UJIAN AKHIR SEMESTER
MENTRANSLATE JURNAL EKOLOGI HEWAN
DISUSUSUN OLEH : Aginta Putri Rehulina Keliat
(140410120037)
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM DEPARTEMEN BIOLOGI
Jurnal Ekologi Hewan
Jurnal Ekologi hewan 2010, 79, 888-896 doi: 10,1111 / j.1365-2656.2010.01686.x
Perbedaan ukuran variabilitas populasi antara
populasi dan spesies dari
keluarga Salmonidae
Ned A. Dochtermann1,2* Dan Mary M. Merak1,2
1Program Ekologi, Evolusi dan Biologi Konservasi, Universitas Nevada, Reno, NV, USA;dan 2DepartemenBiologi, University of Nevada, Reno, NV, USA
Ringkasan
1. Bagaimana ukuran populasi bervariasi dengan waktu adalah pertanyaan ekologis penting baik dengan implikasi praktis dan teoritis. Karena variabilitas ukuran populasi sesuai dengan pengoperasian mekanisme tergantung kepadatan dan kehadiran status yang stabil, banyak peneliti berusaha untuk melakukan perbandingan taksonomi yang luas variabilitas ukuran populasi.
2. Sebagian besar perbandingan ukuran variabilitas populasi menunjukkan kurangnya perbedaan taksonomi. Namun, perbandingan ini dapat menyamakan perbedaan dalam tingkat taksonomi dengan perbedaan antara tingkat taksonomi. Selanjutnya, sejauh mana perbedaan intraspesifik dapat mempengaruhi kesimpulan yang lebih luas secara umum belum diperkirakan dan sebagian besar telah diabaikan.
3. Untuk mengatasi ketidakpastian ini, kami menguji perbedaan intraspesifik dalam ukuran variabilitas populasi untuk total 131 populasi didistribusikan di antara sembilan spesies dari Salmonidae. Kita memperluas ini dibandingkan dengan tingkat interspesifik dengan cara mengembangkan tingkat perkiraan spesies pada populasi dengan banyak variabel.
4. Kami menggunakan pendekatan berlipat (resampling) untuk memperkirakan variasi intra dan interspesifik dalam ukuran variabilitas populasi. Kami menemukan perbedaan intraspesifik yang signifikan dalam bagaimana ukuran populasi bervariasi dengan waktu dalam semua enam spesies salmon di mana itu bisa diuji serta perbedaan interspesifik jelas. Selanjutnya, meskipun variasi antarspesies yang signifikan, mayoritas variasi ini adalah pada tingkat antar jenis. Akhirnya, kami menemukan bahwa langkah-langkah klasik dan yang baru-baru ini dikembangkan dari populasi variabilitas menyebabkan kesimpulan sesuai. 5. Adanya perbedaan signifikan pada intraspesifik di semua spesies diperiksa
menunjukkan bahwa kemampuan untuk mendeteksi pola taksonomi yang luas dalam bagaimana ukuran populasi berubah dari waktu ke waktu mungkin akan terbatas jika varians tidak benar dipartisi antara dan di dalam tingkat taksonomi.
Pengantar
Sejauh mana variasi ukuran populasi dengan waktu terikat pada beberapa pertanyaan kunci dalam ekologi (Connell & Sousa 1983). Misalnya, variabilitas ukuran populasi secara intrinsik terkait dengan peran proses tergantung kepadatan vs. tidak tergantung kepadatan dan apakah proses populasi tersebut stabil. (Mei 1973; Connell & Sousa 1983; Peterson 1984; Hanski 1990). Variabilitas ukuran populasi juga sering dikaitkan dengan pengoperasian mekanisme tergantung kepadatan (Hanski 1990) dan dikenal untuk mempengaruhi kemungkinan kepunahan untuk suatu populasi (Pimm, Jones and Diamond 1988; Bengtsson & Milbrink 1995; Vucetich et al. 2000.; Fagan et al 2001.; Inchausti & Halley 2003).
Untuk alasan ini, perhatian telah diarahkan terhadap menggambarkan pola taksonomi yang luas dalam bagaimana ukuran populasi bervariasi secara temporal. Perbedaan antara taksa dari bunga ekologi dan evolusi karena mereka mungkin mencerminkan perbedaan peran relatif proses tergantung kepadatan dan tidak tergantung kepadatan (Connell & Sousa 1983). Misalnya, Connell & Sousa (1983) menunjukkan bahwa tidak ada bukti perbedaan taksonomi umum dalam variasi ukuran populasi pada berbagai taksa (misalnya tanaman, serangga, parasit dan burung) menunjukkan berbagai jarak variabilitasnya. Baru-baru ini, Inchausti & Halley (2001, 2002) tidak menemukan bukti untuk perbedaan taksonomi dalam variabilitas temporal, menunjukkan bahwa mekanisme tergantung kepadatan tidak spesifik untuk taksa tertentu; meskipun Reed & Hobbs (2004) menyatakan bahwa burung mungkin menunjukkan populasi agak lebih stabil. Stabilitas yang besar ini dapat menunjukkan bahwa mekanisme yang tergantung kepadatan lebih banyak terjadi pada burung. Sayangnya, membandingkan ukuran variabilitas populasi antara taksa sulit karena potensi bias dalam indeks variabilitas, korelasi dengan kepadatan, skala sampling dan keandalan data (McArdle, Gaston & Lawton 1990).
Perbedaan fluktuasi populasi dari waktu ke waktu antara populasi residen serta kalangan congener adalah penting karena dua alasan: Pertama, banyak dari pola taksonomi yang luas atau tidak ada daripadanya- telah dijelaskan berdasarkan hanya beberapa populasi suatu spesies. Dengan demikian, tinggi atau rendah stochasticity lingkungan yang dialami oleh populasi tunggal mungkin akan bias perkiraannya pada variabilitas populasi pada spesies-spesies. Perbedaan dalam taksa memungkinkan adanya perbedaan antara taksa. Kedua, karena hubungan antara variabilitas ukuran populasi dan risiko kepunahan, mengidentifikasi perbedaan antara populasi dari spesies yang sama dapat membantu untuk menjelaskan hal yang mendasari mekanisme kausal pada variabilitas, baik secara alami atau berasal dari antropogenik, yang pada gilirannya dapat digunakan untuk menemukan strategi yang pas (Marsh, 2001).
perbedaan populasi yang umum untuk spesies salmonid. Jika variabilitas ukuran populasi berbeda antara populasi dari spesies yang sama, itu akan menunjukkan bahwa upaya untuk menguji pola taksonomi mungkin sengaja menyamakan perbedaan intra dan interspesifik.
