1;- ~
::
~-
Bul. Agron. (29) (2) 40
-
49 (2001)
tL
')
.-: ';
I' "~
" :j;
,
,i
,
Pemetaan
Marker AFLP untuk Membuat Peta Genetik Bit Gula
,
j, ...:;,:;
i
'I
Mapping of ALP Marker to Construct a Genetic Map Sugar Beet
,'"'I ;
i
,
l' #
I'
; Asep Setiawan!)
!
i'
1 ~
i!
r ABSTRACT
j!
,J One hundred eighty two AFLP marker using primer combination of EcoR//MseI and PstI/MseI were used in this
,
i
study to create a DNA marker genetic map of Sugar beet. In average each primer combination yielded 15.51:
polymorphic
band.
AFLP marker using
primer combination
of EcoR//MseI
signifcantly
yielded more
polymorphic
band
! than PstI/MseI. From this study a high density DNA marker map coverage all nine chromosomes of sugar beet with
I
totally length of 744 cM Haldane
was established.
,
.! i
Ii Key words: AFLP, Genetic map, Restriction enzyme, Polymorphic ,;;il
~ ci
,l ';!
: PENDAHULUAN Botstein, 1989, Yano et al., 1997, Setiawan et al., ;"
,
I
: 2000).
; Peta genetik terbukti bennanfaat sebagai alat untuk Peta keterpautan dengan jarak antar marker lebih ,; studi genetik. Pembuatan peta genetik pada dasanya daTi 5 cM dikategorikan sebagai peta yang berdensitas (jo i
berprinsip pada fenomena keterpautan antara antara dua rendah atau moderat (Tanksley, et al., 1992). :::1
1
gen yang telah lama dikenal oleh Punett daD Bateson DNA marker berbasis PCR Amplified Fragment ;~
': (Suzuki et al., 1986). Length Polymorphism (AFLP) yan menjanjikan hasil -oil
Dua gen dikatakan terpaut apabila allel-allel dari bebas dari artefak daD dapat diandalkan telah ~,
; kedua gen yang berasal dari tetua yang sarna sering atau diperkenalkan oleh Vos et al. (1995). Keunggulan :'
1 :. senantiasa dijumpai pada individu yang sarna. Jarak AFLP selain untuk tanaman telah pula ditunjukkan oleh
antara dua gen yang terpaut dinyatakan dalam cM. Satu Majer et al. (1996) untuk studi biologi molekular daD
~-I
cM didefinisikan sebagai peluang, bahwa 1% rekom- keragaman genetik pada cendawan. ~
,
binan akan dijumpai dari suatu persilangan. Gen dengan Teknik AFLP berbasiskan amplifikasi selektif dari il jarak I cM akan lebih sulit terpisah dibanding gen fragmen-fragmen DNA hasil restriksi dari total genomic
dengan jarak 2 cM. Dengan berkembangnya marka DNA dengan ensim restriksi endonuklease (V os et al.,
molekular, keterpautan genetik kini menjadi semakin 1995). Teknik ini melibatkan tiga tahapan utama yaitu:
penting untuk dikaji daD penggunaannya semakin Restriksi DNA daD ligasi adapter-adapter, amplifikasi
! meluas tidak hanya oleh ahli genetik tapi juga oleh para selektif dari fragment-fragmen restriksi (restriction
pemulia dan ahli biologi molekular. fragments), daD analisis gel dari produk amplifikasi.
Peta keterpautan berdensitas tinggi untuk beberapa
I: tanaman seperti padi dengan 1300 marker (Kurata et al
, 1994), tomat dengan 1030 marker (Tanksley, et al., Tujuan
1992), daD Kedelai dengan 840 marker (Keirn et al.,
ij ] 997). Peta keterpautan dengan densitas tinggi dapat Studi ini bertujuan untuk menilai efisiensi AFLP
berfungsi untuk berbagai keperluan seperti untuk marker dalam menghasilkan marker polimorf,
mem-kromosom walking (Wickman dan Williamson, 1991), bandingkan dua ensim pemotong jarang yaitu Eco RI .
i
marker-based
selection untuk memepercepat
program
dan Pstl dalam menghasilkan
marker polimorf dan
I1
pemuliaan
(Tanksley
et al., 1989; Paterson
et al., 1988)
membuat
peta keterpautan
marker pada tanaman
bit
~l dan untuk mendeteksi dan karakterisasi lokus yang gula yang akan digunakan pada tahap selanjutnya yaitu
mengendalikan sifat kuantitatif atau untuk memfasilitasi sebagai basis pemetaan QTL bagi sifat resistensi
diseksi sifat kuantitatif kedalam faktor-faktor genetik terhadap penyakit bercak daun Cercospora.
yang diskret (Paterson, et al., 1988; Lander daD
-"c ;;;j
I) StafPengajar Jurusan Budidaya Pertanian IPB, Bogor ~
j ..
40
.
Bul. Agron. (29) (2) 40
-
49 (2001)BAHAN DAN METODE Skoring data dilakukan secara manual dengan cara
memberi skor A/C atau BID untuk satu AFLP marker. A
Ekstraksi DNA = tidak acta pita, C = acta pita yang diperoleh dari tetua I.
. . . . B = tidak acta pita, D = acta pita yang diperoleh dari
Metode yang dlgunakan pacta pnnslspnya sepertl t tu 2 ~ yang telah diuraikan oleh Saghai-Maroof et al. (1984). ea.
