PENENTUAN pH DAN SUHU OPTIMUM UNTUK AKTIVITAS
EKSTRAK KASAR ENZIM LIPASE DARI KECAMBAH BIJI
KELAPA SAWIT (Elaeis guineensis Jacq) TERHADAP
HIDROLISIS RBDPO (Refined Bleached
Deodorized Palm Oil)
SKRIPSI
RIZQI AISYAH AMATURRAHIM
080802026
DEPARTEMEN KIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN
ALAM
UNIVERSITAS SUMATERA UTARA
MEDAN
PENENTUAN pH DAN SUHU OPTIMUM UNTUK AKTIVITAS
EKSTRAK KASAR ENZIM LIPASE DARI KECAMBAH BIJI
KELAPA SAWIT (Elaeis guineensis Jacq) TERHADAP
HIDROLISIS RBDPO (Refined Bleached
Deodorized Palm Oil)
SKRIPSI
Diajukan untuk melengkapi tugas dan memenuhi syarat mencapai gelar Sarjana Sains
RIZQI AISYAH AMATURRAHIM
080802026
DEPARTEMEN KIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN
ALAM
UNIVERSITAS SUMATERA UTARA
MEDAN
PERSETUJUAN
Judul : Penentuan pH dan Suhu Optimum untuk Aktivitas Ekstrak Kasar Enzim Lipase dari Kecambah Biji Kelapa Sawit (Elaeis guineensis Jacq) terhadap Hidrolisis RBDPO (Refined Bleached Deodorized Palm Oil)
Kategori : Skripsi
Nama : Rizqi Aisyah Amaturrahim Nomor Induk Mahasiswa : 080802026
Program Studi : Sarjana (S1) Kimia Departemen : Kimia
Fakultas : Matematika dan Ilmu Pengetahuan Alam Universitas Sumatera Utara
Disetujui di
Medan, Oktober 2012
Komisi Pembimbing :
Pembimbing 2 Pembimbing 1
Drs.Firman Sebayang,MS Dr.Yuniarti Yusak,MS NIP. 195607261985031001 NIP. 194901271980022001
Diketahui/Disetujui Oleh
Departemen Kimia FMIPA USU Ketua,
PERNYATAAN
PENENTUAN pH DAN SUHU OPTIMUM UNTUK AKTIVITAS
EKSTRAK KASAR ENZIM LIPASE DARI KECAMBAH BIJI
KELAPA SAWIT (Elaeis guineensis Jacq) TERHADAP
HIDROLISIS RBDPO (Refined Bleached
Deodorized Palm Oil)
SKRIPSI
Saya mengakui bahwa skripsi ini adalah hasil kerja saya sendiri, kecuali beberapa kutipan dari ringkasan yang masing-masing disebutkan sumbernya.
Medan, Oktober 2012
PENGHARGAAN
Alhamdulillah,,Puji dan Syukur Penulis ucapkan atas kehadirat ALLAH SWT yang telah memberikan Rahmat dan Hidayah-Nya sehingga Penulis dapat menyelesaikan skripsi ini. Pada kesempatan ini, Penulis mengucapkan terima kasih dan penghargaan kepada orangtua penulis Bapak Muslihuddin dan Ibu Siti Minawarsa yang telah membesarkan dengan kasih sayang, mendidik, memberikan segalanya untuk Penulis dan untuk adik-adikku Suci Aisyah Amaturrahim, Boy Attaurazaq, Irgie Attaurazaq yang telah memberikan semangat untuk menyelesaikan skripsi ini.
Penulis mengucapkan terimakasih kepada Ibu Dr.Yuniarti Yusak, MS selaku Dosen Pembimbing I, dan Bapak Drs.Firman Sebayang, MS selaku Dosen Pembimbing II dan Bapak Drs. Johannes H.Simorangkir, MS selaku Dosen Penasehat Akademik yang telah sabar dalam membimbing dan memberikan arahan kepada Penulis selama penelitian dan dalam menyelesaikan skripsi ini. Ketua Departemen Kimia Ibu DR.Rumondang Bulan Nasution dan Sekretaris Departemen Kimia Bapak Drs.Albert Pasaribu, M.Sc dan juga kepada Seluruh Dosen Departemen Kimia F-MIPA USU yang telah memberikan ilmu dan pengetahuan kepada Penulis.
Penulis juga mengucapkan terima kasih kepada sahabat Penulis Ghina Fadhilla SP, Ratna, Putri, Linda, Dhea, Rizki Amalia Nst, Fenny, Emi, dan teman-teman kimia stambuk 08 yang tidak bisa disebutkan satu persatu dan kepada kak Fitri, kak Riah, kak Mariana, kak Lisma, kak Fia yang telah banyak membantu Penulis dalam melakukan penelitian dan menyelesaikan skripsi ini.
Akhirnya Penulis menyadari bahwa skripsi ini masih belum sempurna, oleh karena itu Penulis mengharapkan saran dan masukan yang bersifat membangun dari semua pihak demi terciptanya kesempurnaan dari skripsi ini, semoga Allah Subhanallahu Wataa’la membalas semua kebaikan kita dan memberikan kebahagian bagi kita semua. Amin Ya Rabbal Alamin.
ABSTRAK
Telah dilakukan penentuan pH dan suhu optimum untuk aktivitas crude enzim lipase dari kecambah biji kelapa sawit (Elaeis guineensis Jacq). Kecambah biji kelapa sawit dibuat melalui proses pemisahan biji dari buah kelapa sawit, perendaman, pengeringan dan pemanasan, dan perkecambahan biji dilakukan pada suhu 39 – 40oC selama 60 hari, 30ºC selama 21 hari dan 25ºC selama 14 hari Ekstrak kasar enzim lipase diperoleh melalui dua kali proses sentrifugasi selama 30 menit dengan kecepatan putaran 5000 rpm dan 10000 rpm dan dengan penambahan (NH4)2SO4.
PENENTUAN pH DAN SUHU OPTIMUM UNTUK AKTIVITAS EKSTRAK KASAR ENZIM LIPASE DARI KECAMBAH BIJI KELAPA SAWIT (Elaeis
guineensis Jacq) TERHADAP HIDROLISIS RBDPO (Refined Bleached Deodorized Palm Oil)
ABSTRACT
Determination of optimum pH and temperature for crude lipase enzyme activity from oil palm seeds germination had been done. Oil palm germination made by separating seed from palm coconut soaking time process, dried, heated and seed germination in temperature 39-40ºC during 60 days, 30ºC during 21 days and 250C during 14 days. Crude lipase enzyme was obtained by centrifugations during 30 minutes with the speed of rotation 5000 rpm and 10000 rpm, and addition of (NH4)2SO4. Crude lipase enzyme
diluted by buffer phosphate pH 7.0. The activity test of crude lipase enzyme doing by measurement of free fatty acids levels obtained from hydrolysis process of RBDPO (Refined Bleached Deodorized Palm Oil) as substrate by titrimetric methods with temperature variations 300; 350; 400; 450; 500C and pH variations 6.0; 6.5; 7.0; 7.5 and 8.0. In hydrolysis process of refined bleached deodorized palm oil obtained the optimum pH and temperature by crude lipase enzymes were 7.0 and 40ºC with the highest activity
DAFTAR ISI
2.2. Perkecambahan Biji Kelapa Sawit 7
3.2. Bahan – bahan 24 3.3.6. Penentuan pH Optimum untuk Aktivitas Ekstrak Kasar
DAFTAR TABEL
Tabel 2.1. Komposisi Asam Lemak Minyak Sawit 10
Tabel 3.1. Pembuatan Larutan Buffer Fosfat pH 6,0-8,0 25 Tabel 4.1. Hasil Perhitungan Aktivitas Crude Enzim Lipase pada Suhu 30-50o
DAFTAR GAMBAR
Gambar 2.1. Buah Kelapa Sawit 8
Gambar 2.2. Kecambah Biji Kelapa Sawit 9
Gambar 2.3. Reaksi Hidrolisis Trigliserida oleh Enzim Lipase 35 Gambar 4.7. Grafik Penentuan Suhu Optimum untuk Aktivitas Ekstrak Kasar
Enzim Lipase terhadap Hidrolisis RBDPO 36 Gambar 4.8. Grafik Penentuan pH Optimum untuk Aktivitas Ekstrak Kasar
DAFTAR LAMPIRAN
Lampiran 1. Perhitungan Pembuatan Buffer Fosfat 0,05 M 40 Lampiran 2. Perhitungan Kadar Asam Lemak Bebas (ALB)
Hasil Hidrolisis RBDPO 41
ABSTRAK
Telah dilakukan penentuan pH dan suhu optimum untuk aktivitas crude enzim lipase dari kecambah biji kelapa sawit (Elaeis guineensis Jacq). Kecambah biji kelapa sawit dibuat melalui proses pemisahan biji dari buah kelapa sawit, perendaman, pengeringan dan pemanasan, dan perkecambahan biji dilakukan pada suhu 39 – 40oC selama 60 hari, 30ºC selama 21 hari dan 25ºC selama 14 hari Ekstrak kasar enzim lipase diperoleh melalui dua kali proses sentrifugasi selama 30 menit dengan kecepatan putaran 5000 rpm dan 10000 rpm dan dengan penambahan (NH4)2SO4.
