MUTASI GEN xyncmu DAN PENGARUHNYA TERHADAP
STABILITAS ENZIM XILANASE CMU DARI Bacillus
halodurans
DAHMAYANTI SARIDEWI
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Mutasi Gen xyncmu dan Pengaruhnya terhadap Stabilitas Enzim Xilanase CMU dari Bacillus halodurans yang dilakukan di laboratorium Teknologi Bioindustri LAPTIAB-BPPT adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan sebagian hak cipta dari karya tulis saya kepada Laboratorium Pengembangan Teknologi Agroindustri dan Biomedika, Badan Pengkajian dan Penerapan Teknologi (LAPTIAB-BPPT) dan Institut Pertanian Bogor (IPB).
Bogor, Juli 2013
ABSTRAK
DAHMAYANTI SARIDEWI. Mutasi Gen xyncmu dan Pengaruhnya terhadap Stabilitas Enzim Xilanase CMU dari Bacillus halodurans. Dibimbing oleh SURYANI dan NIKNIK NURHAYATI.
Aplikasi enzim xilanase dalam industri kertas memerlukan enzim dengan karakteristik yang tahan terhadap pH dan suhu tinggi. Salah satu alternatif agar dapat mengubah karakteristik enzim tersebut yaitu pendekatan melalui mutasi pada titik yang diarahkan atau site directed mutagenesis PCR sehingga mengubah beberapa titik asam amino pada klon tetua (Klon M3) yang akan berpengaruh pada uji stabilitas enzim. Penelitian ini bertujuan mengubah stabilitas xilanase CMU dan membandingkan karakter antara xilanase CMU mutan dan normal. Langkah awal dari penelitian ini melibatkan plasmid dengan klon M3 yang dimutasi dengan dua pasang primer mutagenik yaitu Q53K dan T194I. Amplikon plasmid mutan pGEMalkxynaq1cmu M3 ditransformasi ke Escherichia coli DH5α dan diseleksi dengan media ampisilin. Hasil seleksi dengan ampisilin dan analisis restriksi mengonfirmasi dua klon mutan yaitu Klon 4 dan Klon 7 positif membawa gen xyncmu. Mutasi gen yang telah berhasil dilakukan sehingga didapatkan Klon 7 yang memiliki karakteristik enzim yang tahan terhadap pH dan suhu tinggi.
Kata kunci: mutagenesis situs terarah (site directed mutagenesis), gen xyncmu, xilanase CMU, stabilitas enzim
ABSTRACT
DAHMAYANTI SARIDEWI. xyncmu Gene Mutation and the Effect of Enzyme Stability Xylanase CMU from Bacillus halodurans. Supervised by SURYANI and NIKNIK NURHAYATI.
Applications of xylanase enzyme on paper industry requires an enzyme characteristic through pH and resistant to high temperatures. A directed point mutations in site directed mutagenesis PCR which being the alternative to changing the enzyme’s characteristic and a several of amino acid on parental clon (M3) which effected with enzyme stability. This study aims to alter the stability of xylanase CMU and comparing the characters between CMU xylanase’s mutant and normal. The first step involves a plasmid with parental clones (Clone M3) which transferred to two pairs of mutagenic primers that Q53K and T194I. The amplicons from mutant plasmid pGEMalkxynaq1cmu M3 were transformed into Escherichia coli DH5α and selected with ampicillin media. Results of selection with ampicillin and restriction analysis were confirmed two positive mutant clones are Clone 4 and Clone 7 which brought of xyncmu gene. Gene mutation has succeeded to done as the result was Clone 7 which had enzyme’s characteristic through pH and resistant to high temperatures.
MUTASI GEN xyncmu DAN PENGARUHNYA TERHADAP
STABILITAS ENZIM XILANASE CMU DARI Bacillus
halodurans
DAHMAYANTI SARIDEWI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
Pada
Departemen Biokimia
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
Judul : Mutasi Gen xyncmu dan Pengaruhnya terhadap Stabilitas Enzim Xilanase CMUdari Bacillus halodurans
Nama : Dahmayanti Saridewi NIM : G84090065
Menyetujui
Dr.Suryani S.P.,M Sc. Pembimbing I
Dr. Niknik Nurhayati Pembimbing II
Mengetahui
Ketua Departemen Biokimia
Dr. Ir. I Made Artika, M.App. Sc Ketua Departemen Biokimia
PRAKATA
Puji syukur penulis panjatkan kehadirat Tuhan Yang Maha Esa atas segala berkat, nikmat dan karunia-Nya sehingga penulis dapat menyelesaikan karya ilmiah ini. Penelitian ini mempunyai tema Biologi Molekuler dan telah dikerjakan sejak Januari 2013 hingga bulan Juli 2013. Penelitian ini berjudul Mutasi Gen xyncmu dan Pengaruhnya terhadap Stabilitas Enzim Xilanase CMU dari Bacillus halodurans.
Penulis mengucapkan terima kasih ibu Dr. Suryani S.P., M Sc. dan ibu Dr. Niknik Nurhayati selaku pembimbing yang selalu memberikan masukan selama penelitian dan karya ilmiah ini, para staf Labotarium Biologi Molekuler non Virus, Bioseparasi, dan Analisis LAPTIAB-BPPT, kepada keluarga penulis: Bapak Dahrizal, Ibu Sri Rusiana, Ibu Nurasiah, Dahvia Nursriyanti, Dharmasetya, Muadzimmahmud, dan Muhtiaistiqamah.
Semoga karya ilmiah ini memberikan nilai tambah bagi perkembangan ilmu pengethuan khususnya bidang Biologi Molekuler.
