YENDAHULUAN
Penelitian dan pengembangan dalam bidang bioteknologi reproduksi yang bertujuan untuk meningkatkan populasi dan mutu genetik ternak
kini
menjadi salah satu prioritas utama dalam pembangunan petemakan di lndonesia. Untuk mengembangkan bioteknologi reproduksi di lndonesia perlu dhkukan penelitian- penelitian dalam rangka penguasaan atau alih teknologi dari peneliti-peneliti di negara maju sehingga hasil penelitian yang dikembangkandi
lndonesia diharap-kan dapat didayagunakan untuk meningkatkan populasi
clan
memperbaiki mutu genetik ternak, khususnya sapi di lndonesia yang pada tahun 1996 populasinya tercatat 11,930,000 ekor sapi potong dan 343,000 ekor sapi perah (Wiryosuhanto, 1997).Setelah bioteknologi inseminasi buatan dan transfer embrio berkembang dengan baik di lndonesia (khususnya pada sapi), pengembangan bioteknologi reproduksi pada ternak dalam kurun waktu lima tahun terakhir ini telah mernasuki era baru yaitu memasuki era pengembangan teknologi fertilisasi in vino (FlV). 'I'eknologi FIV berpotensi untuk meningkatkan daya reproduksi sapi betina, baik semasa maupun setelah habis masa produksinya (diafkir). Oosit yang diambil
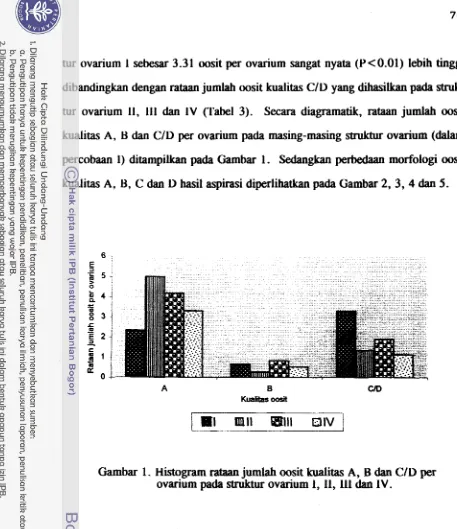
dari ovarium sapi yang berasal dari rumah potong hewan (RPH) &pat dimanfaat-
teknik mikrornanipulasi pada embrio seperti teknik cloninglsplitting untuk memproduksi anak-anak sapi kembar identik dan teknik rekayasa genetik, seperti teknik transfer gen untuk memproduksi sapi transgeniktkhimera serta pengem- bangan penelitian-penelitian dasar dan terapan lainnya. Keberhasilan dalam pembuatan domba klon Dolly dan domba transgenik Polly di lnggris, yang menggemparkan dunia baru-baru ini, tidak terlepas dari pengaplikasian teknologi tertiliasi in vitro.
Menurut Gordon (1W4) ada tiga potensi pengaplikasian teknologi FlV secara bersamaan dengan teknologi 'I'E pada sapi. Pertama, embrio sapi pedaging bangsa Charolais, Limousin dan Simmental yang dihasilkan melalui teknologi FIV dapat ditransfer pa& induk sapi resipien untuk menghasilkan seekor anak sapi pedaging berkualitas tinggi. Yada induk sapi resipien dengan potensi pro- duksi susu yang tinggi (termasuk sapi perah yang mempunyai produksi susu rendah) dengan kondisi pakan dan manajemen yang layak, embrio sapi bangsa HerGord dan Angus yang dihasilkan melalui teknologi FIV &pat digunakan untuk meningkatkan frekuensi kelahiran kembar. Kelahiran kembar dua pada sapi dapat meningkatkan efisiensi biologis dan ekonomi produksi sapi pedaging
semua aspek pembekuan oosittembrio, studi tentang pertumbuhan dan perkem- bangan embrio, pengujian efektivitas embryonic stem cells (ESC) dan pengem- bangan prosedur injeksi gen dan lain-lain. Penerapan teknologi produksi embrio in vitro di laboratorium bersamaan dengan teknologi TE telah memberikan manfaat yang besar &lam meningkatkan pengetahuan di bidang biologi perkem- bangan dan biologi molekular.
Perkembangan teknologi FIV dalam kurun waktu Lima tahun terakhir ini, khususnya di Indonesia, masih berskala percobaan
di
laboratorium. Kendala utama dalam pengembangan teknologi FIV adalah belum sempurnanya kondisi medium yang digunakan untuk pematangan oosit dan kultur embrio in vitro, sehingga produksi embrio melalui teknik tersebut masih rendah dan bervariasi. Salah satu upaya untuk mengatasi kendala tersebut dilakukan pengujian efektivitas penambahan berbagai jenis dan konsentrasi serum sapi serta kokultur selapis sel- sel epitel tuba b'allopii dan kumulus pa& TCM-1YY untuk meningkatkan kemam- puan pematangan oosit dan perkembangan embrio sapi in vitro. Suplementasi serum 5-20% masih dibutuhkan di dalam media pematangan oosit dan kultur embrio in vitro, walaupun sebagian besar kebutuhan nutrisiuntuk
pematangan oosit dan perkembangan embrio sapi in vifro telah tercukupi oleh penggunaan media kompleks seperti tissue culture medium 1W (TCM-199). Pengujian poten- si jumlah dan kualitas oosit yang &pat diaspirasi dari berbagai struktur ovarium dan pengujian potensi spermatozoa sapi yang berasal dari semen segar dan semen beku perlu pula dilakukan untuk m e n g h a s i b embrio daiam jumlah yang tinggi.makro globulin, transferrin dan fetuin), polipeptida, asam lernak (seperti koles- terol) dan mineral; berbagai macam hormon (seperti hormon insulin, steroid dan
lain-lain) serta berbagai macam faktor penumbuh (Malole, 1990; Freshney, 1987; 'I'rounson 1992). Menurut Freshney (1987) albumin bermanfat untuk mengatur t e W osmotik dan penyangga pada medium, fibronektin berguna untuk merangsang perlekatan sel
dm