Untuk ekstraksi DNA sebanyak 5 gram daun segar P t t'
k
d. b'l d " F D d. b.
t
enyusunan pe a gene Ilam I arl setlap tanaman 2. aun yang lam I
.
diusahakan yang belum terlalu tua dan tidak kotor. Untuk menyusun peta genetik dari populasi F2Selanjutnya daun di masukkan dalam kantong plastik basil silangan dua tetua yaitu no 93 164P dan no. 95098P
dan dikeringkan dengan pengering vakum (vacuum- digunakan program Mapmaker/EXP 3.0 (Lander et al.,
dryer) selama 3 hari. Daun yang telah kering 1987; Lincoln et al., 1993). Marker-marker dalam
selanjutnya disimpan pacta suhu 20 ° C menunggu saat kromosom dikatakan terpaut apabila mereka tidak
ekstraksi tiba. bersegregasi secara bebas. Perintah group dengan LaD
Tepung daun yang acta didalam tabung 50 ml skore = 4 dan jarak 25 cM Haldane ditetapkan sebagai
selanjutnya dicampur dengan buffer untuk ekstraksi kriteria untuk menetapkan keterpautan antar marker.
DNA dan diinkubasi pacta suhu 65° C selama 30 - 60 Prosedur detail penggunaan Mapmaker dapat dilihat
menit. Selanjutnya campuran kloroform-isoamil pacta manual Mapmaker/exp 3.0.
alkohol (24: I) dengan volume sebanding dengan buffer untuk ekstraksi DNA ditambahkan dalam setiap tabung.
Pemisahan DNA dengan komponen set lain dilakukan HASIL DAN PEMBAHASAN
dengan sentrifugasi (20 °C 7500 RPM, 20 menit,
senti fugal tipe J2-HC Beckmann). Larutan jernih yang, Dalam penelitian ini 189 DNA total dari tanaman
mengandung DNA selanjutnya dipindahkan ke tabung F2 dan tetuanya dianalisa menggunakan teknik AFLP.
50 ml lain yang baru. Isopropanol ditambahkan lalu Delapan primer kombinasi Eco RI/Msel (ElM) dan
tabung ditaruh dalam es selama 30 menit. Setelah DNA enam primer kombinasi PstI/Msel (PIM) digunakan
terpresipitasi, selanjutnya DNA dipindahkan kedalam dalam penelitian ini. Ke 15 primer kombinasi ini
t
tabung reaksi 2ml. Pelarutan DNA selanjutnya diperoleh d'iifi basil screening primer yang dilakukan; dilakukan dengan menggunakan I - 2 ml O. I x TE. terhadap 7 tanaman F2. Ke 8 ElM primer kombinasi
Konsentrasi DNA selanjutnya diturunkan hingga 10 - menghasilkan 151 pita polimorf atau rata-rata 18.8 pita
12 ngiuL. polimorf per primer kombinasi. Kombinasi PIM primer
Untuk menguji mutu DNA dilakukan uji restriksi. menghasilkan 66 pita polimorf dengan rata-rata 11 pita
i Secara acak 20 sample DNA diambil untuk direstriksi polimorf per primer kombinasi. Secara-rata-rata jumlah
r dengan EcoRI. 100 ng DNA di cerna (digesting) dengan
-
pita polimorf yang dihasilkan oleh kombinasi primerI menggunak~ 5 unit ~nsim Eco RI. Hasil analisa de~gan ElM nyata lebih tinggi dari pacta yang dihasilkan oleh
I
elektroforesls menunJukkan seluruh sample DNA tldak PIM. (TabeI1.).mengalami partial digestion. Pacta studi ini 217 lokus AFLP marker telah
dianalisa. Contoh pola pita AFLP dapat dilihat pacta
f AnalisisAFLP gambar 1. Secara keseluruhan 14 primer kombinasi
i yang digunakan menghasilkan secara rata-rata 15.5 pita
, AFLP dilaksanakan sesuai uraian Vos et al. I' rf
(1?95). Pre.amplifikasi dil~ukan dengan dua primer po Im~e~nggulan AFLP telah dilaporkan oleh banyak
ollgonukleotld Eco RI-A (pnmer y~g homolog dengan penulis setelah teknologi AFLP diintroduksikan oleh
adapter Eco RI dan Msel~C (pr~mer yang. ~o~olog Vos et al. (1995). Banyak peta marker telah disusun
dengan adapter Msel), setlap prImer memlllkl satu dengan menggunakan AFLP marker. Pacta bit gula
.
n~kleotida selektif. Preamplifikasi ~an amplifikasi misalnya oleh Schondelmaier et al. (1996) dan Barnesdllakukan d~ngan .menggunakan PerkIn Elmer 9~00. et al. (1996). Penyusunan peta marker berdensitas tinggi
- Pelabelan prImer dlla~uk~ dengan cara melabel prImer pada kentang juga telah dilakukan oleh Qi dan Lindhout
!
r Eco ~ +3.de.n~an radloaktlfgamma P33~ATP. P~oduk (1997) serta van Eck et al. (1995). Keunggulan AFLP f ampllfikasl dlpls~kan pada polyacryl~lde gel 6 Yo, 60 salah satunya adalah karena marker ini efisien dalam. " w.att . selama leblh. k~rang 1?0 memt. Setelah gel menghasilkan polimorfisme dibanding marker lainnya
i dlkenngkan d~teksl smyal dll~uan dengan meng- (Schondelmaier, et al., 1996). Studi ini juga
gunakan film smar X (Kodak Blomax MR-I, Eastman menunjukan hal serupa AFLP dinilai efisien
Kodak) selama 2 - 3 hari dalam kaset film sinar X. menghasilkan pita polimorf:
, I
:
I
I Asep Setiawan 41
r~
.
"
j----;
!;
Bul. Agron. (29) (2) 40 - 49 (2001)j
,.