Selanjutnya, kami memperkirakan proporsi variasi menjelaskan tingkat interspesifik vs tingkat intraspesifik. Akhirnya, kami melakukan uji demonstratif perbedaan antarspesies antara ikan Lahontan dan populasi Steelhead, yang bentuk anadromous dari ikan rainbow
(Oncorhynchus mykiss), yang terletak di Sungai Keogh, Kanada. Steelhead adalah yang paling terkait erat dengan salmonid tersedia di GPDD. Karena studi mengenai populasi ikan Lahontan di sini adalah terbatas pada daerah terpencil kecil dengan tingkat tinggi variabilitas lingkungan, kami memprediksikan bahwa populasi ikan akan menunjukkan ukuran sementara variabel yang lebih besar kemampuan daripada populasi Steelhead anadromous.
Bahan dan metode IKAN LAHONTAN
POPULASI PENELITIAN
Ikan Lahontan saat ini terdaftar sebagai salah satu hewan terancam punah oleh United States Endangered Species Act dengan sangat erat kisaran terfragmentasi relatif terhadap pendistribusian yang meluas (Coffin & Cowan 1995; Dunham & Vinyard 1997). Secara historis populasi dari ikan Lahontan hidup di beberapa sungai jaringan besar namun sebagian besar populasi termasuk dalam penelitian ini kini terisolasi dalam satu aliran atau jalur (Dunham et al., 1999; Neville, Dunham & Merak 2006). Neville dkk. (2006) menunjukkan bahwa baik bentang darat dan proses metapopulasi memainkan peran dalam ketekunan populasi jangka populasi ikan Lahontan dalam sistem aliran yang saling berhubungan yang besar. Penciptaan kembali seperti jaringan sungai besar tidak mungkin, dengan demikian memahami potensi resiko kepunahan untuk populasi yang berbeda dan pemahaman dalam membedakan antara faktor-faktor ekstrinsik dan intrinsik yang mempengaruhi variabilitas temporal yang akan memungkinkan arah yang tepat upaya konservasi.
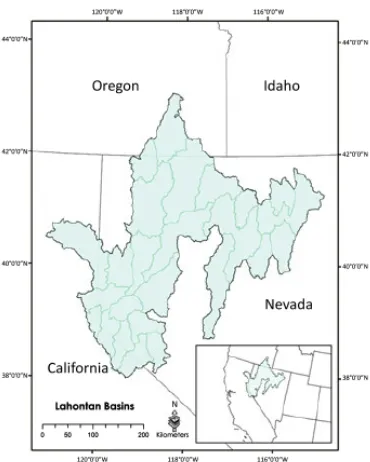
Kami mempelajari dinamika populasi dari lima populasi ikan Lahontan dari tahun 1996 sampai 2000. Populasi ini terletak di Mohawk, Tierney, India, Abel dan 3-Mile sungai. Sungai ini pertama atau kedua sungai dari Sungai Humboldt dan terletak di bagian timur cekungan hidrografi serta ikan Lahontan lebih besar (Gambar. 1). Populasi dalam sungai ini terisolasi ke hulu mencapai ke hilir mengganggu keterkaitan aliran. Hambatan mencakup diversi air, suhu air tidak cocok dan hambatan buatan manusia dirancang untuk meminimalkan hibridisasi dan persaingan ancaman yang ditimbulkan oleh salmon non-pribumi (Dunham et al 1999;. Peacock & Kirchoff 2004).
POPULASI SAMPEL
Gambar. 1. cekungan hidrografi ikan Lahontan, yang mana ikan Lahontan ikan pembunuh endemik, meliputi empat negara bagian di Amerika barat Amerika.
pergantian yang sempurna dari omset populasi ikan Lahontan (Ray, Peacock & Dunham, 2007). Ukuran populasi sebuah sungai yang diperkirakan menggunakan multiple-pass dengan pendekatan deplesi sampling pada tujuh lokasi sepanjang jangkauan diduduki dalam sungai. Lokasi pengambilan sampel adalah 25 m dan dipisahkan oleh 300 m (sesuai protokol standar sampel untuk spesies, misalnya Dunham & Vinyard 1996, 1997; Dunham, Cade & Terrell 2002). Sebelum pengambilan sampel, situs yang diblokir di bawah dan ujung hulu dengan pukat jaring untuk mencegah ikan melarikan diri selama sampling. Suatu situs telah disampling menggunakan unit ransel elektro-ikan sementara yang bergerak hilir ke hulu antara blok-jaring, yang dinyatakan 'lulus sampel'. Unit elektro-fishing mengejutkan individu ikan sementara menakjubkan mereka dengan sedikit efek pada kelangsungan hidup langsung (Mitton & McDonald 1994), meskipun beberapa mempengaruhi pada pertumbuhan yang telah diusulkan (Dwyer, Shepard & White 2001). Passes berlanjut sampai tidak ada ikan baru yang terdeteksi. Massa (g) dan panjang standar (Mm) dicatat untuk setiap sampel individu ikan.
Jumlah ikan Lahontan dalam setiap situs diperkirakan menggunakan MicroFish (Van Deventer & Platts 1989), sebuah pendekatan maksimum kemungkinan bahwa memperkirakan jumlah ikan ini berdasarkan pada jumlah ikan yang ditangkap selama setiap pengambilan yang lulus sampel. Jika semua sampel individual tertangkap selama pass pertama, kami menggunakan total jumlah yang ditangkap sebagai perkiraan kami. Ukuran populasi dari sungai itu kemudian diperkirakan dengan ekstrapolasi kepadatan ikan di lokasi di seluruh panjang yang diduduki sungai (Dunham et al. 1999).
Juga variabilitas ukuran populasi salmon lainnya (Lihat di bawah) tidak dapat dibagi menjadi komponen ruang dan waktu. Dengan demikian, partisi varians spasial untuk ikan Lahontan akan membuat perbandingan dengan spesies lain yang tidak informatif.