Bogor, Juli 2013
DAFTAR ISI
PRAKATA i
DAFTAR ISI ii
DAFTAR GAMBAR iii
DAFTAR LAMPIRAN iii
PENDAHULUAN 1
BAHAN DAN METODE 2
Alat 2
Bahan 2
Metode 3
HASIL 5
Mutan Plasmid pGEMalkxynaq1cmu M3 5
Urutan Nukleotida Klon Mutan Plasmid pGEMalkxynaq1cmu M3
6
Sifat Mutan Xilanase CMU Rekombinan 7
Stabilitas Enzim Xilanase CMU 8
PEMBAHASAN 9
MutanPlasmid Mutan pGEMalkxynaq1cmu M3 9
Urutan Nukleotida Klon Mutan Plasmid pGEMalkxynaq1cmu M3
10
Sifat Mutan Xilanase CMU Rekombinan 11
Stabilitas Enzim Xilanase CMU 11
SIMPULAN DAN SARAN 12
Simpulan 12
Saran 12
DAFTAR PUSTAKA 12
LAMPIRAN 15
RIWAYAT HIDUP 20
DAFTAR GAMBAR
1 Analisis fragmen menggunakan elektroforesis gel agarosa hasil SDM-PCR plasmid rekombinan pGEMalkxynaq1cmu M3 dan hasil restriksi dengan DpnI
5
2 Hasil analisis restriksi plasmid rekombinan pGEMalkxynaq1cmu M3 yang dimutasi
5
3 Hasil alignment dari klon tetua M3, galur liarnya yaitu klon P39, dan klon mutannya (klon 4 dan 7) dengan menggunakan ClustalW
6
4 Hasil pengukuran aktivitas spesifik enzim (U/mg) dengan pengaruh pH dan suhu yang berbeda
6
5 Hasil uji termostabilitas klon mutasi 7 pada suhu 60˚C, pH 9 dan 10 pada menit ke 0, 10, dan 20
6
DAFTAR LAMPIRAN
1 Strategi penelitian 13
2 Alignment gen alkxynaq1cmu M3 dengan program ClustalW (www.genome.jp)
14
3 Alignment mutasi gen pGEMalkxynaq1cmu M3 dengan klon 4 dan 7 16
PENDAHULUAN
Salah satu enzim yang dipakai dalam proses pemutihan kertas adalah enzim xilanase. Enzim ini sedang dikembangkan agar dapat membantu pemutihan kertas dengan efektif dan ramah lingkungan. Sebagian besar negara berkembang telah menggunakan enzim ini, namun di Indonesia belum terlalu banyak diaplikasikan dengan kendala biaya dan sebagainya (Batubara 2006).
Produksi kertas dengan bantuan enzim xilanase akan memiliki tingkat keputihan yang tinggi, lebih lentur, dan permukaan lebih halus (Rifaat et al. 2005). Enzim xilanase yang digunakan dalam industri kertas bersifat termostabil dan tahan pada pH basa. Suasana basa dapat membantu degradasi lignin oleh hidrolisis glukosa pada pulp. Xilanase termostabil dapat mengurangi kontaminasi mikroorganisme. Perbaikan suhu xilanase untuk mencapai termostabil dapat dilakukan dengan mutagenesis situs terarah (site-directed mutagenesis (SDM)) yaitu salah satu program dalam reaksi berantai polymerase (polymeraze chain reaction (PCR)) (Zhang et al 2010).
Program ini ditemukan oleh Charles Weissman pada tahun 1973 dengan analog nukleotida N4-hidroksisitidina yang mengubah transisi basa nitrogen GC menjadi AT. Program SDM pada PCR disingkat menjadi SDM-PCR. Metode biologi molekuler ini digunakan untuk membuat perubahan spesifik maupun intensional pada urutan nukleotida pada gen. Metode ini juga dilakukan untuk meneliti struktur dan aktivitas biologi pada DNA, RNA, dan molekul protein. Awalnya metode ini menggunakan radiasi yang kurang spesifik atau mutagen kimia (Schaffner dan Weissmann 1973).
Kelebihan spesifik penggunaan SDM-PCR adalah dapat meminimalisir adanya kontaminan selain amplikon dari cetakan dan primer yang digunakan, hanya memerlukan jumlah primer yang sedikit, serta mudah memperlihatkan fragmen besar pada DNA. Ukuran fragmen yang terpecah dari proses insersi maupun substitusi dari fragmen lainnya bergantung pada panjang urutan basa nukleotida yang terdapat pada primer (Zheng et al. 2004). Manipulasi genom yang dilakukan oleh Storici dan Resnick pada tahun 2006 dapat mengurangi kesulitan dalam menemukan titik mutasi yang diinginkan dan menghilangkan urutan asam amino heterolog pada gen yang diinginkan.
Penggunaan enzim xilanase sebagai alternatif mengurangi penggunaan klor serta lebih ramah lingkungan memerlukan karakter enzim yang stabil terhadap suhu tinggi dan pH basa. Hal ini disebabkan produksi kertas berlangsung pada pH yang lebih besar dari 8 dan suhu tinggi yaitu lebih dari 65. Mikroba sebagai sumber potensial penghasil enzim ini selain arkaea jarang ditemui yang dapat hidup dalam kondisi ekstrim. Umumnya enzim ini dihasilkan kurang stabil terhadap kondisi ekstrim. Contohnya enzim xilanase Aq1 yang dihasilkan bakteri mesofilik Bacillus subtilis Aq1 memiliki pH 7 dan suhu optimum 55˚C (Wahyuntari et al 2009).
Tim peneliti di Pusat Teknologi Bioindustri BPPT telah memiliki beberapa galur mikroba potensial penghasil xilanase, salah satunya Bacillus halodurans CMU yang didapatkan dari Cimanggu (Ulfah et al 2011). Xilanase dari B. halodurans CMU tersebut telah berhasil diklon dan diekspresikan pada sel E. coli
pH dan suhu tinggi telah banyak dilaporkan. Salah satunya peningkatan thermostabilitas xilanase dari Geobacillus stearothermophilus dengan pendekatan Site Directed Mutagenesis-PCR (Zhang et al 2010). Berdasarkan laporan tersebut tim peneliti Bioindustri-BPPT telah merancang mutasi beberapa titik asam amino pada xilanase CMU dan mensintesis beberapa pasang primer mutagenik pada titik-titik asam amino dimaksud. Lima klon mutan tunggal dan satu klon mutan ganda telah berhasil diperoleh melalui SDM-PCR menggunakan beberapa pasang primer mutagenik pada gen xilanase CMU (Pratiwi 2013).
Penelitian ini bertujuan untuk mengubah asam amino glutamina pada posisi 53 menjadi lisina (Q53K) dan treonina pada posisi 156 menjadi isoleusina (T194I) masing-masing pada deret asam amino mutan ganda M3 menjadi 2 mutan dengan mutasi 3 titik dengan pendekatan SDM-PCR. Pengaruh mutasi tiga titik terhadap karakter enzim xilanase CMU dianalisa serta dibandingkan dengan xilanase CMU mutan ganda (M3) dan normal. Manfaat dari penelitian ini adalah dengan menemukan ketahanan enzim xilanase yang lebih tinggi, enzim tersebut dapat digunakan di dalam industri kertas yang relatif menggunakan suhu dan pH tinggi dalam proses pembuatan kertas.