[image:3.611.60.538.128.270.2]Cc
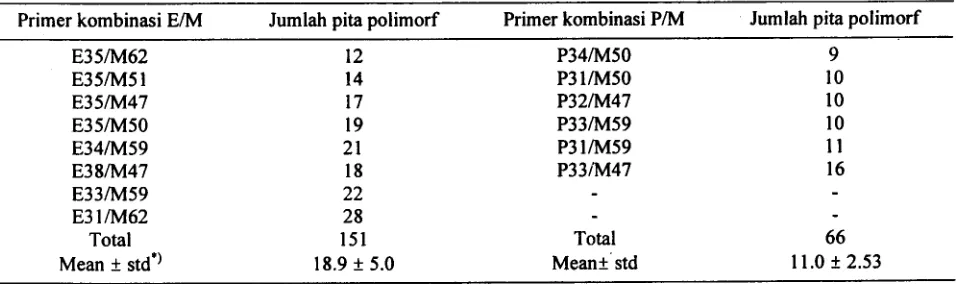
Table 1. Pita polimorf
AFLP pada 189 tanaman
F2. Menggunakan
kombinasi
primer E/M = EcoRI/MseI
clan P/M =
PstI/MseI).
Primer kombinasi E/M Jumlah pita polimorf Primer kombinasi P/M Jumlah pita polimorf
E35/M62 12 P34/M50 9
.
E35/M51 14 P31/M50 10
E35/M47 17 P32/M47 10
E35/M50 19 P33/M59 10 ~
E34/M59 21 P31/M59 11
~~~~~~ ~~ P33~47 1_6 1.!11
E31/M62 28
-
-Total 151 Total 66
i
Mean .t std') 18.9.t 5.0 Mean.tstd 11.0 + 2.53
-
'') Std
= standar
deviasi; Berdasar t-student
test (a = 0.01), jumlah rata-rata
pita polimorftk yang dihasilkan
oleh
kombinasi primer E/M nyata lebih tinggi dari pada P/M (thinmg = 3.864 > t.abel = 2.90).Dalam percobaan dengan tanaman sugar bit, dari 5 harus dilakukan. DNA yang tidak mumi, biasanya
gram daun segar rata-rata dapat diperoleh sebanyak 200 akibat adanya kontaminasi fenol atau bahan penyusun
ug DNA.. Digunakannya 5 gram daun mengingat selain sellain. Hal ini seringkali menjadi penyebab terjadinya
AFLP analisa RFLP juga akan dilakukan pada populasi partial digestion.
ini. Sebelum PCR, jumlah clan kualitas dari DNA Langkah pertama dalam AFLP adalah restriksi
hendaknya diperiksa. Hanya diperlukan nanogram DNA ganda dari DNA menggunakan dua ensim berbeda yaitu
untuk pelaksanaan AFLP. Akan tetapi DNA hendaknya ensim pemotong jarang clan pemotong sering yang
diupayakan memiliki kualitas yang memadai untuk diikuti dengan pemasangan adapter yang akan mengapit
menghindari adanya artefak. Kontaminasi dengan DNA fragmen DNA. Ensim pemotong jarang yang digunakan
asing harus dihindari selama penanganan DNA, adaliih ensim yang mengenal 6 basa pada situs
kontaminasi DNA sedikit saja dapat berakibat pemotongan dalam hal ini Eco RI clan ensim pemotong
timbulnya artefak karena sistem marker ini berbasis sering yang di pakai adalah MseI yang mengenal 4
. PCR. Satu fragmen DNA asing saja berpotensi untuk sekuen basa pada situs pemotongan.
r diamplifikasi. Pemotong sering Mse 1 menghasilkan frgamen
': Fase kritis dalam AFLP adalah saat pemotongan DNA berukuran kecil dalam jumlah banyak. Kombinasi
i dengan ensim restriksi. Digesti parsial harus dihindari ensim pemotong jarang clan pemotong sering
sebab hal ini dapat berakibat timbulnya artefak (V os et diharapkan dapat menghasilkan jumlah optimum
al., 1995). Untuk menghindari hal ini maka uji restriksi fragmen DNA yang,dihasilkan sehingga pola pita AFLP
!
perlu dilakukan sebelum
tahapan
pemotongan
ganda
nantinya dapat dlskor dengan baik. Eco RI banyak
dengan menggunakan dua jenis ensim dilakukan. DNA digunakan sebab ensim ini dinilai relatif murah clan
yang terpotong
sempuma
akan terlihat sebagai
smear
masalah
restriksi
partial relatifsedikit (Vos
et al., 1995).pada elektroforesis. Apabila ada indikasi DNA tidak
Penggunaan
PstIyang juga hexameric
dimaksudkan
terpotong dengan sempuma maka pemumian DNA untuk mendapatkan penyebaran.
~~I'~!~~
.";,f,,M;;! ,!;.2i.i, 'i?\"¥ T,;i'
42 Pemetaan Marker AFLP untuk Membuat Peta Genetik Bit Gula
c~ ~ "'" '
.
y- ,
-.~"'r~ i
Bul. Agron. (29) (2) 40
-
49 (2001) II , f
llic .I
~.
I
II
III
'If !~
I
; ~:~~~547130
+
'1c .
0.0
E335910c
~~:~
~~55019c
1~:~
~~~
'*"
!.