a-2 makro globulin &pat berfungsi meng- hambat kerja enzim proteolitik (seperti tripsin). Fetuin yang terdapat di &lam serum fetus m a 1 bovine serum atau FBS) berperan dalam meningkatkan perlekat- an sel, mencegah pecahnya zona pelusida dan meningkatkan kemampuan fertilisa- si oosit in vitro (Schroeder et al., 1990; Zhang et al., 1991a; Yamazaki dan Ishibashi, 1990).Menurut Y ounis et al. (1989), Lu et al. (1987) dan Henricks et al. (1972) kandungan hormon (khususnya hormon steroid) di dalam serum sapi sangat bervariasi dan dipengaruhi fase siklus estrus. Serum yang diambil pada hari sapi estrus (SHO) mengandung estrogen yang tinggi, sebaliknya
serum
yang diambil pada hari ke tujuh setelah estrus (SH7) mengandung progesteron yang tinggi. Sedangkan keberhasilan pematangan oosit dipengaruhi oleh keseimbangan atau rasio estrogen dan progesteron di dalam medium permtangan oosit in vitro (Younis et al., 1989; Gordon, 1994; Lorenzo et al., 1997). Namun para peneli- ti tersebut menyatakan bahwa peranan progesteron secara fisiologik dalam proses pematangan omit belum diketahui secara jelas.krdasarkan informasi tersebut di atas perlu dilaloukan pengujian efektivi- tas penambahan jenis serum FBS, S& dan SH7 pada konsentrasi 5, 10, 15 dan
ratorium sendiri) pa& konsentrasi yang diujikan dapat menggantikan peranan FBS di dalam media pematangan oosit dan kultur embrio in virro. Serum FBS sebagai bahan impor biologik, memiliki potensi pembawa bibit penyakit eksotik ke lndonesia yang sulit terdeteksi disebabkan oleh virus, mikoplasma dan klami- dia, disamping ketersediaan FBS yang tidak kontinyu di pasatan &ngan harga yang semakin rnahal. Untuk mencapai maksud tersebut di atas dilakukanlah penelitian ini dalam empat seri percobaan dengan judul : "Efektivitas Penambah- an Berbagai Jenis dan Konsentrasi Serum serta Kokultur Sel-Sel Tuba Fallopii dan Kumulus pada
'EM-199
dalam Produksi Embrio Sapi In Vitro". 'Tersedia nya serum secara bersinambung sepanjang tahun dengan harga yang murah danterhindar dari resiko terkontaminasi oleh bibit penyakit yang tidalr ada di lndone- sia serta berdayaguna meningkatkan produksi embrio di laboratorium diharapkan menjadi salah satu faktor penting bagi perkembangan bioteknologi dalam bidang reproduksi ternak, khususnya teknologi FlV
,
di Indonesia.Hipotesis
Hipotesis yang diuji melalui serangkaian percobaan di dalam penelitian ini adalah sebagai berikut :
(1) Ovaria yang memiliki folikel dominan (FD) menghasilkan rataan jumlah oosit kualitas A dan B yang lebih tinggi dibandingkan dengan struktur ovarium lainnya.
(3) Kemampuan spermatoma yang berasal dari semen segar untuk memfertilisasi oosit in vitro lebih tinggi dibandingkan dengan spermatozoa yang berasal dari semen beku.
(4) Penggunaan kokultur selapis sel-sel epitel tuba Fallopii
dan
kumulus sapi dapat meningkatkan kemampuan perkembangan embrio in vitro.'I'ujuan yang ingin dicapai di dalam penelitian ini adalah sebagai berikut : (1) Mengetahui jumlah oosit kualitas A dan f3 yang dapat diaspirasi per ovarium
pada masing-masing struktur ovarium.
(2) Mencari jenis dan konsentrasi serum yang terbaik di antara jenis serum FBS, SHO dan
SH7
pada tingkat konsentrasi 5, 10, 15 dan 20% untuk pematangan oosit dan kultur embrio in vitro.(3) Menguji pengaruh kemampuan spermatozoa yang berasal dari jenis semen segar dan semen beku terhadap angka fertilisasi dan perkembangan embrio in vitro.
(4) Menguji pengaruh medium kokultur selapis sel epitel tuba Fallopii atau sel kumulus terhadap perkembangan embrio in vitro.
Manfaat yang diharapkan dari hasil penelitian ini adalah :
(1) Serum dapat diproduksi di laboratorium sendiri untuk kegunaan operasional FIV secara rutin, sehingga pengaplikasian teknologi F1V dapat dikembangkan secara lebih luas guna meningkatkan produktivitas temak (sapi) di lndonesia
.
banyak lagi oosit yang dirnatangkan
in
vitro.(3) Dapat menghasilkan embrio pada berbagai taraf perkembangan untuk keper- luan penelitian dan pengembangan bioteknologi yang lebih mutakhir dalam bidang reproduksi ternak, ataupun untuk ditransfer pada induk sapi resipien. (4) Dapat meningkatkan daya reproduksi sapi-sapi betina baik semasa maupun
setelah habis masa produksinya (diafkir) dan dipotong di KPH.
(5) Dapat menghasilkan data dan informasi baru bagi penelitian-penelitian selan- jutnya, khususnya mengenai potensi jumlah dan kualitas oosit yang dapat
'TINJAUAN YUSTAKA
Keberhasilan Teknologi Pertilisasi In Vitro pa& Sapi
Istilah in vitro dalam bahasa Latin berarti di luar tubuh. Ketika percobaan- percobaan fertilisasi di luar tubuh pertama kali dikerjakan, kedua macam gamet ditempatkan di dalam tabung atau cawan petri, dan semenjak itu istilah in vitro populer digunakan (Seidel dan Elsden, 1989).
