PENGGUNAAN
FEED ADDITIVE
TEPUNG DAUN SIRIH
(
Piper betle
L.) PADA TERNAK RUMINANSIA SEBAGAI
SALAH SATU UPAYA MITIGASI METAN
INDRI YANI MARCELINA
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Penggunaan Feed Additive Tepung Daun Sirih (Piper betle L.) pada Ternak Ruminansia sebagai Salah Satu Upaya Mitigasi Metan adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Februari 2014
ABSTRAK
INDRI YANI MARCELINA. Penggunaan Feed Additive Tepung Daun Sirih (Piper betle L.) pada Ternak Ruminansia sebagai Salah Satu Upaya Mitigasi Metan. Dibimbing oleh ASEP SUDARMAN dan ANURAGA JAYANEGARA.
Penelitian In vitro selama 48 jam inkubasi ini bertujuan untuk mengevaluasi penggunaan tepung daun sirih (Piper betle L.) dalam ransum terhadap produksi gas metan dalam rumen ternak ruminansia. Produksi metan di dalam rumen diestimasi dengan menggunakan persamaan stoikiometri dari proporsi asam lemak terbang yaitu asam asetat (C2), asam propionat (C3) dan
asam butirat (C4). Rancangan yang digunakan adalah RAK 3 ulangan dan 4
perlakuan dengan level penambahan tepung daun sirih 0%, 1%, 2%, dan 3%. Perbedaan perlakuan diuji dengan menggunakan uji polynomial. Hasil penelitian menunjukkan produksi gas metan dan total gas pada ransum dengan penambahan 2% tepung daun sirih lebih rendah dibanding dengan perlakuan yang lain. Tepung daun sirih cenderung menurunkan jumlah protozoa (P<0.1) dan meningkatkan proporsi asam propionat (C3) dan asam butirat (C4) secara nyata (P<0.05). Dosis
penggunaan tepung daun sirih yang tepat untuk diaplikasikan ke ternak adalah 2% dalam ransum.
Kata kunci: gas metan (CH4), produksi gas, tepung daun sirih, VFA
ABSTRACT
INDRI YANI MARCELINA. The Use of Betel Leaf Meal (Piper betle L.) on Ruminants for Mitigating Methane Emission. Supervised by ASEP SUDARMAN and ANURAGA JAYANEGARA.
This research was conducted using In vitro technique for 48 hours of incubation time. The aim was to evaluate supplementation of Betel leaf meal to decrease methane production. Methane produced in the rumen was estimated by using data of volatile fatty acids (VFA) partial concentration, particularly three main acids, i.e. acetic acid, propionic acid and butyric acid. The experiment used block randomized design with three replicates and four treatments. The treatments were 0%, 1%, 2%, and 3% of betel leaf meal in feed. The results showed that suplementation of Betel leaf meal decreased methane and total gas production. Betel leaf meal also tended to decrease protozoa population (P<0.1), and increased propionic acid (C3) proportion and butiric acid (C4) proportion (P<0.05).
The best dose of Betel leaf meal to reduce methane and to maintain optimum rumen condition was 2%.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Peternakan
pada
Departemen Ilmu Nutrisi dan Teknologi Pakan
PENGGUNAAN
FEED ADDITIVE
TEPUNG DAUN SIRIH
(
Piper betle
L.) PADA TERNAK RUMINANSIA SEBAGAI
SALAH SATU UPAYA MITIGASI METAN
INDRI YANI MARCELINA
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNKAN
INSTITUT PERTANIAN BOGOR BOGOR
Judul Skripsi : Penggunaan Feed Additive Tepung Daun Sirih (Piper betle L.) pada Ternak Ruminansia sebagai Salah Satu Upaya Mitigasi Metan Nama : Indri Yani Marcelina
NIM : D24090057
Disetujui oleh
Dr Ir Asep Sudarman, MRurSc Pembimbing I
Dr Anuraga Jayanegara, SPt,MSc Pembimbing II
Diketahui oleh
Prof Dr Ir Panca Dewi MHK, MSi Ketua Departemen
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Juli 2013 ini ialah mitigasi metan, dengan judul Penggunaan Feed Additive Tepung Daun Sirih (Piper betle L.) pada Ternak Ruminansia sebagai Salah Satu Upaya Mitigasi Metan.
Tepung daun sirih yang sudah terbukti khasiatnya sebagai salah satu herbal penyembuh mastitis subklinis pada ternak perah diharapkan dapat menjadi hijauan sumber feed additive yang dapat memenuhi kebutuhan nutrisi ternak serta mampu meminimalisir produksi gas metan pada saluran pencernaan ternak ruminansia. Oleh karena itu dilakukan pengujian secara In vitro untuk mengukur produksi gas metan dan total gas yang dihasilkan ternak dengan penambahan tepung daun sirih dalam ransum.
Skripsi ini merupakan salah satu syarat untuk kelulusan dan memperoleh gelar Sarjana Peternakan di Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor. Semoga karya ilmiah ini bermanfaat dan menambah amal kebaikan penulis dan pembimbing di akhirat kelak. Amin.
Bogor, Februari 2014
DAFTAR ISI
DAFTAR TABEL x
DAFTAR GAMBAR x
DAFTAR LAMPIRAN x
PENDAHULUAN 1
METODE 2
Bahan 2
Alat 2
Lokasi dan Waktu Penelitian 2
Prosedur Penelitian 2
Rancangan Percobaan 6
HASIL DAN PEMBAHASAN 7
Nilai pH Rumen 7
Produksi Volatile Fatty Acid (VFA) Total 7
Proporsi Volatile Fatty Acid (VFA) Parsial 8
Populasi Protozoa 12
Produksi Gas Metan 13
Kinetika Produksi Gas 15
Estimasi DBO dan Nilai EM 17
SIMPULAN DAN SARAN 18
Simpulan 18
Saran 18
DAFTAR PUSTAKA 18
LAMPIRAN 21
DAFTAR TABEL
1 Komposisi nutrient ransum perlakuan 5
2 Nilai pH rumen 7
3 Proporsi VFA parsial 9
4 Populasi protozoa rumen 12
5 Produksi gas, fraksi yang mudah dicerna, fraksi potensial didegradasi, nilai potensial degradasi dan laju produksi gas per jam 16
6 Nilai DBO dan EM 17
DAFTAR GAMBAR
1 Nilai produksi VFA total masing-masing perlakuan 8 2 Kurva regresi linear dan kuadratik propionat (C3) 10
3 Kurva regresi linear kuadratik rasio C2:C3 10
4 Kurva regresi kuadratik proporsi butirat (C4) 11
5 Estimasi produksi gas metan (CH4) 14
6 Perkembangan produksi gas selama 48 jam inkubasi 15
DAFTAR LAMPIRAN
1 Analisis ragam nilai pH rumen 21
2 Analisis ragam VFA total 21
3 Analisis ragam proporsi asam asetat (C2) 21
4 Analisis ragam proporsi asam propionat (C3) 22
5 Uji polinomial ortogonal proporsi asam propionat (C3) 22
6 Analisis ragam proporsi asam butirat (C4) 22
7 Uji polinomial ortogonal proporsi asam butirat (C4) 22
8 Analisis ragam proporsi isobutirat (IC4) 23
9 Analisis ragam rasio C2:C3 23
10 Uji polinomial ortogonal rasio C2:C3 23
11 Analisis ragam proporsi valerat (C5) 23
12 Analisis ragam proporsi isovalerat (IC5) 24
13 Analisis ragam populasi protozoa 24
14 Uji polinomial ortogonal populasi protozoa 24
15 Analisis ragam produksi gas metan (CH4) 24
16 Analisis ragam produksi gas total 25
17 Analisis ragam koefien (a) 25
18 Analisis ragam koefisien (b) 25
19 Analisis ragam koefisien (a+b) 26
20 Analisis ragam koefisien (c) 26
PENDAHULUAN
Metan (CH4) adalah salah satu gas yang dihasilkan oleh proses pencernaan
fermentatif ternak ruminansia. Metan (CH4) adalah salah satu golongan gas rumah
kaca yang dapat menyebabkan efek gas rumah kaca. Gas rumah kaca di atmosfer memiliki fungsi untuk menahan panas sehingga suhu di bumi ini dapat ideal untuk kelangsungan makhluk hidup. Akan tetapi permasalahan yang muncul adalah ketika konsentrasi gas rumah kaca di bumi ini berlebih sehingga menyebabkan pemanasan global.