20.9
M2
25.7
M28
I~
21.5
P335905C
25.7
P315005C
'23.9
E355010D
28.3
E335917C
i~
24.7
M 1
28.8
E335918c
29.6
E316210D
24.7
E345919C
30.2
M27
29.6
E316204D
25.2
E3162110
41.4
E345906C
40.4
M40
25.7
E3459030
42.8
E335903D
48.0
E335909D
25.7
E384707C
43.1
E3359040
48.4
E335911C
27.2
E384709D
43.2
E316206C
48.7
E355105C
34.7
P334713D
43.4
E345907D
49.2
-
E384713d
36.5
E356207c
43.4
E3162140
50.6
E316213C
38.9
P334704c
43.7
E355110D
51.0
E356204c
40.9
P334705d
45.9
P324710C
53.1
E345901C
43.0
P315907d
46.4
E384705C
56.6
M41
46.4
E384704d
61.7
P334703D
54.8
E384701C
69.4
E335919d
'596
M26
81.7
M42
,:' -
I90.4
cE345908d
!,-92.5
P315011C
97.9
E335920c
97.9
M43
100.2
E316225d
104.0
P315903d
105.3
E345905c
105.3
P315911c
111
.5
M44
118.0
E354707C
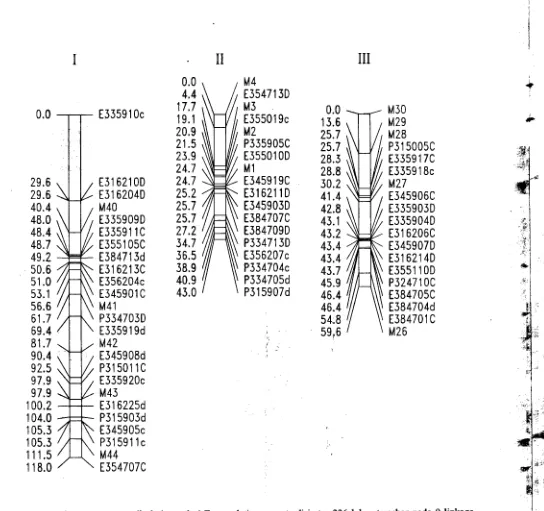
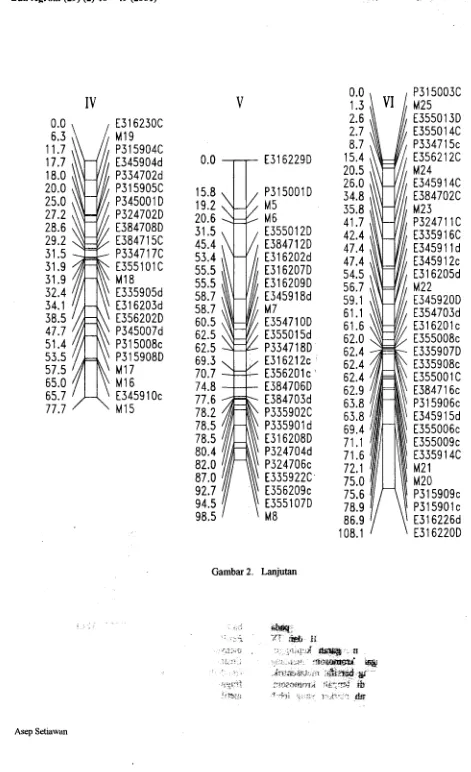
Gambar 2. Peta keterpautan genetik dati populasi F2 population yang terdiri atas 226 lokus tersebar pada 9 linkage
I groups. Kode lokus marker terletak disebelah kanan dan jarak antar marker dalam cM Haldane terletak
I
disebelah
kin. M are RFLP marker, E or P are AFLP marker dengan
primer kombinasi
EcoRI/MseI
or
".
PstI/M.'ieI. Penomoran kromosom mengikuti Schondelmaier dan lung (1977) ! "
!
~
l
44 Pemetaan Marker AFLP untuk Membuat Peta Genetik Bit Gula
.
[image:5.611.62.606.70.581.2]It hili, ~j if!, :,; ,to: ,
Bul. Agron. (29) (2) 40 - 49 (2001) -"
0.0
P315003C
IV
V
1.3 M25..
2.6E355013D
0.0
E316230C
2.7
E355014C
6.3
M19
8.7
P334715c
.~
11.7
P315904C
00
E316229D
15.4
E356212C
17.7
E345904d.
20.5
M24
18.0
P334702d
26.0
E345914C
20.0
P315905C
15.8
P315001D
34.8
E384702C
25.0
P345001
D
19.2
M5 35.8 M2327.2
P324702D
20.6
M6
41.7
P324711C
28.6
E384708D
31.5
E355012D
42.4
E335916C
29.2
E384715C
45.4
E384712D
47.4
E345911d
31.5
P334717C
53.4
E316202d
47.4
E345912c
31.9
E355101C
55.5
E316207D
54.5
E316205d
31.9
M18
55.5
E316209D
56.7
M22
32.4
E335905d
58.7
E345918d
59.1
E345920D
34.1
E316203d
58.7
M7
61.1
E354703d
38.5
E356202D
60.5
E354710D
61.6
E316201c
l
47.7
P345007d
62.5
E355015d
62.0
E355008c
I.
51.4
P315008c
62.5
P334718D
62.4
E335907D
~
53.5
P315908D
69.3
E316212c!
62.4
~
E335908c
!
57.5
M1770.7
E356201c'
62.4
E355001C
65.0
M16
74.8
E384706D
62.9
E384716c
65.7
E345910c
77.6
E384703d
63.8
P315906c
77.7
M15
78.2
P335902C
63.8
E345915d
78.5
P335901d
69.4
E355006c
l 78.5 E316208D 71.1 E355009c
t 80.4 P324704d 71.6 E335914C
82.0
P324706c
72.1
M21
87.0
E335922C'
75.0
M20
92.7
E356209c
75.6
P315909c
94.5
E355107D
78.9
P315901c
98.5
M8
86.9
E316226d
108.1
E316220D
!
I.
Gambar 2. Lanjutan
"
.
,
~:'~
'3i~{1; :;c"',.,; c;':;'[ ,"c:rc;~'l
.
t
!45 Asep Setiawan
.
[image:6.611.79.550.49.821.2]~
.
~Bul. Agron. (29) (2) 40
-
49 (2001)1
!