PENENTUAN pH DAN SUHU OPTIMUM UNTUK AKTIVITAS EKSTRAK KASAR ENZIM LIPASE DARI KECAMBAH BIJI KELAPA SAWIT (Elaeis
guineensis Jacq) TERHADAP HIDROLISIS RBDPO (Refined Bleached Deodorized Palm Oil)
ABSTRACT
Determination of optimum pH and temperature for crude lipase enzyme activity from oil palm seeds germination had been done. Oil palm germination made by separating seed from palm coconut soaking time process, dried, heated and seed germination in temperature 39-40ºC during 60 days, 30ºC during 21 days and 250C during 14 days. Crude lipase enzyme was obtained by centrifugations during 30 minutes with the speed of rotation 5000 rpm and 10000 rpm, and addition of (NH4)2SO4. Crude lipase enzyme
diluted by buffer phosphate pH 7.0. The activity test of crude lipase enzyme doing by measurement of free fatty acids levels obtained from hydrolysis process of RBDPO (Refined Bleached Deodorized Palm Oil) as substrate by titrimetric methods with temperature variations 300; 350; 400; 450; 500C and pH variations 6.0; 6.5; 7.0; 7.5 and 8.0. In hydrolysis process of refined bleached deodorized palm oil obtained the optimum pH and temperature by crude lipase enzymes were 7.0 and 40ºC with the highest activity
BAB 1
PENDAHULUAN
1.1. Latar Belakang
Enzim lipase termostabil atau asilgliserol hidrolase (E.C 3.1.1.3) merupakan enzim
yang dapat menghidrolisis rantai panjang trigliserida. Enzim ini banyak digunakan
pada produksi asam lemak. (Macrae. A .R, 1983). Asam lemak dan gliserol
merupakan produk oleokimia dasar yang sangat diperlukan oleh industri cat, plastik,
detergen, dan sabun. Dewasa ini proses tersebut beroperasi pada suhu 240-2500C dan
tekanan 45-50 atm. Pada proses ini diperlukan energi yang cukup besar untuk
mempertahankan kondisi operasinya dan juga asam lemak yang dihasilkan umumnya
berwarna coklat yang akan mengakibatkan rusaknya komponen-komponen yang
terkandung di dalam minyak, misalnya ᵝ-karoten. (Herawan,T., 1996).
Proses hidrolisis dengan menggunakan enzim lipase, pada umumnya reaksi
beroperasi pada suhu yang relatif rendah yaitu antara 30-600C dan tekanan
atmosferik, sehingga aman bagi lingkungan kerja dan tidak memerlukan energi yang
cukup besar. Disamping itu, produk yang dihasilkan mempunyai kualitas yang relatif
lebih baik dibandingkan produk sejenis yang dibuat dengan proses kimia atau fisika,
karena relatif tidak terjadi kerusakan akibat pemanasan pada suhu tinggi. Sebagai
biokatalis, enzim memiliki sifat-sifat antara lain dapat aktif dalam jumlah yang sangat
kecil dan aktivitas katalitiknya spesifik. (Sri, W.M., 2011)
Enzim lipase dapat diperoleh dari mikroba lokal seperti Pseudomonas,
Aspergillus niger, Mucor miehei, Candida rugosa.(Moentamaria,D., 2009), dan juga
dapat diisolasi dari kecambah biji – bijian, seperti kecambah biji wijen (Sesamun
Indicum).(Arbianti,2008), kecambah biji jarak pagar (Jatropha curcas L).(Abigor,
2002), biji kakao (Theobroma cacao L.).(Permana,M., 2012), dan jarak kepyar
Kelapa sawit berasal dari Afrika Barat, dan ternyata cocok dikembangkan di luar
daerah asalnya, termasuk Indonesia dimana kelapa sawit dapat tumbuh dan
berkembang dengan baik dan paling banyak ditemukan di Sumatera Utara dan
Aceh.(Swadaya,P., 2001). Menurut data Statistik Perkebunan Indonesia 2009-2011,
produksi kelapa sawit di Sumatera Utara tahun 2010 adalah 3.230.488 ton
Ejedegba,dkk (2007) mengkarakterisasi lipase yang diisolasi dari kelapa
(Cocos nucifera linn). Kemudian juga Arbianti,dkk (2008) juga melakukan
pemanfaatan biji wijen sebagai sumber enzim lipase untuk reaksi esterifikasi
gliserol-asam laurat. Chusnul Hidayat,dkk (2008) melakukan penelitian mengenai optimasi
produksi lipase biji kacang tanah (Arachis hypogaea,L) sebagai biokatalis dengan
metode Response Surface Methodology. Enujiugha,dkk (2009) melaporkan bahwa
lipase pada biji kacang minyak African (Pentaclethra macrophylla Benth)
mempunyai aktivitas lebih spesifik terhadap minyak laurat (kandungan asam lemak
rantai pendek).
Berdasarkan uraian diatas, peneliti ingin mengisolasi crude enzim lipase dari
kecambah biji kelapa sawit (Elaeis guineensis Jacq) dan ingin mengetahui pH dan
suhu optimum untuk menghidrolisis RBDPO (Refined Bleached Deodorized Palm
Oil).
1.2. Permasalahan
1. Bagaimana cara mengisolasi crude enzim lipase dari kecambah biji kelapa
sawit (Elaeis guineensis Jacq) ?
2. Bagaimana pengaruh suhu dan pH optimum terhadap aktivitas crude enzim
lipase dari kecambah biji kelapa sawit (Elaeis guineensis Jacq) dalam
1.3. Pembatasan Masalah
Dalam penelitian ini masalah dibatasi sebagai berikut:
1. Kecambah biji kelapa sawit yang digunakan diperoleh dari Pusat Penelitian
Kelapa Sawit (PPKS) varietas Deli Dura x Pisifera Yangambi.
2. Crude enzim lipase diambil dari kecambah yang telah berumur 14 hari.
3. Substrat yang digunakan adalah RBDPO (Refined Bleached Deodorized Palm
Oil).
4. Buffer yang digunakan adalah buffer fosfat dengan variasi pH 6,0; 6,5; 7,0;
7,5 dan 8,0.
5. Variasi suhu pemanasan yang digunakan adalah 300; 350; 400; 450; 500
6. Waktu pemanasan yang digunakan adalah 60 menit.
C.
7. Pengujian aktivitas crude enzim lipase dilakukan dengan metode titrimetri.
1.4. Tujuan Penelitian
Adapun tujuan dari penelitian ini adalah :
1. Untuk mengisolasi crude enzim lipase dari kecambah biji kelapa sawit (Elaeis
guineensis Jacq).
2. Untuk mengetahui suhu dan pH optimum crude enzim lipase dari kecambah
biji kelapa sawit (Elaeis guineensis Jacq) dalam menghidrolisis RBDPO
(Refined Bleached Deodorized Palm Oil) oleh crude enzim lipase.
1.5. Manfaat Penelitian
1. Untuk mendapatkan crude enzim lipase dari kecambah biji kelapa sawit
(Elaeis guineensis Jacq).
2. Untuk mengetahui aktivitas crude enzim lipase yang dihasilkan dari kecambah
3. Sebagai sumber informasi mengenai aktivitas crude enzim lipase dari
kecambah biji kelapa sawit (Elaeis guineensis Jacq) dan juga sebagai bahan
informasi untuk penelitian selanjutnya.
1.6. Metodologi Penelitian
Penelitian ini adalah eksperimen yang dilakukan di laboratorium, yang meliputi :
1. Penyediaan kecambah biji kelapa sawit diperoleh dari Pusat Penelitian Kelapa
Sawit (PPKS) di Jalan Brigjend Katamso,Medan.
2. Pengamatan perkecambahan biji kelapa sawit selama 14 hari
3. Penyediaan crude enzim lipase dari kecambah biji kelapa sawit.
4. Penentuan aktivitas crude enzim lipase dengan pH dan suhu pemanasan yang
berbeda.
1.7. Lokasi Penelitian
Penelitian ini dilakukan di Laboratorium Biokimia FMIPA-USU, Laboratorium
Penelitian Fakultas Farmasi-USU, Laboratorium Kimia Dasar F-MIPA USU dan
BAB 2
TINJAUAN PUSTAKA
2.1 PERKECAMBAHAN
Perkecambahan (germination) merupakan tahap awal perkembangan suatu tumbuhan,
khususnya tumbuhan berbiji. Dalam tahap ini, embrio di dalam biji yang semula
berada pada kondisi dorman mengalami sejumlah perubahan fisiologis yang
menyebabkan ia berkembang menjadi tumbuhan muda. Tumbuhan muda ini dikenal
sebagai kecambah. Kecambah adalah tumbuhan (sporofit) muda yang baru saja
berkembang dari tahap embrionik di dalam biji. Tahap perkembangan ini disebut
perkecambahan dan merupakan satu tahap kritis dalam kehidupan tumbuhan.
Kecambah biasanya dibagi menjadi tiga bagian utama: radikula (akar embrio),
hipokotil, dan kotiledon (daun lembaga). Proses perkecambahan benih merupakan
suatu rangkaian kompleks dari perubahan-perubahan morfologi, fisiologi, dan
biokimia.
