamin E adalah di jaringan adiposa. Seperti halnya lemak, vitamin memerlukan protein pengangkut untuk memindahkannya dari satu tempat ke tempat lain (Winarno, 2004).
Ada berbagai jenis tooferol yang ditemukan di alam. Semua merupakan senyawa 6-hidroksikromana(tokol) yang bersubstitusi- isoprenoid. Ada empat jenis tokoferol yang penting dalam makanan yaitu alfa-, beta-, gama-, delta-tokoferol dan tokotrienol.
Karakteristik kimia utamanya adalah bertindak sebagai antioksdan.
Tokoferol terdiri atas struktur cincin 6-kromonal dengan rantai samping jenuh panjang enam belas karbon fitol (Winarno, 2004).
Perbedaan antar jenis tokoferol pada jumlah dan gugus metil pada struktur cincin (lihat gambar 26).
Tokotrienol mempunyai tiga ikatan rangkap pada rantai samping. Perbedaan struktur ini mempengaruhi tingkat aktivitas vitamin E secara biologik. Tokotrienol tidak banyak terdapat di alam dan kurang aktif secara biologik. Alfa-tokoferol adalah bentuk vitamin E paling aktif, yang digunakan pula sebagai standar
pengukuran vitamin E dalam makanan. Jumlah sintetik vitamin E mempunyai aktivitas biologik 50% daripada alfa-tokoferol yang terdapat di alam (Almatsier, 2001).
Gambar 26. Struktur kimia tokoferol dan tokotrienol
Sumber. Almatsier, S. Prinsip-prinsip Dasar Ilmu Gizi, 2001. Hlm 174
Metabolisme Vitamin E
Komponen vitamin E dalam diit yang memiliki aktivitas antioksidan antara lain α-, β-, dan γ-tocopherol dan tocotrienol.
Semua molekul tersebut memiliki cincin chromanol dengan beragam jumlah gugus methyl dan memiliki ujung phytyl atau ujung unsaturated. Di dalam tubuh, hati mensekresikan α-tokoferol ke plasma dalam pengawasan α-tokoferol transfer protein (α-TTP).
Tidak seperti vitamin larut lemak lainnya, vitamin E tidak disimpan di dalam tubuh dalam jaringan hati atau jaringan lainnya dalam jumlah lebih dari 2-3 kali konsentrasi normal individu yang tidak mengkonsumsi suplemen vitamin E. Di dalam tubuh, bentuk metabolit dari tokoferol adalah CEHC (carboxyethyl hydroxychroman) yang dimetabolisir seperti xenobiotik oleh cytochrome P450s. Hasil metabolisme tersebut dikonjugasikan lalu diekskresi melalui urin dan empedu (Anonim, 2009).
Metabolisme vitamin E merupakan kunci untuk menegah akumulasi bentuk-bentuk vitamin E selain α-tokoferol. Pada pemberian sumplementasi α-, dan γ-tocopherol pada manusia, diketahui bahwa γ-tokoferol lebih cepat dimetabolisme dan dikeluarkan dibandingkan dengan bentuk α-nya. α-tokoferol merangsang metabolisme dan ekskresi non-α-tokoferol dengan meningkatkan kadar CYP3A. Pengaturan ini penting untuk menjaga tubuh dari efek ekumulasi tokoferol yang berlebihan (Anonim, 2009).
Gangguan Metabolisme Vitamin E Bagi Kesehatan
Penyakit kekurangan vitamin E pada manusia jarang terjadi, karena vitamin E terdapat luas di dalam bahan makanan.
Kekurangan biasanya terjadi karena adanya gangguan absorpsi lemak seperti pada cystic fibrosis dan gangguan transpor lipida seperti pada beta-lipopro-teinemia. Kekurangan vitamin E pada manusia menyebabkan hemolisis eritrosit, yang dapat diperbaiki dengan pemberian tambahan vitamin E. Akibat lain adalah sindroma neurologik sehingga terjadi fungsi tidak normal pada sumsum tulang belakang dan retina. Tanda-tandanya adalah kehilangan koordinasi dan refleks otot, serta gangguan penglihatan dan berbicara (Almatsier, 2001).
