POLA PERCABANGAN CABANG BATANG PRIMER DAN
KAITANNYA DENGAN ZAT PENGATUR TUMBUH
TANAMAN JARAK PAGAR (
Jatropha curcas
)
TRIMONOECIOUS
ZULFIKAR ARSA MUJAHIDILLAH
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Pola Percabangan Cabang Batang Primer dan Kaitannya dengan Zat Pengatur Tumbuh Tanaman Jarak Pagar (Jatropha curcas) Trimonoecious adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
ABSTRAK
ZULFIKAR ARSA MUJAHIDILLAH. Pola Percabangan Cabang Batang Primer dan Kaitannya dengan Zat Pengatur Tumbuh Tanaman Jarak Pagar (Jatropha curcas) Trimonoecious Dibimbing oleh TRIADIATI dan DUDI TOHIR.
Tanaman jarak pagar merupakan tanaman monoecious, yaitu bunga jantan dan bunga betina berada pada satu tanaman. Namun, jarang ditemukan jarak pagar andromonoecious (bunga jantan dan hermaprodit pada satu rangkaian bunga) dan trimonoecious (bunga jantan, betina dan hermaprodit pada satu rangkaian bunga). Tipe kelamin bunga jarak pagar diduga dipengaruhi oleh zat pengatur tumbuh (ZPT) dan berhubungan dengan karakter morfologi. Penelitian ini bertujuan menganalisis pola percabangan serta hubungannya dengan kandungan zat pengatur tumbuh sebagai penciri jenis kelamin bunga tanaman jarak pagar khususnya trimonoecious. Pengamatan karakter morfologi berupa tipe percabangan pada nodus pertama dilakukan pada umur 1 hingga 4 minggu setelah persemaian. Jumlah nodus pada 30 cm batang diamati pada umur 9 bulan setelah tanam. Tipe percabangan pada nodus pertama dan jumlah cabang primer jarak pagar trimonoecious asal bunga betina, bunga hermaprodit dan andromonoecious memiliki perbedaan dengan tipe monoecious. Perbedaan pola percabangan batang berhubungan dengan konsentrasi zat pengatur tumbuh. Pola percabangan batang dapat dijadikan penciri awal tipe jenis kelamin jarak pagar. Kata kunci: andromonoecious, jarak pagar, pola percabangan, trimonoecious, zat pengatur tumbuh
ABSTRACT
ZULFIKAR ARSA MUJAHIDILLAH. Primery Shoot Branching Pattern and Correlation with Plant Growth Regulators (PGRs) on Trimonoecious Physic nut (Jatropha curcas) Supervised by TRIADIATI and DUDI TOHIR.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biologi
POLA PERCABANGAN CABANG BATANG PRIMER DAN
KAITANNYA DENGAN ZAT PENGATUR TUMBUH
TANAMAN JARAK PAGAR (
Jatropha curcas
)
TRIMONOECIOUS
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2015
PRAKATA
Alhamdulillahirabbil’alamiin. Puji syukur kehadirat Allah SWT atas limpahan rahmat dan karunia-Nya, sehingga saya dapat menyelesaikan karya ilmiah yang berjudul Pola Percabangan Cabang Batang Primer dan Kaitannya Dengan Kandungan Zat Pengatur Tumbuh Tanaman Jarak Pagar (Jatropha curcas) Trimonoecious.
Penulis ucapkan terima kasih kepada Ibu Dr Triadiati, MSi dan Bapak Drs Dudi Tohir, MS selaku pembimbing atas pengarahan, teladan, kesabaran, waktu, dan nasehat yang diberikan kepada penulis selama penelitian hingga penulisan karya ilmiah ini. Terima kasih juga kepada Ibu Dr Dyah Perwitasari, MSc sebagai penguji wakil Departemen atas nasehat yang diberikan. Terima kasih kepada Ibu, Bapak, Adik, dan keluarga besar penulis. Tak lupa penulis ucapkan terima kasih kepada teman-teman Biologi 47, keluarga Wisma Sawit, keluarga Sabisa Farm dan teman-teman atas kerjasama, dukungan, dan semangatnya.