Tahap pertama suatu perkecambahan benih dimulai dengan proses penyerapan
air oleh benih, melunaknya kulit benih dan hidrasi dari protoplasma. Tahap kedua
dimulai dengan kegiatan-kegiatan sel dan enzim-enzim serta naiknya tingkat respirasi
benih tahap ketiga merupakan tahap dimana terjadi penguraian bahan-bahan seperti
karbohidrat, lemak dan protein menjadi bentuk-bentuk yang melarut dan
ditranslokasikan ke titik-titik tumbuh. Tahap keempat adalah asimililasi dari
bahan-bahan yang telah diuraikan tadi di daerah meristematik untuk menghasilkan energi
baru, pembentukan komponen dan pertumbuhan sel baru. Tahap kelima adalah
pertumbuhan dari kecambah melalui proses pembelahan, pembesaran dan pembagian
Sementara penyerapan air oleh benih terjadi pada tahap pertama biasanya
berlangsung sampai jaringan mempunyai kandungan air 40 – 60 % (atau 67 – 150 %
atas dasar berat kering). Dan akan meningkat lagi pada saat munculnya radikula
sampai jaringan penyimpanan dan kecambah yang sedang tumbuh mempunyai
kandungan air 70 - 90 %.(Sutopo,L., 2002)
Ada sedikitnya tanaman Angiospermae yang dimana terjadi proses
perkembangan zigot menjadi tanaman dewasa secara terus menerus. Perkecambahan
atau pertumbuhan terbuka dari embrio biji dapat terjadi setelah periode dormansi.
Bagaimanapun, sebelum perkecambahan terjadi, kondisi eksternal harus disesuaikan.
Hal yang paling penting adalah kelembapan, oksigen dan suhu.
Kelembapan harus memadai yang secara relatif dibutuhkan sebagai tahap awal
dari perkecambahan. Air membantu lapisan biji dan memfasilitasi pergerakan oksigen
ke dalam biji sehingga air merupakan media dimana material berpindah dari satu
bagian biji ke bagian lainnya yang dibutuhkan tumbuhan seperti pencernaan makanan
dan pernafasan. Jika kecukupan kuantitas oksigen tidak terpenuhi, respirasi akan
dikurangi dan energi yang diperlukan untuk menumbuhkan embrio berkurang. Jarak
temperatur untuk perkecambahan bervariasi, namun perkecambahan biji yang terbaik
terjadi pada suhu 650 F sampai 830 F. (Johnson,W.H., 1995).
Dormansi adalah masa istirahat, artinya kemampuan biji untuk menangguhkan
perkecambahannya sampai pada saat dan tempat yang mengguntungkan baginya
untuk tumbuh.Hal yang menyebabkan terjadinya dormansi yaitu adanya rudimentary
embryo. Di dalam keadaan seperti ini, embrio belum mencapai tahap kematangan
(immature embryo) sehingga memerlukan waktu untuk siap berkecambah.
Faktor lain yang cukup menentukan terhadap keberhasilan perkecambahan
adalah faktor kematangan biji (seed maturity).Hubungan antara faktor kematangan
biji dengan persentase perkecambahan, telah dilakukan penelitian oleh Kinch dan
Termunde (1957) pada biji Perenial Sow Thistle dan Canada Thistle. Dari hasil
(83 %) untuk biji yang diambil pada 9 hari setelah berbunga. Sedangkan untuk
Canada Thistle yaitu 90% untuk biji yang diambil pada 10 hari setelah berbunga.
. Adapun faktor lingkungan yang berpengaruh terhadap proses perkecambahan
yaitu air, udara, temperatur, cahaya, dan zat kimia yang mendukung pada proses
perkecambahan.Air adalah salah satu faktor lingkungan yang sangat diperlukan dalam
perkecambahan. Adanya air sangat penting untuk aktivitas enzim dan penguraiannya,
translokasi dan untuk keperluan fisiologis lainnya.
Faktor lingkungan lain yang berpengaruh dalam proses perkecambahan yaitu
udara. Udara terdiri dari 20 % oksigen, 0,03 % karbon dioksida, dan 80 % nitrogen.
Adanya oksigen di dalam proses respirasi pada perkecambahan, sangat berpengaruh.
Apabila konsentrasi oksigen di udara sangat rendah, menyebabkan terhambatnya
perkecambahan.
Hubungannya dengan temperatur, perkecambahan memerlukan temperatur
yang optimum, yaitu temperatur yang dapat mengakibatkan persentase
perkecambahan yang tinggi dalam waktu yang relatif singkat. Perlu dikemukakan
disini bahwa temperatur minimum, optimum, dan maksimum dikenal dengan
temperatur kardinal. Menurut Copeland (1976), temperatur optimum bagi
perkecambahan sekitar 150-300C, sedangkan untuk temperatur maksimum yaitu 350
-400C.
Cahaya adalah faktor lingkungan lain yang menentukan kemampuan biji
berkecambah. Penelitian pengaruh cahaya terhadap perkecambahan telah dilakukan
oleh Borthwick et al (1952) dan Flint (1936) pada biji lettuce .(Abidin,Z. 1991)
2.2 PERKECAMBAHAN BIJI KELAPA SAWIT
Kelapa sawit merupakan tumbuhan pohon dengan tinggi dapat mencapai 24 meter.
Bunga dan buahnya berupa tandan, serta bercabang banyak. Buah yang masak
berwarna merah kehitaman dengan daging buah padat. Daging dan kulit buah
kosmetik. Ampasnya dapat dimanfaatkan untuk makanan ternak dan tempurungnya
dapat digunakan sebagai bahan bakar. Secara taksonomi, tanaman kelapa sawit dapat
diuraikan sebagai berikut :
Kingdom : Tumbuhan
Divisi : Magnoliophyta
Kelas : Lliliopsida
Ordo : Arecales
Famili : Arecaceae
Jenis : Elaeis
Spesies : E. Guineensis
( Sumber : Diah Muliad,Direktorat Ekspor Produk Pertanian dan Kehutanan,2009)
Gambar 2.1 Buah kelapa sawit
Pada saat ini, telah dikenal beberapa varietas unggul kelapa sawit yang
dianjurkan untk ditanam di perkebunan. Varietas-varietas unggul tersebut dihasilkan
melalui hibridisasi atau persilangan buatan antara varietas Dura sebagai induk betina
dengan varietas Pisifera sebagai induk jantan. Dari hasil pengujian varietas-varietas
tersebut mempunyai kualitas dan kuantitas yang lebih baik dibandingkan varietas
lainnya. Sebagai contoh persilangan buatan varietas unggul kelapa sawit yaitu :
persilangan antara Dura Deli Marihat 434 D dengan Pisifera Yangambi L718T.
Perkecambahan biji kelapa sawit adalah proses yang lambat sepanjang tahun, dalam
pembibitan modern, hal yang pertama dilakukan adalah biji dipanaskan pada suhu
380 C sampai 400 C selama 40 hari dan direndam dengan air untuk mencapai
kondisi yang lembab, (Hussey,G.,1958)
Prosedur ini meniru kondisi natural di Negara Afrika Barat yang merupakan
Negara asal tanaman kelapa sawit dimana biji kelapa sawit berkecambah pada saat
musim penghujan pada permulaan yang diikuti dengan musim kemarau yang
berkepanjangan, sehingga agar proses perkecambahan dapat terjadi dibutuhkan
perlakuan panas sebelumnya.(Rees,A.R.,1962).
Pada saat berkecambah, embrio pecah dan siap untuk membentuk pori
kecambah, kemudian embrio akan membentuk jaringan yang secara cepat
berkembang menjadi plumula (pucuk daun) dan radikula (akar). Aktivitas enzim
lipase terdapat pada saat biji mengalami masa dormansi dan pada saat biji mengalami
proses perkecambahan pada biji Jatropha curcas L. (Abigor, 2002).
Pada saat yang sama, embrio akan membentuk struktur kotiledon yang disebut
dengan haustorium.(Boatman,S.G.; Crumble,W.M., 1958). Haustorium adalah
struktur berongga yang saling membelit pada poros biji. Pada saat biji tumbuh,
haustorium akan mengelilingi endosperm yang pecah dan menyerapnya. Sehingga
setelah tiga bulan, haustorium akan mengisi rongga biji. Setelah itu, daun pertama
akan muncul setelah 20 sampai 40 hari.(Corley,R.H.V.,1976).
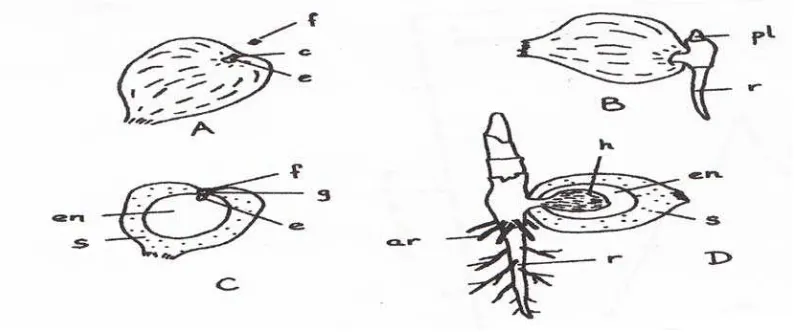
Keterangan :A. Kecambah biji segar ; B. Kecambah berumur 7 hari; C. Penampang
kecambah biji; D. Kecambah berumur 14 hari; ar=penyokong akar,
c=tudung kecambah; e=embrio; en=endosperm; f=serat penyumbat;
g=pori kecambah; h=haustorium; pl=plumula; r=radikula; s=cangkang.