Mengkonsumsi vitamin E secara berlebihan dapat menimbulkan keracunan. Gangguan pada saluran cerna terjadi bila mengkonsumsi lebih dari 600 miligram sehari (60-75 kali kecukupan). Dosis tinggi juga dapat meningkatkan efek obat antikoagulan yang digunakan untuk mencegah penggumpalan darah (Almatsier, 2001).
4) Vitamin K Prinsip
Vitamin yang tergolong ke dalam kelompok vitamin K adalah naftokuinon tersubstitusi-polilisoprenoid. Vitamin K terdapat di dalam alam dalam dua bentuk, keduanya terdiri atas cincin 2- metilnaftakinon dengan rantai samping pada posisi tiga (Gambar 28). Menadion (K3)-senyawa induk seri vitamin K – tidak ditemukan secara alami, tetapi jika diberikan akan mengalami alkilasi ini vivo menjadi salah satu menakuinon (K2). Filokuinon (K1) merupakan bentuk utama vitamin K yang ada pada tanaman. Menakuinon-7 merupakan salah satu anggota seri bentuk tak jenuh poliprenoid vitamin K yang ditemukan pada jaringan hewan dan disintesis oleh bakteri di dalam usus (Murray, dkk 2003).
Menadion (vitamin K3)
Gambar 27. Struktur kimia vitamin K
Sumber. Almatsier, S. Prinsip-prinsip Dasar Ilmu Gizi, 2001. Hlm 180
Metabolisme Vitamin K
Reaksi karboksilase bergantung-vitamin K berlangsung di dalam retikulum endoplasma banyak jaringan dan membutuhkan oksigen molekuler, karbon dioksida, serta bentuk hidrokuinon (bentuk tereduksi) vitamin K. Di dalam retikulum endoplasma hati terjadi siklus vitamin K (Gambar 28), pada siklus ini, produk 2,3- epoksida hasil reaksi karboksilase diubah oleh 2,3-epoksida reduktase menjadi bentuk kuinon vitamin K dengan menggunakan suatu zat pereduksi ditiol (Murray, dkk 2003) .
Gambar 28. Sintesis Vitamin K di dalam hati
Sumber. Modifikasi dari Murray, R.K, Granner, D.K, Mayes, P.A, Rodwell V.W, Biokimia Harper, 2003
Reaksi ini sensitif terhadap inhibisi oleh antikoagulan tipe 4- hidroksidikoumarin (dikumarol), seperti warfarin (Gambar 29).
Reduksi bentuk kuinon menjadi hidrokuinon oleh NADH yang terjadi sesudahnya akan melengkapi siklus vitamin K untuk menghasilkan kembali bentuk aktif vitamin tersebut (Murray, dkk 2003).
Gambar 29. Antagonis vitamin K yang digunakan sebagai antikoagulan: Dikumarol dan Warfarin.
Sumber. Murray, R.K, Granner, D.K, Mayes, P.A, Rodwell V.W, Biokimia Harper, 2003.
Gangguan Metabolisme Vitamin K Bagi Kesehatan
Kekurangan vitamin K menyebabkan darah tidak dapat membeku. Kekurangan vitamin K karena makanan jarang terjadi, sebab vitamin K terdapat secara luas dalam makanan. Kekurangan vitamin K terjadi bila ada gangguan absorpsi lemak. Sedangkan, kelebihan vitamin K terjadi bila vitamin K diberikan dalam bentuk berlebihan berupa vitamin K sinteeik menadin, gejala kelebihan vitamin K adalah hemolisis sel darah merah, sakit kunin (jaundice) dan kerusakan pada otak (Almatsier, 2001).