Sekitar 50% emisi gas metan merupakan hasil aktivitas manusia yang berasal dari pertanian dimana 27% berasal dari sektor peternakan. Untuk Indonesia, perkiraan emisi gas metan dari ternak ruminansia adalah 98% dari total 849 Gg per tahun, sedangkan 2% berasal dari babi dan kuda (Haryanto dan Thalib 2009). Gas metan pada ternak ruminansia berasal dari dua sumber yaitu berasal dari menejemen kotoran (manure) dan dari hasil fermentasi saluran pencernaan (enteric fermentation) yang akan dikeluarkan melalui proses eruktasi 83%, pernafasan 16%, dan anus 1% (McDonald et al. 2002). Fermentasi dari saluran pencernaan ternak menyumbang sebagian besar emisi gas metan yang dihasilkan oleh sektor peternakan. Sebesar 80%-95% metan diproduksi di dalam rumen dan 5%-20% di dalam usus besar (Martin et al. 2008). Produksi gas metan dalam saluran pencernaan ternak ruminansia disebabkan oleh adanya Archaea metanogen yang bersimbiosis dengan protozoa rumen.
Selain mencemari lingkungan, produksi gas metan dari pencernaan ternak juga merugikan ternak, serta peternak dapat mengalami kerugian biaya pemeliharaan dari pakan. Hal tersebut disebabkan karena pembentukan metan merupakan proses pemborosan yang dapat mengurangi 6%-10% gross energy (Jayanegara dan Sofyan 2008) yang seharusnya dapat dikonversi dalam bentuk produk fermentasi untuk mengoptimalkan produktivitas ternak. Mitigasi metan adalah tindakan untuk meminimalisir produksi gas metan. Berbagai upaya telah dilakukan dalam mengurangi emisi metan ruminansia dan strategi yang paling berhasil untuk mereduksi emisi metan salah satunya dengan memodifikasi ekosistem mikroba, tetapi manipulasi komponen mikroba dari sistem tersebut harus terintegrasi (Morgavi et al. 2010) seperti penggunaan tanaman yang mengandung saponin dan tanin sebagai pendefaunasi protozoa di dalam rumen.
Salah satu tanaman yang mengandung tanin dan saponin adalah sirih hijau (Piper betle L.). Menurut hasil uji fitokimia daun sirih memiliki kandungan minyak atsiri, tanin, fenol, saponin, flavonoid, dan asam amino yang cukup lengkap. Kandungan tanin pada daun sirih muda dan daun tua adalah sama (Darwis 1991). Penggunaan tepung daun sirih sebagai salah satu feed additive alami untuk mengurangi jumlah metan dalam saluran pencernaan belum pernah diteliti sebelumnya. Penelitian penambahan tepung daun sirih dalam ransum diharapkan dapat menurunkan produksi metan pada sistem pencernaan ternak ruminansia sehingga dapat meminimalisir kerusakan lingkungan sekaligus mampu meningkatakan produktivitas ternak perah.
2
METODE
Bahan
Bahan yang digunakan dalam penelitian ini adalah rumen segar, larutan McDougall, larutan buffer gas tes, H2SO4 pekat, HCl 0.5 N, larutan H2SO4 15%,
larutan NaOH 0.5 N, larutan indikator phenolphthalein (PP), larutan tryphan blue formaline saline (TBFS), gas CO2, konsentrat komersil sapi perah, rumput gajah,
dan tepung daun sirih (Piper betle L.).
Alat
Alat yang digunakan dalam penelitian adalah alat-alat percobaan In vitro seperti timbangan digital, tabung fermentor bersumbat karet, shaker waterbath, syringe gas test, tabung gas CO2, pH meter, sentrifuse, erlenmeyer, tabung
microtube, counting chamber Fuchs-Rosenthal, mikroskop cahaya, termos, spektrofotometer, dan set alat gas cromatograph.
Lokasi dan Waktu Penelitian
Penelitian ini dilakukan secara In vitro pada bulan Juli 2013 hingga September 2013di Laboratorium Nutrisi Ternak Perah Fakultas Peternakan IPB, Analisa volatile fatty acid (VFA) parsial dan kandungan saponin tepung daun sirih dilakukan di Laboratorium Fisiologi Ternak, Balai Penelitian Ternak, Ciawi Bogor. Pengujian kandungan tanin tepung daun sirih dilakukan di Laboratorium Kimia Balai Besar Pasca Panen, Taman Cimanggu Bogor. Analisa proksimat dilakukan di Laboratorium ITP INTP Fakultas Peternakan IPB.
Prosedur Penelitian
Pengambilan Cairan Rumen
Cairan rumen diambil pada pagi hari dari sapi Friesian holstein berfistula sebelum diberi pakan. Penutup fistula dibersihkan dengan air hangat dan iodin kemudian dibuka dan segera diambil cairan rumen. Setelah koleksi rumen selesai tutup fistula segera dipasangkan kembali serapat mungkin lalu dibersihkan dengan air dan iodin. Cairan rumen yang ditempatkan di dalam termos bersuhu 39 °C dibawa ke laboratorium, disaring menggunakan kain nilon dan dialiri CO2.
Pembuatan Larutan McDougall sebagai Saliva Buatan
Sebanyak 800 ml aquades dimasukan ke dalam tabung erlenmeyer ukuran 2 liter dengan ditambahkan bahan-bahan berikut: 9.8 g NaHCO3, 4.649 g
Na2HPO4.2H2O, 0.57 g KCl, 0.47 g NaCl, 0.12 g MgSO4.7H2O, bahan tersebut
dihomogenkan dengan magnetic stirrer, setelah itu ditambahkan 0.04 g CaCl2 dan
3 Inkubasi In vitro
Sampel pakan konsentrat dan hijauan dikeringkan dalam oven 60 °C, digiling dan disaring menggunakan alat penyaring. Sampel diinkubasi In vitro berdasarkan metode Tilley dan Terry (1963). Sebanyak 500 mg sampel diinkubasi ke dalam tabung fermentor, kemudian ditambahkan dengan 10 ml cairan rumen dan 40 ml larutan McDougall. Lalu tabung fermentor dimasukan ke dalam shaker waterbath dengan suhu 39 °C, tabung dikocok dengan dialiri CO2 selama 30 detik,
kemudian ditutup dengan karet berventilasi. Setelah 4 jam, tutup karet fermentor dibuka lalu rumen inkubasi dicek pH dengan pH meter hingga nilai yang tertera di layar pH meter konstan. Setelah itu diambil sebanyak 2 ml cairan inkubasi dan dimasukan ke dalam tabung microtube lalu ditambah H2SO4 untuk kemudian
dianalisa volatile fatty acid (VFA) parsial. Untuk analisa protozoa diambil sebanyak 1 ml cairan rumen inkubasi dan dicampur ke dalam 1 ml larutan tryphan blue formaline saline (TBFS). Sisa inkubasi kemudian ditetesi dengan HgCl2 dan
disentrifuse pada 4000 rpm selama 15 menit dan disimpan di freezer. Analisa Volatile Fatty Acid (VFA) Total
Konsentrasi VFA total diukur dengan menggunakan teknik destilasi uap (General Laboratory Procedure, Department of Dairy Science University of Wisconsin 1966). Sebanyak 5 ml supernatan ditempatkan ke dalam corong destilasi dan ditambah 1 ml H2SO4 15% sambil dimasukan perlahan ke dalam
tabung destilasi dengan dibilas menggunakan aquades kemudian corong ditutup. Uap air ditampung di ujung tabung destilasi dengan erlenmeyer yang berisi NaOH 0.5 N sebanyak 5 ml. Setelah itu hasil destilasi ditetesi indikator PP dan dititrasi dengan HCl 0.5 N. Konsentrasi VFA total dihitung dengan rumus:
Keterangan : a = volume titran blanko b = volume titran sampel Analisa Volatile Fatty Acid (VFA) Parsial
Analisa menggunakan alat set gas cromatograph Bruker® Scion 436-GC, dan SHO-40 dengan sistem auto inject. Sampel rumen yang telah diasamkan ditambah dengan 0.003 gram asam sulfo 5 salisilat dehidrat. Kemudian campuran dalam tabung microtube tersebut disentrifus selama 10 menit pada 12000 rpm dan suhu 7 °C. Selanjutnya supernatan dimasukan ke dalam tabung berpenutup dan ditempatkan ke dalam rak Bruker® SHO-40. Pada saat analisa berlangsung akan muncul zona peak, peak tersebut kemudian dibaca pada menit ke 6.93 untuk asetat, 7.75 untuk propionat, 8.08 untuk isobutirat, 8.82 untuk butirat, 9.46 untuk isovalerat, dan menit ke 10.64 untuk valerat. Hasil kromatogram dan nilai VFA parsial muncul di layar monitor lalu kemudian keluar dalam bentuk print out. Populasi Protozoa Rumen
4
protozoa dilakukan dengan mikroskop cahaya pada pembesaran 100 kali. Populasi protozoa dihitung dengan rumus:
Keterangan: n = jumlah protozoa yang terhitung dalam counting chamber FP = faktor pengencer
Produksi Gas Metan
Produksi gas metan dapat dihitung dengan pendekatan stoikiometri proporsi VFA parsial. Menurut Moss et al. (2000) produksi metan dapat dihitung melalui persamaan:
Pembuatan larutan buffer diperlukan bahan 0.1 ml larutan mineral mikro (13.2 g CaCl2.2H2O+ 10 g MnCl2.4H2O+ 1 g CoCl2.6H2O + 8 g FeCl3.6H2O + dicampur dengan larutan buffer hingga homogen. Campuran tersebut kemudian disimpan di dalam waterbath dan dialiri CO2. Sebanyak 30 ml campuran media
cairan rumen dimasukan ke masing-masing syringe, udara yang ada dalam syringe dikeluarkan dan klep syringe ditutup. Posisi piston dibaca pada waktu sebelum inkubasi (Gb0), lalu diinkubasi dalam waterbath bersuhu 39 °C selama 48 jam.