SALMONIDAE LAINNYA
Kita memperluas analisis kita tentang pola perbedaan variabilitas ukuran populasi dengan spesies lain dalam keluarga Salmonidae berdasarkan pada populasi tersedia di GPDD.
DATA GPDD
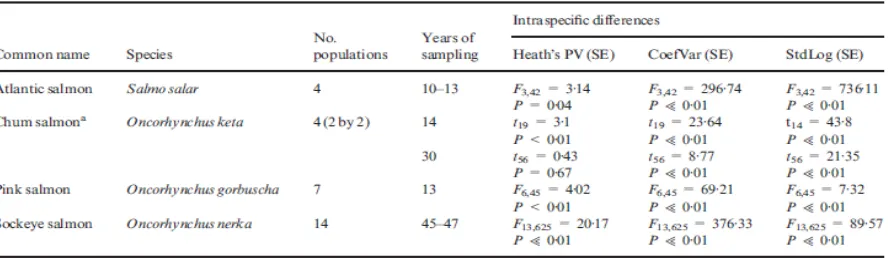
Kami menggunakan populasi di GPDD yang perkiraan ukuran populasi ditentukan tahunan dan berdasarkan jumlah individu. Populasi yang dimasukkan dalam analisis kami tanpa pertimbangan untuk ‘kualitas data'. Pendekatan ini konsisten dengan kegunaan lain data GPDD (Misalnya Fagan et al 2001;. Inchausti & Halley 2002, 2003) dan memaksa asumsi bahwa kualitas data tidak terarah mempengaruhi populasi perkiraan ukuran. Kami tidak, bagaimanapun, termasuk set data dengan kesenjangan dalam tahun ke tahun memperkirakan lebih dari satu periode sampling. Tambahan lagi untuk lima populasi ikan Lahontan, ini mengakibatkan dimasukkannya tambahan 126 populasi yang didistribusikan di antara delapan spesies (Lampiran S1, informasi pendukung). Sedangkan, jumlah populasi yang termasuk dan panjang pengambilan sampel bervariasi dengan spesies (Tabel 1).
aHeath PV, bCoefVar atau cStdLog berbeda secara signifikan antara populasi; aataubatau
kemudianc Hasil Anova disajikan. * Ini intraspesifik dif ferences tercampur dengan efek
panjang time series.
ANALISIS DATA
Perbedaan intraspecific dalam ukuran variabilitas populasi
paling menjadi indeks umum variabilitas populasi tetapi keduanya sensitif pelanggaran asumsi yang mendasarinya seperti mendasari distribusi ukuran populasi (McArdle & Gaston 1995; Heath 2006). Jadi kita juga menghitung indeks nonparametrik temporal variabilitas yang diusulkan oleh Heath (2006). Heath variabilitas populasi (PV Heath) menghitung variabilitas sebagai proporsional rata perbedaan antara semua kelimpahan diukur. Simulasi menunjukkan bahwa PV Heath adalah kurang sensitif terhadap distribusi non-normal dan lebih akurat memperkirakan variabilitas jangka panjang dari jangka pendek set data (Heath 2006). Kami menggunakan semua tiga indeks untuk memungkinkan menjadi lebih besar dan baik dan karena Heath PV mungkin tidak langsung tertandingi pada perkiraan yang diterbitkan sebelumnya. Kami tidak menghitung spektral memerah karena menujukan pertanyaan yang berbeda dari yang kita meminta disini.
Untuk membandingkan poin perkiraan antara kelompok dalam hal ini ukuran variabilitas populasi indeks antara populasi, kami menggunakan pendekatan 'menghapus satu jackknife' (Roff 2006). Prosedur jackknife memproduksi sebuah 'nilai semu' untuk setiap tahun sampling untuk setiap populasi dan indeks. Sebagai contoh, jika populasi telahdi sampling pada 20 kesempatan, 20 nilai semu akan dihitung untuk setiap indeks yang diperkirakan. Nilai semu dihitung dengan terlebih dahulu memperkirakan suatu parameter yang partikular di mana n adalah ukuran sampel. Nilai dihapus kemudian kembali ke kumpulan data dan pengamatan selanjutnya akan dihapus untuk menghitung detik psuedovalue, S1 :
dimana n adalah ukuran sampel. Nilai dihapus kemudian kembali ke kumpulan data dan pengamatan selanjutnya akan dihapus untuk menghitung detik nilai semu (S2). Ini menghapus, estimasi dan mengganti prosedur adalah populasi tunggal. Untuk spesies mana hanya populasi tunggal terus berlanjutnya 'n' kali, sekali untuk setiap titik data.
Rata-rata nilai semu pada sampel, estimasi berlipat untuk indeks tertentu, nilai ini kemudian diestimasi oleh jackknife parameter of interest . Demikian juga, standar error dari nilai semu adalah standar error dari estimasi berlipat (Roff 2006). Selain menghitung standar eror untuk perkiraan, nilai semu juga dapat digunakan untuk pengujian hipotesis (Roff 2006). Hal ini sangat berguna untuk memperkirakan variabilitas ukuran populasi karena memungkinkan perbandingan kuantitatif dari satu populasi yang lain.
analisis pada subset dari data dengan waktu yang sebanding dengan timbangan pengambilan sampel (lihat di bawah).
Untuk menentukan apakah keragaman hadir, ukuran variabilitas antara populasi spesies yang pengambilan sampel adalah dilakukan pada frame skala waktu yang sama baik menggunakan analisis variansi atau t -tests. Misalnya, tujuh populasi Pink Salmon (On corhynchus gorbuscha) masing-masing sampel selama periode 13-tahun. Untuk populasi ini, tiga analisis varians dilakukan dengan menggunakan perkiraan berlipat untuk setiap indeks dengan populasi sebagai independen faktor. Pendekatan umum ini diulangi untuk Atlantic salmon (Salmo salar), Chum salmon (Oncorhynchus keta) dan Sockeye sal mon
(Oncorhynchusnerka) Populasi spesifik dan panjang sampling dilaporkan dalam Tabel 2.