Semoga karya ilmiah ini dapat menambah wawasan dan pengetahuan bagi pembaca.
DAFTAR ISI
DAFTAR TABEL viii
DAFTAR GAMBAR viii
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
METODE 2
Waktu dan Tempat 2
Alat dan Bahan 2
Pengamatan Pola Percabangan Cabang Batang Primer 2
Analisis Kandungan Zat Pengatur Tumbuh 3
HASIL DAN PEMBAHASAN 3
Hasil 3
Pembahasan 6
SIMPULAN DAN SARAN 7
Simpulan 7
Saran 8
DAFTAR PUSTAKA 8
DAFTAR TABEL
1 Rata-rata jumlah nodus batang jarak pagar pada 30 cm dari ujung
pucuk. 4
DAFTAR GAMBAR
1 Pola percabangan batang jarak pagar (a) tipe trimonoecious asal bunga betina, asal bunga hermaprodit, dan tipe andromonoecious (b)
tipe monoecious. 3
2 Rata-rata konsentrasi zat pengatur tumbuh IAA jarak pagar fase vegetatif pada bulan April, Mei, dan Juni 2014 4 3 Rata-rata konsentrasi zat pengatur tumbuh kinetin jarak pagar fase
vegetatif pada bulan April, Mei, dan Juni 2014. 5 4 Rata-rata konsentrasi zat pengatur tumbuh GA3 jarak pagar fase
PENDAHULUAN
Latar Belakang
Tanaman jarak pagar (Jatropha curcas) diketahui bersifat monoecious yaitu tanaman berumah satu dengan bunga jantan dan betina terletak pada tanaman yang sama (Dellaporta dan Urrea 1993). Saat ini diketahui adanya tanaman jarak pagar tipe andromonoecious dan trimonoecious. Jarak pagar andromonoecious memiliki bunga jantan dan hermaprodit pada satu rangkaian bunga. Bunga hermaprodit memiliki serbuk sari dan putik pada satu bunga. Hal ini memberikan kemudahan saat terjadinya polinasi, sehingga besar kemungkinan terjadinya polinasi tanpa bantuan polinator. Tipe trimonoecious memiliki bunga hermaprodit, bunga jantan dan betina pada satu rangkaian bunga. Tipe tanaman andromonoecious dan trimonoecious pada jarak pagar sangat jarang ditemukan dibanding tipe monoecious.
Perbedaan ciri morfologi dapat dipengaruhi oleh perbedaan susunan gen ataupun karena faktor lingkungan. Ciri ini dapat digunakan sebagai penciri suatu individu tanaman, sehingga dapat dengan mudah membedakan antar individu satu dengan lainnya. Ciri morfologi tumbuhan termasuk karakter pola percabangan bisa digunakan untuk penanda ciri tanaman. Kurniati (2014) melaporkan bahwa adanya perbedaan karakter percabangan antara jarak pagar tipe monoecious dan tipe trimonoecious berupa jumlah cabang, ukuran batang dan diameter pangkal batang pada umur 2 hingga 6 bulan setelah tanam. Waktu yang digunakan untuk mengenali perbedaan tipe trimonoecious, monoecious dan andromonoecious paling cepat saat tanaman berumur 2 bulan setelah penanaman (Kurniati 2014). Oleh karena itu perlu analisis lebih awal perbedaan karakter yang muncul untuk mempersingkat waktu seleksi tanaman.
2
tanaman mensintesis ZPT dalam konsentrasi yang berbeda-beda. Permasalahan dari penelitian ini adalah belum ada informasi terkait hubungan pola percabangan batang dengan kandungan ZPT pada jarak pagar berdasarkan jenis kelamin bunga, khususnya pada jarak pagar trimonoecious.