~ .,
;'" i!\ ~
0.0
E316223d
6.8
E316221
c
6.8
E3562060
+
8.9
E356210c
i14.9
P3359060
VII
VIII
15.7 IX
M35
00
M34
15.7
E3550050
. 15.8 E3459090
8.0 M33 0.0 P324708C 158 E345913C
14.1
P334706c
4.9
P315010C
16'8
E354716
14.5
E355004C
5.5
P334707C
16'8
E335915c
14.5
E355011c
6.3
M14
17'6
E354717~
15.9
E354708c
7.3
P3150090
19'1
E384710
16.3
E3547140
19.6
M13
19'1
~ E355113~
16.3
P315910d
20.7
E345902c
19:1
E335906d
16.3
M32
22.6
E355018d
22.3
M36
16.6
E354702d
24.9
P324701d
22.3
E355106c
."
21,.5
E316231d
33.4
M12:
22.7
E335921c
II ' 21.5 E356205C 49.6 P324703c 23 1 E335902C
~~! 22.1 E316218c 51.5 M11 .
P3
I
!;:
32.9
P3347120
61.5
E355016c
29,.1
15902c
!
36.1
P335908C
63.2
M10
30;7
E355003d
ii,
63 2
.
E335901
31.1
M37
c
31' 1
E355002c
64.3
E3847170
.
~
59.8
M31
65.2
E38471-80
32.6
E355111
C
66.5
E356211C
42.1
E345917d
68.3
E3562030
44.8
P335909c
,ft
72.6
E316224c
69.1
E316222C
46.8
E3547040
71.2
E355115C
4,6.8
E384719C
1
73.0
E354709d
~~:~
~~15OO60
I:
73.9
E354705d
55.9
E355109c
~
78.5
E355108d
59.1
E356208C
85.4
E355114c
672
M39
85.4
M9.
I
86.0
P334711c
88.3
P3347090
92.7
P334710c
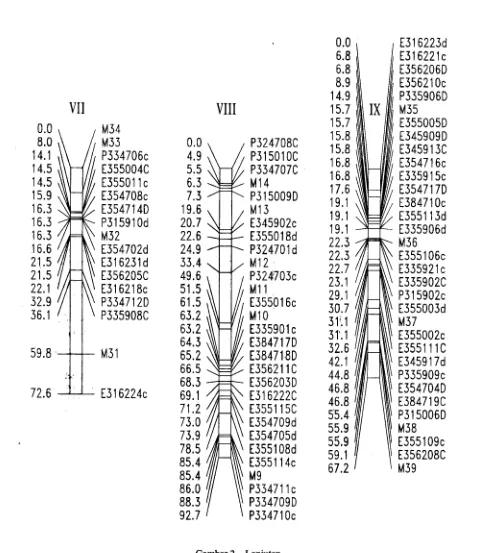
Gambar 2. Lanjutan
r
Klustering marker secara umum terjadi pada baik. Menumt Barnes et. al. (1996) AFLP menggunakan ...
bagian tengah peta. Kromosom VII, VIII dan IX EcoRI menghasilkan marker yang relatif kurnng
cenderung memiliki klustering mengarah kepinggir. menyebar dibanding AFLP menggunakan PstI. ;
~lustering pada bagian tengah kro~osom mem~g EnsimPst~ yang. ~ikenal sebag.ai sen~itive me~lasi ;;~
dlharapkan pada kromosom yang bersifat metasentrik, (methylation sensItive enzyme) diketahUl menghasilkan 't,
dimana sentromernya berada di tengah kromosom. fragmen lebih sedikit dibanding ensim non sensitif ~
Sangat beralasan bahwa didaerah marker yang lebih metilasi EcoRI. lung (1992) melaporkan bahwa
"
~
,j
I
;
46 Pemetaan Marker AFLP untuk Membuat Peta Genetik Bit Gula ~.:: . '1
zc -~:~
[image:7.611.55.542.88.641.2],
.c~~=-Bul. Agron. (29) (2) 40
-
49 (2001)
genome
bit gula sangat
kayadengan
sekuen
berulang
al., 1997)
peta keterpautan
berdasarkan
populasi
aktual
clan metilasi paling banyak terjadi pada daerah ini. yang ada setiap waktu harus dibuat lagi karena populasi
Kidwell (1998) dalam studi diversitas melaporkan aktual tersebut mungkin mengekspresikan suatu sifat
bahwa kombinasi Pst II Msel menghasilkan tingkat tertentu yang diinginkan. Dalam studi ini sifat
... diversitas yang lebih rendah dibanding menggunakan ketahanan terhadap Cercospora menjadi target. Jadi
kombinasi EcoR11 Msel. Tingkat diversitas rendah ini penyusunan peta marker ini sebenarnya merupakan
kemungkinan berkaitan dengan lebih sedikitnya bagian awal daTi usaha pemetaan sifat kuantitatif untuk
~ polimorf yang dihasilakan oleh AFLP berbasis Pstl ketahanan terhadap penyakit bercak daun (Cercospora
dibanding AFLP berbasis Eco RI. beticola Sacc.) pada bit gula.
Kesalahan dalam scoring data merupakan hal Dari studi ini berhasil dibuat peta sepanjang 744
yang kritis dalam studi yang melibatkan banyak cM Haldane yang meliputi seluruh seluruh kromosom
individu clan marker. Untuk mengurangi kesalahan haploid bit gula yang berjumlah sembi Ian kromosm (n =
maka penelitian harus dipersiapkan dengan perencanaan 9). Peta molekuler bit gula lain yang telah ada saat ini
yang baik. Kemungkinan tertukarnya contoh DNA antara lain memiliki panjang 1057 cM Haldane (Pillen
individu tanaman clan kontaminasi DNA harus dicegah. et al., 1993),688 Haldane cM (Schumacher, 1997) clan
i Kesalahan dalam skaTing masih dapat dikoreksi akan 557 cM Haldane (Schondelmaier et al., 1996). Hal ini
i
tetapi kesalahan
akibat tertukarnya
DNA contoh tidak
mengindikasikan
bahwa panjang peta daTi studi ini
lagi mungkin diperbaiki. Karena ini robotisasi menjadi secara umum sebanding dengan peta yang telah ada.