Sreenan (1970) termasuk salah satu peneliti awal yang mencoba melaku- kan fertilisasi in vitro (FIV) pada sapi. Oosit sapi yang dimatangkan in vitro difertilisasi dengan sperma sapi yang telah diinkubasi di dalam medium yang mengandung enzim a-amilase. Namun keberhasilan FIV pertama pa& sapi dilaporkan oleh lritani dan Niwa (1977) di Jepang. Beberapa tahun kemudian (9 Juni 1981) lahir anak sapi pertama yang berasal dari oosit yang diovulasikan dan difertilisasi in vitro di Amerika Serikat (Brackett et al., 1982). L)ua ekor anak sapi hasil FlV lainnya dilahirkan di Georgia pada tahun 1983 (Brackett et al., 1984). lli Kanada, dihasilkan enam ekor
anak
sapi hasil FIV yang berasal dari oosit sekunder yang diambil dengan teknik laparoskopi (Lambert et al., 1983). 'I'eknologi FlV untuk menghasilkananak
sapi tersebut di atas menggunakan salur- an telur kelinci sebagai sistem kultur embrio in vivo.Yotensi Penggunaan 'I'eknologi Pertilisasi In Vitro pada Sapi
'I'eknologi fertilisasi in vitro (FIV) merupakan teknik yang efektif untuk menghasilkan embrio di laboratorium dalam jumlah besar dengan biaya yang relatif murah. Hal ini telah mendorong pengaplikasian teknologi transfer embrio
Kedua, teknologi F l V berpotensi untuk meningkatkan produktivitas susu pada sapi perah. Teknologi FIV dapat menghasilh embrio menggunakan ovaria sapi (perah) bermutu genetik tinggi setelah masa produksinya habis dan diotong di KPH. Silsilah (pedigree) dari beberapa anak sapi yang berasal dari sejumlah embrio ini mash dapat dipertahankan. Di samping itu, teknologi FIV memung- kinkan embrio dihasilkan dari oosit sapi perah bermutu genetik tinggi yang masih hidup, baik yang mempunyai siklus estrus maupun yang bunting pada peride awal (dan kadang-kadang dari sapi-sapi dara sebelum pubertas), dengan menggu- nakan teknik aspirasi folikel melalui vagina yang dipandu dengan peralatan ultra- sonografi. Moreno et a1 (1993) melaporkan bahwa induk sapi bunting mengha- silkan jumlah oosit yang lebih tinggi dari sapi-sapi yang tidak bunting. Embrio yang dihasiUran dengan teknik F l V ini &pat dijadikan sebagai alternatif untuk mengatasi beberapa kelemahan yang a& pada perlakuan-perlakwn superovulasi pada sapi untuk mendapatkan embrio m a konvensional seperti respons supero- vulasi yang bervariasi, jumlah dan kualitas embrio yang juga bervariasi di antara individu dan sebagainya.
tentang cloning pada sapi, pengujian keefektivan embryonic stem cells, pengem- bangan prosedur injeksi gen, dan penelitian berbagai
aspek
pengenalan induk (maternal recognition) terhadap kebuntingan.Potensi dan Perkembangan Oosit pada Ovarium Sapi
Populasi folikel primer yang mengandung bakat sel telur (gem cells atau oBgonium) pa& ovarium rnarnalia mu&, khususnya sapi, sudah tertentu sesuai dengan sifat genetik masing-masing temak yang merefleksikan potensi dari materi genetik yang dapat dieksploitasi (Hunter, 1980 dalam Xu, 1987). Folikel primer tidak dibentuk selama kehidupan dewasa ('Toelihere, 1985a). Setelab pubertas terjadi pertumbuhan dari folikel primer mejadi folikel sekunder, tetapi tidak
sernua folikel primer berkembang menjadi folikel sekunder. Menurut Errickson
(1 966) jumlah folikel primer yang terdapat pa& ovaria sapi potong relatif konstan (dengan rataan 133,000) sampai sapi tersebut berumur empat sampai enam tahun, dan menurun secara cepat menjadi no1 pada umur 15 sampai 20 tahun. Walaupun jumlah folikel primer sangat besar, tetapi tidak lebih dari 300 yang mencapai fase ovulatoris. Menurut Lusier et al. (1987) ha1 tersebut disebabkan karena sebagian folikel pada ovaria mengalami atretik dalam berbagai fase perkembangannya. Pembentukan antrum pada sapi muncul pada berbagai kisaran diameter folikel (0.14 sampai 0.28 mm). Dijelaskan pula bahwa pertumbuhan awal folikel dise- babkan oleh peningkatan jumlah sel-sel granulosa clan peningkatan permukaan lapisan granulosa.
folikel, dan sel-sel kumulus yang menyelimuti oosit. Suatu jembatan sel-sel granulosa menghubungkan oosit yang diselimuti sel-sel kumulus dengan membra- na sel-sel granulosa di sekeliling folikel. Kedua tip sel ini diketahui berbeda dalam distribusi reseptor-reseptornya dan kharateristik steroidogeniknya. Pada folikel yang belum berovulasi, kedua t i p sel ini juga mempunyai respons yang berbeda terhadap gonadotropin. Gonadotropin merangsang sel-sel kumulus untuk memproduksi dan mensekresikan asam hialuronik tetapi tidak berpengaruh sama sekali terhadap sel-sel granulosa. Dengan kata lain hanya sel-sel kumulus yang mempun yai reseptor terhadap gonadotropin.
menghasilkan androgen, tetapi hanya sedikit yang berkemampuaa mentransforma- si androgen menjadi estrogen. Terdapat bukti bahwa perolehan sistem aromatase, yang diatur oleh follicle stimulating hormone
(FSH),
mungkin menrpaluur faktor penentu dalam seleksi dan pembentukan folikel dominan (FD).Pemahaman tentang mekanisme yang mengatur pertumbuhan folikel sampai ovulasi sangat penting artinya bagi pematangm oosit
sapi in
vitro. Lebih dari 30 tahun yang lalu, Rajakoski (1960) telah melaporkan bahwa perkembangan folikel pada sapidara
muncul dalam dua gelombang. Gelombangpertama
dimu-
lai pada hari ketiga dan berakhir pada pertengahan siklus, menghadlm satu folikel dominan (PI)) yang kemudian mengalami regresi. Gelombang kedua terjadi setelah pertengahan siklus dan berakhir menjadisebuah
folikel yang diovu- lasikan. Assey et al. (1993, 1994) menyatakan bahwa siklusestrus
pada sapi secara normal ditandai oleh dua atau tiga gelombang pertumbuhandan
perkem- bangan folikel, yang masing-masing menghasilkan satu FD dan beberapa buah folikel pendamping.ditentukan oleh lamanya regresi sel-sel luted. Pertumbuhan folikel pa& sapi dikontrol oleh endokrin dan parakrin, dan faktor-faktor lokal lainnya seperti steroid, faktor penumbuh, cytokines dan molekul pengatur lainnya. Faktor-faktor tersebut bereaksi pada sel-sel ovarium disebabkan oleh pengaturan respons terha- dap gonadotropin yang bersirkulasi di dalam darah.