(Stumpf,P.K., 1983)
2.3. ENZIM
Enzim mempunyai tenaga katalitik yang luar biasa dan biasanya jauh lebih besar dari
katalisator sintetik. Spesifitas enzim sangat tinggi terhadap substratnya. Enzim
mempercepat reaksi kimia secara spesifik tanpa pembentukan produk samping.
Enzim merupakan unit fungsional untuk metabolisme dalam sel, bekerja menurut
urutan yang teratur. Sistem enzim terkoordinasi dengan baik menghasilkan suatu
hubungan yang harmonis diantara sejumlah aktivitas metabolik yang berbeda.
Kebanyakan enzim diberi nama dengan penambahan akhiran –ase pada kata
yang menunjukkan senyawa asal yang diubah oleh enzim atau pada nama jenis reaksi
kimia yang dikatalisis enzim.(Winarno,1983)
Semua enzim murni yang telah diamati sampai saat ini adalah protein dan
aktivitas katalitiknya bergantung kepada integritas strukturnya sebagai protein.
Sebagai contoh, jika suatu enzim didihkan dengan asam kuat atau diinkubasi dengan
tripsin, yaitu perlakuan yang memotong rantai polipeptida, aktivitas katalitiknya
biasanya akan hancur ; hal ini memperlihatkan bahwa struktur kerangka primer
protein enzim dibutuhkan untuk aktivitasnya. Enzim, seperti protein lain, mempunyai
berat molekul yang berkisar dari kira-kira 12000 sampai lebih dari
1000000.(Lehninger,1997).
Molekul protein terdiri dari ribuan atom. Satuan dasar penyusun protein
adalah asam amino. Setiap molekul asam amino paling tidak mengandung karbon,
hidrogen, oksigen, nitrogen, serta kadang juga mengandung belerang. Sintesis protein
Rantai asam amino ini disebut dengan polipeptida. Molekul protein dapat
terdiri dari 1 atau lebih rantai polipeptida dimana masing-masing rantai polipeptida
terdiri dari ratusan unit asam amino. Komposisi dan ukuran setiap molekul protein
tergantung pada asam-asam amino penyusunnya .Umumnya pada setiap molekul
protein dapat dijumpai 18-20 jenis asam amino. Protein tumbuhan umumnya
mempunyai berat molekul lebih dari 40000g/mol.(Lakitan,B.,2011)
Enzim mempunyai kekhasan yaitu hanya bekerja pada satu reaksi saja. Suatu
enzim ukuran yang lebih besar daripada substratnya. Oleh karena itu tidak seluruh
bagian enzim dapat berhubungan dengan substrat, bagian enzim yang mengadakan
hubungan dengan substrat disebut bagian aktif daripada enzim.
Fungsi suatu enzim adalah sebagai katalis untuk proses biokimia yang terjadi
dalam sel maupun di luar sel. Suatu enzim dapat mempercepat reaksi 108 sampai 1011
kali lebih cepat daripada apabila reaksi tersebut dilakukan tanpa katalis. Jadi enzim
dapat berfungsi sebagai katalis yang sangat efisien, di samping itu mempunyai derajat
kekhasan yang tinggi. (Poedjiadi, 1994).
2.3.1. Sifat – Sifat Enzim
1. Spesifitas
Aktivitas enzim sangat spesifik. Pada umumnya enzim tertentu hanya dapat
mengkatalisis satu reaksi. Sebagai contoh, laktase menghidrolisis gula laktosa tetapi
tidak berpengaruh terhadap disakarida yang lain. Hanya molekul laktosa saja yang
akan sesuai dalam sisi aktif molekul.
2. Pengaruh suhu
Aktivitas enzim sangat dipengaruhi oleh suhu. Suhu optimalnya adalah antara
35oC dan 40o
3. Pengaruh pH
C, yaitu suhu tubuh. Pada suhu diatas dan dibawah optimalnya, aktivitas
enzim berkurang.
Masing – masing reaksi yang dikatalisis oleh enzim paling cepat terjadi pada pH yang
tertentu. Untuk kebanyakan enzim pH optimal adalah sekitar pH 7 (netral) dan jika
4. Ko-enzim dan aktivator
Enzim sering kali memerlukan bantuan substansi lain agar berfungsi secara efektif.
Ko-enzim adalah substansi bukan protein yang mengaktifkan enzim (Gaman, 1992).
2.3.2. Faktor – Faktor yang Mempengaruhi Aktivitas Enzim
1.Pengaruh Suhu
Aktivitas enzim sangat dipengaruhi oleh suhu. Untuk enzim hewan suhu optimal
antara 35°C dan 40°C, yaitu suhu tubuh. Pada suhu di atas dan di bawah optimalnya,
aktivitas enzim berkurang. Di atas suhu 50°C enzim secara bertahap menjadi inaktif
karena protein terdenaturasi. Pada suhu 100°C semua enzim rusak. Pada suhu yang
sangat rendah, enzim tidak benar-benar rusak tetapi aktivitasnya sangat banyak
berkurang (Gaman & Sherrington, 1994). Enzim memiliki suhu optimum yaitu sekitar
18-230C atau maksimal 400C karena pada suhu 450C enzim akan terdenaturasi karena
merupakan salah satu bentuk protein. (Tranggono,B.S.,1989)
Suhu yang tinggi akan menaikkan aktivitas enzim namun sebaliknya juga
akan mendenaturasi enzim (Martoharsono, 1994). Peningkatan temperatur dapat
meningkatkan kecepatan reaksi karena molekul atom mempunyai energi yang lebih
besar dan mempunyai kecenderungan untuk berpindah. Ketika temperatur meningkat,
proses denaturasi juga mulai berlangsung dan menghancurkan aktivitas molekul
enzim. Hal ini dikarenakan adanya rantai protein yang tidak terlipat setelah
pemutusan ikatan yang lemah sehingga secara keseluruhan kecepatan reaksi akan
menurun (Lee, 1992)
2.Pengaruh pH
pH optimal enzim adalah sekitar pH 7 (netral) dan jika medium menjadi sangat asam
atau sangat alkalis enzim mengalami inaktivasi. Akan tetapi beberapa enzim hanya
beroperasi dalam keadaan asam atau alkalis. Sebagai contoh, pepsin, enzim yang
Enzim memiliki konstanta disosiasi pada gugus asam ataupun gugus basa terutama
pada residu terminal karboksil dan asam aminonya. Namun dalam suatu reaksi kimia,
pH untuk suatu enzim tidak boleh terlalu asam maupun terlalu basa karena akan
menurunkan kecepatan reaksi dengan terjadinya denaturasi. Sebenarnya enzim juga
memiliki pH optimum tertentu, pada umumnya sekitar 4,5–8, dan pada kisaran pH
tersebut enzim mempunyai kestabilan yang tinggi (Williamson & Fieser, 1992).
3.Konsentrasi substrat dan konsentrasi enzim
Katalisis terjadi hanya jika enzim dan substrat membentuk suatu kompleks. Oleh
sebab itu, laju reaksi bergantung pada jumlah enzim dan substrat yang berhasil
membentuk kompleks. Jika konsentrasi keduanya tinggi, jumlah kompleks yang
mungkin terbentuk juga tinggi. Jika substrat cukup tersedia, penggandaan konsentrasi
enzim menyebabkan laju reaksi meningkat dua kali lipat. Jika kemudian substrat
menjadi faktor pembatas, maka penambahan enzim selanjutnya tidak lagi
mempengaruhi laju reaksi.
4.Pengaruh produk reaksi
Laju reaksi enzimatik dapat diketahui dengan cara mengukur laju pengurangan
substrat atau dengan laju terbentuknya produk. Dengan kedua pendekatan ini
diketahui bahwa laju reaksi berlangsung semakin lama semakin lambat. Penurunan
laju reaksi ini, kadang disebabkan oleh denaturasi protein selama pengukuran
berlangsung, tetapi faktor lain juga berperan. Satu faktor yang paling penting adalah
pengaruh dari penurunan konsentrasi substrat dan penimbunan produk reaksi.
Akumulasi produk reaksi kadang mencapai konsentrasi yang cukup tinggi
untuk menyebabkan berlangsungnya reaksi balik (reverse reaction). Ini terjadi jika
potensi kimia relatif antara produk dan substrat memungkinkan. Dalam beberapa
kasus, produk menghambat laju reaksi dengan cara menyatu dengan enzim
5.Pengaruh Unsur atau Senyawa Penghambat Enzim (Inhibitor)
Beberapa bahan asing dapat menghalangi efek katalitik enzim. Beberapa diantaranya
adalah unsur-unsur anorganik seperti beberapa kation logam dan beberapa senyawa
organik tertentu. Kedua kelompok penghambat ini dibedakan berdasarkan
pengaruhnya yang bersifat kompetitif dan non-kompetitif dengan substrat.