Tujuan Penelitian
Menganalisis pola percabangan cabang batang primer serta hubungannya dengan kandungan Zat Pengatur Tumbuh (ZPT) sebagai penciri tipe tanaman jarak pagar (Jatropha curcas) berdasarkan jenis kelamin bunga khususnya tipe trimonoecious.
METODE
Waktu dan Tempat
Penelitian ini dilakukan pada bulan Oktober 2013-September 2014, di Rumah Kaca Departemen Biologi IPB, Kebun Penelitian Cikabayan IPB, Laboratorium Fisiologi Tumbuhan Departemen Biologi IPB dan Laboratorium Kimia Organik Departemen Kimia IPB.
Alat dan Bahan
Alat yang digunakan pada penelitian ini, diantaranya kamera, mortar, penguap putar, corong pisah dan spektrofotometer UV-Vis. Bahan yang digunakan yaitu benih tanaman jarak pagar tipe trimonoecious asal bunga betina, trimonoecious asal bunga hermaprodit, monoecious dan andromonoecious sebagai pembanding, metanol, NH4OH 2 N, kloroform, etil asetat, NaOH 7 N, dan HCL 7 N.
Pengamatan Pola Percabangan Cabang Batang Primer
3 Analisis Kandungan Zat Pengatur Tumbuh
Analisis kandungan zat pengatur tumbuh yang dilakukan berjumlah 3 individu setiap masing-masing tipe tanaman. Analisis kandungan zat pengatur tumbuh dilakukan dengan metode Unyayar et al. (1996). Sampel daun dewasa diambil seberat ± 5 g dari daun ketiga hingga kelima dan diekstrak dengan cara digerus. Sampel kemudian dimaserasi dengan 10.0 mL pelarut metanol, kloroform, dan NH4OH 2 N (12:5:3 v/v/v). Ekstrak dimasukkan dalam corong pisah, lalu di ekstraksi cair-cair dengan 22.4 mL air selama 24 jam. Fraksi air dipisahkan, dan diatur pHnya menjadi 2.5 dengan HCL 7 N, lalu diekstraksi dengan 15 mL etil asetat sebanyak 3 kali. Ekstrak didiamkan beberapa menit agar fraksi air terpisah dari fraksi etil asetat. Fraksi etil asetat diambil dan fraksi air dilanjutkan untuk perlakuan berikutnya. Fraksi etil asetat yang diperoleh diharapkan mengandung IAA dan GA3. Fraksi air diatur pH nya menjadi 7 dengan NaOH 7 N kemudian diekstraksi dengan 15 mL etil asetat 3 kali ulangan. Perlakuan ini akan menghasilkan kinetin bebas. Ekstrak IAA, GA3, dan kinetin yang diperoleh selanjutnya dievaporasi. Ekstrak kering yang diperoleh kemudian dilarutkan dengan 10 mL metanol kemudian diukur absorbannya pada panjang gelombang 280, 263, dan 269 nm masing-masing untuk IAA, GA3, dan kinetin. Konsentrasi sampel yang diperoleh dihitung dengan persamaan kurva standar. Analisis dilakukan pada bulan April, Mei, dan Juni 2014.
HASIL DAN PEMBAHASAN
Hasil
Hasil pengamatan pola percabangan cabang batang primer jarak pagar tipe trimonoecious asal bunga betina, asal bunga hermaprodit, monoecious dan andromonoecious, menunjukkan adanya perbedaan pada jenis percabangan yang muncul dari nodus pertama pada waktu 1 bulan setelah semai (Gambar 1).
4
Jarak pagar tipe trimonoecious asal bunga hermaprodit dan trimonoecious asal bunga betina memiliki kesamaan dengan tipe andromonoecious pada tipe percabangan yang muncul pada nodus pertama tanaman. Cabang primer pertama (No. 1) muncul berhadapan pada nodus pertama pada umur dua minggu setelah semai. Selanjutnya pada nodus tersebut muncul cabang primer selanjutnya (No. 4) pada umur empat minggu setelah semai, sehingga pada satu nodus berjumlah empat cabang. Berbeda halnya dengan monoecious, cabang pertama (No. 1) muncul berhadapan pada umur dua minggu setelah semai namun setelah itu tidak diikuti munculnya percabangan baru pada nodus tersebut. Hal ini dapat dijadikan ciri pembeda untuk menseleksi tipe jarak pagar pada waktu lebih awal. Karakter berbeda juga terlihat pada perbedaan jumlah nodus pada 30 cm batang dari pucuk (Tabel 1).