Tabel 2. Ketika variabilitas ukuran populasi dihitung untuk populasi dari berbagai panjang, perbedaan antar jenis yang digabungkan dengan kecenderungan yang diamati untuk ukuran variabilitas populasi meningkat dengan panjang pengamatan (Tabel 1 perbedaan berlabel '*'). Kami dievaluasi perbedaan intraspecific dalam populasi ini berdasarkan himpunan bagian dari data panjang dibandingkan menggunakan salah satu arah ANOVA (dengan penduduk sebagai faktor tetap) atau dua arah t –tests
a Untuk Chum Salmon, kami membandingkan dua pasang populasi: satu pasang telah
dipantau selama 14 tahun (baris atas) dan pasangan lainnya selama 30 (baris bawah).
Perbedaan antarspesies dalam ukuran variabilitas populasi
Untuk menghasilkan perkiraan tingkat spesies variabilitas ukuran populasi dan memungkinkan perbandingan antarspesies, kami menggunakan beberapa
perbedaan pendekatan. Pendekatan yang harus digunakan berbeda berdasarkan pada data yang tersedia.
variabel bawah). Kami menggunakan pendekatan ini untuk Steelhead trout (O. mykiss), Coho salmon (Oncorhynchus kisutch) dan Chinook salmon (Oncorhynchus tshawytscha) (Tabel 1).
Beberapa populasi, panjang pengambilan sampel yang sama. Untuk spesies di mana beberapa populasi sampel masing-masing sampel untuk jangka waktu yang sama, pertama kita menghitung indeks diperkirakan berlipat setiap populasi. Indeks rata-rata antara populasi dan standar eror yang kemudian digunakan sebagai perkiraan tingkat spesies variabilitas ukuran populasi. Kami menggunakan pendekatan ini untuk ikan Lahontan (masing-masing lima populasi dipantau selama 5 tahun) dan sungai ikan forel (Salvelinus fontinalis) (Tabel 1). Selama enam dari tujuh populasi sungai ikan forel tersedia di GPDD dijadikan sampel selama 7 tahun. Populasi terakhir adalah sampel selama 6 tahun. Karena sebagian besar populasi dijadikan sampel untuk rentang waktu yang sama dan satu sampel untuk hanya satu tahun kurang, kita mengabaikan perbedaan ini untuk spesies. Karena pendekatan ini untuk menghitung perkiraan spesies juga memproduksi perkiraan varians, perbedaan antarspesies dapat diuji.
Beberapa populasi variabel panjang pengambilan sampel. Untuk spesies di mana beberapa populasi dijadikan sampel tetapi dengan panjang bervariasi dari periode sampling, spesies menentukan tingkat perkiraan lebih rumit karena variabilitas ukuran populasi meningkat dengan jumlah pengulangan pengambilan sampel (Lawton 1988; Pimm & Redfearn 1988; Arin~o & Pimm 1995; Inchausti & Halley 2002). Untuk mengatasi masalah ini, kita menghitung perkiraan berlipat untuk setiap indeks untuk masing-masing populasi. Kami kemudian memperkiraan populasi ini mundur terhadap panjang rangkaian waktu. Mencegah setiap kemunduran model dan standar eror kemudian digunakan perkiraan tingkat spesies variabilitas ukuran populasi dengan efek panjang rangkaian waktu dihapus. Kami menggunakan pendekatan ini untuk Atlantic salmon (S. salar), Chum salmon (O. keta), Pink salmon (O. gorbuscha) dan Sockeye salmon (O. nerka). Di antara spesies ini, panjang ukuran populasi waktu diperkirakan berkisar antara 7 sampai 111 tahun (Tabel 1).
Kami menganggap pendekatan ini menjadi yang paling dipakai untuk interspesifik perbandingan sebagai spesies yang berbeda mungkin menunjukkan waktu yang berbeda: hubungan variabilitas. Dengan demikian, perkiraan mencegah memungkinkan perbandingan tanpa hubungan pengganggu mungkin panjang pengambilan sampel. Memang, perkiraan ini mungkin bisa dianggap sebagai variabilitas dasar untuk spesies. Namun, penting untuk dicatat bahwa perkiraan yang dihasilkan dengan cara ini dapat dibandingkan dengan masing-masing lainnya, mereka tidak dapat langsung dibandingkan dengan perkiraan dari kedua metode 'populasi tunggal' atau 'beberapa populasi, sampling yang sama panjang 'dijelaskan di atas.
yang tersisa karena antar keragaman. Untuk setiap dua spesies, standar pendekatan statistik membuat filogenetik yang tepat asumsinya, jadi kami juga menunjukkan pengujian perbedaan antarspesies menggunakan estimasi variabilitas ukuran populasi untuk ikan Lahontan dan ikan Steelhead.
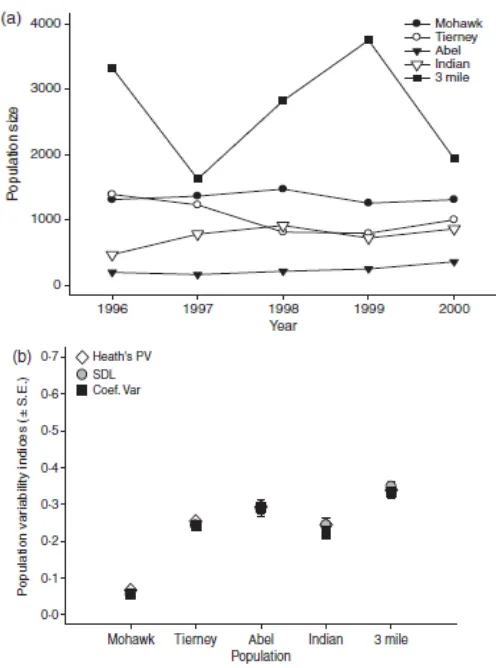
Gambar. 2. (a) perkiraan ukuran pada lima populasi ikan Lahontan diperkirakan selama 5 tahun. Semua populasi yang terletak di bagian timur bagian dari Lahontan Basin dan terisolasi dari hulu tanpa imigrasi di antara mereka. Ukuran populasi diperkirakan menggunakan MicroFish (Van Deventer & Platts 1989). (B) ukuran populasi perkiraan variabilitas untuk lima populasi ikan Lahontan. Perkiraan variabilitas yang dihasilkan untuk ikan Lahontan berdasarkan 5 tahun pengambilan sampel dan pendekatan berlipat.