BAHAN DAN METODE
Alat
Alat-alat yang digunakan adalah mikropipet, alat-alat gelas, jarum ose, elektroforesis (Mupid-exu), Mini Spin (Eppendorf), thermal cycler PCR (Eppendorf), sentrifus dingin (Sorvale Fresco), konsentrator (Eppendorf), dokumentasi gel kodak, thermomixer (Eppendorf), shaker incubator (Kuhner), water purification system (Millipore), laminar air flow class II BSC (ESCO), freezer -85°C (NUAIRE), microwave, autoklaf (IWAKI), alat pengaduk (vortex mixer), spektrofotometer UV-Fis (Hitachi U-2001), pH meter digital (WTW series inolab), neraca analitik (Radwag), dan inkubator (Memert).
Bahan
Bahan-bahan media yang digunakan yaitu media Luria bertani (LB), super optimal broth (SOB), dan super optimal broth with catabolite repression (SOC). Klon yang akan dijadikan perbandingan dengan klon mutan (Klon 4 dan 7) saat diuji aktivitas dan stabilitasnya adalah klon tetua (M3) dan galur liar (P39). Primer yang digunakan adalah dua pasang primer mutagenik Q53K dan T194I. Bahan yang digunakan untuk SDM yaitu Phusion high-fidelity DNA polymerase (Thermo), 5× bufer HF, enzim restriksi DpnI, EcoRI, dan XbaI, dan sel kompeten Escherichia coli DH5α. Bahan kimia yang digunakan adalah ampisilin, asam dinitro salisilat (DNS) Miller, bufer TrisCl pH 8 dan 9, bufer glisin + NaOH pH 10 dan 11, xilan Beechwood, kit untuk ekstraksi DNA (Fermentas).
Metode Penelitian
dari penelitian sebelumnya (Pratiwi 2013). Plasmid rekombinan pGEMalkxynaq1cmu M3 merupakan mutan ganda yang 2 asam aminonya dimutasi menggunakan primer mutagenik Q62L dan E146V dengan pendekatan SDM – PCR. Penelitian ini akan menambahkan mutasi ke-3 pada plasmid mutan ganda M3 menggunakan pasangan primer mutagenik Q53K atau T194I sehingga didapatkan 2 mutan dengan tiga titik (M3+Q53K dan M3+T194I) yang diharapkan memiliki termostabilitas yang tinggi.
Mutasi Plasmid Rekombinan pGEMalkxynaq1cmu M3 dengan Metode SDM-PCR (Zheng et al. 2004). Plasmid pGEMalkxynaq1cmu M3 dimutasi menggunakan metode SDM-PCR. Komposisi reaksi SDM-PCR sebagai berikut: 5× bufer HF 5 µl, 10 mM dNTP mix 0.5 µl, DMSO 0.75 µl, 2.5 U/µl Phusion high-fidelity DNA polymerase (Thermo), 0.5 µl primer forward, 0.5 µl primer revers, plasmid pGEMalkxynaq1cmu M3 0.5 µl, dan ddH2O 17 µl. Kondisi SDM-PCR sebagai berikut: program denaturasi (hot start) selama 2 menit pada suhu 98°C dan dilanjutkan 26 siklus. Setiap siklus SDM-PCR terdiri dari tahapan: denaturasi DNA cetakan selama 30 detik pada suhu 98°C, penempelan primer selama 30 detik pada suhu annealing (bergantung primernya), dan pemanjangan primer selama 1 menit 30 detik pada suhu 72°C. Suhu annealing masing-masing primer sama yakni Q53K dan T194I adalah 64.6°C. Pasca SDM-PCR dilakukan selama 5 menit pada suhu 72°C. Hasil amplifikasi kemudian dipotong dengan enzim restriksi DpnI.
Pemotongan Plasmid Mutan pGEMalkxynaq1cmu M3 dengan Enzim Restriksi DpnI (Zheng et al. 2004). Mutan plasmid pGEMalkxynaq1cmu M3 yang telah didapatkan dari hasil SDM-PCR dipotong dengan enzim restriksi DpnI yang hanya akan memotong DNA yang termetilasi, yaitu plasmid M3 yang diisolasi dari E. coli DH5α yang dijadikan sebagai DNA cetakan pada reaksi SDM-PCR. Komposisi reaksinya sebagai berikut: 17.8 µl produk PCR, 0.2 µl enzim restriksi DpnI 20 U/ µl, dan 2 µl bufer 4 dicampurkan dan diinkubasi pada suhu 37°C selama tiga jam, kemudian diinkubasi pada suhu 80°C selama 20 menit untuk menginaktifasi enzim restriksi. Hasil restriksi dengan enzim restriksi DpnI lalu ditransformasi ke dalam sel kompeten E. coli DH5α untuk perbanyakan dan diisolasi plasmidnya. Transformasi mengikuti metode heat shock (Zheng et al. 2004).
Produksi Enzim Xilanase CMU dari Plasmid Mutan pGEMalkxynaq1cmu M3 (Huang et al. 2006). Sebanyak satu ose koloni transforman plasmid mutan pGEMalkxynaq1cmu M3 ditumbuhkan lebih kurang 18 jam dalam media LB-ampisilin. Setelah itu dilakukan kultivasi koloni mutan dalam 100 ml media LB-ampisilin. Kultur tersebut disentrifugasi selama 15 menit pada 6000 rpm 4°C. Bagian supernatan dibuang dan pada bagian pelet ditambahkan bufer natrium phosphat 20 mM, pH 7 yang mengandung merkaptoetanol 1 mM sebanyak 5 ml. Pelet diresuspensi dengan vortex, kemudian dilisis dengan sonikator tiap 20 detik selama 3 menit. Proses lisis sel dilakukan dalam keadaan dingin dan hasil sonikasi disentrifugasi pada 6000 rpm 4°C, selama 10 menit. Supernatan diambil dan dilakukan uji aktivitas.
Uji Aktivitas Xilanase CMU secara Kuantitatif (Bailey 1992). Sebanyak 400 µl larutan bufer pH yang sudah ditentukan dimasukkan ke dalam masing-masing tabung mikro dan ditambahkan 50 µl substrat beech wood xylan 10%, lalu diinkubasi pada suhu yang telah ditentukan selama 2 menit. Ke dalam setiap tabung mikro dimasukkan 50 µl enzim kasar, kemudian diinkubasi dilanjutkan selama 5 menit. Setelah itu, dimasukkan 750 µl asam dinitro salisilat (DNS), dipanaskan pada suhu 100°C selama 5 menit dan didinginkan. Sampel diukur absorbansinya pada panjang gelombang 540 nm. Blanko dibuat dengan cara yang sama kecuali penambahan enzim dilakukan setelah penambahan DNS. Nilai absorbansi sampel dan blanko diukur pada panjang gelombang 540 nm.
