i
DENGAN INDEKS GLIKEMIK RENDAH MELALUI
FERMENTASI DAN SIKLUS PEMANASAN
BERTEKANAN-PENDINGINAN
NURHAYATI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
iii
Dengan ini saya menyatakan bahwa Disertasi Peningkatan Sifat Prebiotik Tepung Pisang dengan Indeks Glikemik Rendah Melalui Fermentasi dan Siklus Pemanasan Bertekanan-Pendinginan adalah karya saya dengan arahan komisi pembimbing dan belum pernah diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Desember 2011
v
NURHAYATI. Improving prebiotic properties of low glycemic index-banana flour by fermentation and autoclaving-cooling cycle. Under direction of BETTY
SRI LAKSMI JENIE, HARSI DEWANTARI KUSUMANINGRUM and SRI
WIDOWATI.
Banana flour is a potential prebiotic source due to its resistant starch (RS) content. In this study the formation of RS type III (RS3) in banana var agung semeru (Musa paradisiaca formatypica) flour was increased in order to improve the prebiotic properties. RS-rich banana flour can be processed further to produce various kinds of functional foods. The modification process of banana flour production was conducted by a spontaneous submerged fermentation of the banana slices for 24 h at room temperature followed by one or two cycles of autoclaving (121oC, 15 min) and cooling process (4oC, 24 h). The results showed that the modification process influenced the physicochemical characteristics of banana flour. Combination of spontaneous fermentation with two cycles of autoclaving-cooling significantly increased the RS content of the modified flour about four times (28.88%). The spontaneous fermentation produced banana flour with higher amylose content. X-ray diffraction pattern of var agung semeru banana flour could be categorised as C- type granule. The native banana flour showed higher crystalinity (18.74% - 20.08%) than the modified banana flour (6.98% - 9.52%). Phenotypic and genotypic identification showed that
Lactobacillus sp associated with the spontaneous fermentation of var agung semeru banana were identified as homofermentative and heterofermentative lactic acid bacteria. They were Lactobacillus salivarius and L. fructivorans. Controlled fermentation using L. salivarius FSnh1 was conducted to produce an equivalent RS content with 24 h spontaneous fermentation. Banana slices were fermented by
L. salivarius FSnh1for 12 and 24 h at room temperature, followed by two cycles of autoclaving-cooling process. The result showed that controlled fermentation using L. salivarius FSnh1 was able to reduce the fermentation time from 24 h by spontaneous fermentation to 12 h and there fore could be recommended to replace the spontaneous fermentation. The modified banana flour showed better prebiotic properties than the native banana flour. The RS3 isolated from modified banana flour was more stable to hydrolysis performed by artificial human gastric juice,
able to modulate the growth of lactobacilli and bifidobacteria, produce butyric acid and increase prebiotic index (PI) score (5.14) than RS2 isolated from native banana flour (4.02). The modification process could also produced lower glycemic index (low GI)-banana flour than native banana flour (moderate GI).
vii
NURHAYATI. Peningkatan Sifat Prebiotik Tepung Pisang dengan Indeks
Glikemik Rendah Melalui Fermentasi dan Siklus Pemanasan Bertekanan-Pendinginan. Dibimbing oleh BETTY SRI LAKSMI JENIE, HARSI
DEWANTARI KUSUMANINGRUM dan SRI WIDOWATI.
Tepung pisang berpotensi untuk dikembangkan sebagai pangan fungsional karena mengandung komponen prebiotik yaitu pati resisten tipe II (RS2). Potensi RS2 untuk dikembangkan sebagai prebiotik memiliki kelemahan karena sifat resisten RS akan hilang jika pati mengalami gelatinisasi selama pengolahan. Oleh karena itu dilakukan modifikasi proses pembuatan tepung pisang kaya pati resisten tipe III (RS3) yang dapat bersifat lebih stabil selama pengolahan melalui fermentasi dan siklus pemanasan bertekanan-pendinginan.
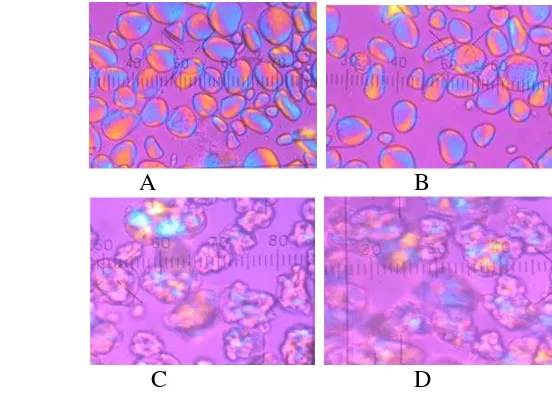
Proses modifikasi dilakukan dengan cara fermentasi spontan selama 24 jam pada suhu ruang yang dikombinasi dengan satu atau dua siklus pemanasan bertekanan (121 oC, 15 menit) dan pendinginan (4 oC selama 24 jam). Dua siklus pemanasan bertekanan-pendinginan mampu meningkatkan kadar RS lebih banyak daripada satu siklus retrogradasi. Kadar RS tepung pisang yang tinggi hingga empat kali (dari 7.24% menjadi 28.88% bk) dihasilkan dari modifikasi fermentasi spontan dengan dua siklus pemanasan bertekanan-pendinginan. Granula pati yang pecah akibat proses gelatinisasi selama pemanasan bertekanan kehilangan sifat
birefringence dan tingkat kristalinitas tepung pisang menurun dari 18.74 - 20.08% menjadi 6.98 - 9.52%. Hasil difraksi sinar X menunjukkan granula pati pisang var agung semeru adalah granula tipe C yaitu campuran granula tipe A dan tipe B yang memiliki puncak difraksi sinar X pada sudut 17o dan 24o.
viii
yang tumbuh kurang dominan (isolat BAL FSnhA) memiliki bentuk koloni bulat kecil berwarna putih bening dengan elevasi seperti tetesan, membentuk gas (heterofermentatif) serta dapat tumbuh pada suhu 15 oC maupun 45 oC. Identifikasi genotip berdasarkan sekuen gen pengkode 16S rRNA menunjukkan isolat BAL FSnh1 dan isolat BAL FSnhA termasuk famili Lactobacillaceae
dengan genus Lactobacillus. Isolat BAL FSnh1 memiliki similaritas dengan L. salivarius dan isolat BAL FSnhA memiliki similaritas denganL. fructivorans.
L. salivarius digunakan sebagai starter dalam pembuatan tepung pisang kaya RS. Fermentasi terkendali pisang oleh L. salivarius FSnh1 selama 12 jam meningkatkan kadar amilosa tepung pisang. Fermentasi 12 jam diikuti dua siklus pemanasan bertekanan-pendinginan menghasilkan kadar RS tepung pisang lebih tinggi (28.53% bk) daripada fermentasi 24 jam (25.72% bk). Penggunaan starter
L. salivarius FSnh1 mampu mempersingkat waktu fermentasi menjadi 12 jam sehingga dapat direkomendasikan sebagai pengganti fermentasi spontan.
RS3 tepung pisang modifikasi terbukti dapat memenuhi beberapa persyaratan sebagai kandidat prebiotik dengan sifat yang lebih baik di antaranya meliputi ketahanannya terhadap hidrolisis asam lambung, mampu meningkatkan pertumbuha laktobasili dan bifidobakteria, menurunkan pertumbuhan bakteri patogen, menghasilkan asam lemak rantai pendek terutama asam butirat dan memiliki indeks prebiotik lebih tinggi daripada RS2 tepung pisang kontrol. RS3 bersifat selektif terhadap pertumbuhan Lactobacillus acidophilus, EPEC dan
Salmonella Typhimurium. Persentase pertumbuhan EPEC lebih rendah daripada
L. acidophilus. Persentase pertumbuhan yang negatif untuk S. Typhimurium menunjukkan bahwa RS3 tidak dapat digunakan oleh bakteri tersebut.
ix
©Hak Cipta milik IPB, tahun 2011
Hak Cipta dilindungi Undang-undang
Dilarang mengutip sebagian atau seluruh disertasi ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
xi
DENGAN INDEKS GLIKEMIK RENDAH MELALUI
FERMENTASI DAN SIKLUS PEMANASAN
BERTEKANAN-PENDINGINAN
NURHAYATI
Disertasi
Sebagai salah satu syarat memperoleh gelar Doktor pada
Program Studi Ilmu Pangan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
xii
Penguji pada Ujian Tertutup:
• Dr. Ir. Sugiyono, M.App.Sc
• Dr. Ir. Utut Widyastuti, MS
Penguji pada Ujian Terbuka:
• Prof. Dr. Ir. Suminar Achmadi
xiii
Judul Disertasi : Peningkatan Sifat Prebiotik Tepung Pisang dengan Indeks Glikemik Rendah Melalui Fermentasi dan Siklus Pemanasan Bertekanan-Pendinginan
Nama : Nurhayati NRP : F261070101
Disetujui: Komisi Pembimbing
Prof. Dr. Ir. Betty Sri Laksmi Jenie, MS Ketua
Dr. Ir. Harsi Dewantari Kusumaningrum Dr. Ir. Sri Widowati, M.App.Sc Anggota Anggota
Mengetahui
Ketua Program Studi Dekan Sekolah Pascasarjana Ilmu Pangan
Dr. Ir. Ratih Dewanti-Hariyadi, M.Sc Dr. Ir. Dahrul Syah
xv
Karya ini kupersembahkan teruntuk orang tuaku, keluargaku,
xvii
Puji syukur kehadirat Allah S.W.T yang telah melimpahkan rahmat dan hidayah-Nya dengan terselesaikannya disertasi yang berjudul ”Peningkatan Sifat Prebiotik Tepung Pisang dengan Indeks Glikemik Rendah melalui Fermentasi dan Siklus Pemanasan Bertekanan-Pendinginan”. Disertasi ini merupakan salah satu syarat untuk mendapatkan gelar Doktor pada Program Studi Ilmu Pangan Sekolah Pascasarjana Institut Pertanian Bogor.