Tabel 1 Rata-rata jumlah nodus batang jarak pagar pada 30 cm dari ujung pucuk. Nama Tipe Jarak pagar Jumlah
Tanaman tipe trimonoecious asal bunga betina, asal bunga hermaprodit, dan andromonoecious berturut-turut sebanyak 55, 56 dan 56 nodus. Sedangkan jarak pagar tipe monoecious hanya memiliki nodus berjumlah 42 nodus. Perbedaan tersebut berhubungan dengan konsentrasi zat pengatur tumbuh tanaman.
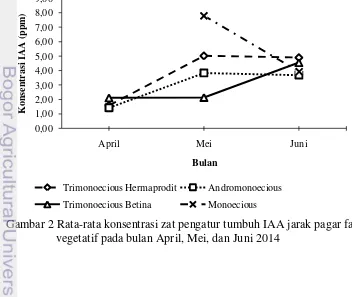
Gambar 2 Rata-rata konsentrasi zat pengatur tumbuh IAA jarak pagar fase vegetatif pada bulan April, Mei, dan Juni 2014
5
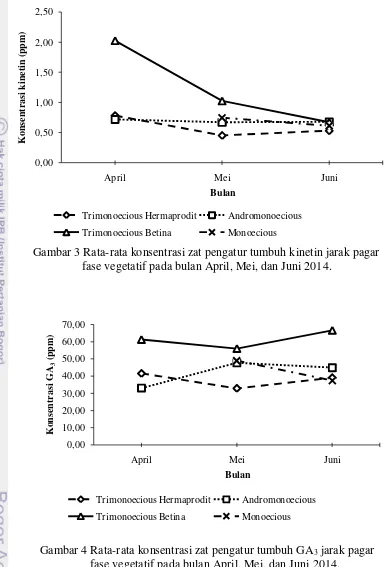
Gambar 3 Rata-rata konsentrasi zat pengatur tumbuh kinetin jarak pagar fase vegetatif pada bulan April, Mei, dan Juni 2014.
Gambar 4 Rata-rata konsentrasi zat pengatur tumbuh GA3 jarak pagar fase vegetatif pada bulan April, Mei, dan Juni 2014.
Kandungan hormon IAA dan kinetin tipe andromonoecious pada bulan Juni sebesar 3.68 ppm dan 0.68 ppm (Gambar 2 dan 3). Konsentrasi hormon GA3 tipe andromonoecious pada bulan Juni terlihat cukup tinggi sebesar 44.91 ppm (Gambar 4). Kandungan IAA, kinetin dan GA3 pada bulan Juni untuk trimonoecious asal bunga betina dan hermaprodit terlihat tinggi. IAA, kinetin, dan GA3 trimonoecious asal bunga betina pada bulan Juni berturut-turut sebesar 4.55
6
ppm, 0.67 ppm, dan 66.10 ppm, sedangkan untuk trimonoecious asal bunga hermaprodit sebesar 4.90 ppm, 0.53 ppm dan 39.04 ppm. Kandungan hormon kinetin pada bulan Juni tipe monoecious cenderung lebih rendah daripada tipe andromonoecious, yaitu sebesar 0.61 ppm, IAA yang cukup tinggi sebesar 3.92 ppm dan GA3 paling rendah dari keseluruhan tipe sebesar 37.37 ppm.