Hasil
IKAN LAHONTAN
Kami membandingkan kelimpahan antara populasi ikan Lahontan menggunakan populasi sebagai faktor acak dan tahun sebagai kovariat. Kelimpahan yang dilog-transformasi setelah itu mereka terdistribusi normal (Shapiro-Wilk W = 0·941; P = 0·16). Kami memperkirakan bahwa rata-rata lebih dari 6000 total ikan Lahontan hadir antara populasi gabungan di Mohawk, Tierney, Abel, Sungai India dan 3-Mile selama setiap tahun sampling (Tidak termasuk muda-of-the-tahun;. Gambar 2a). Namun, ikan Lahontan tidak terdistribusi secara merata di antara populasi. Populasi berbeda dalam kelimpahan (F4,19 = 55·7, P
tahun (F1,19= 0·52, P = 0·48). Secara umum 3 Mile Creek memiliki ukuran populasi terbesar
sementara di Abel dan Sungai India memiliki populasi terendah (Gambar. 2a).
Ikan Lahontan juga berbeda dalam variabilitas temporal. Situs berbeda secara signifikan dalam StdLog (F4,20= 20·21; P> 0Æ01; Gambar. 2b); CoefVar (F4,20= 19·59;
P«0·01; Gambar. 2b) tetapi tidak ketika variabilitas temporal diestimasi dengan menggunakan Heath PV (F4,20= 1·66; P = 0·20; Gambar. 2b). Pasca-hoc (Tukey) perbandingan dilakukan
antara populasi untuk ketiga pengukuran. 3-Mile dipamerkan variabilitasnya yang lebih besar dari semua sungai berdasarkan StdLog dan semua anak sungai kecuali Abel berdasarkan CoefVar. Mohawk Creek dipamerkan variabilitas rendah dari semua sungai lainnya untuk kedua StdLog dan CoefVar. Indian Creek dipamerkan variabilitas rendah dari Abel berdasarkan CoefVar.
PERBEDAAN INTRASPESIFIK SALMONID
Seperti halnya untuk ikan Lahontan, semua spesies di mana yang bisa diuji menunjukkan perbedaan intraspesifik di ukuran variabilitas populasi (Tabel 1). Untuk Atlantic, Chum, Pink dan Sockeye salmon, perbedaan populasi yang sebenarnya adalah berpotensi digabungkan dengan efek dari perbedaan panjang pengambilan sampel. Untuk menguji perbedaan intraspesifk di spesies ini, kami menggunakan subset dari data yang sebanding dengan panjang pengambilan sampel. Analisis subset ini menunjukkan kehadiran keragaman untuk semua empat spesies (Tabel 2). Namun, untuk Chum salmon, PV Heath tidak menunjukkan bahwa dua populasi sampel lebih dari 30 tahun berbeda dari satu sama lain (Tabel 2).
PERBEDAAN INTERSPESIFIK
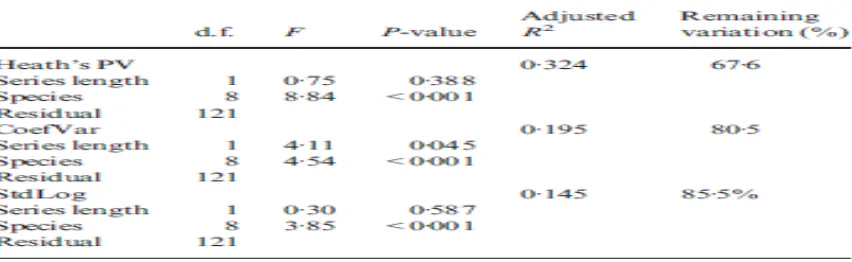
Model linear mengidentifikasi perbedaan spesies yang signifikan untuk ketiga indeks variabilitas ukuran populasi (Tabel 3). Meskipun signifikansi statistik, spesies dan waktu hanya menyumbang 22% dari variasi rata-rata. Jadi untuk tiga indeks 78% dari variasi dalam ukuran variabilitas populasi tetap pada tingkat antar jenis.
Untuk menunjukkan kemampuan menggunakan perkiraan berlipat untuk melakukan perbandingan antarspesies, kita diuji untuk kehadiran
perbedaan antara ikan Lahontan dan ikan Steelhead. Perkiraan populasi jackknifed dihitung untuk ikan Lahontan vs spesies jackknifed perkiraan untuk ikan Steelhead menggunakan t-test [H0= 0·507 (Heath PV), 0Æ643 (CoefVar), 0Æ661 (StdLog)]. Populasi ikan Lahontan
dipamerkan secara signifikan kurang kemampuan variabilitas dalam kelimpahan daripada ikan Steelhead (PV Heath: t4= -5·78, P = 0·004; CoefVar: t4= -8·43, P = 0·001; StdLog: t4=
-8·42, P = 0·001).
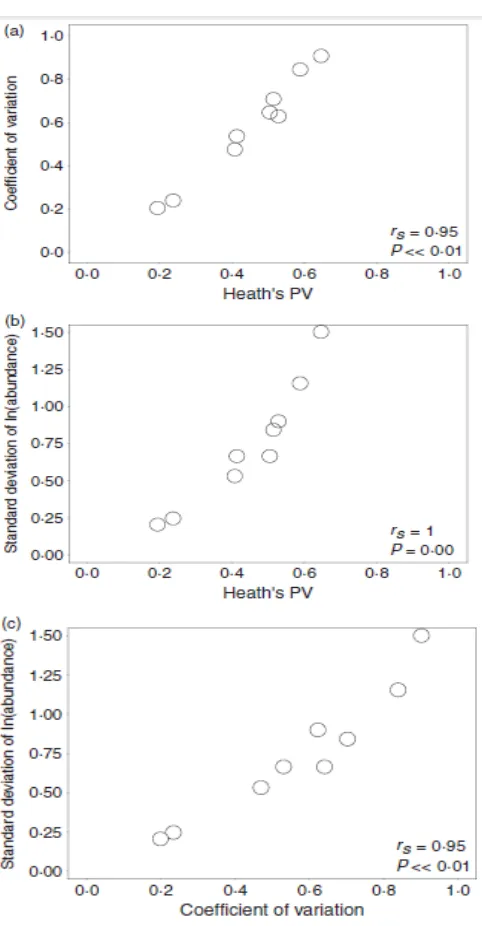
Secara keseluruhan, tiga indeks yang berbeda dari ukuran populasi variabel kemampuan sesuai dalam bagaimana peringkat spesies yang berbeda berkaitan dengan variabilitas ukuran populasi. Spearman peringkat korelasi berkisar antara 0·95 dan 1 untuk tiga indeks yang berbeda (Gbr. 3) menunjukkan bahwa indeks mengukur populasi dengan sifat dinamis yang sama.