Penulis menyampaikan penghargaan dan ucapan terima kasih kepada Komisi Pembimbing; Prof. Dr. Ir. Betty Sri Laksmi Jenie, MS selaku Ketua Komisi Pembimbing, Dr. Ir. Harsi Dewantari Kusumaningrum dan Dr. Ir. Sri Widowati, M.App.Sc selaku Anggota Komisi Pembimbing, yang tulus, arif dan bijak membimbing studi penulis. Terima kasih penulis sampaikan kepada DIKTI yang telah mendanai penelitian ini melalui penelitian Hibah Kompetitif sesuai Prioritas Nasional tahun 2009/2010 yang diperoleh Prof. Dr. Ir. B. Sri Laksmi Jenie, MS, serta Hibah Penelitian Disertasi Doktor tahun 2011. Penulis mengucapkan terima kasih kepada Prof. Dr. Ir. Dedi Fardiaz dan Dr. Ir. Jayus selaku penguji luar komisi pada ujian prakualifikasi lisan. Terima kasih kepada Dr. Ir. Sugiyono, M.App.Sc dan Dr. Ir. Utut Wydiastuti, MS selaku penguji luar komisi pada ujian tertutup. Terima kasih kepada Prof. Dr. Ir. Suminar Setiati Achmadi, M.Sc dan Dr. Ir. Novik Nurhidayat selaku penguji luar komisi pada ujian terbuka. Kepada seluruh staf pengajar Program Studi Ilmu Pangan IPB, penulis mengucapkan terima kasih atas ilmu yang telah diajarkan.
Penulis juga menyampaikan terima kasih kepada Dirjen DIKTI dan pengelola beasiswa I-MHERE Universitas Jember yang telah memberikan beasiswa S3 kepada penulis serta kepada civitas akademika Jurusan Teknologi Hasil Pertanian Fakultas Teknologi Pertanian Universitas Jember atas dukungannya selama studi penulis. Terima kasih kepada rekan-rekan mahasiswa Ilmu Pangan khususnya Angkatan 2006, 2007 dan 2008 atas kebersamaannya serta kepada teknisi yang telah membantu selama di Laboratorium.
Kepada kedua orangtua ibu dan bapak tercinta, penulis sampaikan terima kasih atas iringan doa, restu dan kasih sayang yang meneguhkan ruh pada setiap nafas dan kesuksesan penulis. Terima kasih kepada suami tercinta (Dedy Eko Rahmanto, S.TP, M.Si) dan ananda tersayang (Aisyah Putri Nur Rahmanto) serta adinda terkasih (Nurma Handayani) atas doa dan alunan irama cinta yang menjadikan penulis mampu memelodikan bahtera hidup untuk senantiasa menggapai ridho Allah. Terima kasih kepada keluarga bapak dan ibu mertua atas iringan doa dan restu untuk kesuksesan penulis.
Kesempurnaan merupakan hal yang amat didambakan, meskipun Allah sematalah yang merupakan Dzat Maha Sempurna. Oleh karena itu adanya saran dari pembaca terhadap hasil penelitian ini dengan senang hati akan penulis rekomendasikan pada karya berikutnya. Dengan penuh harapan, semoga penelitian ini memberikan manfaat bagi pembaca serta kemaslahatan umat.
Bogor, Desember 2011
xix
Penulis dilahirkan pada tanggal 10 April 1979 di Desa Krai Kecamatan Yosowilangun, Kabupaten Lumajang Propinsi Jawa Timur sebagai putri pertama dari dua bersaudara pasangan Bapak Satiman dan Ibu Sunarmi. Jenjang pendidikan penulis dimulai dari TK Darma Wanita Desa Krai pada tahun 1983-1985, SD Negeri Krai 02 pada tahun 1985-1991, SMP Negeri I Yosowilangun pada tahun 1991-1994, SMU Negeri I Yosowilangun pada tahun 1994-1997. Penulis menempuh pendidikan sarjana dengan gelar S.TP pada Jurusan Teknologi Hasil Pertanian Fakultas Teknologi Pertanian Universitas Jember (1997-2001) dan sejak April 2004 menjadi sarana penulis mengabdikan sebagian aktivitas keilmuan sebagai staf pengajar. Sebelumnya penulis pernah bekerja sebagai staf pengajar mental aritmatika Lembaga Pendidikan Kazoeru Surya/Primalpha (2001-2002). Penulis juga pernah menjadi staf laboran (QC dan R&D) Tepung Beras Rose Brand PT. Alu Aksara Pratama Mojokerto CV Sungai Budi-Bumi Waras (2003-2004). Pada bulan April 2005 penulis menikah dengan Dedy Eko Rahmanto, S.TP.
Tahun 2007 penulis memperoleh gelar M.Si dari Program Studi Ilmu Pangan SPs IPB melalui beasiswa BPPS (2005-2007). Bulan Agustus 2007 penulis diterima sebagai mahasiswa S3 pada program studi yang sama melalui beasiswa I-MHERE Universitas Jember. Selama studi S3 penulis berperan aktif pada pelatihan maupun seminar ilmiah. Penulis telah mempresentasikan sebagian hasil penelitian disertasi di antaranya pada Seminar International Society for Nutraceuticals & Functional Food 11 – 15 Oktober 2010 di Bali dengan judul poster Effect of lactic acid bacteria fermentation on the plantain (var agung semeru) flour. Penulis mendapatkan juara dua pada Graduate Research Paper Competition (GRPC) tahun 2011 dengan judul Improving prebiotic properties of banana flour by modification process serta mempublikasikannya pada Seminar Nasional PATPI 15 – 17 September 2011 di Manado. Penulis juga mempresentasikan makalah dengan judul Low glycemic index modified plantain flour as functional food pada International Food Conference 28 – 29 Oktober di Surabaya. Bagian dari disertasi telah dipublikasikan pada jurnal ilmiah terakreditasi yaitu Jurnal Ilmu Dasar FMIPA Universitas Jember SK Dikti No. 65a-DIKTI/Kep/2008 Vol. 12 No. 2 Tahun 2011 (210-225) dengan judul
Phenotypic and genotypic identification of lactic acid bacteria isolated from spontaneous fermentation of unripe var agung semeru banana (Musa paradisiaca formatypica). Artikel dengan judul Physicochemical characteristic of modified banana flour by fermentation and autoclaving-cooling process sedang diajukan ke jurnal Agritech FTP-UGM dan Artikel dengan judul Prebiotic properties of modified banana flour as functional foods akan diajukan ke jurnal Internasional
xxi
Halaman
DAFTAR TABEL ... xxiii
DAFTAR GAMBAR ... xxv
DAFTAR LAMPIRAN ... xxvii
1. PENDAHULUAN ... 1
Latar belakang ... 1
Tujuan Penelitian ... 4
Manfaat Penelitian ... 4
Hipotesis Penelitian ... 5
Lingkup Penelitian ... 5
Daftar Pustaka ... 8
2. TINJAUAN PUSTAKA ... 9
Tepung Pisang ... 9
Karakteristik dan Modifikasi Pati ... 12
Indeks Glikemik ... 15
Pati Resisten dan Sifat Prebiotik ... 16
Fermentasi Bakteri Asam Laktat pada Pangan Berpati ... 21
Identifikasi Bakteri Asam Laktat ... 22
Daftar Pustaka ... 24
3. METODOLOGI PENELITIAN SECARA UMUM ... 29
Waktu dan Tempat ... 29
Alat dan Bahan ... 29
Tahap Penelitian ... 30
Daftar Pustaka ... 34
4. KARAKTERISTIK FISIKOKIMIA TEPUNG PISANG TERMODI- FIKASI SECARA FERMENTASI SPONTAN DAN SIKLUS PEMANASAN BERTEKANAN-PENDINGINAN ... 35
Abstrak ... 35 LAKTAT YANG BERPERAN SELAMA FERMENTASI SPONTAN PISANG VAR AGUNG SEMERU (Musa paradisiaca formatypica) ... 53
Abstrak ... 53
Abstract ... 53
xxii
Hasil dan Pembahasan ... 58 Kesimpulan ... 65 Daftar Pustaka ... 65
6. PENINGKATAN PATI RESISTEN TEPUNG PISANG MELALUI
FERMENTASI Lactobacillus salivarius FSnh1 DAN DUA SIKLUS
PEMANASAN BERTEKANAN-PENDINGINAN ... 69
Abstrak ... 69
Abstract ... 69 Pendahuluan ... 70 Bahan dan Metode ... 71 Hasil dan Pembahasan ... 73 Kesimpulan ... 76 Daftar Pustaka ... 77
7. EVALUASI SIFAT PREBIOTIK DAN INDEKS GLIKEMIK TEPUNG PISANG MODIFIKASI ... 79
Abstrak ... 79
Abstract ... 79 Pendahuluan ... 80 Bahan dan Metode ... 81 Hasil dan Pembahasan ... 86 Kesimpulan ... 94 Daftar Pustaka ... 95
8. PEMBAHASAN UMUM ... 99
9. KESIMPULAN DAN SARAN ... 109
DAFTAR PUSTAKA ... 111
xxiii
Halaman Tabel 2.1 Sifat fisik dan kimia tepung pisang dari beberapa varietas ... 10 Tabel 2.2 Komposisi pati, gula dan suhu gelatinisasi berdasarkan tingkat
kematangan warna kulit pisang ... 11 Tabel 2.3 Efek fisiologis dan klaim manfaat kesehatan oligosakarida ... 20 Tabel 4.1 Nilai pH, konsentrasi asam laktat dan tekstur pisang selama
fermentasi spontan pisang ... 41 Tabel 4.2 Pengaruh fermentasi spontan dan siklus pemanasan bertekanan- pendinginan terhadap komposisi proksimat tepung pisang .. ... 42 Tabel 4.3 Pengaruh fermentasi spontan dan siklus pemanasan bertekanan-
pendinginan terhadap komposisi pati dan daya cerna tepung pisang.. ... 43 Tabel 5.1 Karakteristik bakteri asam laktat yang diisolasi dari fermentasi
