14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 3 segment shows that the primary genitalia of the male must have opened on the tenth segment of the abdomen. Also in Machilis it has been shown by Qadri (1940) that the primary phallic lobes develop independently of the appendages of the ninth segment.
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS
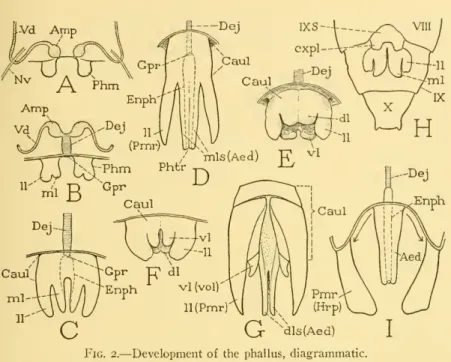
Teontogenetic origin of the phallus from a single pair of primary phallic lobes has been observed in almost all the larger orders of insects of Thysanurato Hymenoptera, with the exception of Mecoptera and Diptera. The six-lobed type of phallic structure appears to be fundamental in Orthoptera, as it is in Hymenoptera, but in the Orthoptera (E) there is Httle uniformity in the development of the lobes, and only in Acrididae do the lobes of the dorsal pair partially unite to to form an aedeagus (see Snodgrass, . 1937)-.
NO. 14 MALE GENITALIA OF IIYMENOPTERA SNODGRASS 9 unite to form an aedeagus, the endophallus is extended into the latter
THE TERMINAL BODY SEGMENTS AND THE PHALLUS OF HYMENOPTERA
THE TERMINAL BODY SEGMENTS
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS II
12 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 which there has been everted from this opening a large vesicle on
THE PHALLUS AND PHALLIC NOMENCLATURE
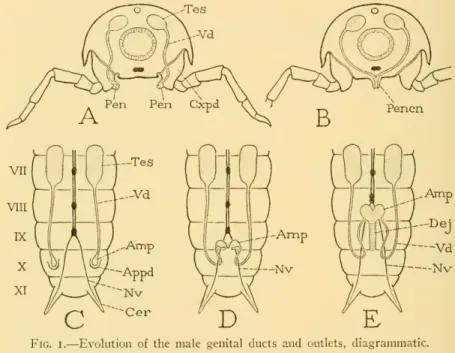
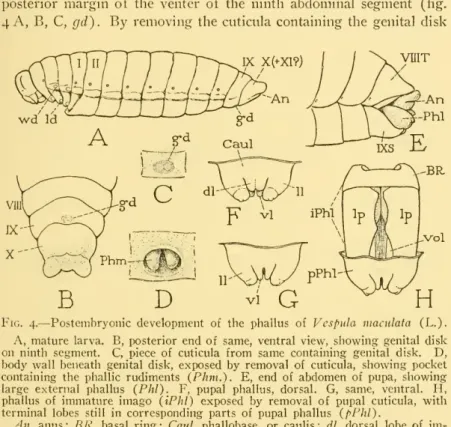
The papillae are primary phallic lobes or phallomeres (Phm) immersed in a "peripodal" pocket of the epidermis. Before molting, the undivided proximal parts of the two appendages fuse, and the dorsomesal lobes merge into a thick central structure.
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 1
The parameral plates (Fig. 5 A, B, Ip) are usually considered the basal parts of the parameres (Pmr), as they are generally con-. Furthermore, the parameres in most Chalastogastra and many Clistogastra are demarcated from the parameral plates of the caulis.
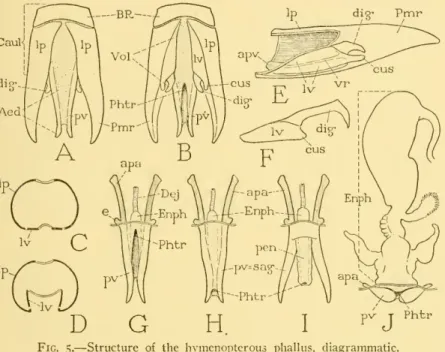
NO. 14 MALE GENITALIA OF HYMENOPTERA — SNODGRASS I9
20 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 The lateral walls of the aedeagus contain a pair of elongate
Although volsellae are completely absent in many bees, they are present as reduced structures in several species. It is the terminal portion of the definitive genital duct, formed between the united phallic rudiments, which is generally regarded by most writers as a continuation of the ductus ejaculatorius.