DISKUSI
Kebanyakan penelitian variabilitas ukuran populasi antara populasi dari satu spesies telah berfokus pada perbedaan konteks kerentanan populasi tertentu untuk kepunahan (Schoener & Spiller 1992; Lima, Marquet & Jaksic 1998; Vucetich et al 2000.; Marsh 2001; Inchausti & Halley 2003; Reed & Hobbs 2004; Legendre et al. 2008). Ini berbeda dari tujuan kita di sini yang adalah untuk mengukur besarnya dan prevalensi perbedaan intrapopulation. Hasil penelitian kami menunjukkan bahwa ada perbedaan yang signifikan dalam kelimpahan berfluktuasi antara populasi spesies salmonid yang berbeda.
Perbedaan ukuran variabilitas populasi antara spesies sulit untuk ditentukan karena metodologi masalah. Masalah-masalah ini tentu mencakup antara perbedaan populasi dalam suatu spesies dan termasuk masalah baik dengan langkah-langkah yang digunakan untuk mengukur variasi temporal (McArdle et al. 1990; McArdle & Gaston 1992, 1995) dan penggabungan tersebut dari berbagai sumber variasi (Stewart-Oaten et al. 1995). Kekhawatiran tentang indeks matematika populasi variabilitas ukuran tampaknya relatif tidak penting untuk salmonid, data yang disajikan di sini karena perkiraan populasi variabilitas ukuran yang sangat konsisten untuk masing-masing tiga tindakan variabilitas (StdLog, CoefVar dan PV Heath). Tiga langkah menyebabkan kesimpulan umum yang sama baik di tingkat sejenis dan congener; Namun, ada beberapa perbedaan kualitatif.
Gambar. 3. Korelasi antara variabilitas populasi Heath (Heath PV) dan koefisien variasi (CoefVar) untuk spesies kelimpahan (a); antara PV Heath dan standar deviasi dari log-transformasi spesies kelimpahan (StdLog) (b) dan antara CoefVar dan StdLog (c) berdasarkan tingkat korelasi Spearman, ketiga indeks tersebut sangat terkait.
Variabilitas ukuran populasi pada umumnya dipengaruhi oleh varietas faktor yang berbeda termasuk faktor ekstrinsik seperti ketersediaan sumber daya (Trzcinski, Walde & Taylor 2005) dan faktor-faktor intrinsik seperti variasi individu dalam kelangsungan hidup dan fekunditas (Uchmanski 1999, 2000). Untuk sebagian besar spesies yang dibahas di sini faktor tertentu yang mempengaruhi populasi variabilitas ukuran yang tidak jelas. Namun, data ikan Lahontan memberikan resolusi yang lebih besar tentang faktor yang menyebabkan variasi intraspesifik.
aliran gradien dan pabrik riparian masyarakat (Dunham et al. 1999). Menggunakan model regresi kuantil , Dunham et al. (2002) menunjukkan variasi pada kepadatan ikan di antara populasi yang berbanding terbalik dengan lebar: rasio kedalaman sungai. Variabel-variabel ini memberikan kontribusi dengan jumlah habitat yang tersedia selama kondisi aliran dasar (yaitu suhu air yang cocok, Dunham, Schroeter & Rieman 2003). Karena kondisi lingkungan di Great Basin sangat bervariasi, variabilitas lebar: rasio mendalam karena perbedaan curah hujan antara tahun mungkin juga berkontribusi pada variabilitas ukuran populasi individu dan resiko kepunahan.
Terlepas dari faktor ekstrinsik yang bertanggung jawab untuk perbedaan intraspesifik antara populasi, keragaman rendah dari ikan Lahontan dibandingkan dengan ikan Steelhead dengan populasi cukup mengejutkan. Kami telah memprediksi bahwa populasi ikan Lahontan yang kami teliti terbatas terisolasi dihulu mencapai ukuran variabilitas populasi yang menunjukan pencapaian yang lebih tinggi akibat paparan yang lebih besar untuk menduga lingkungan. Sebaliknya, individu ikan Steelhead mungkin bisa memilih habitat yang menguntungkan meskipun menduga lingkungan dan akses untuk rentang yang lebih besar dari lingkungan Kondisi ini dapat mengakibatkan ukuran populasi lebih stabil. Perbedaan riwayat hidup antara dua spesies mungkin membantu menjelaskan perbedaan ini. Populasi ikan Steelhead biasanya anadromous dan semelparous dengan investasi pertelur kurang dari ikan Lahontan (Crespi & Teo 2002). Investasi ini yang relatif lebih besar dengan ikan Lahontan menjadi penyangga populasi mereka terhadap fluktuasi yang besar (Winemiller 2005).
Sayangnya, perbandingan interspesifik antara dua spesies terbatas dalam sejauh mana mereka memungkinkan digeneralisasikan secara kesimpulan ekologi atau evolusi (Garland & Adolph 1994). Dengan demikian, kita tidak mempertimbangkan perbedaan yang ditunjukkan di sini antara ikan Steelhead dan ikan Lahontan untuk selalu mewakili perbedaan antara anadromous dan potamodromous salmon. Sebaliknya perbandingan ini harus dipandang sebagai contoh bagaimana proses estimasi berlipat dapat digunakan untuk menghasilkan perkiraan tingkat spesies ukuran variabilitas populasi dan memungkinkan perbandingan interspesifik tanpa campuran dalam perbedaan spesies dengan perbedaan spesies.