Posisi piston dibaca dalam jarak dua jam selama 12 jam pertama, kemudian pada jarak 4 jam pada 12 jam kedua, dan jarak 12 jam hingga 48 jam (Gb48).
5 Kinetika Produksi Gas
Kinetika produksi gas diestimasi melalui persamaan eksponensial yang dideskripsikan oleh Ørskov dan McDonald (1979) sebagai berikut:
p = a + b (1 - e-ct)
Nilai p adalah produksi gas kumulatif pada waktu t jam, sedangkan a, b, dan c merupakan konstanta dari persamaan eksponensial tersebut. Konstanta dapat diintrepretasikan sebagai produksi gas dari fraksi yang mudah larut (a), produksi gas dari fraksi yang tidak larut namun dapat difermentasikan (b) dan laju reaksi pembentukan gas (c), dengan demikian (a + b) dapat diartikan sebagai produksi gas maksimum yang dapat terbentuk selama proses fermentasi pada waktu t mendekati tak hingga. Perhitungan konstanta persamaan eksponensial dilakukan dengan program SPSS.
Estimasi Degradasi Bahan Organik (DBO) dan Energi Metabolis (EM)
Estimasi DBO dan EM dapat dihitung berdasarkan produksi gas 24 jam dan kandungan nutrien pakan mengikuti formula sebagai berikut (Menke et al. 1979):
DBO (%) = 14.88 + 0.889PG + 0.045PK + 0.065XA Keterangan:
DBO = degradasi bahan organik (%) PG = produksi gas (ml) Komposisi nutrien ransum ditampilkan pada Tabel 1.
Tabel 1 Komposisi nutrien ransum perlakuan (% BK)
Kandungan nutrien* Level penambahan tepung daun sirih
6
Uji Kuantitatif Saponin (AOAC 2003)
Proses preparasi sebanyak 0.5 g sampel diekstrak dengan 10 ml MeOH dengan diultrasonik selama 30 menit lalu difilter dan diambil filtratnya untuk dianalisa. Selanjutnya pada proses pengerjaan sebanyak 0.25 ml sampel disimpan dalam waterbath es, lalu ditambah 0.25 ml larutan vanilin dan 2.5 ml H2SO4 72%
lalu dipanaskan pada suhu 60 °C selama 10 menit. Setelah itu didinginkan hingga suhu ruang dan diukur absorbansinya menggunakan spektrofotometer dengan panjang gelombang 544 nm.
Uji Kuantitatif Tanin (AOAC 2003)
Sebanyak 2 g sampel dihaluskan, kemudian dimasukan ke dalam labu didih lalu ditambahkan 350 ml air suling. Larutan kemudian direfluks selama 3 jam, lalu didinginkan. Larutan kemudian disaring dan dipindahkan secara kuantitatif ke dalam labu takar 500 ml dan diimpit dengan air suling hingga tanda tera dan dihomogenkan. Sebanyak 2 ml filtrat dipipet ke dalam labu takar 100 ml dan ditambahkan 2 ml pereaksi Folin Denis, serta 5 ml Na2CO3 jenuh. Larutan filtrat
diimpit dengan air suling lalu dihomogenkan dan didiamkan selama 40 menit, kemudian diukur absorbansinya menggunakan spektrofotometer dengan panjang gelombang 725 nm.
Rancangan Percobaan
Rancangan percobaan yang digunakan dalam penelitian ini adalah Rancangan Acak Kelompok (RAK) yang terdiri dari empat perlakuan dan tiga ulangan. Perlakuan tersebut antara lain:
P0 = ransum basal (konsentrat:hijauan = 1:1) sebagai kontrol P1 = ransum basal dengan penambahan tepung daun sirih 1% P2 = ransum basal dengan penambahan tepung daun sirih 2% P3 = ransum basal dengan penambahan tepung daun sirih 3% Model matematik yang digunakan adalah sebagai berikut:
Yij = µ +τi +βj + εij
Keterangan:
Yij = nilai pengamatan efek perlakuan ke-i, ulangan ke-j
µ = nilai rataan umum τi = pengaruh perlakuan ke-i βj = pengaruh ulangan ke-j
εij = error perlakuan ke-i dan ulangan ke-j
7
HASIL DAN PEMBAHASAN
Nilai pH Rumen
Fermentasi pakan di dalam rumen akan mempengaruhi nilai pH rumen. Nilai pH akan semakin turun ketika aktivitas fermentasi semakin tinggi. Nilai pH rumen adalah faktor yang berpengaruh besar terhadap kondisi rumen, baik untuk pertumbuhan mikroba rumen maupun dalam menghasilkan produk fermentasi berupa VFA dan N-NH3 (McDonald et al. 2002). Berdasarkan hasil pengukuran
pH pada penelitian ini menunjukkan bahwa pemberian tepung daun sirih berlevel dalam ransum tidak berpengaruh nyata (P>0.05) pada nilai pH rumen. Data nilai pH penelitian dapat dilihat pada Tabel 2.
Tabel 2 Nilai pH rumen
*(P=0.271) Analisis Laboratorium Nutrisi Ternak Perah, INTP IPB.
Setelah difermentasi selama 4 jam, nilai pH yang didapat mendekati pH netral yaitu 6.707-6.773 dan masih berada dikisaran pH ideal rumen. Hasil pengukuran pH tersebut menunjukkan hasil yang sama dengan penelitian Yamin et al. (2013) bahwa penambahan tepung daun sirih hingga level 8% menunjukkan pH yang tetap yaitu 6.7. Rataan pH rumen yang normal berada pada kisaran 6-7 (France dan Siddon 1993). Kebanyakan pertumbuhan archaea metanogenik berada pada pH 6.7-7.4, tetapi optimalnya pada kisaran pH 7.0-7.2 dan proses dapat gagal jika pH mendekati 6.0 (Karakashev 2005). Nilai pH yang diperoleh dari masing-masing perlakuan termasuk dalam kategori yang baik untuk berlangsungnya metabolisme di dalam rumen sehingga penambahan tepung daun sirih hingga level 3% aman diberikan kepada ternak ruminansia.