trend dalam teknologi marker yang bekerja dengan Tidak ada informasi berapa panjang sebenarnya peta
populasi dalam jumlah besar. sugar beet dapat dianggap lengkap. Oleh karena itu
AFLP termasuk kodominan marker (V os, et al., adalah lebih baik berfikir konservatif. Bila asumsi 1057
1995) akan tetapi dalam studi ini 80 % AFLP marker cM adalah peta lengkap maka studi ini telah berhasil
diskor sebagai dominan marker. Hal ini dilakukan ; mengcover 70% genom bit gula.
karena seringkali ada keraguan dalam menetapkan Dari 261 polimorfik marker yang digunakan dalam
intensitas sinyal suatu lokus marker sebagai hoterosigot studi ini, sebanyak 226 marker berhasil dikelompokkan
atau homosigot. Dengan diskor sebagai dominan marker kedalam sembilan kelompok (Gambar 2). Pada Gambar
maka lokus heterosigot tidak dapat dibedakan dengan 2 selain 182 AFLP marker terdapat juga 44 RFLP
lokus homosigot dominan. McKill et al. (1996) juga marker. Dalam tulisan ini marker RFLP tidak dibahas
melakukan skor dominan terhadap AFLP marker pada lebih lanjut, hal ini semata-mata dilakukan agar tulisan
tanaman padi akibat sulitnya menskor data AFLP dapat terfokus hanya pada AFLP marker. Seperti terlihat
sebagai marker kodominan, demikian juga Kiem et al., pada Tabel 4. Terdapat dua kromosom yang memiliki
(1997) yang bekerja dengan kedelai. Kesalahan skoring panjang lebih dari 100 cM, clan satu kromosom dengan
juga mempengaruhi presisi peta genetik (Sael dan panjang kurang daTi 50 cM. Kromosom I adalah yang
Nilsson, 1996) oleh karena itu apabila ada keraguan terpanjang (118.2 cM dan Kromosom II adalah yang
maka lebih baik menskor AFLP sebagai dominan terpendek yaitu 43.2 cM (Tabel 2). Meskipun dalam
marker dari pada sebagai kodominan marker. peta marker seperti tersaji pada gambar 2 terdapat tiga
DNA dari populasi F2 basil silangan 93164P dan jarak antar marker yang lebih dari 20 cM, jarak rata-rata
95098P digunakan menyusun peta genetik. Walaupun antara marker pada peta tersebut yang adalah 3.1 cM.
peta genetic bit gula telah banyak dibuat (Barzen, et al., Dengan demikian peta ini dapat dikategorikan dalam
1992; Hallden et al., 1996; Barnes et al., 1996; Pillen et peta dengan densitas tinggi. al., 1993; Schondelmaier et al., 1996; Schumacher et
'1£ r . !;I"~c'~::l""!
~
i,:t;;'~""'!'
:.",:'" ;:" ~:';.- ,c-:i:~,.~'"c;:'"~~
,~~
,~; ",-i.,.. .~ ";,'%li"'~ !~c,j ('"if,' -,!q;;!\ .\V;",T ;'"
':!~
Asep Setiawan 47
.
- "~"
I'! ~
,
Bul. Agron. (29) (2) 40
-
49 (2001) -,-- ,~'
.
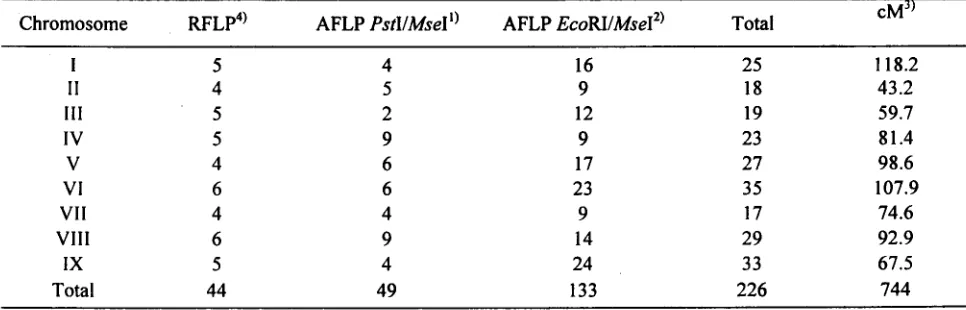
.. c..;Table 2: Penyebaran marker daD panjang peta genetik
cMJ)
Chromosome RFLp4) AFLP PstI/MseII) AFLP EcoRI/Msef) Total
I 5 4 16 25 118.2
II 4 5 9 18 43.2
~
III 5 2 12 19 59.7
IV 5 9 9 23 81.4 ~
V 4 6 17 27 98.6
VI 6 6 23 35 107.9
VII 4 4 9 17 74.6
VIII 6 9 14 29 92.9
IX 5 4 24 33 67.5
Total 44 49 133 226 744
I) AFLP markers menggunakan PstI and MseI. ;;0
2) AFLP markers using EcoRI and MseI
.
3) cM adalah menggunakan fungsi Haldane (Haldane 1919); Penomoran kromosom mengikuti sistem penomoran yang
digunakan oleh Schondelmaier and Jung (1997)
4) RFLP marker tidak dibahas da1am tulisan ini sebagai upaya memfokuskan pembahasan pada AFLP marker sekitar
sentromer memiliki frekuensi frekuensi rekombinasi yang rendah. Bosemark daD Bormotov (1971) melaporkan bahwa kromosom bit gula adalah bersifat metasentri atau sub metasentrik, ini berarti terjadinya klustering yang condong mengarah kepinggir pada kromosom VII, VIII daD IX bilkanlah disebabkan oleh posisi sentromer.
UCAPAN TERIMA KASIH Bo$emark, N.O., Bormotov, V.E. 1971. Chromosome
morphology in a homozygous line of sugar beet.