Identifikasi keragaman yang signifikan antara populasi ikan Lahontan dicerminkan dalam temuan kami untuk salmon lainnya. Dalam semua kasus di mana beberapa populasi dipantau, dengan keragaman yang diidentifikasi signifikan (Tabel 1). Kami juga mengidentifikasi keragaman yang signifikan, di mana perbedaan tersebut tidak akan digabungkan dengan perbedaan panjang pengambilan sampel (Tabel 2). Selain itu, meskipun ada perbedaan yang signifikan antara spesies, lebih banyak variasi dalam bagaimana ukuran populasi sangat berfluktuasi hadir di intra dari tingkat interspesifik (Tabel 3). Hasil ini konsisten dengan orang-orang untuk taksa lainnya. Untuk Misalnya, Schoener & Spiller (1992) menunjukkan cukup dengan perbedaan intraspesifik ukuran variabilitas populasi untuk laba-laba. Marsh (2001) sama menunjukkan bahwa amfibi menunjukkan perbedaan besar dalam ukuran variabilitas populasi antara keluarga taksonomi. Sayangnya tidak satupun dari kasus ini adalah perbedaan intraspesifik eksplisit bunga.
ukuran variabilitas populasi, jadi sementara ikan mungkin secara umum menunjukkan variabilitas yang lebih tinggi daripada burung, hubungan antara ukuran populasi variabilitas salmon vs ikan lain saat ini tidak jelas. Namun, Winemiller (2005) memberikan beberapa dasar untuk menghasilkan prediksi bagaimana salmon mungkin berbeda dari spesies lain: spesies dengan keproduktifan rendah, ukuran telur yang lebih besar dan perawatan orang tua diharapkan memiliki populasi yang relatif lembab ukuran variabilitas. Hasil kami umumnya konsisten dengan harapan ini. Sebagai contoh, Chinook salmon memiliki telur yang relatif lebih besar dan keproduktifan rendah dari Sockeye salmon (Crespi & Teo 2002) dan juga menunjukkan lebih rendah ukuran variabilitas populasi (Tabel 1). Dengan demikian riwayat hidup memungkinkan beberapa prediksi umum bagaimana ikan spesies berbeda, meskipun ini mungkin rumit dengan Status migrasi (misalnya anadromous vs potadromous) dari spesies atau populasi.
Meskipun demonstrasi perbedaan intra dan interspesifik, peran sampel periode panjang yang berbeda dapat memiliki estimasi tingkat spesies yang membutuhkan studi lebih lanjut. Karena hubungan yang kuat antara panjang sampel dan ukuran variabilitas populasi, tampaknya mungkin bahwa variabilitas dapat diremehkan untuk spesies yang sebagian besar sampel telah dilakukan uji selama jangka waktu yang singkat. Sejauh mana hal ini benar dan yang bias ini dapat dihapus secara statistik harus dilakukan tujuan penelitian lebih lanjut.
Kehadiran pola taksonomi bagaimana populasi ukuran bervariasi dengan waktu yang telah sangat diperdebatkan dalam ekologi literatur (Connell & Sousa 1983; Schoener 1985; Inchausti & Halley 2002). Berdasarkan ketersediaan peningkatan jangka panjang data, ukuran variabilitas populasi sekarang tampaknya secara umum independen pengelompokan taksonomi utama (Inchausti & Halley 2002) tetapi bertentangan dengan laporan tetap (misalnya Reed & Hobbs 2004). Namun, penilaian skala besar pola taksonomi dapat mengandalkan hanya beberapa populasi spesies atau beberapa genus spesies. Sebagai hasil kami menunjukkan, ukuran variabilitas populasi dapat berbeda jauh antara populasi spesies tunggal. Dengan demikian setiap pola taksonomi skala besar yang sebenarnya mungkin dikaburkan karena variasi dalam taksa. Misalnya, meskipun ada perbedaan yang signifikan antara spesies salmonid, variasi tetap di tingkat intraspesifik. Dalam analisis konvensional, variasi ini akan tetap dalam denominator sisa F-nilai yang bisa mengaburkan perbedaan taksonomi. Penegasan atau pemberhentian luas pola taksonomi harus dinilai ulang setelah memperhitungkan kekhawatiran ini. Salah satu pendekatan yang potensial untuk menyelesaikan masalah ini adalah model pendekatan berlipat-linear yang digunakan di sini untuk beberapa populasi sampel. Pendekatan ini menghasilkan estimasi spesies yang dapat digunakan dalam perbandingan taksonomi yang luas.
dijelaskan di sini bersama dengan pendekatan baru menggabungkan metode filogenetik dan metaanalisis (Adams 2008; Lajeunesse 2009).
Ucapan Terima Kasih
Kami berterima kasih kepada Complex USFWS Lahontan National Fish Hatchery untuk membiayai, banyak petugas lapangan dan Steve Jenkins untuk diskusi mendalam. Kita juga mengucapkan terima kasih kepada Redaksi dan dua dewan juri yang komentar pada versi sebelumnya naskah ini sangat meningkatkan ruang lingkup dan kejelasan.
Referensi
Adams, D.C. (2008) Phylogenetic meta-analysis. Evolution, 62, 567–572.
Arino, A. & Pimm, S.L. (1995) On the nature of population extremes. Evolutionary Ecology, 9, 429–443.
Bengtsson, J. & Milbrink, G. (1995) Predicting extinctions: interspecific competition, predation and population variability in experimental Daphnia populations. Oecologia, 101, 397–406.
Coffin, P.D. & Cowan, W.F. (1995) Lahontan Cutthroat Trout (Oncorhynchus clarki henshawi) Recovery Plan. U.S. Fish and Wildlife Service, Portland, OR, USA.
Connell, J.H. & Sousa, W.P. (1983) On the evidence needed to judge ecological stability or persistence. American Naturalist, 121, 789–824.
Crespi, B.J. & Teo, R. (2002) Comparative phylogenetic analysis of the evolution of semelparity and life history in salmonid fishes. Evolution, 56, 1008– 1020.
Dunham, J.B. & Vinyard, G.L. (1996) Dysfunction characteristics of small trout populations. Final research report for research joint venture agreement. US Forest Service.
Dunham, J.B. & Vinyard, G.L. (1997) Incorporating stream level variability into analyses of site level fish habitat relationships: some cautionary examples. Transactions of the American Fisheries Society, 126, 323–329.
Dunham, J.B., Peacock, M.M., Rieman, B.E., Schroeter, R.E. & Vinyard, G.L. (1999) Local and geographic variability in the distribution of stream-living Lahontan cutthroat trout. Transactions of the American Fisheries Society, 128, 875–889.