Nilai absorban ini dimasukkan ke dalam kurva standar xilosa. Aktivitas volumetrik enzim dihitung dengan rumus sebagai berikut:
Aktivitas enzim (U/ml) = konsentrasi enzim × 1000 × faktor pengenceran
Bobot molekul xilosa × Volume enzim × waktu reaksi
Setiap satu unit aktivitas xilanase didefinisikan sebagai jumlah enzim yang diperlukan untuk menghidrolisis xilan menjadi 1 µmol xilosa per menit pada suhu dan pH telah dtentukan. Data kadar protein ditentukan dengan menggunakan persamaan dari kurva standar BSA (Bovine Serum Albumin). Nilai kadar protein total diperlukan untuk menentukan aktivitas spesifik enzim yang dihitung dengan cara membagi aktivitas volumetrik enzim dengan berat total protein dan dinyatakan dalam satuan U/mg.
Analisis Restriksi Plasmid Mutan pGEMalkxynaq1cmu M3 (New England Biolabs 2005).
Plasmid mutan yang telah diekstrak lalu dipotong dengan enzim restriksi. Komposisi restriksi sebagai berikut: 2 µl plasmid alkxynaq1cmu rekombinan, 0.1 µl enzim restriksi EcoRI, 0.2 µl XbaI, 10× NEBuffer react 2 dan 2.2 µl ddH2O diinkubasi pada suhu 37°C selama 1 jam. Hasil restriksi dapat dilihat dari potongan fragmennya dengan menggunakan elektroforesis gel agarosa.
Analisis Urutan Nukleotida (sequencing)
Plasmid yang membawa gen mutan dari plasmid rekombinan pGEMalkxynaq1cmu M3 kemudian dianalisis urutan nukleotidanya untuk mengetahui basa yang terdapat pada gen tersebut. Prosedur analisis urutan nukleotida (sequencing) melalui Genetika Science.
Urutan asam amino deduksi ditentukan menggunakan program BLASTn (http://blast.ncbi.nlm.nih.gov/Blast.cgi) dan ClustalW (http://www.genome.jp), serta perangkat lunak Chromas Lite untuk melihat hasil sequencing dan kromatogramnya , dan Clone Manager untuk melihat simulasi kloning, operasi enzim, serta gambar peta grafik dari kloning mutan.
Hasil analisis urutan nukleotida dari Genetica Science, dianalisis kembali menggunakan perangkat lunak BLAST, Chromas Lite, dan Clone Manager. Analisis menggunakan perangkat tersebut dapat menunjukkan urutan nukleotida yang termutasi.
Optimasi suhu dan pH reaksi xilanase (Bailey 1992)
Optimasi reaksi dilaukan dengan mengukur aktivitas enzim ada suhu 60˚C dan 65°C serta pH 9 (bufer Tris-HCl) dan pH 10 (bufer glisin).
Uji Stabilitas Mutan Xilanase (Bailey 1992)
Ketahanan mutan xilanase terhadap suhu dan pH dilakukan dengan cara menginkubasi enzim tanpa substrat pada suhu 60°C selama periode waktu 0, 10, dan 20 menit pada pH 9 dan 10. Aktivitas enzim setelah pemanasan pada masing-masing periode waktu diukur dengan melakukan pengukuran aktivitas xilanase dengan metode Bailey (1992) seperti diuraikan diatas.
HASIL
Mutan Plasmid pGEMalkxynaq1cmu M3
Penelitian sebelumnya (Pratiwi 2013) telah menghasilkan plasmid rekombinan pGEMalkxynaq1cmu M3 yang merupakan mutan ganda yang mengandung 2 titik mutasi asam amino. Plasmid ini dimutasi kembali dengan dua pasang primer melalui SDM-PCR untuk mendapatkan mutan ketiga yang mengandung 3 titik mutasi asam amino. Hasil analisis pada elektroforesis gel agarosa (Gambar 1) menunjukkan proses SDM-PCR berhasil mengamplifikasi plasmid rekombinan M3 dan menghasilkan pita DNA berukuran 4300 pb. Plasmid berukuran 4300 pb terdiri atas vektor pGEMT-Easy yang berukuran 3000 pb dan gen xyncmu yang berukuran 1300 pb. Gen xyncmu terdiri atas sejumlah basa nukleotida berukuran 1100 pb dari klon tetua yaitu Bacillus halodurans dan promotor xynaq1 yang berjumlah 200 pb.
4000 pb
Gambar 1 Analisis elektroforesis gel agarosa produk SDM-PCR plasmid pGEMalkxynaq1cmu M3 (kiri) menggunakan primer Q53K (1), dan primer T194I (2) serta hasil pemotongan dengan enzim restriksi DpnI (kanan) produk SDM-PCR dengan primer Q53K (1), dan primer T194I (2). M: 1 kb DNA Ladder (Fermentas), pb = pasang basa.
multiple cloning site yang mengapit gen xilanase cmu, sedangkan XbaI memotong gen xilanase cmu pada sekitar basa ke 300 dari kodon awal. Gambar 2 menunjukkan beberapa klon hanya terpotong oleh enzim EcoRI menghasilkan fragmen DNA berukuran 1300 pb (xilanase CMU) dan 3000 pb (vektor pGEMT) dan beberapa klon lainnya terpotong oleh kedua enzim restriksi menghasilkan fragmen DNA berukuran 300 pb, 1000 pb dan 3000 pb. Selanjutnya dipilih klon positif yang membawa gen xyncmu yaitu Klon 4 dan Klon 7 dikultivasi untuk
Gambar 2 Hasil analisis enzim restriksi plasmid rekombinan pGEMalkxynaq1cmu M3 yang dimutasi dengan primer Q53K (1-5) dan primer T194I (6-10) M: 1 kb DNA Ladder (Fermentas).
Urutan Nukleotida Mutan Plasmid pGEMalkxynaq1cmu M3
Hasil analisis urutan nukleotida menunjukkan adanya mutasi yang terjadi pada Klon 4 dan Klon 7. Klon 4 merupakan klon dari plasmid rekombinan pGEMalkxynaq1cmu M3 yang dimutasi dengan primer Q53K mengalami perubahan asam amino urutan 53 yaitu asam amino glutamina dengan kodon CAA termutasi menjadi asam amino lisina dengan kodon AAA. Klon 7 merupakan klon dari plasmid yang sama (M3) namun dimutasi dengan primer T194I mengalami perubahan asam amino urutan 156 yaitu asam amino treonina dengan kodon ACA termutasi menjadi asam amino isoleusina dengan kodon ATA (Gambar 3).