Selanjutnya Henricks et al. (1972)
dan
Kajakowski(1960)
menyatakan bahwa terdapat hubungan yang sangat erat antara level estrogen dengan perkem- bangan folikel selama siklus estrus sapi. Folikel berdiameter 5,Omm
atau lebih muncul pada hari ketiga atau keempat, dan digantikan oleh sebuah FD (berdiame- ter 9- 1 3 mm) pada hari ke- 12. Semenjakhari
ketiga atau keempat setelah estrus, level estrogen plasma meningkat dari 3-4 pg/ml menjadi 10 pg/ml pada hari kesembilan.Menurut 'l'oelihere (1985a), dalam proses perkembangan dari folikel ter- tier menjadi folikel yang akan diovulasikan, antrum diisi oleh cairan jernih (liquor folliculi) yang banyak mengandung protein dan estrogen. Estrogen mungkin disekresikan secara langsung oleh sel-sel teka interna
ke
dalam
folikel melalui suatu selubung dasar, mernbrana propria, yang memisahkan teka interna dari membran granulosa. Belum banyak diketahui apakah serum yang banyak mengandung estrogen atau hormon steroid pa& umumnya dapat meningkatkan keberhasilan pematangm oosit dan perkembangan embrio in vitro.Pengambilan Oosit dari Ovarium Sapi
Umur, Siklus Estrus
dan
Kebuntinganpenelitian Lonergan (1990) yang disitir Gordon (1994)
dan
Moreno etd.
(1992) menyatakan bahwa jumlah oosit yang dihasilkan induk sapi lebih rendah di- bandingkan dengan sapi dara yang belum pernah dikawink;aa. Ditambahkan bahwa ovaria sapi dara tanpaCL
menghasilkan jurnlahoosit
kualitas baik paling tinggi. Mermillod et al. (1992a) membukthn bahwa induk sapi yang lebih mu& (berumur satu sampai tiga tahun) menghasilkan jumlah blastosis hasil F1V yang lebih tinggi dibandingkan dengan induk sapi yangberumur
lebih tua.Hasil penelitian yang agak berbeda dilaporkan oleh Zhang et
d.
(1991), yaitu jumlah oosit dengan kualitas baik lebih tinggi pada ovariainduk
sapi di- bandingkan dengan ovaria sapi dara. lnformasi demiHan menimbulkan perta- nyaan, mungkin status nutrisi ternak lebih berpengaruh terhadap jumlah dan kualitas oosit per ovarium dibandingkan dengan umurdan
paritas. Sebagai contoh, induk-induk sapi perah yang diafkir dari perusahaan petemakan sapi perah jelas mempunyai kondisi tubuh tidak sebaik sapid a m
yang disiapkan khusus untuk memenuhi kebutuhan daging di pasaran.ditandai dengan beberapa gelombang pertumbuhan folikel selama satu siklus estrus pada sapi. Seidel dan Elsden (1989) juga menyatakan bahwa fase siklus estrus tidak terlalu berpengaruh terhadap jurnlah oosit yang diaspirasr.
Menurut Machatkova et al. (1993) dalam Oordon (1994) oosit yang diam- bil dari ovaria sapi pada hari 14 sampai 16 siklus estrus mempunyai potensi perkembangan yang lebih tinggi dari pada oosit yang diambil pada hari ketujuh sampai kesembilan atau fase siklus estrus yang tidak diketihi. Dijehkan lebih lanjut bahwa perbedaan pengaruh hi mungkin berkaitan deugm proporsi relatif t;D yang ditemukan pada fase siklus yang berbeda. Namun demikian Tan dm Lu (1990) yang menguji pengaruh tiga fase siklus estrus (fase luteal awal, fase luted dan fase luteal akhir) terhadap angka cleavage dan angka blastosis menyimpulkan bahwa ketiga fase siklus estrus yang diuji tersebut tidak berpengaruh nyata terha- dap angka cleavage dan angka blastosis.
Hasil penelitian Vajta et al. (1992) menujukkmt bahwa, sebagai sumber oosit untuk FIV, induk sapi bunting dapat dianggap sama dengan sapi-sapi yang bersiklus. Sedangkan menurut Moreno et a1 (1993) induk sapi bunting mengha- silkan jumlah oosit yang lebih tinggi dari pada sapi-sapi yang tidak bunting. Bangsa, Nutrisi dan Variasi lndividn Sapi
Jumlah omit yang &pat diambil dari ovaria sapi juga dipengaruhi oleh va- riasi bangsa, nutrisi dan individu ternak sapi. Gordon (1994) menyatakan bahwa sebagian sapi-sapi bangsa persilangan Charolais, Limousin dan Simmental
Maurasse et al. (1985) menemukan bahwa pakan
dengan energi tin&
mempercepat laju pertumbuhan dm pergantiaa folikel. Padg sapi potong setelah melahirkan (postpartum), sapi-sapi yang diberi pakan dengan mtrki ymg rendah mempunyai folikel antral yang lebih sedikit dari pada sapi-sapi yang diberi pakandengan nutrisi yang
tin@
(Prado et al., 1989 d a h t Gordon, 1994). l h h m jangka waktu yang pendek, penurunmt lcuantitas dm kualitas pakan p d a sapipotong muda tidak mempengaruhi populasi folikel padg wariumnya. Dafam kondisi seperti itu hanya level
insulin-like
growth factor 1 (1GF-1) p h yang menurun secara nyata (Spicer et al., 1992).Perbedaan kdmgm
proteinkasar
dalam pakan yang sangat besar dapat m e m p e n g d perkembaqpn folikeldan
perkembangan embriotaraf
selmjutnya (Gar&-&rjdil ezd.,
1991).Adanya p g a r u h variasi individu sapi terhadap p d i h oosit dau jumtgh embrio yang dihasilkan melalui teknik FlV dilaporkan oleh MemiUod et al. (1992b). Yara peneliti lain umumnya menyatakan bahwa variasi jumlah embrio yang dihasilkan melalui teknik FIV sebanding dengan variasi jumlah embrio yang dihasilkan melalui perlakuan-perlakuan superovulasi pada sapi.