Penghambat kompetitif umumnya mempunyai struktur mirip dengan substrat
sehingga dapat berkompetisi untuk mendapatkan sisi aktif enzim. Jika penggabungan
antara enzim dan penghambat terjadi, maka konsentrasi enzim yang efektif menjadi
menurun, sebagai akibatnya tentu laju reaksi juga akan menurun.(Lakitan,B.,2011).
2.3.3. Klasifikasi Enzim
Pada tahun 1956, The International Union of Biochemistry membentuk suatu panitia
untuk menyusun konsep dan mengusulkan klasifikasi dan nomenklatur enzim. Baru
tahun 1961 usul tersebut diterima secara resmi.
Prinsip penamaan tersebut ternyata berdasarkan tipe reaksi yang dikatalisis
dan enzim yang dibagi menjadi enam kelompok utama, yaitu :
1. Oksidoreduktase
Enzim oksidoreduktase adalah enzim yang dapat mengkatalisis reaksi oksidasi atau
reduksi suatu bahan. Dalam golongan ini terdapat 2 jenis enzim yang paling utama
yaitu oksidase dan dehidrogenase.
a. Oksidase adalah enzim yang mengkatalisis reaksi antara substrat dengan
molekul oksigen. Yang termasuk enzim oksidase adalah katalase, peroksidase,
b. Dehidrogenase adalah enzim yang aktif dalam pengambilan atom hidrogen
dari substrat. Contohnya yaitu suksinat dehidrogenase, glutamat
dehidrogenase, dan laktat dehidrogenase.
2. Transferase
Enzim transferase adalah enzim yang ikut serta dalam reaksi pemindahan (transfer)
suatu radikal atau gugus. Enzim yang termasuk dalam golongan ini adalah
transglikosidase, transfosforilase, transaminase, dan transasetilase.
3. Hidrolase
Enzim hidrolase merupakan enzim yang sangat penting dalam pengolahan pangan,
yaitu enzim yang mengkatalisis reaksi hidrolisis suatu substrat atau pemecahan
substrat dengan pertolongan molekul air. Enzim yang termasuk kedalam golongan ini
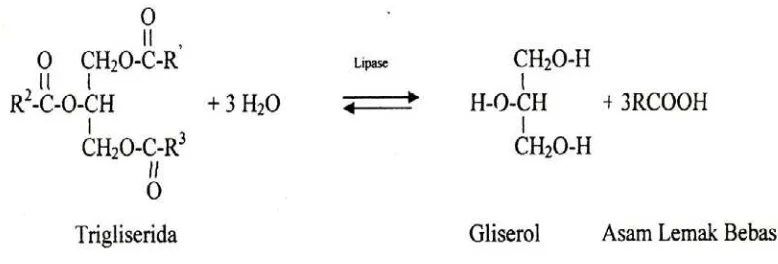
adalah lipase yang menghidrolisis ikatan ester pada lemak alami menjadi gliserol dan
asam lemak, glikosidase menghidrolisis ikatan glikosida dan sebagainya. Disamping
itu masih banyak lagi yang termasuk enzim hidrolase, diantaranya karboksil esterase, pektin metal esterase, selulase, β-amilase, α-amilase dan invertase.
4. Liase
Enzim liase adalah enzim yang aktif dalam pemecahan ikatan C-C dan ikatan C-O
dengan tidak menggunakan melekul air. Yang termasuk dalam golongan enzim ini
adalah enzim dekarboksilase.
5. Isomerase
Enzim isomerase adalah enzim yang mengkatalisis reaksi perubahan konfigurasi
molekul substrat, sehingga dihasilkan molekul baru yang merupakan isomer dari
substrat, atau dengan perubahan isomer posisi. Yang termasuk dalam golongan ini
6. Ligase
Enzim ligase adalah enzim yang mengakatlisis pembentukan ikatan - ikatan tertentu,
misalnya pembentukan ikatan C-O, C-C, dan C-S dalam biosintesis ko-enzim A serta
pembentukan ikatan C-N dalam sintesis glutamin ( Winarno, 1983 ).
2.4. Enzim Lipase
Lipase ( E.C.3.1.1.3 ) adalah enzim yang terutama untuk hidrolisa dari asil gliserida.
Bagaimanapun jumlah berat molekul dari ester baik tinggi maupun rendah tiol ester,
amida, poliol dan lain – lain, dapat diterima sebagai substrat oleh kelompok enzim
lipase ini. Pencampuran dari minyak juga telah dikatalisa dengan lipase, penggunaan
biokatalis ini karena keselektifan dari lipase yang mana memberikan kontrol terhadap
produk ( Gandhi, 1997 ).
Biji yang sedang berkecambah memiliki aktifitas lipolitik yang tinggi.
Aktifitas lipase pada fraksi kecambah tiga kali lebih besar daripada aktifitas enzim
pada fraksi biji. Hal ini disebabkan lipase digunakan untuk memecah substrat berupa
lemak untuk memenuhi kebutuhan energi. Kandungan triasilgliserida (TAG)
menurun dan kandungan monoasilgliserida (MAG) dan asam lemak bebas (FFA)
meningkat dan diasilgliserida (DAG) tidak banyak berubah selama perkecambahan.
Komponen lemak dan lemak netral biji borage (Borago officinalis L.) diubah menjadi
glikolipid dan phospolipid selama perkecambahan dalam gelap suhu 250 C selama 10
hari.(Sennanayake dan Shahidi,2000) .
Enzim adalah suatu biokatalisator yang dapat bertindak menguraikan molekul
yang rantainya panjang menjadi lebih sederhana, serta dapat juga membantu
mekanisme reaksi yang mana tergantung pada enzimnya. Walaupun enzim ikut serta
dalam reaksi dan mengalami perubahan fisik selama reaksi, enzim akan kembali
Enzim mempunyai tenaga katalitik yang luar biasa dan biasanya jauh lebih besar dari
katalisator sintetik. Spesifitas enzim sangat tinggi terhadap substratnya. Enzim
mempercepat reaksi kimia secara spesifik tanpa pembentukan produk samping.
Enzim merupakan unit fungsional untuk metabolisme dalam sel, bekerja menurut
urutan yang teratur. Sistem enzim terkoordinasi dengan baik menghasilkan suatu
hubungan yang harmonis diantara sejumlah aktivitas metabolik yang berbeda.
Kebanyakan enzim diberi nama dengan penambahan akhiran –ase pada kata
yang menunjukkan senyawa asal yang diubah oleh enzim atau pada nama jenis reaksi
kimia yang dikatalisis enzim.
Enzim – enzim yang bekerja dalam hidrolisis lemak dan minyak dapat
dikelompokkan menjadi dua kelompok besar yaitu enzim lipase dan enzim esterase.
Keduanya terlihat baik dalam proses metabolisme lemak maupun penguraian dan
kerusakan lemak. Enzim lipase dan enzim esterase sukar dibedakan karena daya
kerjanya yang sangat mirip, yaitu mengkatalisis hidrolisis ester karbohidrat. Pada
preparat murni enzim diekstraksi dari bahan alami sering terkandung enzim lipase
maupun esterase.
Secara fisiologik, enzim ini penting artinya karena dengan menghidrolisis
lemak dihasilkan asam lemak bebas dan gliserol yang penting peranannya dalam
metabolisme dalam tubuh.
Di bidang industri lemak dan minyak, enzim – enzim ini juga sangat penting
karena peranannya dalam mengendalikan proses produksi minyak dan lemak;
misalnya pada minyak goreng dan margarin dalam proses menyingkirkan cita rasa
dan bau – bauan yang tidak dikehendaki atau sebaliknya dengan enzim tersebut
beberapa cita rasa yang dikehendaki dapat diatur untuk ditampilkan.
Berdasarkan nomenklatur dari International Union of Biochemistry, enzim
2.4.1. Sifat – Sifat Enzim Lipase
Tergantung dari asal dan substratnya, keaktifan optimum lipase sangat tergantung
pada pH dan suhu. Enzim lipase pada pankreas misalnya mempunyai pH optimal
antara 8 dan 9, tetapi dapat menurun menjadi antara 6 – 7 bila substratnya berbeda.
Keaktifan optimal enzim lipase tegantung juga dari senyawa pengemulsi yang
digunakan dan ada tidaknya garam dalam substrat. Enzim lipase yang berasal dari
susu mempunyai pH optimal sekitar 9.
Suhu optimal enzim lipase pada umumnya berkisar antara 30o – 40oC.
Meskipun telah ditemukan adanya lipase yang masih aktif pada suhu -29oC, terutama
2.4.2. Sumber – Sumber Enzim Lipase
Lipase biasanya diproduksi oleh pankreas babi dan sapi, ragi Candida, Aspergillus,
Rhizopus, dan Mucor sp.(Ghandi,1997). Pada umumnya sumber lipase adalah
mikrobia(Ghosh dkk,1996) dan jamur (Nelson dkk, 1996). Lipase tedapat juga pada
biji dan buah tanaman seperti palma, selada, bekatul, beras, barley, gandum, oat,
kapas, jagung, mentimun, dan kacang-kacangan(Abigor,2002;Sennayake dan Shahidi,
2000 ; Mohammed, 2000 ; Dundas, 1998).