Dunham, J.B., Cade, B.S. & Terrell, J.W. (2002) Influences of spatial and temporal variation on fish-habitat relationships defined by regression quantiles. Transactions of theAmerican Fisheries Society, 131,86–98.
Dunham, J., Schroeter, R. & Rieman, B. (2003) Influence of maximum water temperature on occurrence of Lahontan cutthroat trout within streams. North American Journal of Fisheries Management, 23, 1042–1049.
Dwyer, W.P., Shepard, B.B. & White, R.G. (2001) Effect of backpack electroshock on westslope cutthroat trout injury and growth 110 and 250 days posttreatment. North American Journal of Fisheries Management, 21, 646–650.
Fagan, W.F., Meir, E., Prendergast, J., Folarin, A. & Karieva, P. (2001) Characterizing population vulnerability for 758 species. Ecology Letters, 4,132– 138.
Garland, T., Bennett, A.F. & Rezende, E.L. (2005) Phylogenetic approaches in comparative physiology. Journal of Experimental Biology, 208, 3015–3035.
Hanski, I. (1990) Density dependence, regulation and variability in animal populations. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 330, 141–150.
Heath, J.P. (2006) Quantifying temporal variability in population abundances. Oikos, 115, 573–581.
Inchausti, P. & Halley, J. (2001) Investigating long-term ecological variability using the global population dynamics database. Science, 293, 655–657.
Inchausti, P. & Halley, J. (2002) The long-term temporal variability and spectral colour of animal populations. Evolutionary Ecology Research, 4,1033– 1048.
Inchausti, P. & Halley, J. (2003) On the relation between temporal variability and persistence time in animal populations. Journal of Animal Ecology, 72, 899–908.
Lajeunesse, M.J. (2009) Meta-analysis and the comparative phylogenetic method.American Naturalist, 174, 369–381.
Lawton, J.H. (1988) Population dynamics: more time means more variation. Nature, 334, 563–563.
Legendre, S., Schoener, T.W., Clobert, J. & Spiller, D.A. (2008) How is extinction risk related to population-size variability over time? A family of models for species with repeated extinction and immigration American Naturalist, 172, 282–298.
Lima, M., Marquet, P.A. & Jaksic, F.M. (1998) Population extinction risks of three Neotropical small mammal species. Oecologia, 115, 120–126.
Marsh, D.M. (2001) Fluctuations in amphibian populations: a meta-analysis. Biological Conservation, 101, 327–335.
May, R.M. (1973) Stability and Complexity in Model Ecosystems. Princeton University Press, Princeton, NJ, USA.
McArdle, B.H. & Gaston, K.J. (1992) Comparing population variabilities. Oikos, 64, 610– 612.
McArdle, B.H. & Gaston, K.J. (1995) The temporal variability of densities – back to basics. Oikos, 74, 165–171.
McArdle, B.H., Gaston, K.J. & Lawton, J.H. (1990) Variation in the size of animal populations: patterns, problems and artifacts. Journal of Animal Ecology, 59, 439– 454.
Mitton, C.J.A. & McDonald, D.G. (1994) Consequences of pulsed DC electrofishing and air exposure to rainbow trout (Oncorhynchus mykiss). Canadian Journal of Fisheries and Aquatic Sciences, 51, 1791–1798.
NERC Centre for Population Biology, I. C. (1999) The Global Population Dynamics Database. Available at: http://www.sw.ic.ac.uk/cpb/cpb/ gpdd.html.
Neville, H.M., Dunham, J.B. & Peacock, M.M. (2006) Landscape attributes and life history variability shape genetic structure of trout populations in a stream network. Landscape Ecology, 21, 901–916.
Peterson, C.H. (1984) Does a rigorous criterion for environmental identity preclurde the existence of multiple stable points. American Naturalist, 124, 127–133.
Pimm, S.L. & Redfearn, A. (1988) The variability of population densities. Nature, 334, 613– 614.
Pimm, S.L., Jones, H.L. & Diamond, J. (1988) On the risk of extinction.American Naturalist, 132, 757–785.
Ray, C., Peacock, M.M. & Dunham, J.B. (2007) Demography and population dynamics of Lahontan cutthroat trout (Oncorhynchus clarkii henshawi) stream populations in Eastern Nevada. Final research report for cooperative research agreement. USFWS. Reed, D.H. & Hobbs, G.R. (2004) The relationship between population size and temporal
variability in population size. Animal Conservation, 7,1–8.
Roff, D.A. (2006) Introduction to Computer-Intensive Methods of Data Analysis in Biology. Cambridge University Press, Cambridge.
Schoener, T.W. (1985) Are lizard population sizes unusually constant through time? American Naturalist, 126, 633–641.
Schoener, T.W. & Spiller, D.A. (1992) Is extinction rate related to temporal variability in population size? An empirical answer for orb spiders. American Naturalist, 139, 1176– 1207.
Stewart-Oaten, A., Murdoch, W.W.&Walde, S.J. (1995) Estimation of temporal variability in populations. American Naturalist, 146, 519–535.
Trzcinski, M.K., Walde, S.J. & Taylor, P.D. (2005) Stability of pitcher-plant microfaunal populations depends on food web structure. Oikos, 110,146–154.
Uchmanski, J. (1999) What promotes persistence of a single population: an individual-based model. EcologicalModelling, 115, 227–241.
Uchmanski, J. (2000) Individual variability and population regulation: an individual-based model. Oikos, 90, 539–548.
Van Deventer, J.S. & Platts, W.S. (1989) Microcomputer software system for generating population statistics from electrofishing data: User guide for microfish 3.0. USDAGen. Tech. Rep. INT-254.
Vucetich, J.A., Waite, T.A., Qvarnemark, L. & Ibarguen, S. (2000) Population variability and extinction risk. Conservation Biology, 14, 1704–1714.
Winemiller, K.O. (2005) Life history strategies, population regulation, and implications for fisheries management. Canadian Journal of Fisheries and Aquatic Sciences, 62, 872– 885.
Informasi Pendukung
Tambahan Informasi Pendukung dapat ditemukan pada versi online artikel ini.
Lampiran S1. GPDD mengidentifikasi nomor panggilan untuk dataset yang digunakan dalam penelitian ini.