'Teknik Pengambilan Oosit dari Ovaria Sapi
kemiringan (bevel) dan ketajaman ujung jarum serta besarnya tekamm.apirasi. Umumnya peneliti menggunakan ukuran jarum 18 O untuk mengasphi w i t dari folikel berdiameter dua sampai enam mm pada ovarium sapi yang berasal dari W H (Ball et al., 1983). Menurut Yurwantara (1994) sudut kemiringan atau keruncingan ujung jamm yang disarankan adalah 25 sampai 45 derajat tergantung ukuran folikel yang diaspirasi. Sudut kemitingan yang besar cocok untuk folikel kecil karena dapat menghindari kebocoran clan keluamya cairan folikel, sedang- kan sudut yang kecil lebih cocok untuk folikel besar. D i mbahwa aspira- si dapat dilakukan dengan menggunakan alat vakum kompresi atau dengan penye- dotan manual. 'Manan yang digunakan bervariasi antara emun sampai 36 rnl air per menit. Peneliti lain menggunakan t e b 30 sampai 40 mf air per menit tergantung ukuran diameter jamm. Sehjutnya dijelaskan bahwtl diperlukan pengkajian yang lebih intensif tentang tekanari aspirasi yang optimal tanpa menimbulkan pengaruh negatif terhadap kompleks mit-kumulus.
Katska (1984) berhasil mengambil 43% w i t dari ovaria dengan te- aspirasi dan 100% dengan teknik diseksi. Hanya 45% dari oosit yang diambil dengan teknik aspirasi merniliki morfologi normal. Leibfried-Rutledge et al. (1985) berhasil mengambil 60% oosit dari ovaria dengan rataan lima sampai delapan oosit berkualitas baik per pasang ovarium. Suss et al. (1988) mendapat- kan rataan 23 oosit per pasang ovarium menggunakan teknik aspirasi, dengan 47% oosit (10.6 per pasang ovarium) yang memiliki sel-sel kumulus utuh. Menurut Vergos (1990) dalam Gordon (1994) hanya 50 sampai 60% oosit yang dapat diambil dengan teknik aspirasi (menggunakan jarum berukuran 18 G)
Menurut Godon dan Lu (1990) dan Lu et al. (1987) rataan jumlah oosit yang diperoleh dengan teknik aspirasi 6.9 oosit/ovarium (dari 646 ovaria) dan dengan teknik penyayatan adalah 6.5 oosit/ovarium (dari 892 ovaria). Sedan*
Sergejew dan Baibekow (1992) dengan teknik aspirasi mendapatkan 507 kom- pleks oosit-kumulus dari 121 ovaria (dengan rataan 4.2 wsit/ovarium) sapi FH yang berasal dari KYH.
Keuntungan penggunaan teknik aspirasi folikel adalah pengambilan oosit dapat dilakukan dengan cepat, tiga kali lebih cepat dibandingkan dengan teknik diseksi. Kecepatan pengambilan oosit sangat penting
artinya dalam
sistem pro- duksi embrio secara komersial (Gordon, 1994).Untuk mengatasi kelemahan teknik aspirasi tersebut di atas (jumlah oosit yang diperoleh relatif rendah) penusukan jarum dilakukan pada bagian stroma ovarium terlebih dulu kemudian baru menusuk folikel demi folikel sekaligus (YuMlantara, 1994). Cara tersebut bertujuan untuk menghindari terlontarnya oosit ke luar folikel bersama cairan folikel, mempersingkat waktu kerja, dan memperkecil peluang terjadinya kontarninasi.
'I'eknik Penyayatan Ovarium. Menurut Suss dan Madison (1983) 20 sampai 30 oosit dapat diambil dari ovarium sapi dengan menggunakan teknik penyayatan. Xu et ul (1992) melaporkan bahwa dari dua ekor induk sapi dapat dihasilkan rataan 55 oosit per pasang ovaria dengan teknik penyayatan. Menurut Carolan (1993, 1994) teknik penyayatan menghasilkan perolehan oosit tiga kali lipat lebih banyak dibandingkan dengan
teknik
aspirasi. Ditambahkan bahwa oosit yang diperoleh dengau teknik penyayatan dan aspirasi mempunyai kemam- puan yang sama untuk dimatangkan in vino. Penggunaan teknik penyayatan dapat dikombinasikan dengan teknik aspirasi. Namun demikian Takagi et al. (1992) menyatakan bahwa penggabungan teknikaspirasi
dengan teknik penyayat- an tidak diperlukan untuk mendapatkan jumlah atau W t a s oosit yang tinggi dari ovaria sapi.Carolan et al. (1993) melaporkan bahwa bahwa dengan rnenggtmkan tekoik penyayatan pada 12 pasang ovaria sapi dihasillran rataan jumlah blastosis 15.4 per pasang ovaria. Teknik penyayatan mernbutuhkan walrtu tiga kali lebih
lama dibandingkan dengan teknik aspirasi, walaupun dalam waktu yang sama teknik penyayatan menghasilkan jumlah oosit yang sama dengan teknik aspirasi (dari jumlah ovaria yang lebih sedikit). Bila jumlah ovaria yang tersedia relatif banyak maka teknik aspimi lebih tepat untuk digunakan.
Medium Pengambilan Oosit
dari
Ovaria Sapimedium yang mefigandung konsentrasi Ca2+ b MgZ' suboptimal seperti PBS, dalam jangka waktu yang pendek, dapat menimbuhn pengaruh yang merusak terhadap oosit itu sendiri.
Suhu dan Batas Waktu Penyimpanan Ovaria
Umumnya oosit diambil dari ovaria dalam waktu satu sampai dua jam setelah pemotongan sapi, dengan suhu penyimpanan sekitar
WC
(Schleger et al., 1990; Sekine et al., 1992). Oosit yang dikumpulkan pada suhu jauh di bawah 30°c mengalami pematangan inti yang tidak sempwna (Firstdan
Parrish, 1987a). Penunrnan suhu oosit yang diaspirasi dari folikel sampai jauh di bawah suhu fisiologik tubuh (suhu rektal sapi 38 sartlpai 39,3Oc) berpen%aruh mgatif pada viabilitas oosit, dalam bentuk abnormalitas pada semua fase meiosis (Lenz et al., 1983; Pollard et al., 1996). Namun hal ini masih menjadi pcakbtm, sebab oosit yang diasphsi pada suhu 2 0 0 ~ tidak selalu memiliki viabilitas yang rendah setelah difdisasi in vino atau setelah embrio dikulturin
W o (Zhang et al., 1990). Kruip dan Ferwoy (1982) menyatakan bahwa p e n g m b h oosit pada suhu 18 sampai 21°c mentpakan yang p a h g layak.Zhang et al. (1990) meneliti pengaruh tiga tingkat suhu ovaria selama transportasi dari RPH ke laboratorium &&a@ produksi embrio in vitro. Ovaria yang c&mmphn pada suhu no1
sampai
dua OC men- mgka fertilisasidan perkembangan embrio yang nyata lebih redah dibandingbn dengan ovaria yang ditraospotkan pada suhu 18 sampai 20% atau 30 sampai 32%. Lenz et
al.