2.4.3. Aktifitas Enzim Lipase
Keaktifan enzim dapat ditentukan secara kualitatif dengan reaksi kimia yaitu dengan
substrat yang dapat dihidrolisis oleh enzim tersebut, dan secara kuantitatif ditentukan
dengan mengukur laju reaksi tersebut. Aktivitas enzim lipase mempunyai satuan unit
(U). Satu unit aktivitas enzim lipase setara dengan 1µ mol asam lemak bebas yang
dihasilkan dari hidrolisis substrat yang dikatalisis oleh enzim lipase tiap satuan menit
(Handayani, 2005).
Untuk menentukan aktivitas optimum pada kondisi optimum dari enzim lipase
maka dilakukan pengukuran aktivitas enzimatik pada variasi suhu dan pH. Sehingga
akan diketahui berapa aktifitas lipase di setiap rentang suhu dan pH yang ditentukan.
Seperti protein lainnya, enzim dapat terdenaturasi pada suhu tertentu, perilaku
kimia, dan kondisi ekstrim lainnya. Apabila terjadi proses denaturasi, maka bagian
aktif enzim akan terganggu dan dengan demikian konsentrasi efektif enzim menjadi
berkurang dan reaksinya pun akan menurun. Dengan demikian, perubahan pH
lingkungan akan berpengaruh terhadap efektivitas bagian aktif enzim dalam
membentuk kompleks enzim substrat. (Poedjiadi, 1994)
Selain itu enzim mempunyai pH optimum yang spesifik, yaitu pH yang
menyebabkan aktivitas enzim maksimal. pH optimum enzim tidak perlu sama dengan
pH lingkungan normalnya, dengan pH yang mungkin sedikit di atas atau dibawah pH
2.5 ISOLASI DAN PEMURNIAN ENZIM
Enzim merupakan suatu protein sehingga untuk mengisolasi enzim, protein harus
diisolasi dalam bentuk murni, protein yang diinginkan harus dipisahkan dari semua
jenis protein yang lain dan biomolekul yang lainnya. Protein seringkali diisolasi dari
jaringan hewan atau tumbuhan, cairan biologi, sel mikrobiologi yang sebelumnya
harus diubah terlebih dahulu sebagai sel homogenat
Ekstrak yang mengandung ribuan jenis protein yang berbeda dan juga
biomolekul yang lainnya dipisahkan berdasarkan sifat-sifat protein, yaitu polaritas,
muatan, ukuran (massa molekul) dan kemampuan untuk berikatan dengan molekul
yang lain.(Boyer,2006).
Setelah sel homogen, protein dapat diekstraksi dengan larutan buffer encer
pada pH yang sesuai dengan pH darimana enzim lipase diisolasi, dimana jika enzim
diisolasi dari tumbuhan pH yang sesuai adalah sekitar 6,0-7,0. Metode yang biasanya
digunakan untuk memisahkan protein adalah presipitasi differensial, kromatografi
penukar-ion, elektroforesis, filtrasi gel, dan ultrasentrifuga si.
Metode ini diperkenalkan pertama kali oleh Svedberg (1925) dengan prinsip
menggunakan gaya sentrifugal. Jika larutan yang mengandung makromolekul
sejenis, maka mereka akan turun kebawah tabung sentrifuge pada kecepatan yang
sama dan apabila larutan mengandung campuran makromolekul yang mempunyai
bentuk dan ukuran yang berbeda, akan terjadi perbedaan penempatan karena adanya
perubahan indeks bias dalam larutan.(Cole,A.S,1977).
Pemutaran homogenat di dalam sentrifuge akan memisahkan bagian-bagian
sel ke dalam dua fraksi, yaitu pelet, yang terdiri atas struktur-struktur lebih besar yang
terkumpul di bagian bawah tabung sentrifuge, dan supernatan, yang terdiri atas
bagian-bagian sel yang lebih kecil yang tersuspensi dalam cairan di atas pelet
tersebut.Supernatan dapat disentrifugasi kembali dengan kecepatan yang lebih tinggi
Enzim lipase yang dihasilkan dalam bentuk cair harus dipekatkan terlebih dahulu
untuk mendapatkan ekstrak enzim. Proses pemekatan enzim dapat dilakukan dengan
pengendapan protein melalui penambahan garam mineral. Metode ini merupakan
bagian dari proses isolasi enzim dengan metode ekstraksi. Metode ekstraksi
digunakan untuk memisahkan enzim (protein) yang terkandung dalam larutan dengan
menggunakan garam mineral, sehingga enzim yang merupakan fraksi berat akan
terendapkan di bawah.(Sri,W.M., 2011).
Menurut Belter dkk (1988), dalam pemilihan jenis garam mineral tersebut
terdapat beberapa hal yang harus diperhatikan : (1) Anion efektif dalam urutan
sebagai berikut:citrate > PO43- > SO42- > CH3COO- > Cl- > NO3-. (2) Kation efektif
dalam urutan sebagai berikut : NH4+ > K+ > Na+ . (3) Dipilih garam yang murah, jika
akan digunakan dalam jumlah yang banyak. (4) Dipilih garam yang densitasnya
berbeda dari densitas larutan, sehingga dapat dilakukan pemisahan dengan proses
sentrifugasi.
Amonium sulfat merupakan garam mineral yang paling umum digunakan
dalam proses pengendapan enzim, karena solubilitasnya di dalam air amat tinggi,
tidak mengandung zat-zat yang toksik terhadap kebanyakan enzim, harganya relatif
murah dan dalam jumlah banyak dapat bertindak sebagai stabilisator enzim itu sendiri
(Darwis dan Sukara,1990).
2.6. RBDPO (Refined Bleached Deodorized Palm Oil) sebagai substrat.
Refined Bleached and Deodorized Palm Oil (RBDPO) adalah minyak sawit yang
telah mengalami proses penyulingan untuk menghilangkan asam lemak bebas serta
penjernihan untuk menghilangkan warna dan penghilangan bau. Proses pengolahan
kelapa sawit menjadi minyak goreng sawit dimulai dari proses pengolahan tandan
buah segar menjadi Crude Palm oil (CPO). Minyak sawit yang digunakan sebagai
produk pangan biasanya dihasilkan dari minyak sawit maupun minyak inti sawit
Dewasa ini, produksi CPO (Crude Palm Oil) Indonesia sebagian besar difraksinasi
sehingga dihasilkan fraksi olein cair dan fraksi stearin padat. Fraksi olein itulah yang
digunakan untuk memenuhi kebutuhan minyak goreng domestik sebagai pelengkap
minyak goreng dari minyak kelapa.(Swadaya,P., 2001)
Setelah kelapa sawit berubah menjadi CPO, maka proses selanjutnya adalah
mengolah CPO menjadi minyak goreng sawit. Secara garis besar proses pengolahan
CPO menjadi minyak goreng sawit, terdiri dari dua tahap yaitu tahap pemurnian
(refinery) dan pemisahan (fractionation).
Setelah kelapa sawit berubah menjadi CPO, maka proses selanjutnya adalah
mengolah CPO menjadi minyak goreng sawit. Secara garis besar proses pengolahan
CPO menjadi minyak goreng sawit, terdiri dari dua tahap, yaitu tahap pemurnian
(refinery) dan pemisahan (fractionation).Tahap pemurnian terdiri dari penghilangan
gum/getah (degumming), pemucatan (bleaching), dan penghilangan bau
(deodorization).
CPO yang berasal dari tangki penampungan CPO dipompa melalui rainer
menuju refinery. Pada proses ini terjadi pemanasan CPO untuk mempermudah
pemompaan CPO ke tangki berikutnya. Hasil dari proses ini disebut DPO
(Degummed Palm Oil), kemudian di pompa menuju drier dengan kondisi vakum lalu
dipompakan ke reaktor yang terlebih dahulu melewati static mixer kemudian turun ke
slurry tank yang didalamnya terjadi pemanasan sampai temperature 90-1200 C dan
penambahan H3PO4, CaCO3, dan Bleaching Earth.
Slurry Oil dari slurry tank mengalir ke bleacher dan dipompa ke filter untuk
filtrasi. Hasil dari filtrasi ini adalah DBPO (Degummed Bleached Palm Oil) yang
selanjutnya dialirkan ke intermediate tank untuk tahap deodorizing. DBPO yang
dihasilkan dialirkan ke deaerator lalu dipompa ke Spiral Heat Exchanger (SHE).
Dalam proses ini terjadi penambahan panas dengan temperatur 185-2000C, lalu
dialirkan ke flash vessel dan turun ke packed column dan dialirkan lagi menuju
deodorize yang didalamnya terjadi penghilangan zat-zat yang menimbulkan bau
DBPO yang sudah hilang baunya dipompa kembali ke SHE untuk mengalami
pertukaran panas.Dan dalam hal ini minyak sudah dalam bentuk RBDPO (Refined
Bleached Degummed Palm Oil).