spontan pisang var agung semeru.. ... 59 Tabel 5.2 Pola fermentasi isolat BAL FSnh1dan isolat BAL FSnhA pada
Kit API 50CHL ... 60 Tabel 5.3 Hasil analisis sekuen DNA Pengkode 16SrRNA dari isolat BAL
FSnh1 dan FSnhA menggunakan program BLAST-N.. ... 62 Tabel 6.1 Populasi bakteri asam laktat, nilai pH dan konsentrasi asam laktat selama fermentasi pisang.. ... 73 Tabel 6.2 Pengaruh lama fermentasi pisang oleh L. salivarius FSnh1dan
dua siklus siklus pemanasan bertekanan-pendinginan terhadap komposisi pati tepung pisang modifikasi .. ... 75 Tabel 7.1 Populasi beberapa jenis mikroflora dan konsentrasi asam lemak
rantai pendek selama fermentasi pati resisten oleh kultur fekal…... 91 Tabel 7.2 Indeks glikemik tepung pisang pada beberapa perlakuan
xxv
Halaman
Gambar 1.1 Diagram alir penelitian ... 7 Gambar 2.1 Pisang Agung (Musa paradisiaca formatypica) ... 11 Gambar 2.2 Struktur granula pati yang menunjukkan daerah amorf dan semi
kristal ... 14 Gambar 2.3 Granula pati (A) sebelum dan (B) sesudah difermentasi oleh
bakteri asam laktat amilolitik ... 22 Gambar 2.4 Granula pati pada media MRS cair (A) sebelum diotoklaf , (B)
sesudah diotoklaf, (C) setelah difermentasi oleh L. amylophilus
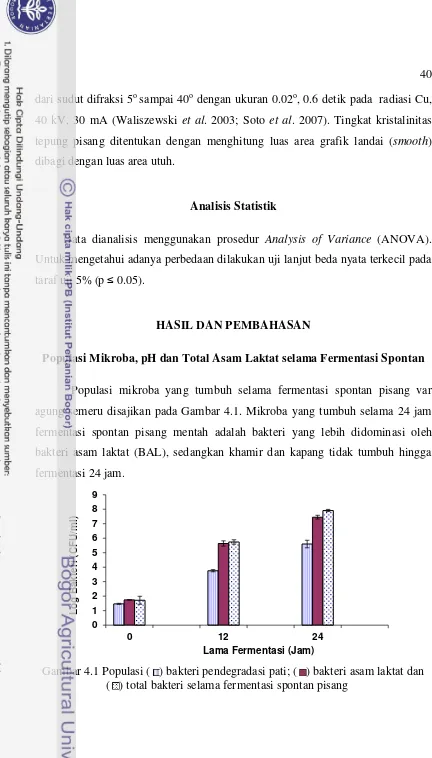
GV6 (C) ... 22 Gambar 4.1 Populasi ( )bakteri pendegradasi pati; ( ) bakteri asam laktat
dan ( ) total bakteri selama fermentasi spontan pisang ... 40 Gambar 4.2 Pengaruh fermentasi spontan dan dua siklus siklus pemanasan
bertekanan- pendinginan terhadap sifat birefringence granula pati pisang. (A) alami (kontrol); (B) fermentasi; (C) dua siklus siklus pemanasan bertekanan-pendinginan; (D) fermentasi spontan dengan dua siklus siklus pemanasan
bertekanan-pendinginan pada perbesaran 400× ... 45 Gambar 4.3 Pengaruh fermentasi spontan terhadap intensitas difraksi tepung
pisang; ( ) kontrol, ( ) fermentasi ... 47 Gambar 4.4 Pengaruh dua siklus siklus pemanasan bertekanan-pendinginan
terdapat intensitas difraksi tepung pisang. ( ) tanpa fermentasi
spontan, ( ) dengan fermenasi spontan ... 48 Gambar 5.1 Hasil elektroforesis agarosa 1% dan amplifikasi DNA pengkode
16S rRNA dengan PCR M = marka DNA 1kb DNAladder.
a = BAL FSnh1; b = BAL FSnhA ... 61 Gambar 5.2 Pohon filogenetik berdasarkan sekuen DNA pengkode 16S rRNA
isolat BAL FSnh1 dan BAL FSnhA yang dibandingkan dengan sekuen DNA pengkode 16S rRNA bakteri asam laktat genbank
dalam satu famili Lactobacillacea ... 63 Gambar 5.3 Pohon filogenetik berdasarkan sekuen DNA pengkode 16S rRNA
dari isolat BAL FSnh1 dan BAL FSnhA yang dibandingkan dengan sekuen DNA pengkode 16S rRNA bakteri asam laktat
genbank dalam satu genus Lactobacillus ... 64 Gambar 6.1 Pengaruh lama fermentasi oleh Lactobacillus salivarius FSnh1
xxvi
(A) kontrol (tanpa modifikasi), (B) dengan fermentasi spontan, (C) dua siklus pemanasan bertekanan-pendinginan, (D) fermentasi spontan dengan dua siklus pemanasan
bertekanan-pendinginan ... ... 87 Gambar 7.2 Persentase Pertumbuhan ( ) L. acidophilus (MRSB), ( ) EPEC
(MRSB), ( ) L. acidophilus (MRSB basis + RS), ( ) EPEC (MRSB basis + RS), (PBP = pemanasan
bertekanan-pendinginan) ... 88 Gambar 7.3 Persentase Pertumbuhan ( ) L. acidophilus (MRSB), ( ) S.
Typhimurium (MRSB), ( ) L. acidophilus (MRSB basis + RS), ( ) S. Typhimurium (MRSB basis + RS), (PBP = pemanasan
bertekanan-pendinginan) ... 88 Gambar 7.4 Indeks prebiotik pati resisten tepung pisang; ( ) tanpa
modifikasi (kontrol), ( ) modifikasi secara fermentasi yang dikombinasikan dengan dua siklus pemanasan bertekanan-
xxvii
1.
PENDAHULUAN
Latar Belakang
Pangan fungsional adalah makanan atau bahan pangan yang dapat memberikan manfaat tambahan bagi kesehatan manusia di samping fungsi gizi dasar pangan tersebut. Komponen pangan fungsional dapat berupa suatu komponen gizi makro yang memiliki efek fisiologis spesifik seperti pati resisten dan asam lemak n-3 atau suatu komponen gizi mikro yang asupannya lebih besar dari asupan harian atau tidak bernilai gizi seperti mikroorganisme dan komponen bioaktif tanaman (Roberfroid 2000).
Permintaan konsumen terhadap pangan fungsional semakin meningkat dan mendapat respon positif dari produsen. Produsen makanan dan minuman fungsional di Amerika Serikat, Eropa dan Asia Pasifik dapat meraih keuntungan $72.3 milyar dan diperkirakan akan terus meningkat 5.7% per tahun dalam tahun 2007 sampai 2012. Pangan fungsional tersebut didominasi oleh pangan probiotik dan prebiotik (Datamonitor Newswire 2008).
Pada tahun 2007 jumlah produk makanan prebiotik yang terlisensi lebih dari 400 macam serta lebih dari 20 perusahaan memproduksi oligosakarida dan serat sebagai prebiotik. Produsen prebiotik di Eropa telah meraih keuntungan sebesar €87 juta dan naik menjadi €179.7 juta pada tahun 2010 (FAO 2007). Peningkatan pertumbuhan pasar prebiotik dalam lima hingga sepuluh tahun mendatang membuka kesempatan bagi ahli teknologi pangan untuk senantiasa mengkaji sumber-sumber prebiotik baru yang dapat bermanfaat bagi status kesehatan manusia.
keseimbangan mikroflora intestinal (Gibson & Roberfroid 1995; Roberfroid 2007). Mikroflora yang digolongkan sebagai probiotik diantaranya adalah yang memproduksi asam laktat yaitu laktobasili dan bifidobakteria serta bakteri jenis lain. Beberapa produk komersial prebiotik adalah FOS (fruktooligo sakarida), inulin, GOS (galaktooligosakarida), laktulosa dan laktitol. Bahan-bahan lain yang memenuhi kriteria prebiotik yaitu xilosa, soya (rafinosa dan stakiosa kedelai), serta manosa (Collin & Gibson 1999; FAO 2007).
Sekelompok bahan yang dalam beberapa tahun terakhir memperoleh perhatian besar karena berpotensi sebagai kandidat prebiotik adalah pati resisten (resistant starch/RS). Menurut Sajilata et al. (2006) RS adalah pati yang tidak dapat dicerna oleh enzim pencernaan pada usus kecil sehingga dapat mencapai usus besar dan dapat difermentasi oleh mikroflora pada usus besar. Kondisi demikian akan mampu menstimulasi pertumbuhan mikroflora probiotik seperti
Lactobacillus spp dan Bifidobacteria dan dapat menurunkan pH usus sehingga mencegah pertumbuhan bakteri patogen seperti Escherichia coli, Salmonella sp,
Staphylococcus aureus, dan Clostridium sp. RS pati jagung yang dimodifikasi secara kimiawi terbukti dapat menstimulasi pertumbuhan bifidobakteria.
Kadar amilosa yang tinggi berperan dalam meningkatkan kadar RS3 yang terbentuk akibat proses retrogradasi. Tepung jagung dengan kadar amilosa 25% memiliki kadar RS sebesar 3g/100g berat kering, sedangkan tepung jagung dengan kadar amilosa 70% memiliki kadar RS sebesar 20 g /100 g berat kering (Sajilata et al. 2006).
Lumajang 2009). Pisang var agung semeru adalah jenis pisang plantain yang perlu diolah terlebih dahulu sebelum dikonsumsi seperti dikukus, digoreng, dikolak, diolah menjadi keripik dan bentuk olahan lainnya. Pisang plantain
memiliki kandungan pati lebih banyak sehingga baik untuk dikembangkan menjadi tepung pisang sebagai bentuk produk setengah jadi (intermediet product) yang dapat diolah lebih lanjut menjadi produk pangan.
Pengembangan tepung pisang menjadi pangan fungsional dapat berdasarkan pertimbangan kandungan RS yang banyak ditemukan pada buah pisang mentah. RS tersebut merupakan RS tipe II (RS2) yang bersifat mudah rusak selama proses pengolahan terutama pemanasan basah yang menyebabkan gelatinisasi pati sehingga struktur granula pati rusak dan kehilangan sifat resistennya terhadap enzim pencernaan. Oleh karena itu dilakukan upaya meningkatkan kandungan RS yang bersifat lebih stabil selama proses pengolahan. Soto et al. (2004) melaporkan bahwa proses retrogradasi berulang pada pati pisang mampu meningkatkan pati resisten tipe III (RS3) yang bersifat lebih stabil selama pengolahan.