Pendinginan ovarium sapi (no1 sampai dua OC) sesaat akan diproses dapat menimbulkan kerusakan yang lebih tinggi, walaupun oosit &ngan sel-sel kumulus yang utuh yang didinginkan sampai suhu kamar (18 sampai 20'~) selama di- proses di laboratorium tidak menimbulkan akibat seperti yang terjadi pada pendi- nginan ovarium tersebut di atas.
Pollard et al. (1996) mengaspirasi oosit selarna 30 menit pada suhu 2 5 O ~ , 30°c dan 35OC, kemudian diproses pada suhu lebih tinggi atau sama dengaa 35OC. Hasil penelitian menunjukkan bahwa suhu yang readah berpengaruh nyata menghambat perkembangan produksi embrio, walaupun angka fertilisasi in vitro tidak lebih rendah dibandingkan dengan suhu kontrol (suhu aspirasi 35'~). Persentase blastosis pada suhu kontrol 33.7 %
,
lebih tinggi dibandingkan dengan 22.4 dan 19.5 % pada suhu 2 5 ' ~ dan 30'~.Data hasil penelitian Yang et al. (1990) menunjukkan bahwa angka clea-
vage dan angka blastosis yang tinggi dihasilkan hanya jika ovaria disimpan pada suhu 2 4 ' ~ dan 3 7 ' ~ selama no1 sampai empat jam. Walaupun penyimpanan ovaria selama delapan jam masih memberikan angka cleavage dan angka blastosis yang cukup tingggi, tetapi hams melakukan penurunan suhu menjadi 24Oc. Kematian ternak secara cepat akan diikuti oleh perubahan-perubahan degeneratif pada ovaria, dan perubahan-perubahan tersebut berlangsung lebih cepat pada suhu tubuh (30°C sampai 37'~) dibandingkan suhu yang lebih rendah (20°c sarnpai 25'~).
lndikator Kualitas Oosit
kualitas oosit secara morfologik dewasa ini didasarkan
pada
kekompakm dan
kuantitas sel-sel kumulus yang mengelilingi oosit (Looset
ul., 1989). KlasShsi oosit berdasarkan kriteria morfologi menurut Loos etul.
(1989) adalah sebagai berikut : (1) Oosit dikelilingi oleh multi lapisan kumulus yangkompak,
ooplasma homogen, kompleks oosit-kumulus (KOK) secara keseluruhan terlihat terang dan transparan; (2) Oosit dikelilingi oleh multi lapisan kumulus yang kompak, oo- plasma homogen tetapi dengan penampilan yangkasar, daerah
pinggiran oosit yang lebih gelap, dan KOK secara keselmhan terlihat lebih gelap dan kurang transparan; (3) Oosit dikelilingi oleh sel-sel kumulus yang kurangkompak,
oo- plasma tidak beraturan dengan bercak-bercak hitam, dan KOK lebih gelap lagi dari klasifikasi (1) dan (2); (4) Sel-sel kumulus yang mengelilingi oosit mengem- bang, sel-sel kumulus terpencar-pencar membentuk gumpalan-gumpalan gelap yang kental, ooplasma tidak teratur membentuk gumpalan-gumpalan berwarna gelap, dan KOK secara keseluruhan menghitam dan tidak teratur. Monaghan (1993) dalam Gordon (1994) menyatakan bahwa kehomogenan ooplasm tidak selalu dapat dipedornani sebagai kriteria seleksi oosituntuk
dimatangkan in vitro. Klasifikasi oosit dengan nilai prediksi yang baik sangat penting artinya dalam sistem produksi embrio in vitro.Ukuran Folikel dan Kualitas Oosit
Kisaran ukuran folikel yang diambil dari oosit sapi untuk kegiatan FIV umumnya dua sampai enam mm. Tan dan Lu (1990) membuktikan adanya kore- lasi antilra ukuran folikel dan kualitas oosit. Oosit yang diambil dari folikel berdiameter kurang dari 2 mm menghasilkan angka cleavage dan embrio yang lebih rendah dibandingkan dengan oosit yang diambil dari folikel berdiameter dua sampai enam mm, dan oosit yang diambil dari folikel berdiameter lebih dari enam mm menghasilkan embrio yang lebih tinggi. Hal yang sama juga dinyata- kan oleh Fuhrer et al. (1989) dan Pavlok et al. (1993)
Assey et al. (1992, 1994) membandingkan lingkungan daa stmktw oosit yang berasal dari FD dengan dua folikel pendamping yang terbesar, yang diuji beberapa hari setelah ovulasi. Para peneliti ini menyatakan bahwa oosit di dalam FL) yang lingkungannya didorninasi oleh estrogen mempunyai sel-sel kumulus yang sehat, sedangkan oosit di dalam folikel pendamping yang lingkungannya di- dominasi oleh progesteron mempunyai sel-sel kumulus yang telah berdegenerasi.
Yematangan Oosit
In
VitroPisiologi Yematangan Oosit
Semenjak kehidupan fetal atau neonatal, oosit mamalia telah mengalami pembelahan meiosis 1, dan perkembangan inti sel tertahan pada tingkat dictyate
pecahnya selubung inti (gemiml vesicle breakdown =
GVBD)
diiringi oleh khromosom yang berkondensasi menjadi bentuk yang Irompak, pembentukm spindle meiosis yang menurut gilirannya diikuti oleh keluarnya badan kutub(polar body) 1 dan pembentukan cakram metafase 11 (M-ll) (Jagiello et al., 1975). Di samping khromosom badan kutub I mengandung berbagai organel termasuk mitokhondria, ribosom dan granula kortikal (Gordon
,
1994).Lanzendorf et al. (1990) menyatakan ha1 yang serqa bahwa dalam per- kembangannya sel telur dapat dibedakan antara stadium inti (germinal vesicle = GV) dimana terlihat inti dan dikelilingi lapisan korona pekat, stadium metafase 1 (M-1) dimana terjadi peleburan inti sehingga tidak terlihat inti di dalam sel dan stadium metafase 11 (M-ll), yang merupakan sel telur matang dirnana terlihat adanya benda kutub I.