Minyak sawit terdiri dari gliserida campuran yang merupakan ester dari
gliserol dan asam lemak rantai panjang. Dua jenis asam lemak yang paling dominan
dalam minyak sawit yaitu asam palmitat, C16:0 (jenuh), dan asam oleat, C18:1 (tidak
jenuh). Umumnya, komposisi asam lemak minyak sawit dapat dilihat pada Tabel di
bawah ini:
Tabel 2.1 Komposisi Asam Lemak Minyak Sawit
Nama Asam Lemak Rumus Asam Lemak Komposisi
Laurat C12:0 0,2 %
Miristat C14:0 1,1 %
Palmitat C16:0 44,0 %
Stearat C18:0 4,5 %
Oleat C18:1 39,2 %
Linoleat C18:2 10,1 %
Lainnya - 0,9 %
(Sumber: Pahan,I., 2008)
Kusumo,D.P.(2008) dalam penelitiannya “Sintesis dan Karakterisasi Minyak
Kaya DAG (MK-DAG) Berbahan Baku RBDPO Dengan Metode Gliserolisis
Enzimatis ” menunjukkan bahwa hasil analisis terhadap RBDPO (Refined Bleached
Deodorized Palm Oil) memiliki kadar air sebesar 0,08 % (b/b), nilai bilangan
peroksida sebesar 1,97 meq/kg, bilangn iod sebesar 52,38 % dan nilai ALB (Asam
BAB 3
- Neraca Analitis Mettler Toledo
- Centrifuge 5000 rpm Hitachi
- Centrifuge 10000 rpm Hitachi
- Pipet Tetes
- Statif dan Klem
- Botol Akuades
- pH meter Walklab
- Pipet Volumetri Pyrex
- Ball-Pipet
- Indikator Fenolftalein p.a.(E.Merck)
p.a.(E.Merck)
- KOH(s)
- Asam oksalat
p.a.(E.Merck)
- RBDPO (Refined Bleached Deodorized Palm Oil)
- Air
- Kecambah Biji Kelapa Sawit (Elaeis guineensis Jacq)
- Fungisida Dithane M-40
- Kotak Gray
3.3. Prosedur Penelitian
3.3.1. Pembuatan Larutan Pereaksi
3.3.1.1. Indikator Fenolftalein 1%
Sebanyak 1 g indikator Fenolftalein ditimbang dan dilarutkan dengan etanol dalam
labu takar 100 mL sampai garis tanda.
3.3.1.2. Pembuatan Larutan KOH 0,0906 N a. Pembuatan larutan KOH 0,0906 N
Sebanyak 5,61 g KOH ditimbang dan dimasukkan kedalam labu takar 1000 mL,
kemudian dilarutkan dengan akuades hingga garis tanda, setelah itu dihomogenkan.
b. Standarisasi Larutan KOH 0,0906 N dengan Asam Oksalat
Sebanyak 0,63 g asam oksalat ditimbang dengan teliti (BM = 126), kemudian
dilarutkan kedalam 100 mL akuades dan dipipet sebanyak 10 mL kemudian
ditambahkan 3 tetes indikator Fenolftalein kemudian dititrasi dengan larutan KOH
yang akan distandarisasi hingga warna merah rose. Hal yang sama dilakukan 3 kali
ulangan.
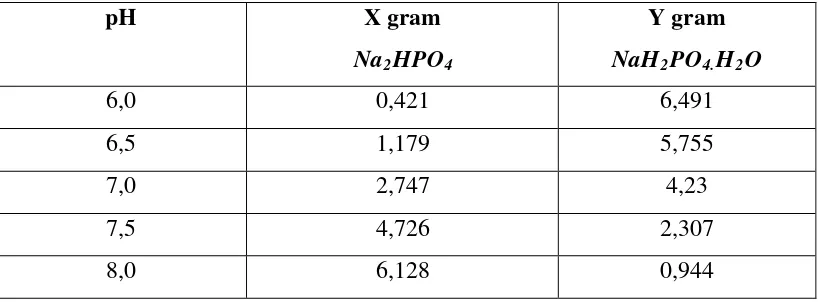
3.3.2. Pembuatan Buffer Fosfat 0,05 M
A = X gram Na2HPO
B = Y gram NaH
4 2PO4.H2
A + B dimasukkan kedalam labu takar 1000 mL dan diencerkan sampai garis tanda.
O
Tabel 3.1 Pembuatan Larutan Buffer Phosfat pH 6,0-8,0
Perhitungan pembuatan buffer fosfat 0,05 M dapat dilihat pada lampiran 1
3.3.3. Pembentukan Kecambah Dari Biji Kelapa Sawit
Tandan buah kelapa sawit varietas DxP dipisahkan bagian buah dari tandannya lalu
dikupas. Kemudian biji kelapa sawit direndam dengan air selama 7 hari lalu dibilas
dengan larutan Dithane M-40 0,4 % dan dikeringanginkan selama 1 hari. Kemudian
dimasukkan kedalam germinator selama pada suhu 400C selama 60 hari lalu direndam
kembali dengan air selama 3 hari dan dibilas kembali dengan larutan Dithane M-40
0,4% ,setelah itu dimasukkan ke dalam ruangan pada suhu 300C selama 21 hari.
Kemudian dimasukkan kecambah biji kelapa sawit ke dalam plastik dan direndam
dengan air pada suhu 25º C selama 14 hari.
3.3.4 Penyediaan Ektrak Kasar Enzim Lipase Dari Kecambah Biji Kelapa Sawit.
Sebanyak 90 kecambah biji kelapa sawit (416,5501 g) dan dipisahkan cangkang
dengan biji bagian dalam, biji bagian dalam (121,179 g) ditambahkan dengan buffer
fosfat pH 7,0 dan diblender hingga halus, kemudian disaring. Filtrat disentrifugasi
pada 5000 rpm selama 30 menit. Supernatan ditambahkan dengan NH4(SO4)2
sebanyak 100,68 gram, kemudian didiamkan selama 1 malam pada suhu 40C.
Suspensi yang terbentuk disentrifugasi pada 10000 rpm selama 30 menit dan endapan
yang dihasilkan dilarutkan dengan buffer fosfat pH 7,0.
3.3.5. Penentuan Suhu Optimum Untuk Aktivitas Ekstrak Kasar Enzim Lipase Pada Hidrolisis RBDPO
Ditimbang RBDPO sebanyak 1 gram dan masing-masing dimasukkan ke dalam 5
gelas Erlenmeyer, ditambahkan 2 mL buffer fosfat pH 7,0 ; ditambahkan 1 mL crude
enzim lipase dan dipanaskan gelas Erlenmeyer suhu 300C selama 60 menit, setelah itu
ditambahkan 6 mL etanol:aseton (1:1) dan ditambahkan 3 tetes indikator
Phenolphtalein, setelah itu dititrasi dengan KOH 0,0906 N sampai terjadi perubahan
warna menjadi merah lembayung, dicatat volume KOH 0,0906 N yang terpakai,
dihitung % ALB dan aktivitasnya. Diulangi perlakuan yang sama dengan variasi suhu
350C; 400C; 450C; 500C.
3.3.6. Penentuan pH Optimum Untuk Aktivitas Ekstrak Kasar Enzim Lipase Pada Hidrolisis RBDPO
Ditimbang RBDPO sebanyak 1 gram dan masing-masing dimasukkan ke dalam 5
gelas Erlenmeyer, ditambahkan 2 mL buffer fosfat pH 6,0 dan ditambahkan 1 mL
crude enzim lipase dan dipanaskan pada suhu 400C selama 60 menit, setelah itu
ditambahkan 6 mL etanol: aseton (1:1) dan ditambahkan 3 tetes indikator
Phenophtalein, setelah itu dititrasi dengan KOH 0,0906 N sampai terjadi perubahan
warna menjadi merah lembayung, dicatat volume KOH 0,0906 N yang terpakai,
dihitung % ALB dan aktivitasnya. Diulang perlakuan yang sama dengan variasi
3.4 Bagan Penelitian
3.4.1 Pembentukan Kecambah Dari Biji Kelapa Sawit
Dipisahkan buah kelapa sawit dari tandan
(Pusat Penelitian Kelapa Sawit Medan) Kecambah kelapa sawit berumur 1 hari Buah kelapa sawit (9,056 kg)
Tandan kelapa sawit varietas DxP (±16,00 kg)
Tandan kosong kelapa sawit
dikupas
Direndam dengan air selama 7 hari
Dibilas dengan larutan Dithane M-40 0,4 %
Dikering anginkan selama 1 hari
Diletakkan pada kotak gray
Dimasukkan ke dalam germinator pada suhu 39-40ºC selama 60 hari
Direndam dengan air selama 3 hari
Dibilas dengan larutan Dithane M-40 0,4 %
Diletakkan pada kotak gray
Dimasukkan dalam ruangan pada suhu 30ºC selama 21 hari
Biji kelapa sawit (2,704 kg) Daging buah kelapa sawit (7,525 kg)
Dimasukkan dalam plastik
Direndam dengan air
Diletakkan dalam ruangan pada suhu 25 º C selama 14 hari
ditambahkan dengan 100,68 g (NH4)2SO4
didiamkan selama 1 malam pada suhu 4ºC
disentrifugasi pada 10.000 rpm selama 30 menit
Dilarutkan dengan buffer fosfat pH 7,0 dalam labu
takar 50 mL
3.4.2 Penyediaan Ekstrak Kasar Enzim Lipase dari Kecambah Biji Kelapa Sawit
90 buah kecambah biji kelapa sawit (416,5501 g)
ditambahkan dengan 250 mL buffe
fosfat pH 7,0
diblender
disaring
dipisahkan cangkang dengan biji bagian dalam
Cangkang (295,380 g ) Biji Bagian Dalam (121,179 g)
Residu Filtrat (198,5 mL)
disentrifugasi pada 5000 rpm selama 30 menit
Endapan Supernatan (167,8 mL)
Suspensi (215,5 mL)
Supernatan Endapan (1,52 g )
3.4.3. Penentuan Suhu Optimum Untuk Aktivitas Ekstrak Kasar Enzim Lipase Terhadap Hidrolisis RBDPO
Dilakukan perlakuan yang sama untuk variasi suhu 35ºC; 40ºC; 45ºC; dan 50ºC 1 g RBDPO
dimasukkan masing-masing ke dalam gelas Erlenmeyer 250 mL
ditambahkan 4 mL buffer fosfat pH 7,0
ditambahkan 1 mL crude enzim lipase
dipanaskan gelas Erlenmeyer pada suhu 30ºC selama 60 menit
ditambahkan 6 mL etanol : aseton (1:1)
ditambahkan 3 tetes indikator fenolftalein
dititrasi dengan KOH 0,0906 N sampai terjadi perubahan warn
menjadi merah lembayung
dicatat volume KOH 0,0906 N yang terpakai
dihitung % ALB nya
dihitung aktivitasnya
3.4.4. Penentuan pH Optimum Untuk Aktivitas Ekstrak Kasar Enzim Lipase Pada Hidrolisis RBDPO
Dilakukan perlakuan yang sama untuk variasi buffer fosfat pH 6,5; 7,0; 7,5; dan 8,0.