Penulis mengucapkan terima kasih kepada : Hereditas 69:205-212.
I. Prof. C. Jung daD Dr. Koch dari Dept. of crop Sci. Haldane JBS. 1919. The combination of linkage value
and Plant Breeding, Kiel University Germany atas and the calculation of distance between the loci of
segal a bimbingan daD bantuannya selama penelitian linked factors. J. Genet. 8:299-309.
ini berlangsung .
2. Ms. M. Bruisch atas bantuan teknisnya Hallden, C., Hjerdin, A., Rading, 1. M., SaIl, T.,
I 3. DAAD yang telah membiayai penulis selama Fridlundh, B.1 Johannisdottir, G., Tuvesson, S., penelitian ini berlangsung. Akesson, C., Nilsson, N-O. 1996. A high density
!
RFLP linkage map of sugar beet. Genome39:634-645. DAFTARPUSTAKA
; Keirn, P., Schupp, J.M., Travis, S.E., Clayton, K., Zhu,
Barnes S., Massaro, G., Lefebvre, M., Kuiper, M., T., Shi, L., Ferreira, A., Webb, D.M. 1997. A
Verstege, E. 1996. A Combined RFLP and AFLP High-Density Soybean .Genetic Map Based on
genetic map for sugar beet. Proceedings of the 59th AFLP Markers. Crop SCI. 37: 537-543.
IIRB Congress. Kurata, N., Nagamura, Y., Yamamoto, K., Harus Ima,h
.
Barret, B.A., Kidwell, K.K., Fox, P.N. 1998. Y., Sue, N., Wu, J., Antonio, B.A., Shomura, A.,
1
Comparison ofAFLP and Pedigree-Based Genetic Sh!mizu, T., Lin~ .S.Y., Inoe, T., Fuk~da, A.,
Diversity Assessment Methods Using Wheat Shlm~n.o, T., Kublkl, Y., Toyam~, T., Miyamoto,
Cultivars from the Pacific Northwest. Crop Sci. Y., Kmhara, T., Hayasaka, K., Mlyao, A., Monna, 'to'
38:1271-1278. L., Zhong, H.S., Tamura, Y., Wang, ZX., Momma,
T., Umehara, Y., Yano, M., Sasaki, T., Minobe, Y.
. Barzen, E., Mechelke, W., Ritter, E., Kappert, E.S. 1994. A 300-kilobase-interval genetic map of rice
1995. An extended map of the sugar beet genome including 883 expressed sequences. Nature Genet.
containing RFLP and RAPD loci. Theor. Appl. 8:365-372.
I
Genet. 90:189-193.li
48 Pemetaan Marker AFLP untuk Membuat Peta Genetik Bit Gula"'" .
[image:9.611.57.540.118.273.2]Bul. Agron. (29) (2) 40
-
49 (2001)Lander, E.S., Green, P., Abrahamson, J., Barlow, A., Schondelmaier, J., Jung, C. 1997. Chromosomal
Daly, M.J., Lincoln S.E. Newburg, L. 1987. assigment of the nine groups of sugar beet (Beta
Mapmaker: An interactive computer package for vulgaris L.) using primary trisomics. Theor. Appl.
constructing primary genetic linkage maps of Genet. 95:590-596.
experimental and natural populations. Genomics I:
4f: 174-181. Schumacher, K., Schondelmaier, J., Barzen, E.,
SteinrUcken, G., Borchardt, D., Weber, W.E.,
Lincoln, S.E., Daly, M.J., Lander, E.S. 1993. Jung, C., Salamini, F. 1997. Combining different
~ Costructing genetic linkage maps with linkage maps in sugar beet (Beta vulgaris L.) to
MAPMAKER/EXP version 3.0: A tutorial and make one map. Plant Breeding 116: 23-38.
reference manual.
Setiawan, A., G. Koch, S.A. Barnes, C. Jung. 2000.
Majer, D., Mithen, R., Lewis, B.G.,Vos, P., Oliver, R. Mapping Quantitative trait loci (QTLs) for
P. 1996. The use of AFLP finger printing for the resistance to cercospora leaf spot disease
detection of genetic variation in fungi. Mycol. Res. (Cercospora beticola Sacc.) in Sugar beet (Beta
100(9): 1107-1111. vulgaris L.). Theor. Appl. Genet. 100 (8):
1176-1182 Mckill, DJ., Zhang,Z., Redona, E.D., Co low it, P.M.
1996. Level of polymorphism and genetic Shagai-Maroof, M.A., Soliman, K.M., Jorgensen, R.A.,
mapping of AFLP markers in rice. Genome Allard, R. W. 1984. Ribosomal DNA spacer-length
39:969-977. polymorphisms in barley: Mendelian inheritance,
chromosomal location, and population dynamics.
Paterson, A. H., Lander, E. S., Hewitt, J.D., Peterson, Proc. Nat!. Acad. Sci. 81: 8014-8018.
S., Lincoln, S. E., Tanksley, S. D. 1988.
Resolution of quantitative traits into Mendelian' Tanksley, S.D., Young, N. D., Paterson, A. H.,
factors by using a complete linkage map of Bonierbale. 1989. RFLP mapping in plant
restriction fragment length polymorphisms. Nature breding: new tools for an old science. Bio
335:721-726 Technology 7: 257- 264.
1 Pillen, K., SteinrUcken, G., Herrmann, R.G., Jung, C. Van Eck H:J., van der Voort, J. R., Draaistra, J., van
1
1993. An extended
linkage map of sugar beet
Zandvoort, P., van Encvort, E., Segers, B.,
t
(Beta vulgaris L.) including nine putative lethal Peleman, J., Jacobsen, E., Helder, J., Bakker, J.genes and their restorer gene X. Plant Breeding 1995. The inheritance and chromosomal
III: 265-272. localization of AFLP markers in a non-inbreed
potato offspring. Molecular Breeding I: 397-410. Qi, X., Lindhout, P. 1997. Development of AFLP 1995.
markers in barley. Mol Gen Genet 254: 330-336.