Kadar RS pada pisang cavendish (Musa cavendishii) yang sudah tua tetapi belum matanghanya sebesar 1.51 ± 0.1% berat kering. Kadar RS akan meningkat sebesar dua kali jika pati pisang dihidrolisis dengan asam dan meningkat sebesar 7 – 10 kali jika pati pisang dipanaskan pada suhu 121 oC selama 1 jam (Saguilan et al. 2005). Soto et al. (2007) juga melaporkan bahwa kadar RS meningkat pada pati pisang jenis plantain dengan adanya pemanasan yang dikombinasi dengan pendinginan (retrogradasi). Menurut Sajilata et al. (2006) pati teretrogradasi (RS3) cenderung sulit dicerna dalam saluran pencernaan, akan tetapi dapat digunakan oleh mikroflora usus sebagai sumber energi bagi pertumbuhannya.
diikuti pendinginan (4 oC, 24 jam). Modifikasi tersebut mampu meningkatkan kadar RS3 sekitar 2 kali lipat (Jenie et al. 2009).
Fermentasi secara spontan selama 24 jam berperan dalam meningkatkan kandungan RS3 tepung pisang, akan tetapi memiliki kelemahan di antaranya ialah jenis mikroba yang tumbuh dapat bervariasi dan sangat tergantung pada kondisi dan lingkungan sehingga sulit dikontrol. Populasi awal BAL yang rendah dapat menyebabkan bakteri pembusuk serta bakteri patogen tumbuh lebih cepat mendahului pertumbuhan BAL (Antara et al. 2002). Penggunaan kultur starter indigenus dari bahan aslinya lebih memudahkan dalam mengendalikan proses fermentasi serta memberikan hasil fermentasi yang lebih baik dan sesuai dengan karakteristik produk yang diinginkan (Antara 2010). Oleh karena itu perlu dilakukan penelitian lebih lanjut untuk menerapkan proses fermentasi terkendali dengan menggunakan isolat BAL asal fermentasi spontan pisang dengan lama fermentasi yang optimal. Dalam upaya peningkatan RS tepung pisang, dilakukan modifikasi proses yang meliputi kombinasi fermentasi pisang dengan siklus pemanasan bertekanan-pendinginan (retrogradasi).
Tujuan Penelitian
Tujuan umum penelitian adalah meningkatkan sifat prebiotik tepung pisang dengan indeks glikemik rendah melalui proses fermentasi dan siklus pemanasan bertekanan-pendinginan. Secara rinci tujuan dari penelitian adalah:
1. Mengevaluasi karakteristik fisikokimia tepung pisang modifikasi secara fermentasi spontan dan siklus pemanasan bertekanan-pendinginan dalam upaya meningkatkan pati resisten (RS) sebagai kandidat prebiotik terhadap.
2. Mengidentifikasi fenotip dan genotip isolat bakteri asam laktat yang berperan selama fermentasi spontan pisang var agung semeru (Musa paradisiaca
formatypica).
4. Mengevaluasi sifat prebiotik isolat RS dan indeks glikemik tepung pisang modifikasi.
Manfaat Penelitian
Tepung pisang modifikasi yang dihasilkan dapat dikembangkan sebagai pangan fungsional yang memiliki sifat prebiotik yang baik dan nilai indeks glikemik rendah sehingga dapat dimanfaatkan bagi kesehatan pencernaan manusia dan sebagai pangan diet. Galur bakteri asam laktat yang diperoleh dari fermentasi spontan pisang var agung semeru dapat digunakan sebagai stater dalam pembuatan tepung pisang modifikasi kaya RS sehingga proses fermentasi pisang lebih terkendali.
Hipotesis Penelitian
Hipotesis penelitian adalah:
1. Proses modifikasi secara fermentasi spontan dan siklus pemanasan bertekanan-pendinginan mempengaruhi karakteristik fisikokimia tepung pisang.
2. Bakteri asam laktat yang berperan selama fermentasi spontan memiliki karakteristik fenotip dan genotip yang spesifik.
3. Fermentasi pisang menggunakan starter isolat BAL indigenus dapat mempersingkat waktu fermentasi dalam pembuatan tepung pisang modifikasi kaya RS.
4. Proses modifikasi secara fermentasi dan siklus pemanasan bertekanan-pendinginan mampu meningkatkan sifat prebiotik tepung pisang.
Lingkup Penelitian
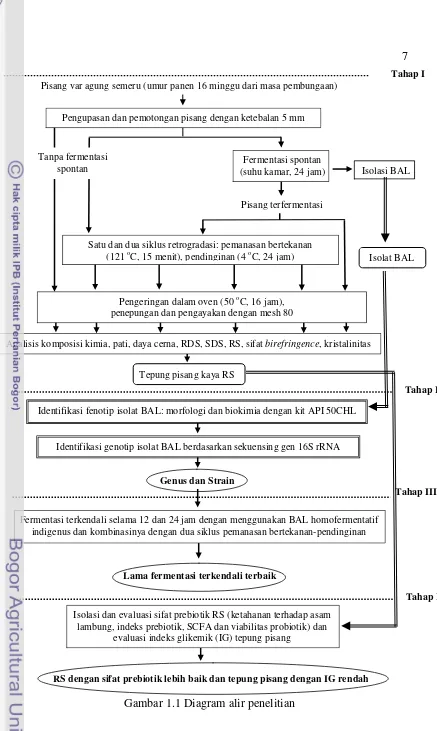
Gambar 1.1Diagram alir penelitian
Tahap I
Pengeringan dalam oven (50 oC, 16 jam), penepungan dan pengayakan dengan mesh 80
Analisis komposisi kimia, pati, daya cerna, RDS, SDS, RS, sifat birefringence, kristalinitas
Identifikasi fenotip isolat BAL: morfologi dan biokimia dengan kit API 50CHL
Tahap III
Satu dan dua siklus retrogradasi: pemanasan bertekanan (121 oC, 15 menit), pendinginan (4 oC, 24 jam)
Fermentasi terkendali selama 12 dan 24 jam dengan menggunakan BAL homofermentatif indigenus dan kombinasinya dengan dua siklus pemanasan bertekanan-pendinginan
Pisang var agung semeru (umur panen 16 minggu dari masa pembungaan)
Pengupasan dan pemotongan pisang dengan ketebalan 5 mm
Fermentasi spontan (suhu kamar, 24 jam)
Pisang terfermentasi
Tahap IV Tahap II
Lama fermentasi terkendali terbaik
Identifikasi genotip isolat BAL berdasarkan sekuensing gen 16S rRNA
Isolasi dan evaluasi sifat prebiotik RS (ketahanan terhadap asam lambung, indeks prebiotik, SCFA dan viabilitas probiotik) dan
evaluasi indeks glikemik (IG) tepung pisang
Isolasi BAL Tanpa fermentasi
spontan
Isolat BAL
Genus dan Strain
Tepung pisang kaya RS
DAFTAR PUSTAKA
Antara NS. 2010. Peran bakteri asam laktat strain lokal untuk memperbaiki mutu dan keamanan produk pangan lokal. [Orasi Ilmiah]. Fakultas Teknologi Pertanian Universitas Udayana.
Antara NS, Sujaya IN, Yokota A, Asano K, Aryanta WR, Tomita F. 2002. Identification and succession of lactic acid bacteria during fermentation of ‘urutan’, a Balinese indigenous fermented sausage. World J Microbiol & Biotechnol. 18: 255–262, 2002.
[DN] Datamonitor Newswire. 2008. Functional food. http://www.google.com. [13 Nov 2008].
[FAO] Food and Agricultural Organization. 2007. Technical meeting on preobitics. http://www.fao.org/ag/agn/agns/files/Prebiotics_Tech_Meeting_ Report.pdf. Accessed on 22 November 2008.
Gibson GR, Roberfroid M. 1995. Dietary modulating of the human colonic microbiota: introducting the concept of prebiotics. J Nutr. 125: 1401-1412. http:/www.ajcn.org/cgi/content/full/69/5/1052S [12 Nov 2008].
Jenie BSL, Widowati S, Nurjannah S. 2009. Pengembangan Produk Tepung Pisang Dengan IG Rendah dan Sifat Prebiotik Sebagai Bahan Pangan Fungsional. Laporan Akhir Hibah Kompetitif Penelitian Sesuai Prioritas Nasional Batch II. LPPM, IPB
[RPJMD] Kabupaten Lumajang. 2009. Rencana Pembangunan Jangka Kabupaten Menengah Daerah Kabupaten Lumajang 2010 - 2014.
Roberfroid M. 2000. Concepts and strategy of functional food science: the european perspective. Am J Clin Nutr. 71(suppl):1660S–4S.
Roberfroid M. 2007. Prebiotics: The Concept Revisited. The Journal of Nutrition Effect of Probiotics and Prebiotics.137:830S-837S [01 Juni 2008]
Saguilan AA, Flores-Huicochea E, Tovar J, Garcia-Suarez F, Guiterrez-Meraz F, Bello-Perez LA. 2005. Resistant starch rich-powders prepared by autoclaving of native and lintnerized banana starch: partial characterization.
J Starch/Starke. 57:405-412.
Sajilata MG, Rekha SS, Puspha RK. 2006. Resistant starch a review. J Comprehensive Rev in Food Sci and Food Safety. 5: 1-17.
Soto RAG, Acevedo EA, Feria JS, Villalobos RR, Bello-Perez LA. 2004. Resistant starch made from banana starch by autoclaving and debranching. J Starch/Stärke. 56: 495–499.
2. TINJAUAN PUSTAKA
Tepung Pisang
Tepung pisang merupakan produk antara yang cukup prospektif dalam pengembangan sumber pangan lokal. Tepung pisang dibuat dari buah pisang yang masih mentah yang sudah cukup tua namun belum masak. Manfaat pengolahan pisang menjadi tepung antara lain yaitu lebih tahan disimpan, lebih mudah dalam pengemasan dan pengangkutan, lebih praktis untuk diversifikasi produk olahan, mampu memberikan nilai tambah buah pisang, mampu meningkatkan nilai gizi buah melalui proses fortifikasi selama pengolahan, dan menciptakan peluang usaha untuk pengembangan agroindustri pedesaan.
Tepung pisang banyak dimanfaatkan sebagai campuran pada pembuatan roti, cake, kue kering, campuran tepung terigu, dan campuran makanan bayi. Pada dasarnya semua jenis buah pisang mentah dapat diolah menjadi tepung, tapi warna tepung yang dihasilkan beragam, karena dipengaruhi oleh tingkat ketuaan buah, jenis buah dan cara pengolahan. Buah pisang kepok mempunyai warna tepung yang paling baik yaitu putih.