22 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 to one of the free volsellar lobes, or have named the lobes and the
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 23 of each plate usually forms an apoclemal inflection (apv) for the
24 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 meral plate or in the base of the paramere, inserted anteriorly on the
CHALASTOGASTRA
14 MALE GENITALIA OF HYMENOPTERA — SNODGRASS 25comparative study of the male organ in all other Hymenoptera. The aedeagus of most Chalastogastra is shaped like an inverted trough (secondarily open above in strophandriose forms) with thephallotreme at its base; in the Cephoidea, however, the aedeagus.
26 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 The possible grouping of the chalastogastrous famiHes on a basis
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 2/
28 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 Crampton (1919) gives illustrations of the phallic structure in 12
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 29
IXS)
The broad articulated parameres (Pmr) are heavily muscled, each bearing distally a large oval cupping disc (I,v) provided with a wider retractor muscle arising at the base of the paramere. The lateral sclerites of the aedeagus (E, pv) are each produced proximal to a short aedeagal apodeme (apa).
32 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99
ICHNEUMONOIDEA
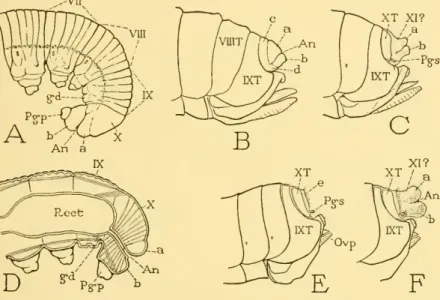
Only three families of Ichneumonoidea are discussed here, namely Ichneumonidae, Evaniidae, Braconidae, and of these the main attention is given to the last because the genital structures of ichneumonids were well described by Peck (1937) and their taxonomic value was discussed by Pratt (1939). The last fully exposed dorsal plate of the ichneumonid abdomen is. tergum VIII; tergal plates of segments IX and X either partially exposed behind tergum VIII or completely retracted below it.
34 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 posterior margin of the tenth tergum or tergites, but they are directly
CHALCIDOIDEA
The chalcidoid abdomen is variable in shape and in the relative size of its segments, as shown in Berecyntus bakeri (pi.8G), Callimome. I), Pteromalus piiparum (N), Tetrasticus bruchophagi (P), Leucospis afUnis (Q, R), Ormyrus sp. There are only two pairs of abdominal spiracles, the first on the pro-podium (pi. 8F, ISp), the other on segment VIII of the “post-.
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 37
CYNIPOIDEA
38 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99
SERPHOIDEA
NO. 14 MALE GENITALIA OF IIYMENOPTERA SNODGRASS 39
40 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 h) are united with the entire ventral surface of the aedeagus, though
FORMICOIDEA DoRYLiDAE
The last exposed segmental plates of the abdomen of male ants are tergum VIII and sternum IX (pi. ii A, I; pi. 12A, I, P; pi. 13D), but the membranous tenth segment may extend beyond the eighth tergum. It may be a transverse plate in the dorsum at the base of the tenth segment (pi. 12H), but generally it is strongly constricted medially (pi. The volsellae are usually hidden from below by the ventral lobes of the parameral plates (N , o), but they have the usual position against the inner surfaces of the basiparameral lobes (L).
Thevolsellae of the first species are of typical structure (pi. 12 N), and are freely exposed between the aedeagus and parameres (L, M). In the second species the volsellar cuspids appear to be wanting (pi. 13B), each volsellar plate {Iv) bearing only a large hooked digitus {dig) lying against the inside of the corresponding paramere {Pmr). In Pscudoniyrma gracilis the volsella is greatly reduced (pi. 13 G), the basal plate of each organ being a slender sclerite {Iv) attached to the lower edge of the corresponding parameral plate (//'), bearing only a single , small, setiger -ous lobe,the digitus {dig).