Hasil pensejajaran memperlihatkan adanya mutasi pada klon normal mengalami perubahan asam amino urutan 62 yaitu asam amino glutamina dengan kodon CAA termutasi menjadi asam amino leusina dengan kodon CTA. Mutasi dari klon normal lainnya yang terbawa pada klon normal dan mutannya mengalami perubahan asam amino urutan 108 yaitu glutamat dengan kodon GAG menjadi valina dengan kodon GTG.
6
M 1 2 3 4 5 6 7 8 9 10 M
Gambar 3 Hasil pensejajaran (alignment) dari klon tetua M3, galur liarnya yaitu klon P39, dan klon mutannya (Klon 4 dan Klon 7) dengan menggunakan ClustalW, 1: Klon 4 dengan perubahan asam amino glutamina (CAA) menjadi lisina (AAA), 2: terjadi perubahan asam amino glutamina (CAA) menjadi leusina (CTA) pada Klon P39, 3: terjadi perubahan asam amino glutamat (GAG) menjadi valina (GTG) pada Klon P39, 4: Klon 7 dengan perubahan asam amino treonina (ACA) menjadi isoleusina (ATA)
Plasmid yang telah mengandung gen xyncmu dan promotor xilanase Aq1 diekspresikan pada vektor pGEMT-Easy. Penggunaan vektor tersebut dapat menghasilkan produktivitas tinggi.
Sifat Mutan Xilanase CMU Rekombinan
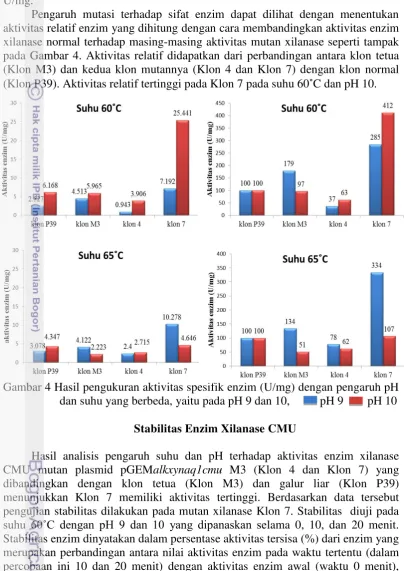
Mutasi xilanase tiga titik yang positif yaitu dari Klon 4 dan Klon 7 serta xilanase mutan ganda dari Klon M3 dan xilanase normal dari Klon P39 masing-masing diukur aktivitasnya pada suhu 60˚C dan 65˚C dan pH 9 dan 10. Klon yang memiliki nilai aktivitas tertinggi adalah Klon 7 yaitu klon mutan dari plasmid
aktivitas pada suhu 60˚C untuk pH 9 sebesar 4.513 U/mg dan pH 10 sebesar 5.965 U/mg; pada suhu 65˚C untuk pH 9 sebesar 4.122 U/mg dan pH 10 sebesar 2.223 U/mg.
Pengaruh mutasi terhadap sifat enzim dapat dilihat dengan menentukan aktivitas relatif enzim yang dihitung dengan cara membandingkan aktivitas enzim xilanase normal terhadap masing-masing aktivitas mutan xilanase seperti tampak pada Gambar 4. Aktivitas relatif didapatkan dari perbandingan antara klon tetua (Klon M3) dan kedua klon mutannya (Klon 4 dan Klon 7) dengan klon normal (Klon P39). Aktivitas relatif tertinggi pada Klon 7 pada suhu 60˚C dan pH 10.
Gambar 4 Hasil pengukuran aktivitas spesifik enzim (U/mg) dengan pengaruh pH dan suhu yang berbeda, yaitu pada pH 9 dan 10, pH 9 pH 10
Stabilitas Enzim Xilanase CMU
Hasil analisis pengaruh suhu dan pH terhadap aktivitas enzim xilanase CMU mutan plasmid pGEMalkxynaq1cmu M3 (Klon 4 dan Klon 7) yang dibandingkan dengan klon tetua (Klon M3) dan galur liar (Klon P39) menunjukkan Klon 7 memiliki aktivitas tertinggi. Berdasarkan data tersebut pengujian stabilitas dilakukan pada mutan xilanase Klon 7. Stabilitas diuji pada
suhu 60˚C dengan pH 9 dan 10 yang dipanaskan selama 0, 10, dan 20 menit. Stabilitas enzim dinyatakan dalam persentase aktivitas tersisa (%) dari enzim yang merupakan perbandingan antara nilai aktivitas enzim pada waktu tertentu (dalam percobaan ini 10 dan 20 menit) dengan aktivitas enzim awal (waktu 0 menit), seperti yang ditunjukkan pada Gambar 5. Aktivitas tersisa ensim setelah pemanasan pada suhu 60˚C selama 10 menit dengan pH 9 tampak masih tinggi hingga mencapai 91%, kemudian menurun hingga mencapai 39% ketika pemanasan diperpanjang hingga 20 menit.
Aktivitas menurun drastis hingga 21% setelah pemanasan pada suhu 60˚C pH 10 selama 10 menit.
Gambar 5 Hasil uji termostabilitas klon mutasi 7 pada suhu 60˚C, pH 9 dan 10 pada menit ke 0, 10, dan 20 pH 9 pH 10
PEMBAHASAN
Mutan Plasmid pGEMalkxynaq1cmu M3
Proses amplifikasi dengan SDM-PCR umumnya sama seperti PCR biasa yaitu mengamplifikasi cetakan dengan primer oligonukleotida spesifik. Cetakan yang digunakan adalah Klon M3, sedangkan primer oligonukleotida spesifiknya adalah Q53K dan T194I. Metode ini menggunakan enzim polimerase yang bersifat termostabil seperti Phusion high-fidelity DNA polymerase (Thermo) karena terdapat langkah awal yang melibatkan suhu tinggi yaitu 98˚C. Proses SDM-PCR digunakan pada penelitian sebelumnya (Zhang et al 2010) memperlihatkan pengaruhnya terhada aktivitas enzim dan termostabilitas yang diuji dengan ekstrak kasar enzim. Pemakaian metode ini juga bertujuan agar saat plasmid mutan pGEMalkxynaq1cmu M3 rekombinan ditransformasi, parentalnya tidak tercampur sehingga didegradasi oleh DpnI (Zhang et al 2010).