Pembahasan
Pola percabangan yang muncul pada nodus pertama tanaman tipe trimonoecious asal bunga betina dan asal bunga hermaprodit adalah sama dengan tipe andromonoecious, namun memiliki perbedaan dengan monoecious. Pola percabangan yang terbentuk berupa munculnya cabang berhadapan pada nodus pertama dan diikuti cabang berikutnya muncul pada nodus yang sama. Menurut Bangerth (1994) rasio antara IAA dan kinetin akan mempengaruhi pola percabangan suatu tanaman. Rendahnya kandungan kinetin dan IAA yang tinggi dapat menyebabkan terhambatnya pertumbuhan cabang pada nodus tanaman, sehingga jumlah cabang rendah (Darmanti 2008). Hal ini dapat diasumsikan bahwa kandungan kinetin dan IAA yang disintesis berbeda akan memunculkan karakter morfologi yang berbeda.
Kandungan IAA dan kinetin juga berpengaruh terhadap jumlah nodus pada 30 cm dari pucuk batang. Jumlah nodus juga berpengaruh terhadap jarak antar nodus. Keseimbangan konsentrasi kinetin tinggi dan IAA rendah akan memacu terjadinya pembelahan dan diferensiasi sel pada nodus untuk membentuk primordia cabang lateral (Darmanti 2008). Jarak antar nodus dipengaruhi oleh hormon IAA dan GA3 yang menyebabkan pemanjangan sel. Menurut Davenport et al. (2000), GA3 dapat mempengaruhi aktivitas IAA endogen serta dapat mempengaruhi pemanjangan batang. Penambahan hormon IAA yang digabungkan dengan GA3 efektif meningkatkan pemanjangan batang pada tanaman (Naeem et al. 2004).
Pola percabangan jarak pagar trimonecious asal bunga betina dan bunga hermaprodit adalah sama dengan pola andromonoecious. Diduga, pola percabangan berkaitan dengan keberadaan bunga hermaprodit yang dimiliki oleh setiap tipe tersebut. Pada trimonoecious asal bunga betina kemungkinan membawa sifat yang berasal dari bunga hermaprodit karena bunga betina dan bunga hermaprodit berasal dari tanaman induk yang sama. Selain faktor tersebut, diduga sintesis hormon kinetin yang lebih tinggi menyebabkan munculnya cabang lanjutan pada nodus yang sama pada tipe andromonoecious, trimonoecious asal bunga betina dan asal bunga hermaprodit. Hal ini terlihat dari hasil analisis kinetin tipe andromonoecious dan trimonoecious asal bunga betina yang lebih tinggi dari monoecious. Kandungan hormon kinetin dan rendahnya konsentrasi IAA pada tipe andromonoecious dibandingkan dengan tipe monoecious menyebabkan jumlah cabang yang lebih banyak. Menurut Dasumiati (2014) bahwa jarak pagar tipe andromonoecious memiliki jumlah cabang batang primer yang banyak serta jarak antar nodus yang dekat.
7 (2001) mengemukakan bahwa kinetin akan merangsang pembelahan sel pada tanaman dan sel-sel yang membelah tersebut akan berkembang menjadi tunas, cabang, dan daun. Lebih jauh Sato et al. (2009) menjelaskan bahwa kandungan IAA yang rendah dapat merangsang protein IPT terekspresi sehingga terjadi biosintesis kinetin pada batang. Kinetin kemudian di transportasikan menuju daerah perkembangan tunas aksilar, sehingga tunas aksilar mulai tumbuh.
Pada tipe trimonoecious asal bunga betina dan hermaprodit, kandungan IAA dan kinetin kedua tipe ini terlihat tinggi sedangkan percabangan yang dicirikan oleh jumlah nodus adalah sama dengan andromonoecious. Hal ini mengindikasikan bahwa hormon tidak berpengaruh terhadap percabangan kedua tipe tersebut. Percabangan yang terbentuk diduga dipengaruhi oleh kesediaan unsur hara tanaman. Kesediaan unsur hara pada tanaman akan mempengaruhi pertumbuhan cabang lateral tanaman. Tanaman dapat mengkonsentrasikan energi dan nutrisi pada salah satu pertumbuhan saja, seperti tinggi atau percabangan (Hidayati 2009).