Wicking, C., Williamson B. 1991. From linked marker
Sail, T., Nilsson, N-O. 1994. The robustness of to gene. Trends Genet. 7: 288-293.
recombination frequency estimates in intercrosses
with dominant markers. Genetics (137):589-596 Zuzuki D.T., Griffiths, A. J. F., Lewontin, R.C. 1981.
An Introduction to Genetic Analysis. 2nd ~. W.H.
Scholdelmaier, J., SteinrUcken, G. and Jung, C. 1996. Freeman and Co. San Francisco. 911p.
Integration of AFLP markers into a linkage map of sugar beet (Beta vulgaris L). Plant Breeding 115: 231-237.
.
,
.
'(c.§{~'
':;'" "'r;~"";1,,,,'1""
j
;1
Asep Setiawan 49
1;- ~
::
~-
Bul. Agron. (29) (2) 40
-
49 (2001)
tL
')
.-: ';
I' "~
" :j;
,
,i
,
Pemetaan
Marker AFLP untuk Membuat Peta Genetik Bit Gula
,
j, ...:;,:;
i
'I
Mapping of ALP Marker to Construct a Genetic Map Sugar Beet
,'"'I ;
i
,
l' #
I'
; Asep Setiawan!)
!
i'
1 ~
i!
r ABSTRACT
j!
,J One hundred eighty two AFLP marker using primer combination of EcoR//MseI and PstI/MseI were used in this
,
i
study to create a DNA marker genetic map of Sugar beet. In average each primer combination yielded 15.51:
polymorphic
band.
AFLP marker using
primer combination
of EcoR//MseI
signifcantly
yielded more
polymorphic
band
! than PstI/MseI. From this study a high density DNA marker map coverage all nine chromosomes of sugar beet with
I
totally length of 744 cM Haldane
was established.
,
.! i
Ii Key words: AFLP, Genetic map, Restriction enzyme, Polymorphic ,;;il
~ ci
,l ';!
: PENDAHULUAN Botstein, 1989, Yano et al., 1997, Setiawan et al., ;"
,
I
: 2000).
; Peta genetik terbukti bennanfaat sebagai alat untuk Peta keterpautan dengan jarak antar marker lebih ,; studi genetik. Pembuatan peta genetik pada dasanya daTi 5 cM dikategorikan sebagai peta yang berdensitas (jo i
berprinsip pada fenomena keterpautan antara antara dua rendah atau moderat (Tanksley, et al., 1992). :::1
1
gen yang telah lama dikenal oleh Punett daD Bateson DNA marker berbasis PCR Amplified Fragment ;~
': (Suzuki et al., 1986). Length Polymorphism (AFLP) yan menjanjikan hasil -oil
Dua gen dikatakan terpaut apabila allel-allel dari bebas dari artefak daD dapat diandalkan telah ~,
; kedua gen yang berasal dari tetua yang sarna sering atau diperkenalkan oleh Vos et al. (1995). Keunggulan :'
1 :. senantiasa dijumpai pada individu yang sarna. Jarak AFLP selain untuk tanaman telah pula ditunjukkan oleh
antara dua gen yang terpaut dinyatakan dalam cM. Satu Majer et al. (1996) untuk studi biologi molekular daD
~-I
cM didefinisikan sebagai peluang, bahwa 1% rekom- keragaman genetik pada cendawan. ~
,
binan akan dijumpai dari suatu persilangan. Gen dengan Teknik AFLP berbasiskan amplifikasi selektif dari il jarak I cM akan lebih sulit terpisah dibanding gen fragmen-fragmen DNA hasil restriksi dari total genomic
dengan jarak 2 cM. Dengan berkembangnya marka DNA dengan ensim restriksi endonuklease (V os et al.,
molekular, keterpautan genetik kini menjadi semakin 1995). Teknik ini melibatkan tiga tahapan utama yaitu:
penting untuk dikaji daD penggunaannya semakin Restriksi DNA daD ligasi adapter-adapter, amplifikasi
! meluas tidak hanya oleh ahli genetik tapi juga oleh para selektif dari fragment-fragmen restriksi (restriction
pemulia dan ahli biologi molekular. fragments), daD analisis gel dari produk amplifikasi.
Peta keterpautan berdensitas tinggi untuk beberapa
I: tanaman seperti padi dengan 1300 marker (Kurata et al
, 1994), tomat dengan 1030 marker (Tanksley, et al., Tujuan
1992), daD Kedelai dengan 840 marker (Keirn et al.,
ij ] 997). Peta keterpautan dengan densitas tinggi dapat Studi ini bertujuan untuk menilai efisiensi AFLP
berfungsi untuk berbagai keperluan seperti untuk marker dalam menghasilkan marker polimorf,
mem-kromosom walking (Wickman dan Williamson, 1991), bandingkan dua ensim pemotong jarang yaitu Eco RI .
i
marker-based
selection untuk memepercepat
program
dan Pstl dalam menghasilkan
marker polimorf dan
I1
pemuliaan
(Tanksley
et al., 1989; Paterson
et al., 1988)
membuat
peta keterpautan
marker pada tanaman
bit
~l dan untuk mendeteksi dan karakterisasi lokus yang gula yang akan digunakan pada tahap selanjutnya yaitu
mengendalikan sifat kuantitatif atau untuk memfasilitasi sebagai basis pemetaan QTL bagi sifat resistensi
diseksi sifat kuantitatif kedalam faktor-faktor genetik terhadap penyakit bercak daun Cercospora.
yang diskret (Paterson, et al., 1988; Lander daD
-"c ;;;j
I) StafPengajar Jurusan Budidaya Pertanian IPB, Bogor ~
j ..
40
.