Produksi Volatile Fatty Acid (VFA) Total
Proses pencernaan karbohidrat di dalam rumen ternak ruminansia akan menghasilkan energi berupa VFA. Degradasi karbohidrat di dalam rumen dilakukan dengan dua tahapan yaitu 1) karbohidrat kompleks (polisakarida: selulosa, pati dan lain-lain) dihidrolisa menjadi gula sederhana (monosakarida glukosa) oleh enzim-enzim mikroba rumen, 2) monosakarida menjadi piruvat yang selanjutnya akan diubah menjadi produk akhir yaitu VFA (asetat, propionat dan butirat) (McDonald et al. 2002). VFA sangat penting karena sebagai sumber energi yang memenuhi sekitar 50%-70% dari kebutuhan energi ternak ruminansia
Level penambahan tepung daun sirih Nilai pH*
0% 6.737±0.06
1% 6.773±0.04
2% 6.743±0.04
8
(Damron 2006). Hasil pengukuran VFA total dengan penambahan tepung daun sirih dengan level yang berbeda dalam ransum dapat dilihat pada Gambar 1.
Pada Gambar 1 menunjukkan penambahan tepung daun sirih tidak berpengaruh nyata (P>0.05) terhadap produksi VFA total pada masing-masing perlakuan. Konsentrasi VFA total yang layak bagi kelangsungan hidup ternak yaitu 70-150 mM (McDonald et al. 2002). Nilai produksi VFA total pada penelitian adalah 129.005-153.568 mM. Sehingga ransum dengan penambahan tepung daun sirih dapat mencukupi kebutuhan energi untuk ternak.
Gambar 1 Nilai produksi VFA total masing-masing perlakuan *(P=0.184) Analisis Laboratorium Nutrisi Ternak Perah INTP-IPB.
Produksi VFA mengalami peningkatan pada penambahan tepung daun sirih dan VFA tertinggi yaitu pada penambahan 2%, akan tetapi produksi VFA mengalami penurunan pada level 3%. Hal tersebut diduga karena kandungan zat aktif daun sirih yang lebih banyak, salah satu zat aktif tersebut adalah eugenol. Eugenol dapat menurunkan serta mengubah profil VFA karena eugenol dapat membunuh bakteri pencerna serat. Hasil penelitian Castillejos et al. (2006) penggunaan eugenol dengan dosis 500 mg l-1 dapat menurunkan VFA total dan proporsi asetat. Selain itu daun sirih mengandung flavonoid, salah satunya adalah fenol. Fenol daun sirih ini memiliki kemampuan membunuh bakteri 5 kali lipat dari fenol biasa (Moeljanto 2003). Menurut Yamin et al. (2013) penambahan level tepung daun sirih berpengaruh nyata terhadap produksi VFA total. Kadar VFA yang tertinggi yaitu pada perlakuan 2%, tetapi kadar VFA menurun pada level 4%, 6% dan 8%.
Proporsi Volatile Fatty Acid (VFA) Parsial
Konsentrasi VFA total umumnya memberikan nilai yang kurang akurat dibandingkan dengan proporsi VFA parsialnya (Church 1976). VFA terdiri dari asetat, propionat, butirat, valerat, kaproat, isobutirat, isovalerat, 2-metilbutirat dalam jumlah sedikit, serta beberapa jenis asam lainnya yang diproduksi di rumen
9 sebagai hasil akhir fermentasi mikroba rumen (France dan Dijkstra 2005). Secara umum asam asetat merupakan komponen terbesar yaitu sekitar 65%, asam propionat 21%, asam butirat 14% (McDonald et al. 2002), isobutirat 1%, isovalerat dan valerat di bawah 3% (Hungate 1966). Sapi perah yang diberi pakan hijauan dan konsentrat 1:1 menghasilkan VFA total sebesar 109 mmol l-1 dengan proporsi asetat 67%, propionat 21% dan butirat 12% (France dan Dijkstra 2005). Hasil pengukuran proporsi VFA parsial pada penelitian ini ditampilkan pada Tabel 3.
Tabel 3 Proporsi VFA parsial
Proporsi VFA
parsial (%)*
Level penambahan tepung daun sirih P
Value Kuadratik
**
0% 1% 2% 3%
Asetat (C2) 66.044±3.483 58.026±0.141 53.910±9.062 58.625±5.548 0.238 NS
Propionat (C3) 19.016±1.126a 24.202±0.741b 22.332±1.559b 22.281±0.539b 0.011 0.010
Butirat (C4) 9.076±0.645a 13.133±0.585b 13.749±2.198b 11.129±1.135ab 0.029 0.008
Isobutirat(IC4) 1.379±0.517 1.244±0.015 2.237±1.404 2.103±1.547 0.671 NS
Valerat (C5) 0.822±0.137 1.634±0.118 4.488±3.547 3.572±1.980 0.247 NS
Isovalerat(IC5) 1.020±0.217 1.761±0.021 3.283±2.279 2.290±0.799 0.276 NS
Rasio C2/C3 3.475±0.098b 2.399±0.079b 2.419±0.428a 2.636±0.308a 0.014 0.010
*Analisis Laboratorium Fisiologi Ternak, BPT Ciawi-Bogor (2013); Angka-angka pada baris yang sama yang
diikuti oleh huruf berbeda menunjukkan beda nyata (P<0.05); ** P Value pengaruh perlakuan secara kuadratik,
NS menunjukkan P>0.05.
Pada Tabel 3 penambahan tepung daun sirih menunjukkan pengaruh nyata (P<0.05) terhadap proporsi propionat, butirat dan rasio C2:C3, tetapi tidak
menunjukkan pengaruh nyata (P>0.05) terhadap proporsi asetat, isobutirat, valerat, dan isovalerat. Proporsi asetat yang dihasilkan pada penelitian ini adalah 53.910%-66.044%. Menurunnya produksi asetat dengan penambahan tepung daun sirih diharapkan dapat menurunkan produksi gas metan. Menurut Jayanegara dan Sofyan (2008) produksi asetat diikuti dengan produksi H2 yang digunakan sebagai
substrat pada reaksi metanogenesis. Proporsi yang paling tinggi dihasilkan dalam penelitian ini adalah asetat. Asetat banyak dihasilkan atau merupakan produk utama fermentasi bakteri selulolitik (Hobson dan Jouany 1988).
10
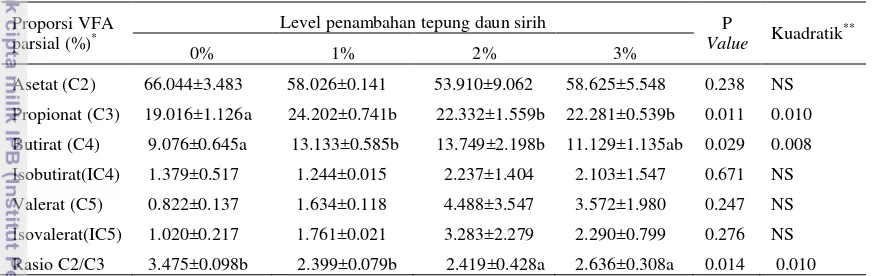
Gambar 2 Kurva proporsi propionat (C3)
Di dalam rumen heksosa dihidrolisis oleh bakteri amilolitik dan sakarolitik, sehingga produksi asam laktat meningkat seiring dengan meningkatnya aktivitas Streptococcus. Peningkatan asam laktat menyebabkan laju proses di jalur akrilat mengalami peningkatan, sehingga menghasilkan propionat lebih tinggi (Arora 1989). Penurunan proporsi propionat dengan level pemberian lebih dari 2% diduga karena kandungan kavicol daun sirih yang lebih tinggi sehingga menurunkan aktifitas Streptococcus untuk menghasilkan laktat, sehingga laju di jalur akrilat menurun yang menyebabkan produksi propionat menurun. Kavicol mampu menghambat pertumbuhan bakteri Streptococcus. Menurut penelitian Nalina dan Rahim (2007) ekstrak daun sirih mampu menghambat pertumbuhan bakteri Streptococcus mutans.