b. Vitamin Larut dalam Air
Vitamin larut dalam air adalah vitamin yang hanya dapat disimpan dalam jumlah sedikit dan biasanya akan segera hilang bersama aliran makanan. Sebaian besar vitamin larut air merupakan komponen sistem enzim yang banyak terlibat dalam membantu metabolisme energi. Vitamin larut air biasanya tidak disimpan dalam tubuh dan dikeluarkan melalui urine dalam jumlah kecil. Oleh sebab itu vitamin larut air perlu dikonsumsi setiap hari untuk mencegah kekurangan yang dapat mengganggu fungsi tubuh normal (Almatsier, 2001).
Vitamin larut air dikelompokkan menjadi vitamin C dan vitamin B-kompleks. Vitamin B-kompleks terdiri dari sepuluh faktor yang saling berkaitan fungsinya didalam tubuh dan terdapat didalam bahan makanan yang hampir sama. Fungsinya terkait dalam proses metabolisme sel hidup, baik pada tumbuh-tumbuhan maupun hewan sebagai koenzim dan kofaktor (Almatsier, 2001).
1) Vitamin B Kompleks
Vitamin B yang esensial bagi nutrisi manusia adalah tiamin (vitamin B1), riboflavin (vitamin B2), niasin (vitamin B3), asam pantotenat (vitamin B5), pyridoksin (vitamin B6), Biotin, Kobalamin (vitaminB12), asam folat (Murray dkk, 2003).
a) Vitamin B1 (Tiamin) Prinsip
Vitamin B1 (thio-vitamine, thiamine, thiamin) adalah vitamin yang terlarut dalam air. Tiamina terdiri atas cincin pirimidina dan cincin thiazola (mengandung sulfur dan nitrogen) yang dihubungkan oleh jembatan metilen. Turunan fosfatnya ikut serta dalam banyak proses sel. Tiamina disintesis dalam bakteri, fungi dan tanaman (Murray dkk, 2003).
Metabolisme Vitamin B1
Tiamin berperan sentral dalam metabolisme penghasil energi, dan khususnya metabolisme karbohidrat. Tiamin difosfat adalah koenzim untuk tiga kompleks multi-enzim yang mengatalisis reaksi dekarboksilasi oksidatif: piruvat dehidrogenase dalam metabolisme karbohidrat. 𝛼-ketoglutarat dehidrogenase dalam siklus asam sitrat dan asam sitrat keto dehidrogenase rantai bercabang pada metabolisme leusin, isoleusin, dan valin. Pada masing-masing kasus, tiamin difosfat menyediakan sebuah karbon reaktif pada gugus tiazol yang membentuk suatu karbanion, yang kemudian menambah gugus karbonil pada, misalnya piruvat.
Senyawa tambahan kemudian mengalami dekarboksilasi dan mengeluarkan CO2. Tiamin trifosfat juga merupakan koenzim untuk transketolase, pada jalur pentosa merupakan fosfat (Gambar 30) (Murray dkk, 2009).
A) B)
C)
Gambar 30. A) Tiamin, B) Tiamin difosfat, dan C) bentuk karboniat
Sumber. Modifikasi dari Murray, R.K, Granner, D.K, Mayes, P.A, Rodwell V.W, Biokimia Harper, 2003
Tiamin difosfat memiliki peran penting dalam hantaran saraf;
senyawa ini memfosforilasi (sehingga mengaktifkan) kanal klorida di membrane saraf.