Pola percabangan monoecious diduga karena adanya pengaruh dari hormon IAA yang lebih tinggi dan kinetin yang rendah. Dugaan tersebut didasarkan pada hasil analisis kinetin tipe monoecious yang cenderung lebih rendah dan IAA yang lebih tinggi dibanding tipe andromonoecious. IAA yang disintesis pada ujung batang ini akan ditranspor secara basipetal ke bagian batang bagian basal (Hidayati 2009). Hal ini menyebabkan terakumulasinya IAA pada ketiak daun dibawahnya yang berakibat inisiasi pembentukan tunas lateral pada ketiak daun terhambat. Menurut Sato et al. (2009) transpor IAA secara basipetal menyebabkan pertumbuhan tunas aksilar terhenti (dorman) karena IAA dapat menghambat kerja protein adenosin fosfat Isopenteniltransferase (IPT) sebagai pengontrol sintesis hormon kinetin.
Pengaruh kandungan IAA dan kinetin juga terlihat pada jumlah cabang yang dihasilkan tipe ini paling rendah dibanding tipe lainnya. Namun, pengaruh dari GA3 tidak terlihat, karena jumlah cabang yang rendah akan menyebabkan jarak antar nodus yang tinggi sehingga hormon GA3 seharusnya tinggi pada monoecious. Menurut Lakitan (1996) fungsi fitohormon bergantung pada jenis tanaman, musim yang berbeda, interaksi dengan metabolit lain, dan sensitivitas jaringan terhadap fitohormon yang bersangkutan.
Selain itu juga pengaruh hormon GA3 tidak berpengaruh terhadap jumlah percabangan setiap tipe tanaman. Hal ini diduga karena GA3 memiliki efektifitas yang rendah terhadap pemanjangan sel pada nodus batang jarak pagar. Menurut Srivastava (2002) bahwa aksi hormon GA3 berbeda di setiap tanaman bergantung jenis GA3 yang aktif. Selain itu faktor lingkungan seperti suhu dan intensitas cahaya sangat berpengaruh terhadap respon tanaman terhadap GA3.
SIMPULAN DAN SARAN
Simpulan
8
andromonoecious, yaitu terbentuknya 4 cabang lateral primer pada nodus pertama pada umur 1 bulan setelah semai, dan jumlah nodus pada 30 cm batang. Karakter tersebut berbeda dengan tipe monoecious yang hanya membentuk 2 cabang lateral primer pada nodus pertama tanaman, dan jumlah nodus pada 30 cm batang yang lebih sedikit dari ketiga tipe lainnya. Karakter yang muncul dipengaruhi oleh kandungan zat pengatur tumbuh. Pola percabangan batang yang berbeda dapat dijadikan ciri pembeda antara tanaman jarak pagar trimonoecious asal bunga betina, asal bunga hermaprodit, dan andromonoecious dengan tipe monoecious.
Saran
Penelitian terkait jarak pagar perlu dilakukan lebih lanjut terlebih pada tingkat molekular. Penelitian tingkat molekular akan memberikan informasi yang lebih pasti guna menyelidiki perbedaan setiap tipe jarak pagar.
DAFTAR PUSTAKA
Bangerth F. 1994. Response of cytokinin concentration in the xylem exudate of bean (Phaseolus vulgaris L) plants to description and auxin treatment and relationship to apical dominance. Planta 194: 439-442.
Darmanti S. 2008. Pembentukan cabang lateral jarak pagar (Jatropha curcas) setelah perlakuan girdling. BIOMA 10 (1): 7-11.
Dasumiati. 2014. Karakterisasi dan peningkatan jumlah bunga hermaprodit tanaman Jarak Pagar (Jatropha curcas) andromonoecious. [disertasi]. Bogor: Sekolah Pascasarjana, IPB.