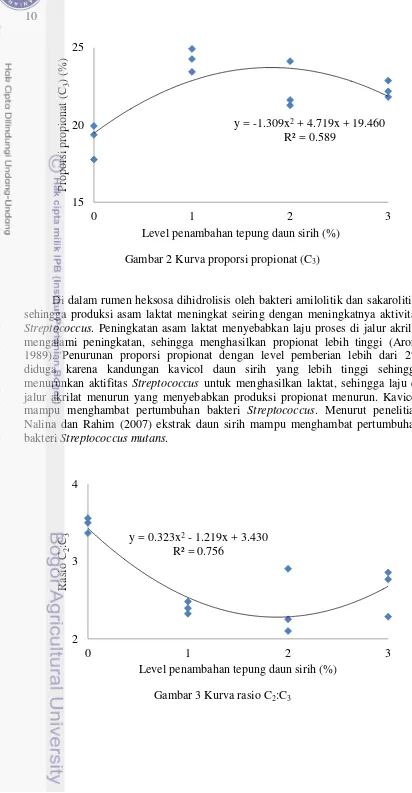
Gambar 3 Kurva rasio C2:C3
y = -1.309x2 + 4.719x + 19.460
R² = 0.589
15 20 25
0 1 2 3
P
ropor
si propionat (
C3
) (
%
)
Level penambahan tepung daun sirih (%)
y = 0.323x2 - 1.219x + 3.430
R² = 0.756
2 3 4
0 1 2 3
R
asio C
2
:C
3
11 Peningkatan proporsi propionat dapat disebabkan oleh penggunaan H2 hasil
fermentasi untuk produksi propionat akibat rendahnya aktifitas metanogen dalam pembentukan gas metan (CH4). Dengan meningkatnya proporsi propionat
diharapkan produksi metan yang dihasilkan dalam saluran pencernaan ruminansia berkurang, selain itu meningkatnya proporsi propionat diharapkan mampu meningkatkan produksi susu.
Proporsi propionat yang meningkat menyebabkan rasio asetat:propionat menurun secara nyata (P<0.05). Gambar 3 menunjukkan penambahan tepung daun sirih nyata menurunkan rasio C2:C3. Rasio C2:C3 dalam rumen dapat
memberikan indikasi tentang pemanfaatan hasil fermentasi tersebut lebih ke arah produksi susu atau untuk perbaikan kualitas susu. Selain itu rasio C2:C3 juga dapat
digunakan untuk menentukan produksi metan. Jika Rasio C2:C3 rendah maka
metan yang dihasilkan dalam rumen rendah.
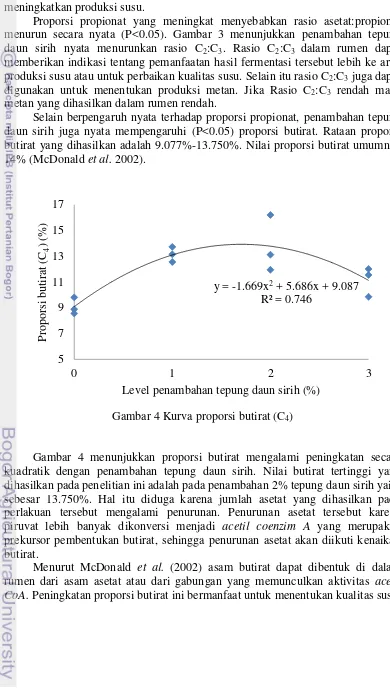
Selain berpengaruh nyata terhadap proporsi propionat, penambahan tepung daun sirih juga nyata mempengaruhi (P<0.05) proporsi butirat. Rataan proporsi butirat yang dihasilkan adalah 9.077%-13.750%. Nilai proporsi butirat umumnya 14% (McDonald et al. 2002).
Gambar 4 Kurva proporsi butirat (C4)
Gambar 4 menunjukkan proporsi butirat mengalami peningkatan secara kuadratik dengan penambahan tepung daun sirih. Nilai butirat tertinggi yang dihasilkan pada penelitian ini adalah pada penambahan 2% tepung daun sirih yaitu sebesar 13.750%. Hal itu diduga karena jumlah asetat yang dihasilkan pada perlakuan tersebut mengalami penurunan. Penurunan asetat tersebut karena piruvat lebih banyak dikonversi menjadi acetil coenzim A yang merupakan prekursor pembentukan butirat, sehingga penurunan asetat akan diikuti kenaikan butirat.
Menurut McDonald et al. (2002) asam butirat dapat dibentuk di dalam rumen dari asam asetat atau dari gabungan yang memunculkan aktivitas acetil CoA. Peningkatan proporsi butirat ini bermanfaat untuk menentukan kualitas susu,
y = -1.669x2 + 5.686x + 9.087
12
asam butirat merupakan salah satu prekursor pembentuk lemak susu meskipun hanya menyumbang 10%.
Selain menghasilkan asetat, propionat dan butirat, pencernaan rumen juga menghasilkan isobutirat (IC4), valerat (C5) dan isovalerat (IC5) meskipun dalam
jumlah proporsi yang kecil. Penambahan tepung daun sirih hingga level 3% tidak mempengaruhi proporsi VFA tersebut (P<0.05). Namun terdapat pola yang sama antara valerat (C5) dengan isovalerat (IC5) yang meningkat hingga level 2% dan
proporsinya menurun pada level 3%. Meningkatnya proporsi valerat dan isovalerat ini bedampak positif, karena dalam proses pembentukannya menggunakan H2 yang dihasilkan dalam pembentukan asetat dan butirat, sehingga
H2 yang berikatan dengan CO2 untuk membentuk metan semakin sedikit. Hal itu
akan mengakibatkan produksi metan dalam rumen menurun.
Populasi Protozoa
Protozoa merupakan salah satu mikroba yang hidup secara anaerob di dalam rumen dan ikut mempengaruhi fermentasi rumen. Keberadaan protozoa dalam rumen sering menggangu ekosistem bakteri karena memiliki sifat memangsa bakteri. Selain itu keberadaan protozoa mempengaruhi jumlah metanogen di dalam rumen, karena metanogen hidup bersimbiosis dengan protozoa. Sebanyak 20% (Bryden dan Annison 1998) sampai 37% (Newbold et al. 1995) metanogen bersimbiosis dengan protozoa, dan sisanya beraktivitas bebas di dalam rumen. Keberadaan protozoa di dalam rumen cukup penting, tetapi tidak mutlak dan cenderung dapat merugikan sehingga perlu adanya proses defaunasi. Hasil perhitungan populasi protozoa pada penelitian ini disajikan pada Tabel 4.
Tabel 4 Populasi protozoa rumen
Level penambahan tepung daun sirih Populasi protozoa (log sel ml-1)*
0% 3.966±0.088b
1% 3.904±0.085ab
2% 3.828±0.113ab
3% 3.784±0.190a
*(P=0.076) Analisis Laboratorium BFMN INTP IPB (2013).
13 yang kompleks dengan sterol pada permukaan membran sel protozoa, sehingga menyebabkan membran sel protozoa pecah, lalu sel mengalami lisis dan akhirnya mengalami kematian (Wallace et al. 2002). Tanin memiliki potensi menurunkan jumlah protozoa dan produksi metan dengan tidak mempengaruhi kondisi normal rumen. Menurut hasil penelitian Daning (2010) penambahan limbah teh hitam (Bohea bulu) yang setara dengan kandungan tanin 3, 6, dan 12 mg g-1 substrat pada fermentasi rumput gajah dan dedak halus secara In vitro nyata menurunkan populasi protozoa, produksi gas dan metan.