Gangguan Metabolisme Vitamin B1 Bagi Kesehatan
Defisiensi tiamin dapat menyebabkan tiga sindrom tersendiri: suatu neuritis perifer kronik, beriberi, yang dapat berkaitan atau tidak dengan gagal jantung dan edema; beriberi penisiosa (fulminan) akut (beriberi shoshin) dengan gejala yang predominan berupa gagal jantung dan kelainan metabolik tanpa neuritis perifer; dan ensefalopati Wernicke disertai psikosis Korsakoff, yang terutama berkaitan dengan penyalahgunaan alcohol dan narkotik. Peran tiamin difosfat dalam piruvat dehidrogenase memiliki arti bahwa pada defisiensi terjadi gangguan perubahan piruvat menjadi asetil KoA. Pada orang dengan diet karbohidrat yang relatif tinggi, hal ini menyebabkan meningkatnya kadar laktat dan pirufat plasma, yang dapat menyebabkan asidosis laktat yang mengancam jiwa (Murray dkk, 2009).
b) Vitamin B2 (Riboflavin) Prinsip
Vitamin B2 diperlukan untuk berbagai ragam proses seluler. Seperti vitamin B lainnya, riboflavin memainkan peranan penting dalam metabolisme energi, dan diperlukan dalam metabolisme lemak, zat keton, karbohidrat dan protein. Vitamin ini juga banyak berperan dalam pembetukkan sel darah merah, antibodi dalam tubuh, dan dalam metabolisme pelepasan energi dari karbohidrat (Murray dkk, 2009).
Gambar 31. Struktur Riboflavin
Sumber. Winarno, F.G. Kimia Pangan dan Gizi, 2002.
Metabolisme Vitamin B2
Vitamin B2 (Riboflavin) Berperan Penting dalam Metabolisme Penghasil-Energi. Riboflavin menyediakan gugus-gugus reaktif koenzim flavin mononukleotida (FMN) dan flavin adenine dinukleotida (FAD). FMN dibentuk oleh fosforilasi riboflavin dependen-ATP;
sementara FAD disintesis oleh reaksi lebih lanjud ATP dengan gugus AMP yang dipindahkan le FMN. Sumber utama riboflavin dalam makanan adalah susu dan produk susu. Selain itu, karena warnanya yang kuning terang, riboflavin sering digunakan sebagai zat aditif makanan (Murray dkk, 2003).
Gangguan Metabolisme Vitamin B2 Bagi Kesehatan
Meskipun riboflavin berperan sentral dalam metabolisme lipid dan karbohidrat, dan defisiensi riboflavin terjadi di banyak Negara, namun defisiensi ini tidak mematikan karena penghematan riboflavin di jaringan sangat efisien. Defisiensi riboflavin ditandai oleh keilosis, deskuamasi dan peradangan lidah, dan dermatitis seboroik. Status gizi riboflavin dinilai dengan mengukur pengaktivan glutation reduktase eritrosit oleh FAD yang ditambahkan in vitro (Murray dkk, 2003).
c) Vitamin B3 (Niasin) Prinsip
Niasin ditemukan sebagai nutrien sewaktu penelitian tentang pellagra dilakukan. Niasin bukan suatu vitamin sejati karena zat ini dapat disintesis dalam tubuh dari asam amino esensial triptofan. Dua senyawa, asam nikotinat dan nikotinamida, memiliki aktivitas biologis niasin; fungsi metaboliknya adalah sebagai cincin nikotinamida pada koenzim NAD dan NADP dalam reaksi oksidsi/reduksi. sekitar 60 mg triptofan setara dengan 1 mg niasin dalam makanan. Kandungan niasin dalam makanan dinyatakan sebagai:
Mg niasin ekuivalen = mg niasin yang sudah ada + 1/60 x mg triptofan. Karena sebagian besar niasin dalam sereal tidak dapat digunakan secara biologis, jumlah ini tidak diperhitungkan.