Davenport TL, Pearce DW, Rood SB. 2000. Correlation of endogenous gibberellic acid with initiation of mango shoot growth. JPGR 19 (4): 445-452. doi: 10.1007/s003440000029.
Dellaporta SL, Urrea AC. 1993. Sex determination in flowering plants. The Plant Cell. 5: 1241-1251.
Hidayati Y. 2009. Kadar hormon IAA pada tanaman kenaf (Hibiscus cannabicus L.) bercabang dan tidak bercabang. AGROVIGOR 2 (2): 89-96.
Kurniati. 2014. Karakter biologi jarak pagar (Jatropha curcas L .) Trimonoecious. [tesis]. Bogor : Fakultas Matematika dan Ilmu Pengetahuan Alam, IPB. Komariah S. 2012. Kandungan zat pengatur tumbuh daun dan pola infloresen
bunga pada jarak pagar (Jatropha curcas L.) andromonoecious.[skripsi].
Bogor : Fakultas Matematika dan Ilmu Pengetahuan Alam, IPB.
Lakitan, B. 1996. Fisiologi Pertumbuhan dan Perkembangan Tanaman. Jakarta(ID): PT Raja Grafindo Perkasa.
Naeem M, Bhatti I, Ahmad RH, Ashraf MY. Effect of some growth hormones (GA3, IAA and kinetin) on the morphology and early or delayed initiatin of bud of lentil (Lens culinaris medik). Pak J Bot. 36(4): 801-809.
9 Salisbury FB, Ross CW. 1995. Fisiologi Tumbuhan Jilid 3. Diah RL, Sumaryono,
penerjemah. Bandung (ID): ITB Pr. Terjemahan dari: Plant Physiology. Sato SS, Tanaka M, Mori H. 2009. Auxin–cytokinin interactions in the control of
shoot branching. Plant Mol Biol 69: 429–435. doi: 10.1007/s11103-008-9416-3.
Srivastava LM. 2002. Plant Growth and Development : Hormones and Environment. California (US): Academic Pr.
Takei K, Sakakibara H, Sugiyama T. 2001. Identification of genes encoding adenylate isopentenyltransferase, a cytokinin biosynthesis enzyme, in Arabidopsis thaliana. J Biol Chem 276 (28): 26405-26410. doi: 10.1074/jbc.M102130200.
10
RIWAYAT HIDUP
Penulis dilahirkan di Sukabumi pada tanggal 14 Pebruari 1993 dari ayah Saepulloh dan ibu Ai Rohyati. Penulis merupakan anak pertama dari tiga bersaudara. Tahun 2010 penulis lulus dari SMA Negeri 3 Sukabumi. Pada tahun yang sama, penulis lulus seleksi masuk IPB melalui jalur Seleksi Nasional Masuk Perguruan Tinggi Negeri (SNMPTN) di Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam. Penulis mendapatkan beasiswa Bantuan Belajar Mahasiswa (BBM) dari DIKTI pada tahun 2012-2014.
Pada tahun 2012 penulis aktif sebagai anggota divisi Pengembangan Sumberdaya Mahasiswa (PSDM) Himpunan Mahasiswa Biologi IPB dan pada tahun 2013 menjadi ketua di divisi yang sama.Penulis aktif mengikuti beberapa kegiatan dalam masa studi seperti menjadi peserta kompetisi olimpiade biologi pada OSN Pertamina (2012), serta aktif kegiatan Organisasi Mahasiswa Daerah (OMDA) Sukabumi. Penulis juga aktif mengikuti lomba Futsal dan basket SPIRIT FMIPA IPB (2012-2013). Penulis juga menjadi ketua kegiatan olahraga dan seni Grand Biodiversity Biologi (2012) dan pernah menjadi ketua acara Masa Perkenalan
Departemen Biologi “MORFOLOGI” tahun 2013, serta beberapa kepanitiaan lainnya.
Pada Tahun 2013-2015 penulis aktif sebagai pengelola di Perkebunan Buah Naga SABISA Farm IPB.