Secara struktural, pematangan sitoplasma oosit digambarkan oleh sejumlah perpindahan dan transformasi organel, termasuk peningkatan jumlah granula kortikal dan pergerakan sentrifugalnya (Hyttel et al., 1986a). Perubahan- perubahan sitoplasma mencerminkan adanya variasi dalam aktivitas metabolik ooplasma (Kruip et al., 1983). Pematangan oosit juga diasosiasikan dengan satu seri perubahan kuantitatif dalam pola sintesis protein dan transkripsi mKNA (Schultz dan Wassarman, 1977 dalam Xu, 1987).
kompleks (wsit-kumulus dengan massa sel kumulus yang terang) dijadikan seba- gai indikasi yang kuat untuk kematangan oosit sebab hanya oosit dengan sel-sel kumulus aktif yang dapat difertilisasi in vitro (Greve et al., 1984).
Sistem lnkubasi Pematangan Oosit I n Vitro
Salah satu media kompleks yang paling banyak digunakan untuk pema- tangan oosit in vitro adalah tissue culture medim 199
(TCM-199)
(Gordon,1 994). '1'CM- 199 terdiri atas garam-garam Earle yang mengandung N-(2-hyd- roxyethy1)-piperazine-N '-(2-ethanesulphonic acid) (HEPES) dan natrium bikarbo- nat sebagai penyangga dan disuplementasi dengan piruvat, laictat, asam amino- asam amino, vitamin, purin dan substansi lain yang terdapat di dalam serum.
Kisaran optimum tekanan osmotik medium untuk pema-tangan oosit in vitro
adalah 275 sampai 285 mosm (isotonik terhadap cairan folikel/plasma semen). Air merupakan bahan utama penyusun media pematangan oosit in vitro.
Penggunaan air ultra murni dan bebas dari kontaminan merupakan falctor yang sangat penting untuk dipersiapkan (Savister et al., 1992). Beberapa macam kelompok kontaminan yang mungkin ditemui adalah bahan-bahan padat dan gas yang terionisasi dan tidak terionisasi yang terlarut di dalam air, partikel-partikel zat, mikroba dan pirogen. Dengan menggunakan filter berdiameter 0.22 pm semua bakteri dapat dihilangkan, karena bakteri paling kecil berukuran 0.3 pm.
dan gas, serta memudahkan pemeriksaan selama penginkubasian (Gordon, 1994). Menurut Sirrad et al. (1989) pematangan oosit sapi in vi?ro memerlukan waktu 18 sampai 24 jam. Fase germinal vesicle (GV) terlihat jelas dari no1 sampai 6.6 jam, GVBD muncul dari 6.6 sampai 8.0 jam, kondensasi khromatin pada 8.0 sampai 10.3 jam, metafase I pa& 10.3 sampai 15.4 jam,
anafase
1 pa&1 5.4 sampai 16.6 jam, telofase 1 pada 16.6 sampai 18 -0 jam dan metafase 11 pada 18.0 sampai 24.0 jam semenjak awal pematangan oosit in vitro. Menurut Gordon (1994) periode pematangan oosit 24 jam menghasilkan angka pematangan dan angka cleavage yang hampir sama dengan periode pematangan oosit 18 jam, tetapi angka blastosis pada pada hari ketujuh dan kesembilan pada periode pema- tangan oosit 24 jam nyata lebih tinggi.
Jenis antibiotik yang digunakan dalam pematangan oosit sapi in vitro adalah anti bakteri yang berspektrum luas dan tidak toksik seperti penisilin (50 sampai 100 1UIml) dan streptomisin (50 sampai 100 pglrnl). Penggunaan genta- misin sulfat (50 pglml) dapat dijadikan sebagai altematif pengganti penisi- linlstreptomisin (Younis et al., 1989). Kelebihan gentamisin adalah tahan diau- toklaf pada suhu tinggi. Namun demikian Mizoshita et al. (1993) menyarankm untuk ti& menggunakan gentamisin &lam produksi embrio sapi in vitro.
Menurut Gordon (1994) suhu optimum untuk pematangan oosit in vitro adalah 38 sampai 39'~, dan suhu optimum untuk fertilisasi dan kultur embrio in vitro adalah 39%. Penggunaan suhu 4 0 ' ~ memberi pengaruh yang merusak terhadap fertilisasi oosit dan perkembangan embrio taraf awal.
dalam udara lebih tinggi dibandingkan dengan fase gas 5.0% C02, 5.0% O2 dan
90 % N2. Menurut Trounson (1992) penwunan konsentrasi 0 2 dari 20% menjadi 5.0% menghasilkan perkembangan embrio yang lebih baik. Sedan- Suzuki et al. (1997) melaporkan bahwa embrio sapi dapat dihasilkan secara in vitro meng- gunakan inkubator C02 sederhana dengan tekanan udara negatif.
Penggunaan Serum Sapi dalam Pematangan Oosit I n Vitro
Menurut Malole (1990) dan Freshney (1987) serum mengandung zat-zat nutrisi seperti bermacam-rnacam protein, asam lemak, hormon insulin, hormon steroid, mineral dan faktor penumhuh yang dapat dimanfaatkan untuk pematangan oosit dan perkembangan embrio. Secara umum, komposisi serum telah diuraikan oleh Freshney (1985) dan Malole (1990). Protein merupakan komponen serum yang terbesar, dan protein yang telah terbukti bermanfaat adalah albumin
dari
globulin. Albumin berguna untuk membawa vitamin, lemak atau lipid (asam lemak/kholesterol), hormon dan mineral; dan untuk mengatur tekanan osmotik serta sebagai penyangga. Fibronektin (globulin tak terlarut) berguna untuk merangsang perlekatan sel, sedangkan a-2 makroglobulin berfungsi meng- hambat tripsin yang merupakan enzim proteolitik. Fetuin yang terdapat dalam serum fetus meningkatkan perlekatan sel, sedangkan transferin berfungsi mengi- kat unsur besi (Fez+). Protein lain mungkin masih banyak tetapi belum jelas karakterisasinya. Menurut Malole (1990), di antara hormon yang a& &lam serum, insulin merupakan unsur yang esensial bagi pertumbuhan hampir semua sel dalam kultur.juga bovine senun albwnin (BSA). Beberapa jenis serum yang sering dicobakan dalam studi pematangan oosit sapi antara lain fetal calf serum (FCS) atau fetal bovine senun (FBS), bovine senun albumin (BSA), steer serum, serum sapi proestrus, serum sapi estrus, dan serum sapi estrus yang disuperovulasi (Gordon et al., 1994).