Ada beberapa jenis pisang yang warnanya berbeda-beda, tetapi hampir semua yang dijual di pasar atau supermarket berwarna kuning ketika sudah matang dengan bentuk mayoritas melengkung. Deptan (2009) mengklasifikasikan jenis pisang menjadi empat yaitu:
1. Pisang yang dimakan dalam keadaan segar setelah buahnya masak yaitu
Musa paradisiaca var.sapienium, M. nana L atau M. cavendishii, dan M. sinensis. Misalnya pisang ambon, susu, raja, barangan dan mas.
2. Pisang yang dimakan setelah diolah yaitu M. paradisiaca formatypicaatau
M. paradisiaca normalis. Misalnya pisang nangka, tanduk dan kepok. 3. Pisang berbiji yaitu M. brachycarpa yang di Indonesia dimanfaatkan
daunnya.Misalnya pisang batu dan kluthuk.
Komposisi kimia buah pisang bervariasi tergantung pada varietasnya. Pada umumnya daging buah pisang mengandung energi, protein, lemak, berbagai vitamin serta mineral seperti yang ditunjukkan pada Tabel 2.1.
Tabel 2.1 Komposisi zat gizi pisang per 100 g buah segar (Aurore et al. 2009)
Senyawa Komposisi
Energi (Kkal) 91.00
Air (g) 63.00
Karbohidrat (g) 24.30
Protein (g) 0.80
Lemak (g) 0.10
Ca (mg) 7.00
Mg (mg) 33.00
P (mg) 35.00
Fe (mg) 0.50
Cu (mg) 0.16
Β karoten ekuivalen (µg) 0.03 – 1.20
Vitamin B1 (mg) 0.05
Vitamin B2 (mg) 0.05
Vitamin B6 (mg) 0.07
Vitamin C (mg) 20.00
Asam pantotenat (mg) 0.37
Asam folat (mg) 0.16
Serotonin (mg) 45.00
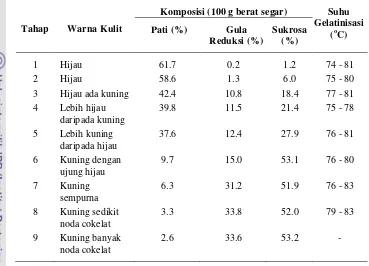
Tabel 2.2 Komposisi pati, gula dan suhu gelatinisasi berdasarkan tingkat kematangan warna kulit pisang (Zhang et al. 2005)
Tahap Warna Kulit
Komposisi (100 g berat segar) Suhu Gelatinisasi
Pisang var agung semeru (Musa paradisiaca formatypica) merupakan jenis pisang yang biasa dimakan setelah diolah misalnya dikukus, digoreng, direbus, diolah menjadi kolak, kripik, dan lain sebagainya. Pisang var agung semeru mempunyai ukuran buah yang besar dan bentuk yang menyerupai tanduk sehingga di beberapa tempat menyebutnya sebagai pisang tanduk. Pisang ini mempunyai panjang dapat lebih dari 10 cm. Setiap tandan hanya mempunyai satu hingga tiga sisir.
Pisang var agung semeru banyak dibudidayakan di Kabupaten Lumajang Jawa Timur. Berdasarkan data dari Unit Pelaksana Teknis Dinas (UPTD) Pertanian Kecamatan Senduro Kabupaten Lumajang, populasi pisang var agung semeru di awal tahun 2004 mencapai 323 hektar dari luas total Kecamatan Senduro yaitu 52,000 hektar. Produksi pisang var agung semeru di Kecamatan Senduro mencapai 33 ribu ton per tahun, dan tahun 2004 meningkat menjadi 37 ribu ton. Data UPT Dinas Pertanian Kecamatan Senduro tahun 2009 menunjukkan produksi pisang var agung semeru masih lebih tinggi yaitu mencapai 4,095,000 kg dibandingkan produksi pisang jenis lainnya seperti pisang mas (1,123,850 kg), pisang ambon (1,757,520 kg) dan pisang kepok (526,500 kg). Populasi tanaman pisang var agung semeru di desa Jambe Arum Kecamatan Pasrujambe mencapai 612.5 hektar atau sekitar 1.53 juta pohon pisang, dengan asumsi rata-rata 2,500 pohon pisang per hektar (RPJMD Kabupaten Lumajang 2009).
Karakteristik dan Modifikasi Pati
Pati merupakan homoglikan yang terdiri atas satu jenis unit D-glukosa yang dihubungkan oleh ikatan glukosida. Unit glukosa pati membentuk dua jenis polimer yaitu amilosa dan amilopektin. Pada umumnya pati mengandung 15-30% amilosa, 70-85% amilopektin dan 5-10% bahan lain seperti lipid, protein dan mineral (Emanual 2005).
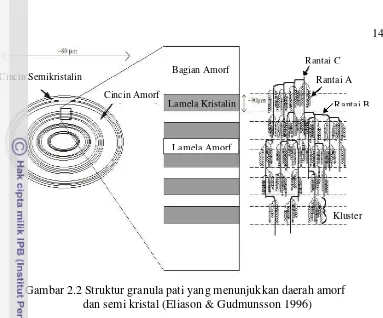
Bentuk pati secara alami berupa butiran-butiran kecil yang sering disebut granula. Bentuk dan ukuran granula setiap jenis pati mempunyai karakteristik tersendiri sehingga dapat digunakan untuk identifikasi. Selain ukuran granula, karakteristik lain dari pati adalah bentuk dan keseragaman granula, lokasi hilum serta permukaan granula pati. Dalam keadaan murni granula pati berwarna putih, mengkilat, tidak berbau dan tidak berasa. Secara mikroskopik terlihat bahwa granula pati dibentuk oleh molekul-molekul yang membentuk lapisan-lapisan tipis dan tersusun secara terpusat.
Gambar 2.2 Struktur granula pati yang menunjukkan daerah amorf dan semi kristal (Eliason & Gudmunsson 1996)
Pati pisang memiliki ukuran diameter rata-rata 24.31 µm untuk pati yang tidak dimasak dan 59-66 µm untuk pati yang dimasak (Nunez-Santiago et al. 2004). Tingkat kematangan pisang juga mempengaruhi komposisi kimia daging buah seperti kadar pati, gula reduksi, sukrosa dan suhu gelatinisasi (Zhang et al. 2005).
Lawal (2004) menjelaskan cara modifikasi pati di antaranya melalui proses hidrolisis asam, oksidasi, ikat silang (cross-linking atau cross bonding), subtitusi dan gelatinisasi. Pati termodifikasi asam dibuat dengan menghidrolisis pati menggunakan asam di bawah suhu gelatinisasi (sekitar 52 oC) sehingga terjadi pemotongan ikatan α-1,4-glukosidik dari amilosa dan α-1,6-D-glukosidik dari amilopektin membentuk pati dengan ukuran molekul lebih rendah. Pati teroksidasi dibuat dengan penambahan natrium hipoklorid dan banyak digunakan pada pembuatan kertas. Pati pregelatinisasi dibuat dengan cara memanaskan pati sehingga terjadi gelatinisasi dan banyak digunakan pada pembuatan saus, pasta dan jelly. Pati yang diperoleh secara kimia dari reaksi ikat silang bahan kimia seperti boraks, epikloridin, fosfor oksiklorida dan lain sebagainya. Pati jenis ini banyak digunakan sebagai pie filling (pada pengalengan, gravy dan saus),
Cincin Semikristalin
Cincin Amorf
Bagian Amorf
Lamela Kristalin
Lamela Amorf
Rantai C Rantai A
Rantai B
pembuatan makanan bayi dan salad dressing. Pati termodifikasi oleh hidrolisis α– amilase menyebabkan terjadinya pemotongan ikatan glukosidik yang berlangsung dalam dua tahap yaitu serangan enzim secara acak akan mendegradasi pati menjadi maltosa dan maltotriosa dan hidrolisis oligosakarida menjadi glukosa dan maltosa.
Indeks Glikemik
Pati dalam pangan dapat diklasifikasikan berdasarkan nilai glikemik dan ketahanannya terhadap enzim pencernaan. Pati glikemik didegradasi dalam saluran pencernaan oleh enzim amilase. Pati glikemik dapat dikategorikan sebagai pati yang dicerna secara cepat (rapidly digestible starch/RDS) dan pati yang dicerna secara lambat (slowly digestible starch/SDS). RDS dicerna secara cepat dalam usus halus, sedangkan SDS dicerna lebih lambat daripada RDS. Kedua jenis pati ini dapat dicerna secara sempurna. Contoh pati yang dicerna secara cepat adalah pati yang mengalami gelatinisasi seperti kentang rebus. Pati yang dicerna secara lambat banyak terdapat pada bahan pangan seperti pasta (Croghan 2002).
Indeks glikemik (IG) adalah petunjuk tentang faali makanan terhadap kadar glukosa darah dan respon insulin. Pangan yang menaikkan kadar glukosa darah dengan cepat memiliki IG tinggi. Sebaliknya pangan yang menaikkan kadar glukosa darah dengan lambat memiliki IG rendah. Glukosa murni digunakan sebagai pembanding IG yang memiliki nilai IG 100. Beberapa faktor yang dapat mempengaruhi IG suatu pangan di antaranya adalah proses pengolahan, perbandingan kadar amilosa dan amilopektin, kadar gula dan daya osmotik, kandungan serat, kandungan lemak dan protein serta kandungan zat antigizi (Foster-Powell & Miller 1995).
diserap. Oleh karena itu semakin kecil ukuran partikel akan meningkatkan IG pangan. Pangan yang lebih banyak mengandung amilopektin akan lebih lama dicerna menjadi glukosa karena membutuhkan dua macam enzim untuk
mendegradasinya yaitu α-amilase dan α, 1-6 glukosidase sehingga lebih lambat dalam meningkatkan kadar IG (Astawan & Widowati 2011)
Pangan yang mengandung sukrosa dalam jumlah besar memiliki IG mendekati 60. Sukrosa dihidrolisis menjadi D-glukosa dan D-fruktosa oleh enzim sukrase atau invertase. Fruktosa diserap dan diambil langsung, selanjutnya diubah menjadi glukosa dalam hati. Oleh sebab itu respon gula darah terhadap fruktosa murni sangat kecil yaitu 23. Pangan yang mengandung lemak dan protein tinggi cenderung memperlambat laju pengosongan lambung sehingga pencernaan makanan dalam usus halus lebih lambat. Hal ini akan mempengaruhi peningkatan kadar glukosa darah sehingga IG pangan rendah. Kandungan zat antigizi dalam pangan seperti antitripsin dan fitat dalam kedelai dapat mempengaruhi pelepasan glukosa. Zat antigizi dapat membentuk kompleks dengan zat gizi seperti protein sehingga menurunkan daya cerna protein. Zat antigizi lain seperti senyawa polifenol dapat menghambat aktivitas enzim pencernaan sehingga menurunkan daya cerna pati (Fostel-Powell & Miller 1995; Astawan et al. 2006).