Gambar 32. Struktur Kimia Niasin
Sumber. Almatsier, S. Prinsip-prinsip Dasar Ilmu Gizi, 2001 Metabolisme Vitamin B3
Nikotinat merupakan bentuk niasin yan diperlukan bagi sintesis NAD+ dan NADP+ oleh enzim yang terdapat di dalam sitosol sebagian besar sel. Oleh karena itu, setiap nikotinamida di dalam makanan pertama-tam harus mengalami deamidasi menjadi nikotinat. Di dalam sitosol, nikotinat diubah menjadii senyawa desamido-NAD+ melalui reaksi yang semula berlangsung dengan 5-fosforibosil 1-pirofosfat (PRPP), dan kemudian melalui adenilasi dengan ATP. Gugus amido pada glutamin kemudian turut membentuk koenzim NAD+. Koenzim ini bisa mengalami fosforilasi lebih lanjut membentuk NADP+ (Murray dkk, 2009).
Nukleotida nikotinamida memainkan peran luas sebagai koenzim banyak enzim dehidrogenase yang terdapat baik dalam sitosol maupun dalam mitokondria. Oleh karena itu, vitamin ini merupakan komponen kunci pada banyak lintasan metabolik yang mempengaruhi metabolisme karbohidrat, lemak serta asam amino (Murray dkk, 2009).
Gangguan Metabolisme Vitamin B3 (niasin) Bagi Kesehatan Niasin bersifat toksik jika pengunaannya berlebihan. Asam nikotinat digunakan untuk mengobati hiperglikemia dan jika digunakan dalam kisaran 1-6 g/hari dapat menyebabkan ditalasi pembuluh darah dan flushing serta iritasi kulit. Asupan asam nikotinamida yang melebihi 500 mg/hari juga menyebabkan kerusakan hati (Almatsier, 2001).
d) Vitamin B5 (Asam Pantotenat) Prinsip
Asam pantotenat dibentuk melalui penggabungan asam pantoat dengan 𝛽 −alanin (Gambar 33).
Gambar 33. Asam Pantotenat
Sumber. Murray, R.K, Granner, D.K, Mayes, P.A, Rodwell V.W, Biokimia Harper, 2003
Metabolisme Vitamin B5
Asam pantotenat dapat dengan mudah di absorpsi di dalam usus dan selanjudnya mengalami fosforilasi oleh ATP membentuk senyawa 4’-fosfopantotenat. Penambahan system dan pengeluaran gugus karboksilnya mengakibatkan adisi nettotioetanolamin sehingga menghasilkan 4’-fosfopantetein, gugus prostetik pada KoA dan protein pembawa asil (ACP, acyl carrier protein). Sepertinya halnya koenzim aktif banyak vitamin larut-air lain, KoA mengandung nukleotida adenine. Dengan demikian, 4’-fosfopantetein akan mengalami adenililasi oleh ATP membentuk defosfo-KoA. Fosforilasi akhir terjadi dengan ATP yang mengadisikan fosfat ke gugus 3’-hidroksil moietas ribose, menghasilkan KoA (Murray, dkk 2009).
Gangguan Metabolisme Vitamin B5 Bagi Kesehatan
Difisiensi asam pantotenat jarng terjadi karena tersebar luasnya asam pantotenat di dalam berbagai makanan, khususnya dalam jumlah berlimpah di dalam jaringan hewan, sereal utuh, dan kacang-kacangan meskipun demikian, sindrom burning foot pernah terjadi di antara para tawanan perang akibat defisiensi asam pantotenat dan ini berhubungan dengan menurunnya kemampuan asetilasi (Murray, dkk 2002).
e) Vitamin B6 (Pyridoksin) Prinsip
Vitamin B6 terdiri atas tiga derivat piridin yang berhubungan erat yaitu pirikdosin, piridoksal, dan piridoksamin (Gambar 34).
Gambar 34. Bentuk alami vitamin B6
Sumber. Murray, R.K, Granner, D.K, Mayes, P.A, Rodwell V.W, Biokimia Harper, 2003.
Ketiga bentuk ini memiliki aktivitas vitamin yang sama karena dapat melakukan interkonversi satu sama lain di dalam tubuh.