Menurunnya protozoa diharapkan memberikan kesempatan hidup yang lebih baik untuk mikroba rumen sehingga mampu meningkatkan jumlah bakteri amilolitik yang dapat menghasilkan lebih banyak asam propionat. Hal ini sejalan dengan proporsi propionat yang meningkat dengan penambahan tepung daun sirih hingga level 2%. Hasil penelitian Yamin et al. (2013) menunjukkan jumlah bakteri total rumen meningkat pada taraf pemberian tepung daun sirih sebesar 2% dan mengalami penurunan seiring dengan bertambahnya level penggunaan tepung daun sirih. Selain itu menurunnya jumlah protozoa diharapkan mampu menurunkan jumlah metanogen sehingga gas metan yang dihasilkan akan menurun. Menurut Jouany (1991) terdapat hubungan antara populasi protozoa dan produksi gas metan dalam rumen. Semakin tinggi populasi protozoa, maka gas metan yang dihasilkan semakin banyak.
Produksi Gas Metan
Pada ternak ruminansia senyawa-senyawa organik bahan pakan difermentasi oleh mikroba rumen sehingga menghasilkan asam lemak mudah terbang, karbon dioksida (CO2), hidrogen (H2) dan massa mikroba. Pada prinsipnya pembentukan
gas metan di dalam rumen terjadi melalui reduksi CO2 oleh H2 yang dikatalisis
oleh enzim yang dihasilkan Archaea metanogen menurut jalur reaksi seperti berikut:
CO2 + 4H2→ 4 + 2H2O ; ∆ ⁰ = -32.75 kJ mol-1 H2 (Vlaming 2008).
Lebih dari 60 spesies metanogen yang diisolasi dari berbagai habitat yang berbeda, namun hanya lima jenis metanogen yang telah diisolasi dalam rumen yaitu Methanobacterium formicicum, Methanobrevibacter ruminantium, Methanosarcina barkeri, Methanosarcina mazei, dan Methanomicrobium mobile. Diantara kelima spesies tersebut hanya Methanobrevibacter ruminantium dan Methanosarcina barkeri yang telah ditemukan pada populasi >106 koloni ml-1 dalam rumen dan dianggap berperan penting pada proses metanogenesis di dalam rumen (Moss et al. 2000).
Komposisi asam lemak terbang yang dihasilkan selama proses fermentasi pakan di dalam rumen akan sangat berpengaruh terhadap produksi gas metan. Sehingga menurut Moss et al. (2000) produksi gas metan dapat diestimasi dari konsentrasi VFA parsial berupa asam asetat (C2), asam propionat (C3) dan asam
butirat (C4). Menurut Negara (2012) garis persamaan Moss et al. (2000) lebih
mendekati garis ideal, garis ideal adalah garis dimana nilai dari CH4 melalui
estimasi sama dengan CH4 melalui pengukuran, sehingga menunjukkan model
14
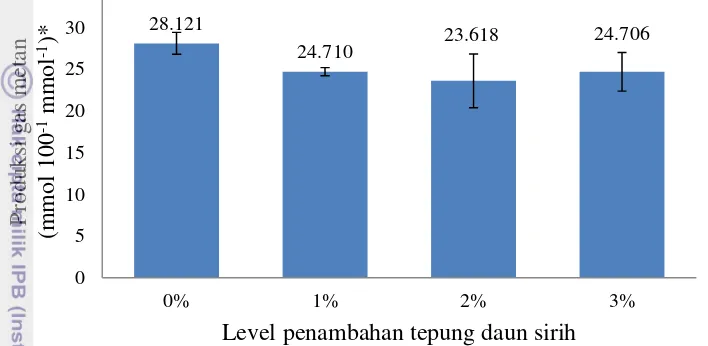
Hasil perhitungan produksi gas metan pada penelitian ini ditampilkan dalam Gambar 5. Penambahan tepung daun sirih dengan level penambahan 1%-3% pada ransum belum optimal mengurangi produksi gas metan secara nyata (P>0.05).
Gambar 5 Estimasi produksi gas metan (CH4) *(P=0.206) Perhitungan
dengan menggunakan metode Moss et al. (2000).
Berdasarkan hasil penelitian, rataan produksi gas metan pada perlakuan yaitu 23.618-28.120 mmol 100-1 mmol-1. Penambahan 2% tepung daun sirih menunjukan hasil produksi gas metan lebih rendah yaitu 23.618 mmol 100-1 mmol-1. Hal ini besar kemungkinan disebabkan karena meningkatnya propionat secara nyata (P<0.05) dalam rumen.
Proporsi asetat pada ransum dengan penambahan 2% tepung daun sirih juga lebih rendah. Pada saat terjadi produksi asam asetat di dalam rumen, dihasilkan H2
dan CO2 yang dimanfaatkan oleh metanogen untuk proses metanogenesis. Jika
jumlah asetat yang dihasilkan menurun maka H2 dan CO2 yang dihasilkan
menurun. Hal tersebut dapat mengganggu proses metanogenesis, sehingga produksi metan mengalami penurunan. Hasil estimasi produksi metan ini sesuai dengan menurunnya nilai perhitungan rasio C2:C3 pada Tabel 3 yang secara nyata
dipengaruhi oleh penambahan tepung daun sirih. Selain itu hasil perhitungan jumlah protozoa dalam penelitian ini menunjukkan penurunan jumlah populasi protozoa dengan semakin tingginya dosis penggunaan tepung daun sirih. Menurunnya jumlah protozoa ini dapat menyebabkan menurunnya jumlah Archaea metanogen yang bersimbiosis dengan protozoa.
Pada penggunaan tepung daun sirih sebanyak 3% produksi gas metan mengalami peningkatan dibanding dengan penggunaan tepung daun sirih sebanyak 2% tetapi tidak lebih tinggi dari ransum kontrol, padahal jumlah protozoa pada taraf 3% tepung daun sirih menunjukkan hasil yang paling kecil. Peningkatan produksi gas metan tersebut diduga karena proporsi asetat pada penggunaan 3% tepung daun sirih menunjukkan hasil yang lebih besar dibanding
15 taraf 2%, selain itu proporsi propionat dan valerat untuk ransum 3% tepung daun sirih proporsinya mengalami penurunan sesuai data Tabel 3.
Kinetika Produksi Gas
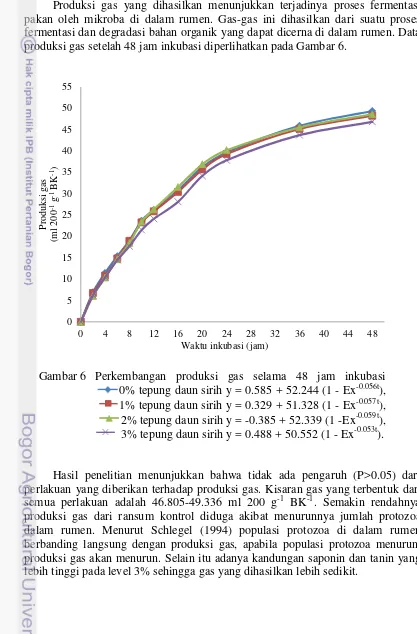
Produksi gas yang dihasilkan menunjukkan terjadinya proses fermentasi pakan oleh mikroba di dalam rumen. Gas-gas ini dihasilkan dari suatu proses fermentasi dan degradasi bahan organik yang dapat dicerna di dalam rumen. Data produksi gas setelah 48 jam inkubasi diperlihatkan pada Gambar 6.
Gambar 6 Perkembangan produksi gas selama 48 jam inkubasi 0% tepung daun sirih y = 0.585 + 52.244 (1 - Ex-0.056t), 1% tepung daun sirih y = 0.329 + 51.328 (1 - Ex-0.057t),
2% tepung daun sirih y = -0.385 + 52.339 (1 -Ex-0.059t), 3% tepung daun sirih y = 0.488 + 50.552 (1 - Ex-0.053t).