Pati Resisten dan Sifat Prebiotik
Pati ada yang dapat dicerna dan ada yang tidak dapat dicerna. Sejumlah besar pati yang tidak dapat dicerna masuk ke dalam usus besar dan merupakan substrat yang penting bagi mikroflora kolon. Pati tersebut bersifat resisten terhadap enzim pencernaan sehingga disebut pati resisten (resistant starch/RS).
tertentu dapat difermentasi oleh mikroflora usus besar yang akan menghasilkan asam lemak rantai pendek yaitu asam propionat, asam asetat dan asam butirat. Komponen tersebut dapat memberikan aspek fungsional bagi kesehatan tubuh. RS juga bisa memodifikasi lingkungan intrakolonik dan secara tepat mengubah fungsi toksikologi serta melindungi terhadap kanker colorectal dengan memperpendek waktu transit dan meningkatkan densitas kamba feses (Kumari & Thayumanavan 1997).
Ada empat macam pati resisten (RS) yang dikelompokkan berdasarkan asal terbentuknya. RS tipe I (RS1) adalah jenis pati yang terperangkap di dalam matriks sel, seperti pati pada polong-polongan. RS tipe II (RS2) adalah pati alami yang berupa granula pati, contohnya pati jagung yang kaya amilosa, pati kentang mentah dan pati pisang mentah. RS tipe III (RS3) adalah pati yang sudah mengalami retrogradasi karena pemanasan dan pendinginan berulang-ulang. RS tipe IV (RS4) adalah pati yang telah dimodifikasi secara kimia (Sajilata et al. 2006).
RS3 merupakan pati resisten yang paling sering digunakan sebagai bahan baku pangan fungsional. Pembentukan RS3 terjadi karena granula pati mengalami gelatinisasi. Granula rusak akibat proses pemanasan basah dan terjadi pelepasan amilosa dari granula ke dalam larutan. Pada saat pendinginan, rantai polimer terpisah sebagai ikatan ganda membelit (double helix) dan mengalami pembentukan kembali ke struktur awalnya secara perlahan membentuk struktur kompak yang distabilkan oleh ikatan hidrogen (Sajilata et al. 2006). Peristiwa ini dikenal dengan istilah retrogradasi (Lawal 2004). Amilosa teretrogradasi (RS3) bersifat lebih stabil terhadap panas, sangat kompleks dan tahan terhadap enzim amilase.
Bifidobacteria sehingga merupakan bahan bifidogenik yang sangat potensial (Hegar 2007).
Jumlah pati resisten pada pisang mentah lebih tinggi yaitu 4.7 gram dibandingkan kentang (3.2 gram) pada takaran penyajian yang sama (Mendosa 2008). Saguilan et al. (2005) menjelaskan bahwa pati pisang cavendish (Musa cavendishii) yang sudah tua tapi belum matang mengandung RS sebesar 1.51 ± 0.1 % berat kering. Kadar RS pada pisang ini akan meningkat sebesar dua kali jika dihidrolisis dengan HCl (litnerized starch), dan meningkat sebesar 7 – 10 kali jika dipanaskan pada suhu 121 oC selama 1 jam (autoclaved starch).
Beberapa riset telah melakukan modifikasi proses untuk meningkatkan kandungan RS tepung pisang. Kecepatan aliran udara selama proses pengeringan
chips (irisan) pisang mentah dapat mempengaruhi kandungan RS pada tepung pisang yang dihasilkan. Pengeringan pada suhu 55 oC dengan kecepatan udara 1.0 m/detik mampu meningkatkan kadar RS sekitar 40% (Tribess et al. 2009). Jenie et al. (2009) melaporkan bahwa modifikasi proses di tingkat pisang secara fermentasi spontan dan satu siklus pemanasan bertekanan-pendinginan mampu meningkatkan RS tepung pisang hingga dua kali lipat. Jenie et al. (2010) mengaplikasikan tepung pisang yang dihasilkan pada pembuatan produk pangan yaitu roti, cookies dan brownies.
Konsumsi bahan prebiotik secara signifikan dapat memodulasi komposisi mikrobiota kolon yang menyebabkan bifidobakteria lebih dominan dalam kolon dan banyak ditemukan dalam tinja (Gibson & Roberfroid 1995). RS3 dari gandum, kentang dan kacang polong dapat menstimulasi pertumbuhan bifidobakteria yaitu Bifidobacteria pseudolongum KSI9, B. breve KN14 dan B. animalis KS20a1 (Wronkowska et al. 2006). RS4 dari jagung juga mampu menstimulasi pertumbuhan Bifidobacteria sp (Hegar 2007). Pati resisten termasuk molekul yang mempunyai panjang rantai (derajat polimerisasi) lebih pendek. Panjang rantai ini sangat berhubungan dengan kecepatan fermentasi. Roberfroid et al. (1997) menjelaskan bahwa derajat polimerisasi suatu oligosakarida dari bahan
polimerisasi (DP) kurang dari 10 seperti inulin akan difermentasi dua kali lebih cepat daripada molekul yang mempunyai DP lebih dari 10.
Pengaruh prebiotik terhadap pertumbuhan probiotik dinyatakan sebagai indeks prebiotik (IP) yang dihitung berdasarkan jumlah logaritmik pertumbuhan probiotik, dan mikroflora usus lainnya seperti klostridia dan bakteroides terhadap jumlah mikroba total. Analisis tersebut dilakukan dengan menumbuhkan mikroba dari feses manusia pada medium yang mengandung prebiotik uji (Manderson et al.2005; Roberfroid 2007).
Prebiotik didefinisikan sebagai suatu karbohidrat yang tidak dapat dicerna dan tidak dapat diserap tetapi dapat difermentasi secara selektif dan mempunyai fungsi regulasi terhadap probiotik dalam usus sehingga dapat memberikan efek kesehatan bagi manusia maupun hewan (Salminen & Wright 2004). Beberapa prasyarat yang dijabarkan FAO (2007) yaitu meliputi karakteristik prebiotik yang mendeskripsikan sumber asal prebiotik, tingkat kemurniannya, komposisi kimia dan strukturnya. Karakteristik lainnya adalah jumlah asupan yang dikonsumsi, sifat fungsional yang dapat menunjukkan bukti ilmiah antara efek fisiologis senyawa prebiotik dalam memodulasi mikrobiota pada daerah/organ spesifik. Kualifikasi prebiotik didasarkan pada penelitian dan metode ilmiah yang representatif dan akurat, serta keamanannya jika prebiotik tersebut dikonsumsi. Prebiotik juga tidak mengandung kontaminan dan impuritis, tidak mengubah mikrobiota yang menyebabkan dampak negatif bagi inang (manusia). Oleh karena itu senyawa prebiotik perlu mendapatkan status GRAS (Generally Recognized As Safe) atau setaranya. Beberapa klaim kesehatan dalam hubungannya dengan efek fisiologis dari senyawa prebiotik golongan oligosakarida dipaparkan pada Tabel 2.3.
dalam tahap pengembangan yaitu pektioligosakarida, laktosukrosa, gula alkohol, gluko-oligosakarida, levan, pati resisten, xilosakarida dan soy-oligosakarida.
Tabel 2.3 Efek fisiologis dan klaim manfaat kesehatan oligosakarida (Macfarlane & Cummings 1999)
Efek Fisiologis Faktor Kesehatan
- Menstimulasi metabolisme karbohidrat; meningkatkan massa sel bakteri, asam lemak rantai pendek
- Selektif terhadap bifidobakteria dan bakteri asam laktat dalam usus besar - Tidak terhidrolisis oleh mikroorganisme
mulut
- Tidak bersifat glikemik
- Menstimulasi dan bersifat tidak spesifik terhadap fungsi imun
- Memodulasi metabolisme karsinogen - Mengurangi sintesis LDL dan trigliserida
serum
- Meningkatkan penyerapan Mg dan Ca
Melalui asam lemak rantai pendek, menyediakan sumber energi untuk epitel kolon dan mengontrol diferensiasi serta menghindari sembelit
Meningkatkan retensi terhadap invasi patogen
Melindungi terhadap karies gigi Bermanfaat bagi penderita diabetes Mencegah infeksi
Bersifat antikarsinogen atau antikanker Mencegah penyakit jantung koroner Mencegah osteoporosis
Prebiotik dapat memodulasi pertumbuhan bakteri yang menguntungkan (probiotik). Peningkatan populasi probiotik memiliki manfaat diantaranya yaitu mencegah kanker karena dapat menghilangkan bahan prokarsinogen dari tubuh dan mengaktifkan sistem kekebalan tubuh. Probiotik tertentu seperti
Bifidobacterium infantis mengandung bahan aktif anti tumor pada dinding sel. Selain itu probiotik juga memproduksi berbagai enzim pencernaan (fosfatase, lisozim) dan vitamin (B1, B2, B6, asam folat, dan biotin) yang akan diserap di dalam usus halus dandimanfaatkan oleh tubuh serta memproduksi asam laktat dan asam asetat sehingga menyebabkan usus menjadi asam dan akhirnya menekan pertumbuhan bakteri patogen penyebab radang usus seperti Escherichia coli dan
Clostridium perfringens. Senyawa asam yang dihasilkan oleh bakteri asam laktat (probiotik) mampu menurunkan pH usus, meningkatkan absorpsi kalsium dan mengurangi penyerapan amonia dan amina sehingga dapat mencegah tekanan darah tinggi, kolesterol dan kanker yang disebabkan oleh nitrosamin.
menghasilkan antioksidan indigenus yaitu superoksid dismutase (Salminen & Wright 2004).