CHRYSIDOIDEA
99partial invagination of the parameral plates (//>) in the basal ring, partial invagination of the parameral plates (//>) in the basal ring, . and the shape of the broadly expanded, petiolate paranieres {Pmr). 13 G), the basal plate of each organ is a slender sclerite {Iv) attached to the lower margin of the corresponding parameral plate (//'), which bears only a single, small, setigerous lobe, the digitus {dig). The larger lobe of each pair (N, O, ciis), which is continuous from the small volsellar plate {Iv), is evidently the cuspis; the slenderer articular lobe {dig) is therefore the digitus, but in Holopyga (Q) it overlaps the base of the apical lobe and has a lateral position.
In Parnopes edwardsii the volsella are reduced to a single pair of small weak setae that project below the base of the broad aedeagus.
AIUTILLOIDEA
14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 43{0,apa) and prominent basal processes {e).. In Parnopes cdwardsii, {0,apa) and prominent basal processes {e). Apart from the absence of digitus, scolias volsella thus has the same character as Dasymutilla (F). Myzine quiuqiiecincta (F.).- Then the ninth abdominal sternum of this species (pi. 14M, IXS) is mostly exposed, as its distal part, proceeding from a small, concealed basal plate (O), consists of a long tapering spine that curves upwards behind the tip of the abdomen. of tergum VIII.
The small pygostyles of the tenth segment projecting out below its posterior marginale. The elongate parameres (Pmr) are produced distally from ventral angles of the parameral plates (Q, S), and the broad cuspid lobes of the volsellae (S,cus) project on the sides of the aedeagus above the parameres. Each has a broad basal plate (R,Iv) produced distal to a large cuspid lobe {cus) and armed on its mesal surface with a dense row of strongly sclerotized, finely toothed ridges. The digus {dig) is articulated dorsally on the volsal plate and bears on its ventral edge a series of strong spines.
XL BETHYLOIDEA
99ScolianohilifataF.—In this species the apex of the abdomen (pi.14 ScolianohilifataF.—In this species the apex of the abdomen (pi.14 G) is formed by tergum VIII above and sternum VIII below, the small three-pointed ninth sternum (G , H, IX) directly IX. above the eighth. Each volsella consists of a single large, strongly muscular lobe (L, ciis), the base of which clearly represents the volsella plate (Iv) because it contains a small intrinsic muscle (21).The parameral plates (P,Q, //') are so deeply separated from the long slender aedeagus (Aed) that their distal parts form a pair of independent basiparameral lobes.
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 45 species show the principal types of phallic structure found in the super-
VESPOIDEA
The exposed part of the male abdomen in Vespoidea ends with tergum VIII above and sternum IX below (pi.. pi. 17A), but sternum VIII is never visible externally, so that the ventriloquium of the "postabdomen" appears to have only seven sternal plates cor- responding (deceptively) with the seven exposed dorsal tergal plates. Sternum VIII is much reduced in size and hides above sternum VII (pi.. 15J) at the base of sternum IX, with which it is either closely connected by a membrane (pi. The phallus of Vespoidea is very variable in structure, but appears that there are no distinguishing features from the group, even though Vespidaethe volsellae attains the type of structure characteristic of this family.
The following examples will illustrate the structural variations and specializations of the vespoid phallus.
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 47
48 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 arm (c) prolonged from the dorsal surface between the lateral apo-
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 49
50 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 from an anatomical study, since all the muscles of each organ are
SPHECOIDEA
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 5
52 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99
APOIDEA
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 53 lobe to form a pair of free lateral arms commonly known as the
54 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 body of the organ, so that there are no free sagittal arms of the
The inconspicuous full cell layers are borne on the ventral edges of the parameral plates below the base of the aedeagus (E, Vol). The sternal plates of the genital region are variable in the degree of their modifications from relatively simple to complex forms, as seen in Anthophora ahrupta (pi. 26H,I,J), Diadasia australis (O, P), Tetralonia atriventris (U, V ), and Melissodes sp. The phallus (pi. 26 K,L,Q, S; pi, 27 C,D) is characterized by the articulation of the parameres on the parameral plates.