Proses amplifikasi bermula pada denaturasi cetakan dari klon tetua (Klon M3) serta mengubah bentuk dari untai ganda menjadi untai tunggal. Proses selanjutnya adalah annealing yaitu proses penempelan antara primer dengan cetakannya. Proses akhir dari langkah ini adalah tahapan pemanjangan (elongation) yaitu pemanjangan hasil untai tunggal yang merupakan gabungan antara primer dan cetakan yang diamplifikasi. Penambahan enzim polimerase pada awal langkah SDM-PCR ini bertujuan mensintesis untai baru DNA yang komplementer terhadap cetakannya (Schaffner dan Weissmann 1973).
kompeten Escherichia coliDH5α kemudian dijadikan sebagai DNA cetakan pada reaksi SDM-PCR. Sel kompeten Escherichia coliDH5α merupakan sel kompeten yang sering digunakan pada aplikasi kloning. Sel kompeten jenis DH5α juga dapat digunakan untuk melakukan seleksi biru dan putih, meningkatkan stabilitas insert atau gen yang akan ditransformasi, serta dapat meningkatkan kualitas plasmid DNA yang akan diekstraksi dengan cara minipreps (Molecular Cloning Laboratories 2010).
Proses inkubasi pada suhu 37˚C selama 3 jam bertujuan memisahkan cetakan dari klon normal (Klon P39) dari klon tetua (Klon M3), sedangkan inaktivasi enzim ini dilakukan pada suhu 80˚C selama 20 menit. Fragmen hasil pemotongan terbentuk bayang atau smear karena proses degradasi cetakan dari klon normal (P39), sehingga fragmen yang terbentuk pada 4300 pb merupakan produk SDM-PCR yang juga merupakan plasmid rekombinan dari klon tetua (Klon M3) (Zheng et al 2010).
Tahapan selanjutnya merupakan transformasi dengan heat shock. Tahap ini merupakan tahap perbanyakan sel pada E. coli DH5α dan menggunakan selectable marker antibiotik ampisilin. Transforman yang ditumbuhi pada media LB (Luria Bertani) yang ditambahkan ampisilin akan menumbuhkan bakteri yang membawa gen resistensi terhadap ampisilin, sehingga hanya E. coli dengan plasmid rekombinan pGEMalkxynaq1cmu M3 yang tumbuh pada media dengan antibiotik pada dosis toksik (Brown 1991). Transforman yang tumbuh pada media selektif kemudian diisolasi plasmidnya oleh kit ekstraksi DNA (Fermentas) untuk dianalisis nukleotida dan analisis restriksi. Analisis nukleotida bertujuan mengetahui urutan asam amino yang telah termutasi dan disejajarkan dengan klon normal dan tetuanya, sedangkan analisis restriksi bertujuan mengetahui ukuran plasmid yang menjadi target berukuran 4300 pb dengan enzim restriksi EcoRI dan XbaI. Enzim restriksi EcoRI memotong plasmid pada daerah multi cloning site yang mengapit gen xilanase CMU, sedangkan XbaI memotong gen xilanase CMU pada sekitar basa ke-300 dari kodon awal (Meijer et al 1986).
Transforman yang didapat dari media selektif kemudian diproduksi untuk diuji aktivitasnya. Pelet hasil produksi transforman kemudian ditambahkan dengan bufer natrium fosfat 20 mM pH 7 yang mengandung merkaptoetanol 1 mM sebanyak 5 ml dan disonikator tiap 20 detik selama 3 menit. Hal ini dapat melisis sel yang mengandung plasmid rekombinan pGEMalkxynaq1cmu M3 sehingga dapat diuji aktivitas enzimnya yang merupakan supernatan dari hasil sonikasi dan pelisisan tersebut (European Molecular Biology Laboratory 2009).
Urutan Nukleotida Klon Mutan Plasmid pGEMalkxynaq1cmu M3 (klon 4 dan 7)
Perubahan ini disebabkan mutasi sebelumnya pada klon tetua (Klon M3) dengan primer Q62L. Perubahan lain urutan asam amino terdapat pada posisi 108 yaitu asam amino glutamat (GAG) menjadi valina (GTG) yang disebabkan mutasi klon M3 dengan primer E146V seperti yang ditunjukkan pada Gambar 5 (Pratiwi 2013). Asam amino glutamina merupakan asam amino yang bersifat conserve atau memiliki kesamaan dengan asam amino posisi ke-53 pada urutan asam amino pada klon normal (Klon P39) dan urutan basa nukleotida pada Bacillus halodurans yang menghasilkan xilanase termostabil (Wahyuntari et al 2009).
Sifat Mutan Xilanase Rekombinan
Parameter pH berpengaruh terhadap kecepatan aktivitas enzim dalam mengatalis suatu reaksi. Konsentrasi ion hidrogen pada larutan bufer dengan pH tertentu mempengaruhi aktivitasnya. Setiap enzim memiliki pH optimum agar membentuk struktur yang kondusif dalam mengikat substrat. Jika konsentrasi ion hidrogen berubah dari titik optimal, aktivitas enzim menjadi tidak fungsional. Aktivitas enzim yang menurun saat perubahan pH disebabkan perubahan keadaan ion substrat dan enzim (Lehninger et al. 2008).
Peningkatan suhu lebih lanjut menurunkan aktivitas enzim karena enzim telah mengalami denaturasi dan perubahan konformasi pada suhu yang terlalu tinggi. Hal ini menyebabkan substrat terhambat dalam memasuki sisi aktif enzim. Pengukuran uji aktivitas enzim xilanase bertujuan mengukur produk hasil hidrolisis substrat xilan yang diubah menjadi 1 µmol gula pereduksi xilosa permenit pada suhu dan pH yang telah ditentukan (U/ml) (Yusriah dan Nengah 2013).
Data kadar protein didapatkan dari kurva standar BSA (Bovine Serum Albumin). Nilai kadar protein total bertujuan menentukan aktivitas spesifik enzim yang terukur dengan membagi aktivitas volumetrik enzim dengan berat total protein dan dinyatakan dalam satuan U/mg. Penambahan asam dinitro salisilat pada campuran substrat dan enzim bertujuan menghentikan aktivitas enzim pada suhu dan waktu yang telah ditentukan (Bailey 1992).
Stabilitas Enzim Xilanase CMU
Salah satu penyebab sifat stabilitas enzim tersebut adalah ketahanan dari denaturasi protein karena pengaruh panas (Muawanah 2006). Penggunaan mutasi dengan bertahap yang menghasilkan 3 titik mutasi pada protein dapat meningkatkan perbedaan termostabilitas pada klon mutan terhadap klon normalnya (Miyazaki et al. 2006). Adapun pendapat lain tentang penelitian termostabilitas enzim xilanase yang diekspresikan dengan Escherichia coli mempunyai aktivitas xilanase yang tinggi pada tingkat fraksi periplasmitnya (Huang J et al. 2006).