Hasil penelitian menunjukkan bahwa tidak ada pengaruh (P>0.05) dari perlakuan yang diberikan terhadap produksi gas. Kisaran gas yang terbentuk dari semua perlakuan adalah 46.805-49.336 ml 200 g-1 BK-1. Semakin rendahnya produksi gas dari ransum kontrol diduga akibat menurunnya jumlah protozoa dalam rumen. Menurut Schlegel (1994) populasi protozoa di dalam rumen berbanding langsung dengan produksi gas, apabila populasi protozoa menurun, produksi gas akan menurun. Selain itu adanya kandungan saponin dan tanin yang lebih tinggi pada level 3% sehingga gas yang dihasilkan lebih sedikit.
16
Menurut hasil penelitian Masruroh (2013) penggunaan saponin dari ekstrak buah lerak mampu menurunkan produksi gas. Suplementasi bahan pakan yang mengandung tanin dapat menurunkan gas total dari sistem fermentasi rumen secara In vitro (Sajati 2012). Selain menurunnya jumlah protozoa, ransum dengan penambahan 3% tepung daun sirih memiliki nilai potensial degradasi yang lebih rendah dibanding ransum perlakuan lain, sehingga tingkat fermentasi dan degradasi bahan organiknya rendah. Rendahnya kemampuan fermentasi tersebut mengakibatkan jumlah gas yang dihasilkan juga semakin sedikit (Firsoni 2005).
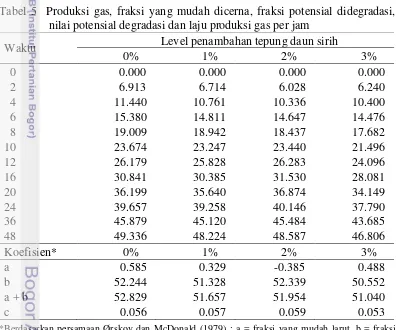
Pola produksi gas dapat menggambarkan laju degradasi bahan organik. Produksi gas terus meningkat dari 2 jam inkubasi hingga akhirnya mulai melambat setelah 24 jam inkubasi. Tabel 5 menunjukkan nilai koefisien fraksi yang mudah larut (a), fraksi yang potensial didegradasi (b), nilai potensial degradasi (a + b), dan laju produksi gas per jam (c). Penambahan tepung daun sirih tidak menunjukkan pengaruh nyata (P>0.05) terhadap nilai koefisien (a), koefisien (b), koefisien (a + b), dan koefisien (c).
Tabel 5 Produksi gas, fraksi yang mudah dicerna, fraksi potensial didegradasi, nilai potensial degradasi dan laju produksi gas per jam
Waktu Level penambahan tepung daun sirih
0% 1% 2% 3%
*Berdasarkan persamaan Ørskov dan McDonald (1979) ; a = fraksi yang mudah larut, b = fraksi yang potensial didegradasi, a + b = nilai potensial degradasi, c = laju produksi gas per jam.
17 kandungan nutrien yang berbeda antara lingkungan baru dengan lingkungan sebelumnya sehingga diperlukan waktu penyesuaian untuk mensintesa enzim-enzim hingga akhirnya bakteri masuk ke dalam fase log (pertumbuhan eksponensial). Hal ini sesuai dengan hasil produksi gas pada ransum dengan penambahan 2% tepung daun sirih yang cenderung lambat pada jam-jam pertama dan mengalami peningkatan laju produksi gas pada jam ke-12 hingga jam ke-24 lebih tinggi dibanding dengan perlakuan lainnya.
Estimasi Nilai DBO dan Nilai EM
Estimasi degradasi bahan organik (DBO) dan energi termetabolis (EM) dapat dihitung berdasarkan formula Menke et al. (1979) yang berdasarkan produksi gas dan komposisi nutrien ransum perlakuan. Hasil perhitungan pada penelitian ini disajikan pada Tabel 6.
Tabel 6 Nilai DBO dan EM
Level penambahan tepung daun sirih DBO (%)* EM (kkal kg-1 DM-1)*
0% 51.162±1.077 2044.364± 39.370
1% 50.807±1.372 2031.849± 50.158
2% 51.596±2.242 2061.179± 81.972
3% 49.501±3.017 1985.042±110.303
P value P=0.222 P=0.229 *Berdasarkan persamaan Menke et al. (1979).
Penambahan tepung daun sirih pada berbagai level tidak menunjukkan pengaruh nyata (P>0.05) terhadap nilai DBO dan nilai EM. Terdapat kesamaan pola antara nilai DBO dan EM pada masing-masing perlakuan. Terjadi penurunan nilai DBO dan EM pada pemberian 1% tepung daun sirih. Pada saat ditambah 2% tepung daun sirih, nilai DBO dan EM meningkat bagus dan turun lebih dari kontrol saat ditambah 3% tepung daun sirih. Hasil tersebut sesuai dengan perhitungan nilai fraksi yang mudah didegradasi (koefisen b) yang menunjukkan nilai koefisien (b) pada level pemberian 2% tepung daun sirih lebih besar nilainya jika dibanding dengan level 0%. Menurut Yamin et al. (2013) penambahan tepung daun sirih berlevel tidak memperlihatkan pengaruh nyata terhadap nilai kecernaan bahan kering (KCBK) dan kecernaan bahan organik (KCBO), namun penambahan 2% tepung daun sirih mampu meningkatkan KCBK dan KCBO, peningkatan level penambahan tepung daun sirih hingga 8% menurunkan KCBK dan KCBO.
18
SIMPULAN DAN SARAN
Simpulan
Penambahan tepung daun sirih dengan level 1%-3% dalam ransum belum nyata menurunkan gas metan yang dihasilkan di saluran pencernaan ternak ruminansia tetapi mampu meningkatkan proporsi propionat dan butirat dengan level optimal 2% tepung daun sirih dalam ransum.
Saran
Perlu adanya identifikasi bakteri rumen dengan teknik poly chain reaction (PCR) sehingga dapat diketahui pasti jumlah metanogen yang masih ada dalam rumen dan metode mitigasi dengan bahan lain yang langsung membunuh metanogen tanpa mengurangi proporsi asetat untuk pembentukan lemak susu.
DAFTAR PUSTAKA
[AOAC] Assosiation of Official Analytical Chemist. 2003. Official Method of Analysis of The Association of Official Analytical of Chemist. Virginia (US):AOAC.
Arora SP.1989. Pencernaan Mikroba pada Rumen. Yogyakarta (ID): UGM Pr. Bryden WL, Annison EF. 1998. Prespectives on Ruminant Nutrition and
Metabolism. Department of Animal Sci. Camden (AU): Univ of Sydney. Castillejos LS, Calsamiglia, Ferret A. 2006. Effect of essential oil active
compounds on rumen microbial fermentation and nutrient flow in In vitro systems. J Dairy Sci. 89:2649-2658.
Church DC. 1976. Digestive Physiology and Nutrition of Ruminants. Oregon (GB): Oxford Univ Pr.
Damron WS. 2006. Introduction to Animal Science. Ohio (US): Prentice Hall. Daning DRA. 2010. Limbah teh hitam (Bohea bulu) sebagai agen defaunasi
terhadap reduksi gas metan pada fermentasi rumen dalam mendukung peternakan ramah lingkungan. [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Darwis SN. 1991. Potensi sirih (Piper betle Linn.) sebagai tanaman obat. Warta Tumb Obat Indones. 1(1):11-12.
Firsoni. 2005. Manfaat tepung daun kelor (Moringa oleifera, Lam) dan glirisidia (Gliciridia Sepium, Jacq) sebagai sumber protein dalam urea molases blok (UMB) terhadap metabolisme pakan secara In vitro dan produksi susu sapi perah. [tesis]. Malang (ID): Univ Brawijaya.
19 France J, Siddon RC. 1993. Volatile Fatty Acids Production. In: Forbers. Quantitative Aspect Ruminant Digestion and Metabolism. Wallingford (GB): CABI.
General Laboratory Procedure. 1966. Department of Dairy Sci.Madinson (WI): Univ of Wiscosin.
Haryanto B, Thalib A. 2009. Emisi metana dari fermentasi enterik: kontribusinya secara nasional dan faktor-faktor yang mempengaruhinya pada ternak. Wartazoa. 19(4):157-165.