The sternal plates of the genital region in Nomada hella are relatively simple (Fig. 27F,G), although the sternum IX bears a long, median, distal alarm. The sagittae articulate on the parameral plates by their widely divergent bases (H), which are closely bridged by the base of the penis. In the usual condition (C), the genito-anal chamber at the end of the abdomen is approximately closed.
NO. 14 MALE GENITALIA OF HYMENOPTERA — SNODGRASS 57
58 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 groups, already noted among the bees in the melectid Triepeolus
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODORASS 59
The fallotrema is a large sub-terminal opening on the inner side of the penis (E, Phtr) and leads to a wide endophallic pouch (Enph). The eighth and ninth hidden sterna (pl.. 31 A, VIIIS, IXS) are closely connected to each other in the ventral part of the genital chamber. NO, 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 6l dorsal wall of genital chamber (pi.. 31 D, IXT) extending posteriorly to sides of proctiger (A').
The base of the penis contains a large dorsal plate (pi.. 31 E,ni) that laterally supports the bases of the sagittae. The opening in the sagitta from the inner part of the aedeagus is reduced to a small hole (pi. 30M) immediately behind the basal lobe (s), mesado from which the short aedeagal apodeme (apa) arises. The ninth tergum is reduced to a pair of small sclerites lying laterally on the dorsal wall of the genitalia.
62 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 The male genital organ of the Apidae is characterized by a reduc-
In incUifcra and indica a marginal ridge of the parameral plate (M, N,z^ li'^ is the connection of a connection between the sternum (N, IXS) and the tergite (IXT). The endophallus is a large and complex organ. in each of three species of Apis studied here The aphid phallus, developed in Apis Horca, appears to be of a generalized structure, as the penile valves are an integral part of the aedeagus.
The development of the ectophallus of Apis inclifcra was followed in detail by Michaelis (1900) and Zander (1900). The late larval instar shows a small oval genital disc with two slight thickenings on the ventral side of the ninth abdominal segment (Fig. 32F, G,gd). It is under the disc, in the envelope of the next stage, which develops inside the outer skin.
64 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 phallotreme between them ; the smaller lateral lobes are evidently
On the so-called "penis" of the bed bug (Cimex lecularius) and on the homologies in general of the male and female genital organs of this insect. Male genitalia and terminal abdominal structures, and terminal structures of chalastogastrous Hymenop larvae. Morphology and development of the genitalia and genital ducts of the Homoptera and Zygoptera as shown in the life histories of Philaenus and Agrion.
Structure and development of the reproductive system of the Coleoptera with notes on its homologies.
70, SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99
EXPLANATION OF PLATES
NO. 14 MALE GENITALIA OF HYMENOPTERA — SNODGRASS 73
74 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99
NO. 14 MALE GENITALIA OF HYMENOPTERA — SNODGRASS 75
76 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99
NO. 14 MALE GENITALIA OF HVMENOPTERA — SNODGRASS ']']
78 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99 Plate 15
NO. 14 MALE GENITALIA OF IIYMENOPTERA — SNODGRASS 79 Plate 17
82 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99
14 MANLIKE GENITALIA VAN IIYMENOPTERA SNODGRASS 83G, Hylaeiisbasalis (F. Sin.), agthabdominale tergum, ventral en dorsale wand G, Hylaeiisbasalis (F. Sin.), agthabdominale tergum, ventral en dorsale wand.
84 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99
NO. 14 MALE GENITALIA OF HYMENOPTERA SNODGRASS 85
86 SMITHSONIAN MISCELLANEOUS COLLECTIONS VOL. 99
CHAUASTOG ASTRA
CHALASTOGASTRA. ICHNEUMONOIDEA
ICHNEUMONOIDEA
VESPOIDEA
SPHECOIDEA
APOIDEA
IXS"