Leibfried-Rutledge et al. (1986) mernbuktikan bahwa FCS lebih baik dibandingkan dengan BSA sebagai suplemen protein di dalam medium. FBS mengandung fetuin, glikoprotein utama yang terdapat di
dalam
FBS, dapat mencegah pecahnya zona pelusida selama pematangan oosit in vitro (Schroeder et al., 1990). Dijelaskan bahwa fetuin dapat mencegah alctivitas enzim proteolitik yang berasal dari penglepasan granula kortikal yang terlalu cepat. Peoelitian pada tikus (Zhang et al., 1991) dan mencit (Yamazaki daa Ishibashi, 1990) juga mernbuktikan bahwa EiSS &pat mencegah pecahnya zona pelusida dan memper- baiki kapasitas fertilisasi oosit.Gordon dan Lu (1990) dan Lu et al. (1987) memodifikasi metode pema- tangan oosit in vitro pada sapi dan domba dengan cara mengeliminasi hormon- hormon dan mengganti FCS yang digunakan dalam studi-studi awal dengan 20% serum sapi estrus sebagai komponen yang memperkaya TCM-199. Angka pematangan oosit dan angka cleavage dalam penelitian ini masing-wing 79% dan 83%. Serum sapi estrus mengandung konsentrasi steroid dan gonadotropin yang sangat bervariasi dibandingkan dengan serum sapi yang diambil pada fase siklus estrus atau dari kategori sapi lainnya (Lu et al., 1987). Menwut Dobson et al. (1973) dalam Gordon (1994) konsentrasi estrogen meningkat secara tajam dari
menurut Henricks et al. (1972) konsentrasi estrogen dalam plasma darah perifera1 sapi sehari sebelum estrus berkisar antara 15 sampai 25 pglrnl
dan
level ini berta- han selama beberapa jam menjelang estrus, dan rataan komentrasi estrogen pada hari ke-3, 6, 12 dan 15 setelah hari estrus kurang dari 7.0 pgtml. Konsentrasi estrogen sedikit meningkat pa& hari kesembilan (13 pg/ml) tetapi tidak menim- bulkan perubahan berarti terhadap populasi folikel. Pada sapi bunting, konsentra- si estrogen rendah (kurang dari 5.0 pg/ml) dari hari ketiga hingga hari ke-39 sete- lah estrus. Sebaliknya konsentrasi progesteron plasma meningkat dari 1.2 nglml pada hari ketiga menjadi 9.9 ng/ml pada hari ke-12 (pada sapi bunting) dan menjadi 8.2 nglml pada hari ke- 12 @a& sapi yang tidak bunting). Setelah hari ke-12 (sel-sel lutein mulai beregresi) konsentrasi progesteron menurun pada sapi yang tidak bunting sampai satu atau dua hari sebelum estrus, dan meningkat menjadi 13.9 nglml pa& sapi yang bunting (Henricks et al., 1972).Pengaruh yang menguntungkan dari serum pada pematangan oosit in vitro munglun bereaksi melalui sel-sel kumulus atau secara langsung pada oosit. Serum juga mengandung sejumlah faktor penumbuh yang memainkan peranan penting dalam pengaturan pernatangan oosit, khususnya melalui sel-sel kumulus (Skinner, 1990 dalam Trounson, 1992). Menurut Toelihere (1985a) sel-sel kumulus berperan sebagai pemelihara
dan
pensuplai zat-zat nutrien yang dibu- tuhkan oleh oosit.memberikan pengaruh yang sama efektifnya dengan serum sapi yang sedang estrus untuk pematangan oosit in vitro pada sapi (Gordon, 1994).
Kmip et al. (1988) menyatakan bahwa persentase abnormalitas pemben- tukan (formasi) spindle dan pemunculan badan kutub meningkat secara tajam pada oosit yang dikultur 12 sampai 20 jam dalam medium yang mengandung konsentrasi estradiol yang tinggi (37 pmolll). Sila oosit dikultur selama no1 sampai enam jam dalam media yang mengandung estradiol, maka persentase abnormalitas pembentukan spindle dan pemunculan badan kutub tidak terjadi.
Menurut Lorenzo et al. (1997) penambahan
faktor
penumbuh seperti epidermal growth factor (EGP) ke dalam medium pematangan oosit umumnya berpengaruh meningkatkan produksi progesteron oleh sel-sel kumulus. Pening- katan konsentrasi EGF dari no1 nglml menjadi 1, 10 dan 50 ngtml maghasilkan peningkatan produksi progesteron dari 1.57 menjadi 3.87,
4.90 dan 10.01 nglml; meningkatkan angka pematangan oosit (M 11)dari
54.1 menjadi 61.1,
73.5 dan 72.0%; dan menghasilkan produksi estrogen yang relatif konstan dari 2.01 menjadi 2.19,
2.10 dan 2.38 nglrnl. lnteraksi antara EGF dengan sel-sel kumulus oosit menimbulkan perubahan rasio produksi estrogen dan progesteron di dalam folikel yang secara tidak langsung berpengaruh terhadap pematangan oosit in vitro. Gordon (1994) juga menyatakan ha1 yang sama bahwa pematangan oosit in vitro dipengaruhi oleh perubahan keseimbangan estrogen dan progesteron di dalam folikel. Namun para peneliti tersebut di atas menyatakan bahwa secara fisiologik, jxranan progesteron dalam proses peatangan oosit itu sendiri tidak diketahui dengan jelas.
disamping itu variasi yang sama juga mungkin timbul pada lrandungan hormon, faktor penumbuh, cytokines, vitamin dan substansi lain yang terdapat di dalam serum (Fallon et al., 1988).
Tabel 1. Konsentrasi asam amino serum sapi
Asam amino
Rataan (pmol/l)
~i sarana
Alanin
Arginin
~ s ~ a r a ~ i
nbSitrulin
Asam glutamatC
Glisin
Hist idin
Hidroksiprolin
Isoleusin
Leusin
Lisin
Metionin
Ornitin
Feni lalanin
Pro1 in
Ser
in
Treon i n
Triptofan
Tirosin
Valin
Asam amino aromat ik
Sumber : Fallon
et
al. (1988) a rataan f 2SD
termasuk
asun
aspartat. termasuk glutamin [image:200.545.45.489.156.749.2]