Metabolisme Vitamin B6
Vitamin B6 penting dalam metabolisme asam amino dan glikogen, juga dalam kerja hormone steroid. Terdapat enzim senyawa yang memiliki aktivitas vitamin B6. piridoksin, piridoksal, piridoksamin, dan turunan 5-fosfatnya. Koenzim aktif adalah piridoksal 5’-fosfat.
Sekitar 80& vitamin B6 total dalam tubuh adalah piridoksal fosfat di otot, sebagian besar berkaitan dengan glikogen fosforilase. Bentuk ini tidak dapat digunakan pada keadaan defisiensi, tetapi dibebaskan jika terjadi kelaparan, saat cadangan glikogen terkuras, dan kemudian dapat digunakan, terutama di hati dan ginjal untuk memenuhi peningkatan kebutuhan glukoneogenesis dari asam amino (Almatsier, 2001).
Vitamin B6 memiliki beberapa peran dalam metabolisme.
Piridoksal fosfat adalah suatu metabolisme asam amino, khusunya transaminasi dan dekarboksilasi. vitamin ini juga merupakan kofaktor glikogen fosforilase, dan gugus fosfat penting untuk katalisis. Selain itu vitamin B6 penting bagi kerja hormone steroid. Piridoksal fosfat mengeluarkan kompleks hormon-reseptor dari ikatan dengan DNA dan menghentikan kerja hormon. Pada defisiensi vitamin B6, terjadi peningkatan kepekaan terhadap kerja estrogen, androgen, kortisol, dan vitamin D konsentrasi rendah (Almatsier, 2001).
Gangguan Metabolisme Vitamin B6 Bagi Kesehatan
Meskipun gejala klinis defisiensi jarang dijumpai, namun terdapat bukti bahwa cukup banyak orang yang status vitamin B6-nya
marginal. Defisiensi tingkat sedang menyebabkan kelainan metabolisme triptofan dan metionin. Peningkatan kepekaan terhadap kerja hormone steroid mungkin penting dalam pembentukan kanker dependen-hormon pada payudara, uterus, dan prostat, dan status vitamin B6 mungkin memengaruhi prognosis (Almatsier, 2009).
Vitamin B6 menyebabkan neuropati sensorik jika berlebihan.
Timbulnya neuropati sensorik pernah dilaporkan pada pasien yang mengonsumsi 2-7 g piridoksin per hari untuk berbagai alasan (terdapat sedikit bukti bahwa vitamin ini efektif dalam mengobati sindrom prahaid). Pengertian pemberian dosis tinggi ini meninggalkan kerusakan residual, laporan lain menyatakan bahwa asupan melebihi 200 mg/hari berkaitan dengan kerusakan saraf (Almatsier, 2001).
f) Biotin Prinsip
Struktur biotin, biositin, dan karboksibiotin (zat perantara metabolik aktif) diperlihatkan gambar 35. Biotin tersebar luas di banyak makanan sebagai biositin (𝜀 -amino-biotinillisin), yang dibebaskan pada proteolisis (Murray dkk, 2009).
Gambar 35. Struktur Kimia Biotin
Sumber. Almatsier, S. Prinsip-prinsip Dasar Ilmu Gizi, 2001 Metabolisme Biotin
Biotin adalah koenzim dari enzim karboksilase. Biotin berfungsi memindahkan karbon dioksida dalam sejumlah kecil reaksi karboksilase (setil-KoA, piruvat, propionil-KoA, dan metilkrotonil-KoA karboksilase). Holokarboksilase sintase mengkatalisis pemindahan biotin menjadi residu lisin dari apo-enzim untuk membentuk residu biositin dari holoenzim. Zat antara reaktif adalah 1-N-karbosi-biotin, yang dibentuk dari bikarbonat dalam sebuah reaksi dependen-ATP.
Kelompok karboksi kemudian diubah menjadi substrat untuk karboksilasi. Biotin juga memiliki peranan dalam mengatur siklus sel, yang bekerja dengan melakukan biotinilasi pada inti protein utama (Murray dkk, 2009).