Fermentasi Bakteri Asam Laktat pada Pangan Berpati
Bakteri asam laktat (BAL) dapat ditemukan pada produk fermentasi spontan seperti ogi dari singkong (Reddy et al. 2008), asinan buah dan sayur (Kusumawaty et al. 2003), urutan yang merupakan sosis khas Bali dari daging babi (Antara et al. 2002). Selain itu BAL juga dapat diisolasi dari daging (Arief et al. 2011), susu (Sujaya et al. 2008), limbah kedelai (Malik et al. 2008), minuman serta buah yang busuk (Plessis et al. 2004). BAL tertentu seperti Lactobacillus plantarum, L. fermentum, L. manihotivorans, L. amylophillus, L. amylovorus, L. amilolyticus, Leuconostoc cellobiosus, L. acidophillus, Leuconostoc sp,
Streptococcus bovis dan S. macedonicus telah dilaporkan memiliki sifat amilolitik yaitu mampu menghasilkan enzim amilase untuk mendegradasi pati (Reddy et al. 2008).
BAL dapat memfermentasi pangan berkarbohidrat seperti jagung, kentang, ubi kayu, serealia dan lain sebagainya. Bakteri ini mampu menghasilkan enzim amilase dan asam yang dapat menghidrolisis sebagian pati seperti pati jagung, kentang, atau singkong dan beberapa substrat berpati lainnya (Reddy et al. 2008).
Spesies terbaru BAL amilolitik adalah Lactobacillus manihotivorans yang diisolasi dari pati asam ubi kayu (Reddy et al. 2008). Olympia et al. (1995) mengkarakteristik strain L. plantarum dari makanan khas Filipina yaitu burong isda yang terbuat dari ikan dan nasi. Strain amilolitik L. fermentum pertama kali diisolasi dari adonan pati jagung Benin (ogi dan mawe) (Agati et al. 1998). Sanni
et al. (2002) menemukan strain BAL amilolitik dan L. fermentum dari pangan terfermentasi khas Nigeria. BAL amilolitik menghasilkan enzim ekstraseluler yaitu amilase dan pululanase yang dapat menghidrolisis sebagian pati alami menjadi gula sederhana dan oligosakarida lain atau dekstrin (Sikorsi 2002).
Vishnu et al (2006) mengidentifikasi enzim amilase dan pululanase dari
0.18 U/g/min untuk pululanase yang difermentasi pada media dedak gandum (wheat bran). Aktivitas α-amilase dalam fermentasi pati oleh Streptococcus bovis
sebesar 1.41 U/ml lebih besar daripada fermentasi glukosa (0.06 U/ml) (Narita et al. 2004). Enzim α-amilase akan memotong karbohidrat pada ikatan endo-α 1,4 menghasilkan maltosa dan dekstrin. Pululanase akan memotong karbohidrat pada ikatan endo-α 1,6 menghasilkan dekstrin linier (Sikorsi et al. 2002).
Wronkowska et al. (2006) menjelaskan bahwa fermentasi pati gandum, pati kentang dan pati kacang polongoleh BA L selama 24 jam menunjukkan
perubahan mikrostruktur yaitu pembentukan struktur globular dan lamelar.
Sajilata et al.(2006) menjelaskan perubahan struktur pati dari kristalin menjadi lebih porus (amorf), meningkatkan kemampuan pelepasan amilosa serta menurunkan suhu gelatinisasi pati. Semakin banyak amilosa yang terlarut selama proses gelatinisasi maka akan semakin tinggi terjadinya retrogradasi pati selama proses pendinginan. Pati yang mengalami retrogradasi akan memiliki sifat lebih resisten terhadap enzim pencernaan. Pati ini sering disebut sebagai pati resisten tipe III (RS3). Perubahan yang terjadi pada granula pati akibat fermentasi BAL dapat diamati dengan menggunakan mikroskop elektron (Gambar 2.3 dan Gambar 2.4).
Gambar 2.3 Perubahan granula pati (A) sebelum dan (B) sesudah difermentasi oleh bakteri asam laktat amilolitik. Sumber: Wijbenga (2000)
Gambar 2.4 Granula pati pada media MRS cair (A) sebelum diotoklaf, (B) sesudah diotoklaf, (C) setelah difermentasi oleh
B
A
A
L. amylophilus GV6. Sumber: Vishnu et al. (2006)
Identifikasi Bakteri Asam Laktat
Bakteri asam laktat (BAL) merupakan kelompok bakteri yang dapat memproduksi asam laktat dengan cara memfermentasi karbohidrat. BAL yang menghasilkan dua molekul asam laktat dari fermentasi glukosa disebut bakteri asam laktat homofermentatif, sedangkan BAL yang menghasilkan satu molekul asam laktat dan satu molekul etanol serta satu molekul karbon dioksida disebut bakteri asam laktat heterofermentatif (Reddy et al. 2008).
Identifikasi BAL dapat dilakukan berdasarkan sifat fenotip dan genotip. Identifikasi fenotip hanya terbatas sampai tingkat spesies yang didasarkan pada hasil pengamatan morfologi seperti bentuk sel, tipe koloni dan pewarnaan Gram, uji fisiologis, metabolik (biokimia) atau kemotaksonomi. BAL merupakan bakteri dengan sifat katalase negatif sehingga pada uji katalase dengan hidrogen peroksida 30% tidak menghasilkan gelembung udara/gas. Identifikasi fenotip dengan pengujian fisiologis berdasarkan pola fermentasi BAL pada beberapa gula terkadang bias untuk beberapa spesies tertentu. Oleh karena itu identifikasi genotip perlu dilakukan untuk uji konfirmasi spesies BAL (Plessis et al. 2004).
Identifikasi genotip dilakukan dengan menggunakan metode molekuler di antaranya yaitu melalui penentuan urutan basa DNA pengkode 16S rRNA pada bakteri dengan metode Polymerase Chain Reactions (PCR)-sekuensing (Ammor
et al. 2005). Aplikasi molekuler DNA pengkode 16S rRNA untuk menganalisis keragaman molekuler suatu bakteri sangat sesuai karena gen ini terdapat pada semua mikroorganisme prokariot. Gen pengkode 16S rRNA bersifat stabil dalam sel bakteri daripada rRNA yang biasanya dapat terdegradasi dan hanya terdapat pada fase-fase tertentu saja (Guttel et al. 1994).
Ammor S, C Rachman, S Chaillou, H Prevost, X Dousset, M Zagorec, E Dufour, I Chevallier. 2005. Phenotypic and genotypic identification of lactic acid bacteria isolated from a small-scale facility producing traditional dry sausages. J Food Microbiol. 22: 373–382
Antara NS, IN Sujaya, A Yokota, K Asano, WR Aryanta, F Tomita. 2002. Identification and succession of lactic acid bacteria during fermentation of ‘urutan’, a Balinese indigenous fermented sausage. World J Microbiol & Biotechnol 18: 255–262, 2002.
Arief II, Jenie BSL, Asyawan M, Witarto AB. 2010. Efektivitas probiotik
Lactobacillus plantarum 2C12 dan Lactobacillus acidophilus 2B4 sebagai pencegah diare pada tikus percobaan. J Media Peternakan. 33 (3): 137-143.
Astawan M, Widowati S. 2011. Evaluation of nutrition and glycemic index of sweet potatoes and its appropriate processing to hypoglycemic foods.
Indonesian J Agricultural Science. Vol 12 (1)
Aurore G, Parfait B, Fahrasmane L. 2009. Bananas, raw materials for making processed food products. J Trends in Food Science & Technology. 20: 78 - 91
Collin MD, Gibson GR. 1999. Probiotics, prebiotics and synbiotics: approaches for modulating the microbial ecology of the gut. American J Clin Nutr. Vol. 69, No. 5. http://www.ajcn.org/cgi/ content/full/69/5/1052S [12 Okt 2008]. Croghan M. 2002. Resistant starch as a functional ingredient in food systems. J
Business Briefing: FoodTech. (Referece Section).
[Deptan] Departemen Pertanian. 2009. Produktivitas Pisang di Kabupaten Lumajang dalam Laporan Departemen Pertanian Kabupaten Lumajang Tahun 2008.
Eliasson AC, Gudmunsson M. 1996. Starch: physicochemical and functional properties aspects. In: Carbohydrates in Food (Edited by Eliasson A.C.), Marcel Dekker, Inc. New York. p 431-504.
Emanuel C. 2005. Pengaruh Fosforilasi dan Penambahan Asam Stearat Terhadap Karakteristik Film Edible Pati Sagu. [Tesis]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
[FAO] Food and Agricultural Organization. 2007. Technical meeting on preobitics. http://www.fao.org/ag/agn/agns/files/Prebiotics_Tech_Meeting_ Report.pdf. Accessed on 22 November 2008.
Foster-Powell K, Miller JB. 1995. International tables of glycemic index. Am J Clin Nutr. 62(suppl 2):S871–S890
Gibson GR, Roberfroid M. 1995. Dietary modulating of the human colonic microbiota: introducting the concept of prebiotics. J Nutr. 125: 1401-1412. http:/www.ajcn.org/cgi/content/full/69/5/1052S [12 Nov 2008].
Guttel RR, Larsen N, Woese CR. 1994. Lesson from evoluation rRNA, 16S rRNA and 23S rRNA strutsfores from a comparative perspective microbes. J Kes. 58: 10-26
Hegar B. 2007. Mikroflora saluran cerna pada kesehatan anak. J Dexa Media. 20 (1). Januari -Maret.
Jenie BSL, Widowati S, Nurjannah S. 2009. Pengembangan Produk Tepung Pisang Dengan IG Rendah dan Sifat Prebiotik Sebagai Bahan Pangan Fungsional. Laporan Akhir Hibah Kompetitif Penelitian Sesuai Prioritas Nasional Batch II. LPPM, IPB
Jenie BSL, Widowati S, Kusumaningrum HD. 2010. Pengembangan Produk Tepung Pisang Dengan IG Rendah dan Sifat Prebiotik Sebagai Bahan Pangan Fungsional. Laporan Akhir Hibah Kompetitif Penelitian Sesuai Prioritas Nasional Batch II. LPPM, IPB
Kumari SK, Thayumanavan B. 1997. Comparative study of resistant starch from minor millets on intestinal responese, blood glucose, serum cholesterol and triglycerides in rats. J Sci Food Agric. 75:296-302.