Hobson PN, Jouany JP. 1988. Models Mathematical and Biological of The Rumen Funcion. London (GB): Elsevier Science.
Hungate RE. 1966. The Rumen and Its Microbes. New York (US): Academic Pr. Jayanegara A, Sofyan A. 2008. Penentuan aktivitas biologis tanin beberapa
hijauan secara In vitro menggunakan hohenheim gas test dengan polietilen glikol sebagai determinan. Med Pet. 31:44-52.
Jouany JP. 1991. Defaunation of The Rumen. In: Rumen Microbial Metabolism and Ruminant Digestion. Paris (FR): INRA.
Martin C, Doreau M, Morgavi DP. 2008. Methane Mitigation in Ruminants: From Rumen Microbes to The Animal. Paris (FR): Herbivores Research Unit. Masruroh S. 2013. Populasi protozoa dan produksi gas total dari rumen kambing
perah yang pakannya di suplementasi ekstrak herbal secara In vitro. JIP. 1(2):420-429.
McDonald PR, Edwards A, Greenhalg JFD, Morgan CA. 2002. Animal Nutrion. Ed ke-6. New York (US): John Willey.
Menke KH, Raab L, Salewski A, Steingab H, Schneider W. 1979. The estimation of digestibility and metabolizable energy content of ruminant feedstuff from the gas production when they are incubated with rumen liquor. J Agric Sci. 93:217-222.
Moeljanto RD. 2003. Khasiat & Manfaat Daun Sirih: Obat Mujarab dari Masa ke Masa. Jakarta (ID): Agromedia Pustaka.
Morgavi DP, Forano E, Newbold CJ. 2010. Microbial ecosystem and methanogenesis in ruminants. NCBI. 4(7):1024-1036.
Moss AR, Jouany JP, Newbold J. 2000. Methane production by ruminants: its contribution to global warming. Ann Zootech. 49:231-253.
Nalina T, Rahim ZHA. 2007. The crude aqueous extract of Piper betle L. and its antibacterial effect towards Streptococcus mutans. Am J Biochemist Biotech. 3(1):10-15.
Negara HP. 2012. Prediksi emisi gas metana pada ransum mengandung tanin dalam sistem rusitec melalui komposisi asam lemak terbang. [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Newbold CJ, Lassalas B, Jouany JP. 1995. The importance of methanogens associated with ciliate protozoa in ruminal methane production In vitro. Lett Appl Microbiol. 21:230-234.
Ørskov ER, McDonald I. 1979. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J Agric Sci. 92:499-503.
20
Schlegel HG. 1994. Mikrobiologi Umum. Penerjemah: T. Baskoro. Yogyakarta (ID): UGM Pr.
Steel RGD, Torrie JH. 1993. Prinsip dan Prosedur Statistik. Suatu Pendekatan Biometrik. Jakarta (ID): Gramedia Pustaka Utama.
Tilley JM, Terry RA. 1963. Two-stage technique for In vitro digestion of forage crop. J Brit Grass Soc. 18:104-111.
Vlaming JB. 2008. Quantifying variation in estimated methane emission from ruminants using the sf6 tracer fechnique. [thesis]. New Zealand (AU):
Doctor of Phylosophy in Animal Sci. Massey Univ.
Wallace RJ, McEwan NR, McLntosh FM, Teferedegne B, Newbold CJ. 2002. Natural product as manipulatiors of rumen fermentation. J Anim Sci. 15(10):1458-1468.
Karakashev D, Batstone DJ, Angelidaki I. 2005. Influence of environtmental conditions on methanogenic compositions in anaerobic biogas reactors. Appl Environ Microbiol. 71:331-338.
21 Lampiran 1 Analisis ragam nilai pH rumen
Sumber Jumlah
Lampiran 2 Analisis ragam VFA total
Sumber Jumlah
Lampiran 3 Analisis ragam proporsi asam asetat (C2)
22
Lampiran 4 Analisis ragam proporsi asam propionat (C3)
Sumber Jumlah
Lampiran 5 Uji polinomial ortogonal proporsi asam propionat (C3)
Sumber Jumlah
Lampiran 6 Analisis ragam proporsi asam butirat (C4)
Sumber Jumlah
Lampiran 7 Uji polinomial ortogonal proporsi asam butirat (C4)
23 Lampiran 8 Analisis ragam proporsi isobutirat (IC4)
Sumber Jumlah
Lampiran 9 Analisis ragam rasio C2:C3
Sumber Jumlah
Lampiran 10 Uji polinomial ortogonal rasio C2:C3
Sumber Jumlah
Lampiran 11 Analisis ragam proporsi valerat (C5)
24
Lampiran 12 Analisis ragam proporsi isovalerat (IC5)
Sumber Jumlah
Lampiran 13 Analisis ragam populasi protozoa
Sumber Jumlah
Lampiran 14 Uji polinomial ortogonal populasi protozoa
Sumber Jumlah
Lampiran 15 Analisis ragam produksi gas metan (CH4)
25 Lampiran 16 Analisis ragam produksi gas total
Sumber Jumlah
Lampiran 17 Analisis ragam koefisien (a)
Sumber Jumlah
Lampiran 18 Analisis ragam koefisien (b)
26
Lampiran 19 Analisis ragam koefisien (a + b)
Sumber Jumlah
Lampiran 20 Analisis ragam koefisien (c)
Sumber Jumlah
Lampiran 21 Analisis ragam estimasi DBO (Degradasi Bahan Organik)
27 Lampiran 22 Analisis ragam estimasi EM (Energi Metabolis)
Sumber Jumlah
kuadrat
Derajat bebas
Kuadrat
tengah F hitung Sig. Faktor terkoreksi 0.796a 5 0.159 5.429 0.031 Intercept 866.239 1 866.239 2.955E4 0.000 Kelompok 0.628 2 0.314 10.705 0.010 Perlakuan 0.168 3 0.056 1.912 0.229
Galat 0.176 6 0.029
Total 867.211 12
28
RIWAYAT HIDUP
Penulis dilahirkan di Lamongan, Jawa Timur pada tanggal 11 Januari 1991. Penulis merupakan anak kedua dari dua bersaudara dari pasangan Bapak H.Soepomo dan Ibu Hj.Siti Mardhiyah. Pada 2006 penulis diterima di SMA Negeri 2 Lamongan. Penulis diterima di Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor pada tahun 2009 melalui jalur Undangan Seleksi Masuk IPB (USMI).
Selama mengikuti perkuliahan penulis aktif dalam
berbagai kegiatan kemahasiswaan, Siswa Leadership and Enterpreneurship School (LES) 2009-2010, Staff public relation di LES BEM KM IPB tahun 2010-2011, Anggota biro kewirausahaan BEM Fakultas Peternakan IPB kabinet
D’S -2011. Kepala biro kewirausahaan BEM Fakultas
D’O s 2011-2012. Penulis pernah mengikuti kegiatan
magang di Balai Besar Inseminasi Buatan Singosari, Malang, Jawa Timur 2012. Beberapa prestasi yang diraih oleh penulis antara lain PKMP didanai DIKTI pada tahun 2012 dan 2013 sebagai peneliti utama, 104 Inovasi IPB dalam PIM selaku dosen pembimbing akademik sekaligus dosen pembimbing skripsi, dan Dr Anuraga Jayanegara, SPt, MSc selaku dosen pembimbing skripsi yang telah mendukung, memotivasi dan membimbing selama penyelesaian tugas akhir dengan penuh kesabaran. Ibu Dr Ir Dwierra Evvyernie A, MS MSc yang telah memberi arahan dan semangat selama menyelesaikan tugas akhir. Ibu Dian Anggraeni dan Ibu Adriani, SSi atas bantuannya di laboratorium. Ir Kukuh Budi Satoto, MSi selaku dosen pembahas dan Dr Iwan Prihantoro, SPt, MSi selaku dosen panitia dalam seminar hasil penelitian. Kepada Dr Ir Idat Galih Permana, MScAgr dan Bapak Ahmad Yani, STP, MSi selaku dosen penguji serta Ibu Dilla Mareistia Fassah, SPt, MSc selaku panitia ujian akhir sarjana.