12 Penelitian lainnya yang dilakukan oleh Helianti et al 2010 menghasilkan xilanase CMU yang diisolasi dari bakteri dengan genus Bacillus dan diperbanyak dengan sel kompeten Escherichia coli DH5α dengan ketahanan pH 6-7 dan suhu 55-60˚C. Berdasarkan dari uji aktivitas enzim terhadap pH dan suhu, enzim yang dihasilkan dari klon 7 memiliki ketahanan pada pH 9 dan suhu 60˚C.
SIMPULAN DAN SARAN
Simpulan
Mutasi glutamina menjadi lisina pada posisi 53 menggunakan pasangan primer (Q53K) dan threonina menjadi isoleusina pada posisi 156 menggunakan pasangan primer T194I telah berhasil dilakukan pada plasmid pGEMalkxynaq1cmu M3 dengan melalui pendekatan mutasi terarah atau PCR site directed mutagenesis dan prinsipnya mensubtitusi beberapa asam amino yang dapat mempengaruhi stabilitas enzim. Aktivitas relatif pada suhu 60 dan 65°C xilanase mutan tiga titik dari Klon 7 memiliki nilai paling tinggi di antara xilanase normal maupun mutan lainnya baik pada pH 9 maupun 10. Aktivitas relatif xilanase mutan tiga titik dari Klon 4 pada kondisi pH dan suhu tersebut memiliki aktivitas relatif yang lebih rendah baik dibandingkan dengan xilanase normal klon P39 maupun mutan ganda klon M3. Hasil pengujian stabilitas enzim pada suhu 60°C, pH 9 dan 10 mengindikasikan mutan xilanase Klon 7 relatif stabil pada pH 9.
Saran
Perlu dilakukan mutasi lebih lanjut pada mutan plasmid pGEMalkxynaq1cmu Klon 7 dengan primer mutagenik lainnya. Stabilitas klon mutan yang mengandung gen xyncmu diharapkan meningkat apabila terjadi perubahan asam amino yang bersifat polar menjadi non-polar sehingga memiliki banyak residu yang berperan dalam peningkatan stabilitas enzim.
DAFTAR PUSTAKA
Bailey MJ, Biely P, Poutanen K. 1992. Interlaboratory testing of methods for assay of xylanase activity. J Biotechnol. 23:257-270.
Balaa BA, Brijz K, Gebruers K, Vandenhaute J, Wouters J, Housen I. 2009. Xylanase XYL1p from Soytalidium acidophilum: site directed mutagenesis and acidophilic adaptation. Bioresource Technology. 100: 6465-6471. Batubara R. 2006. Teknologi bleaching ramah lingkungan [tesis]. Medan:
Program Pascasarjana, Universitas Sumatera Utara.
13
Chen H, Rangasamy M, Tan SY, Wang H, Siegfried BD. 2010. Evaluation of five methods for total DNA extraction from Western Corn Rootworm Beetles. Plos ONE 5.
Dubnovitsky AP, Kapetaniou EG, Papageorgiou AC. 2005. Enzyme adaptation to alkaline pH : atomic resolution (1.08Ả) structure of phosphoserine amino-transferase from Bacillus alcalophillus. Protein Sci 14: 97-110.
[EMBL] European Molecular Biology Laboratory. 2009. Protein purification extraction and clarification: Preparation of cell lysates from E.coli [internet]. [diunduh pada 23 September 2013]. Tersedia pada: http://www.embl.de/ pepcore/pepcore_service/protein_purification/extract_clarification/cell_lysa-tes_from_E.coli/.
Helianti et al. 2010. Constitutive high level expression of an endoxylanase gene from the newly isolated Bacillus subtilis AQ1 in Escherichia coli. Journal of Biomedical and Biotechnology. Page 1-12.
Huang J, Wang G, Xiao L. 2006. Cloning, sequencing, and expression of the xylanase gene from a Bacillus subtilis strain B10 in Escherichia coli. Bioresource Technology. 97(6): 802-806.
Inoue H, Nojima H, Okayama H. 1990. High efficiency transformation of Escherichia coli with plasmids. Gene. 96: 23-28.
Jeong MY, Kim S, Yun CW, Choi YJ, Cho S. 2007. Engineering a de novo internal disulfide bridge to improve the thermal stability of xylanase from Bacillus stearothermophilus No. 236. Journal of Biotechnology. 127: 300-309.
Lehninger AL, David LN, Michael MC. 2008. Principles of Biochemistry. New York: Academic Press.
Liu L, Zeng L, Wang S, Cheng J, Li X, Song A, Wu K, Chen H. 2012. Activity and thermostability increase of xylanase following transplantation with modules sub-divided from hyper-thermophilic CBM9 1-2. Process Biochemistry. 47: 853-857.
Meijer H, Dreesen JC, Van Boven CP. 1986. Molecular cloning and restriction endonuclease mapping of the rat cytomegalovirus genome. J Gen Virol. 67: 1327-42.
Miyazaki K, Takenouchi M, Kondo H, Noro N, Suzuki M, Truda S. 2006. Thermal stabilization of Bacillus subtilis family-11 xylanase by directed evolution. Journal of Biological Chemistry. 281(15): 10236-10242.
[MCL] Molecular Cloning Laboratories. 2010. DH5-Alpha competent Escherichia coli [internet]. [diunduh pada 23 September 2013]. Tersedia pada: www.mclab.com/DH5-Alpha-Competent-E.-Coli.html
Muawanah A. 2006. Produksi enzim xilanase termostabil dari Thermomyces lanuginosus ifo 150 [skripsi]. Bogor (ID): Institut Pertanian Bogor.
New England Biolabs, 2005. Catalog and Technical Reference. Jakarta: Gene Craft Labs.
Pratiwi, Nadia A. 2013. Peningkatan termostabilitas enzim alkalifilik xilanase dengan site directed mutagenesis [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Promega Corporation. 1997. Site-directed mutagenesis. Promega Notes Magazine. P.12
Rapley R. 2000. Recombinant DNA technology. In: Walker JM, Rapley R (eds). Molecular Biology and Biotechnology. Cambridge: The Royal Society of Chemistry.
Rifaat HM, Nagieb ZA, Ahmed YM. 2005. Productions of xylanases by Streptomyces sp. and their bleaching effect on rice straw pulp. Environ. 4: 151-160.
Schaffner W, Weissmann C. 1973. A rapid, sensitive, and specific method for the determination protein in dilute solution. Analytical Biochemistry. Vol (56): 502-514
Sriprapundh D, Vieille C, Zeikus JG. 2000. Molecular determinants of xylose isomerase thermal stability and activity: analysis of thermozymes by site-directed mutagenesis. Protein Engineering. Vol (13): 259-265.