Kusumawati N, Jenie BSL, Siswasetyahadi, Hariyadi RD. 2003. Seleksi bakteri asam laktat indigenus sebagai galur probiotik dengan kemampuan menurunkan kolesterol. J Mikrobiologi Indonesia. 8 (2): 39-43
Lawal OS. 2004. Composition, physicochemical properties and retrogradation characteristics of native, oxidised, acetylated andacid-thinned new cocoyam (Xanthosoma sagittifolium) starch. J Food Chem. 87: 205–218
Macfarlane GT, Cummings JH. 1999. Probiotics and Prebiotics: can regulating the activities of intestinal bacteria benefit health? J Brit Med. April. 10.318(17189):999-1003.http://www.pubmedcentral.nih.gov/articlerender. fcgi?artid=1115424 [21 Agust 2008].
Malik A, Donna M. Ariestanti, Nurfachtiyani A, Yanuar A. 2008. Skrining gen glukosiltransferase (gtf) dari bakteri asam laktat penghasil eksopolisakarida.
J Makara Sains. 12 (1): 1-6
Manderson K, Pinar M, Tuhoy KM, Race WE, Otckiss AT, Widmer W, Yadhav MP, Gibson R, Rastall RS. 2005. In vitro determination of prebiotic properties of oligosaccharides derived from an orange juice manufacturing by-product stream. App and Env Microbiol. 71 (12): 8383-8389,
Mendosa. 2008. Revised international table of glycemic index (GI) and glycemic load (GL) values. www.mendosa.com [11Jan 2009].
Olympia M, Fukuda H, Ono H, Kaneko Y, Takano M. 1995. Characterization of starch-hydrolyzing lactic acid bacteria isolated from a fermented fish and rice food, “Burong Isda,” and its amylolytic enzyme. J Ferment Bioeng. 80:124–30.
Plessis HW, Dicks LMT, Pretorius IS, Lambrechts MG, Toit MD. 2004. Identification of lactic acid bacteria isolated from South African brandy base wines. Intern J Food Microbiol. 91: 19– 29
Reddy G, Altaf M, Naveena BJ, Venkateshwar M, Kumar EV. 2008. Amylolytic bacterial lactic acid fermentation — A review. J Elsevier- Biotechnol Adv. 26: 22–34.
[RPJMD] Kabupaten Lumajang. 2009. Rencana Pembangunan Jangka Kabupaten Menengah Daerah Kabupaten Lumajang 2010 - 2014.
Roberfroid M. 2007. Prebiotics: The Concept Revisited. The Journal of Nutrition Effect of Probiotics and Prebiotics.137:830S-837S [01 Juni 2008]
Saguilan AA, Flores-Huicochea E, Tovar J, Garcia-Suarez F, Guiterrez-Meraz F, Bello-Perez LA. 2005. Resistant starch rich-powders prepared by autoclaving of native and lintnerized banana starch: partial characterization.
J Starch/Starke. 57:405-412.
Sajilata MG, Rekha SS, Puspha RK. 2006. Resistant starch a review. J Comprehensive Rev in Food Sci and Food Safety. 5: 1-17.
Salminen S, Wright AV. 2004. Lactic Acid Bacteria: Microbiology and functional aspect. 2nd Edition. Revised and Expanded. New York: Marcell Dekker, Inc. Sanni A, Morlon-Guyot J, Guyot JP. 2002. New efficient amylase-producing
strains of Lactobacillus plantarum and L. fermentum isolated from different Nigerian traditional fermented foods. Int J Food Microbiol. 72:53–62.
Sikorsi ZE. 2002. Chemical and functional properties of food components. Ed ke-2. CRC Press
Sujaya N, Ramona Y, Widarini NP, Suariani NP, Dwipayanti NMU, Nociaanitri KA, Nursini NW. 2008. Isolasi dan karakterisasi bakteri asam laktat dari susu kuda sumbawa. J Veteriner. 9 (2): 52-59
Tribess TB, Hernandez-Uribe JP, Mendez-Montealvo MGC, Menezes EW, Bello-Perez LA, Tadini CC. 2009. Thermal properties and resistant starch content of green banana flour (Musa cavendishii) produced at different drying conditions. J Food Sci and Technol. 42:1022-1025.
Wijbenga DJ. 2000. Enzymatic modification of starch granules: peeling off versus porosity. TNO Nutr and Food Research. www.voeding.tno.nl [12 Febr 2009].
Wronkowska M, Smietana MS, Krupa U, Biedrzycka E. 2006. In vitro fermentation of new modified starch preparations—changes of microstructure and bacterial end-products. J Enzyme Microbial Technol. 40: 93–99
3. METODOLOGI PENELITIAN SECARA UMUM
Waktu dan Tempat
Penelitian ini dimulai bulan Maret 2009 sampai dengan bulan April 2011. Penelitian dilakukan di Laboratorium Departemen Ilmu dan Teknologi Pangan Fakultas Teknologi Pertanian IPB, Laboratorium South East Asian Food &
Agricultural Science & Technology (SEAFAST) Centre IPB serta Laboratorium pendukung seperti Labotarorium Balitnak Kementerian Pertanian dan Laboratorium Biorin-Bioteknologi IPB.
Alat dan Bahan
Alat-alat yang digunakan meliputi: otoklaf, mikroskop polarisasi (Olympus C-35AD-4 Japan), spektrofotometer, difraksi sinar X (Shimadzu XRD-7000 Maxima), gas kromatografi (Chrompack CP 9002 seri 946253), anoxomat,
anaerobic jar, sentrifuse, elektroforesis, UV transilluminator, alat PCR PTC 100 (MJ Research, Inc), inkubator, otoklaf, oven dan lain sebagainya.
Bahan baku yang digunakan adalah pisang var agung semeru (Musa paradisiaca formatypica) yang diperoleh dari Kecamatan Senduro Kabupaten Lumajang Propinsi Jawa Timur. Buah pisang tua mentah dipanen pada minggu ke 16 dari awal pembungaan.
Media dan bahan kimia yang digunakan antara lain: de Mann Rogosa Sharp Agar (MRSA) dan de Mann Rogosa SharpBroth (MRSB), Brain Heart Infusion
(BHI) agar, Thioglycollate agar,kristal violet, lugol, safranin, alkohol, NaCl, asam tartarat 10%, gliserol, NaOH, buffer fosfat, buffer TAE, Na2HPO4.2H2O,
NaHPO4, CaCl2.2H2O, MgCl2.6H2O, DNS, HCl, K2HPO4, MgSO4.7H2O,
CaCl2.2H2O, NaHCO3, bacto agar, L-cysteine HCl, garam bile, resazurin, vitamin
Tahap Penelitian
Penelitian meliputi empat tahap yaitu: 1) proses modifikasi secara fermentasi spontan serta kombinasinya dengan satu dan dua siklus pemanasan bertekanan yang dilanjutkan dengan pendinginan, 2) identifikasi fenotip dan genotip bakteri asam laktat (BAL) asal fermentasi spontan pisang var agung semeru, 3) proses modifikasi secara fermentasi oleh isolat BAL indigenus dan kombinasinya dengan siklus pemanasan bertekanan-pendinginan dalam pembuatan tepung pisang kaya RS, 4) isolasi pati resisten (RS) serta evaluasi sifat prebiotik dan nilai indeks glikemik tepung pisang.
Modifikasi Pembuatan Tepung Pisang secara Fermentasi Spontan dan Kombinasinya dengan Siklus Pemanasan Bertekanan-Pendinginan
Proses modifikasi dilakukan pada irisan pisang yaitu pisang diiris dengan ketebalan ± 5mm. Irisan pisang diberi perlakuan fermentasi spontan yang dilanjutkan dengan satu atau dua siklus retrogradasi yaitu pemanasan bertekanan yang diikuti pendinginan. Proses pemanasan bertekanan dilakukan dengan menggunakan otoklaf pada suhu 121 oC selama 15 menit yang kemudian didinginkan pada suhu 4 oC selama 24 jam. Tepung pisang kontrol disiapkan tanpa modifikasi yaitu pisang diiris dengan ketebalan ± 5mm dan dikeringkan pada suhu 50 oC selama 16 jam selanjutnya dihaluskan serta diayak dengan ayakan 80 mesh.
siklus pemanasan bertekanan-pendinginan maka irisan pisang terfermentasi diberi perlakuan satu dan dua siklus pemanasan bertekanan-pendinginan. Perlakuan diulang sebanyak dua kali dengan dua kali ulangan teknik sampling bahan baku di lahan budidaya pisang var agung semeru.
Keenam jenis tepung pisang yang dihasilkan selanjutnya dianalisis komposisi kimia yang meliputi proksimat (air, protein, lemak, mineral dan karbohidrat) dengan menggunakan metode AOAC (1999), kadar pati dan kadar amilosa (AACC 2000), pati cepat tercerna (rapidly digestible starch/RDS), pati lambat tercerna (slowly digestible starch/SDS), pati resisten (resistant starch/RS) (Englyst et al. 1992), serta daya cerna (AACC 2000). Karakterisasi fisik seperti bentuk granula pati dan tingkat kristalinitas tepung pisang juga dievaluasi pada tepung pisang kontrol dan tepung pisang modifikasi yang banyak mengandung pati resisten.
Identifikasi Fenotip dan Genotip Bakteri Asam Laktat Asal Fermentasi Spontan Pisang var Agung Semeru
Selama fermentasi spontan 24 jam diketahui populasi bakteri asam laktat (BAL) tumbuh dominan. Oleh karena itu dilakukan identifikasi BAL yang berperan selama fermentasi. Identifikasi fenotip berdasarkan sifat Gram positif, katalase negatif, bentuk sel kokus atau batang dengan bentuk/tipe koloni tertentu (Ammor et al. 2005). Selanjutnya isolat diinokulasikan dalam media MRS cair dan diinkubasi pada suhu 37 oC selama 24 jam untuk diidentifikasi lanjut atau disimpan dalam larutan gliserol (30% v/v) pada suhu -20 oC. Sifat fermentatif isolat BAL dianalisis dengan menggunakan kit API 50CHL.
Identifikasi genotip dilakukan dengan menggunakan metode PCR dan sekuensing DNA pengkode 16S rRNA. DNA isolat BAL diekstraksi dengan menggunakan metode Thompson et al. (1995).