Storici F, Resnick MA. 2006. The delitto perfetto approach to in vivo site-directed mutagenesis and chromosome rearrangements with synthetic oligonucleotides in yeast. Methods Enzymol. 409:329-345.
Ulfah M, Nurcholis M, Nurhayati N, Helianti I, Wahyuntari B, Wardani AK. 2012. Cloning of α-L-arabinofuranosidase Genes and Its Expression in Escherichia coli: A Comparative Study of Recombinant Arabinofuranosidase Originatingin Bacillus subtilis DB104 and Newly Isolated Bacillus licheniformis CW1. Microbiology Indonesia. Vol. 6 No.1 Wahyuntari B, Mubarik NR, Setyahadi S. 2009. Effect of pH, temperature and
medium composition on xylanase production by Bacillus sp. AQ-1 and partial characterization of the crude enzyme. Microbiol Indonesia Vol. 3, No. 1.
Yusriah, Nengah DK. 2013. Pengaruh pH dan suhu terhadap aktivitas protease Penicillium sp. Jurnal Sains dan Seni Pomits. Vol 2: 2337-2340.
Zhang et al. 2010. Improving the thermostability of Geobacillus stearothermophilius xylanase XT6 by directed evolution and site-directed mutagenesis. Biosource Technology. 101: 9272-78.
Zheng L, Baumann U, Reymond JL. 2004. An efficient one-step site-directed and site-saturation mutagenesis protocol. Nucleic Acids Res. 21, el15.
[WBC] Worthington Biochemical Corporation. 2013. Pectinase. WBC [Internet]. [diunduh 2013 Juli 19]. Tersedia pada : http://www.worthington-biochem.com/PASE/.
15
Lampiran 1 Strategi Penelitian
Analisa aktivitas mutan pGEMalkxynaq1cmu M3 rekombinan pada suhu 60 dan
65ºC
Analisa aktivitas mutan pGEMalkxynaq1cmu M3 rekombinan pada pH 9 dan 10
Studi thermostabilitas enzim xilanase CMU rekombinan pada pH optimal dan suhu optimal ± suhu 5ºC selama 0,
10, dan 20 menit
Mutagenesis situs terarah mutan tunggal plasmid pGEMalkxynaq1cmu M3 dengan
Phusion high-fidelity DNA polymerase (Thermo)
Pemotongan klon mutan plasmid pGEMalkynaq1cmu M3
Uji aktivitas xilanase rekombinan dengan metode Bailey Ekstraksi plasmid rekombinan
pGEMalkxynaq1cmu M3 (Minipreps)
Analisis enzim restriksi mutan plasmid pGEMalkxynaq1cmu M3
Sequencing mutan plasmid pGEMalkxyna1cmu M3 rekombinan
Transformasi mutan pGEMalkxynaq1cmu M3ke dalam E. coli DH5α kompeten
Primer mutagenik
Lampiran 2 Alignment Gen alkynaq1cmu M3 dengan program ClustalW (www.genome.jp)
Keterangan:
1. Perubahan asam amino glutamina (CAA) menjadi lisina (AAA) pada urutan asam amino posisi 53 yang disebabkan mutasi klon tetua (M3) dengan primer Q53K
2. Perubahan asam amino glutamina (CAA) menjadi leusina (CTA) pada urutan asam amino posisi 62 yang disebabkan mutasi klon tetua (M3) dengan primer Q62L
16
Lampiran 2 (lanjutan)
Keterangan:
1. Perubahan asam amino glutamat (GAG) menjadi valina (GTG) pada urutan asam amino posisi 108 yang disebabkan mutasi klon tetua (M3) dengan primer E146V
2. Perubahan asam amino treonina (ACA) menjadi isoleusina (ATA) pada urutan asam amino posisi 156 yang disebabkan mutasi klon tetua (M3) dengan primer T194I
17
1
Lampiran 3 Alignment mutasi gen pGEMalkxynaq1cmu M3 dengan klon 4 dan 7
M3
Klon 4
Klon 7
Keterangan:
Perubahan asam amino terlihat dengan menggunakan software Chromas Lite. Perubahan terlihat dari perbandingan kromatogram yang terdeteksi pada software tersebut. Perubahan asam amino terdapat di posisi 53.
Lampiran 3 (lanjutan)
M3
Klon 4
Klon 7
Keterangan:
Perubahan asam amino terlihat dengan menggunakan software Chromas Lite. Perubahan terlihat dari perbandingan kromatogram yang terdeteksi pada software tersebut. Perubahan asam amino terdapat di posisi 156.
RIWAYAT HIDUP
Penulis dilahirkan di DKI Jakarta pada tanggal 15 Juli 1991 dari ayah Dahrizal dan ibu Sri Rusiana. Penulis adalah putri kedua dari lima bersaudara. Tahum 2009 penulis lulus dari SMA Sekolah Indonesia Kuala Lumpur dan pada tahun yang sama penulis lulus seleksi masuk Institut Pertanian Bogor (IPB) melalui Ujian Talenta Mandiri IPB dan diterima di Departemen Biokimia, Fakultas Matematika dan Ilmu Pengetahuan Alam.
Selama mengikuti perkuliahan, penulis mengikuti kursus bahasa Jerman di dalam lingkungan perkuliahan dan lulus dengan nilai yang memuaskan pada tahap Einsfuhrung atau pemula. Selain bidang akademis, penulis pernah aktif dalam Himpunan Profesi CREBs (Community of Research and Education in Biochemistry) pada departemen Biokimia dan memegang divisi Wakil Bendahara umum pada tahun ajaran 2010/2011 dan mengikuti kepanitiaan (MPF) Masa Perkenalan Fakultas dan (MPD) Masa Perkenalan Departemen dengan menjabat sebagai anggota divisi MOD ( Master of Discipline) pada tahun ajaran 2011/2012. Penulis pernah mengikuti praktik lapang di Laboratorium Biotek-BPPT di Serpong, Tangerang Selatan selama 2 bulan.
Selain di bidang organisasi dan perkuliahan, penulis juga aktif dalam mengikuti cabang olahraga basket putri dan pernah mengikuti pertandingan antar fakultas dalam OMI (Olimpiade Mahasiswa IPB) pada tahun ajaran 2011/2012 dan pernah mengikuti perlombaan cabang seni drama musikal dalam acara-acara seminar kesehatan yang diadakan oleh Departemen Biokimia maupun antar departemen pada tahun 2010/2011.