1 g RBDPO
dimasukkan masing-masing ke dalam gelas Erlenmeyer 250 mL
ditambahkan 4 mL buffer fosfat pH 6,0
ditambahkan 1 mL crude enzim lipase
dipanaskan gelas Erlenmeyer pada suhu 40ºC selama 60 menit
ditambahkan 6 mL etanol : aseton (1:1)
ditambahkan 3 tetes indikator fenolftalein
dititrasi dengan KOH 0,0906 N sampai terjadi perubahan warn
menjadi merah lembayung
dicatat volume KOH 0,0906 N yang terpakai
dihitung % ALB nya
dihitung aktivitasnya
BAB 4
HASIL DAN PEMBAHASAN
4.1. Hasil Penelitian
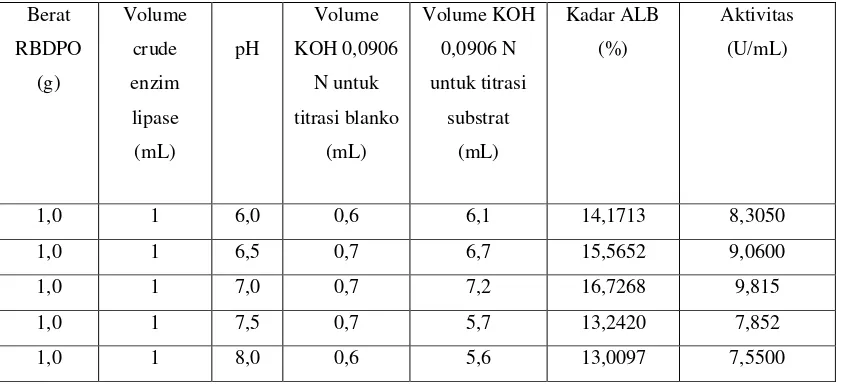
4.1.1 Penentuan Suhu Optimum Untuk Aktivitas Ekstrak Kasar Enzim Lipase Terhadap Hidrolisis RBDPO (Refined Bleached Deodorized Palm Oil)
Data hasil pengamatan aktivitas ekstrak kasar enzim lipase dalam menghidrolisis
RBDPO pada suhu 30º - 50º C dapat dilihat pada tabel dibawah ini :
Tabel 4.1. Hasil Perhitungan Aktivitas Ekstrak Kasar Enzim Lipase pada Suhu
30º - 50º
Kadar ALB (asam lemak bebas) dapat diketahui berdasarkan volume (mL) KOH
0,0906 N yang dipakai untuk membebaskan 1 mg asam lemak bebas RBDPO yang
dihidrolisis oleh crude enzim lipase.
Pengolahan data untuk perhitungan kadar ALB (asam lemak bebas) dapat dilihat
Dari perhitungan kadar ALB pada lampiran 2, maka aktivitas crude enzim lipase
dapat diketahui berdasarkan kadar ALB nya. Dimana aktivitasnya dinyatakan sebagai
jumlah asam lemak bebas yang dihasilkan (U/mL) dari hidrolisis substrat RBDPO.
Pengolahan data untuk perhitungan aktivitas crude enzim lipase dapat dilihat pada
lampiran 3.
4.1.2. Penentuan pH Optimum Aktivitas Ekstrak Kasar Enzim Lipase terhadap Hidrolisis RBDPO
Data hasil perhitungan Aktivitas Crude Enzim Lipase dalam menghidrolisis RBDPO
pada pH 6,0 – 8,0 dapat dilihat pada tabel di bawah ini :
Tabel 4.2. Hasil Perhitungan aktivitas crude enzim lipase pada pH 6,0 – 8,0
Berat
Kadar ALB (asam lemak bebas) dapat diketahui berdasarkan volume (mL) KOH
0,0906 N yang dipakai untuk membebaskan 1 mg asam lemak bebas RBDPO yang
dihidrolisis oleh crude enzim lipase.
Pengolahan data untuk perhitungan kadar ALB (asam lemak bebas) dapat dilihat pada
lampiran 2.
Dari perhitungan kadar ALB pada lampiran 2, maka aktivitas crude enzim lipase
jumlah asam lemak bebas yang dihasilkan (U/mL) dari hidrolisis substrat minyak
RBDPO.
Pengolahan data untuk perhitungan aktivitas crude enzim lipase dapat dilihat pada
4.2. Pembahasan Hasil Penelitian
4.2.1. Isolasi Ekstrak Kasar Enzim Lipase dari Kecambah Biji Kelapa Sawit
Ekstrak kasar enzim lipase diperoleh dari kecambah biji kelapa sawit (Elaeis
guineensis Jacq) yang telah berumur 14 hari. Kemudian isolasi dilakukan dengan
metode ekstraksi yaitu metode pengendapan protein melalui penambahan garam
(NH4)2SO4. Sehingga, enzim (protein) yang terkandung dalam larutan terpisah
dengan partikel non enzim dan enzim yang merupakan fraksi berat akan terendapkan
dibawah larutan. Karena adanya perbedaan densitas antara garam mineral (NH4)2SO4
dengan larutan enzim, maka pemisahan enzim dengan (NH4)2SO4 dilakukan dengan
sentrifugasi.
4.2.2. Penentuan Suhu Optimum untuk Aktivitas Ekstrak Kasar Enzim Lipase
dari Kecambah Biji Kelapa Sawit terhadap Hidrolisis RBDPO
Gambar 4.6. Hidrolisis trigliserida oleh enzim lipase
Pengaruh suhu terhadap aktivitas ekstrak kasar enzim lipase dari kecambah biji
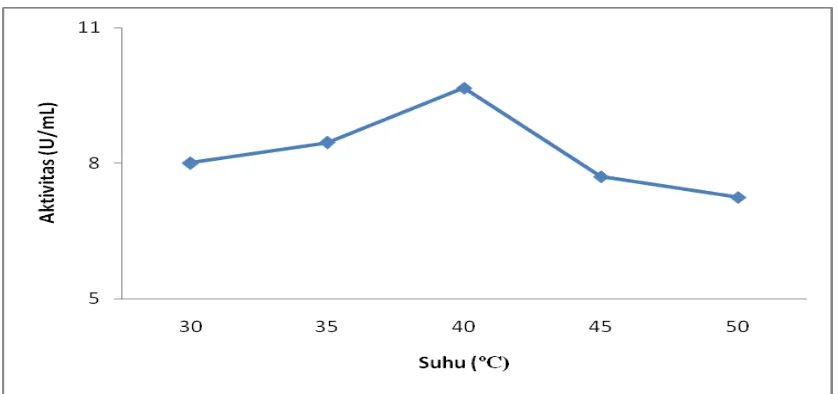
kelapa sawit terhadap hidrolisis RBDPO dengan rentang suhu 30-50o dapat dilihat
Gambar 4.7.Penentuan suhu optimum untuk aktivitas ekstrak kasar enzim lipase
terhadap hidrolisis RBDPO
Pada gambar diatas dapat dilihat bahwa aktivitas crude enzim lipase bertambah
dengan bertambahnya suhu. Suhu optimum dicapai pada suhu 40oC dengan aktivitas
sebesar 9,664 U/mL . Aktivitas ekstrak kasar enzim lipase mulai menurun pada suhu
45oC sebesar 7,701 U/mL dan semakin menurun pada suhu 50ºC dengan aktivitas
sebesar 7,248 U/mL.
Hal ini dikarenakan enzim lipase mengalami kerusakan pada suhu yang lebih
tinggi. Enzim merupakan protein, maka pada suhu tinggi dapat menyebabkan
denaturasi protein, yaitu kerusakan pada struktur protein yang menyebabkan
terganggunya fungsi enzim sebagai katalis, dimana kerja suatu enzim terhadap
substrat dianalogikan sebagai gembok dan kunci, sehingga gembok dan kunci tidak
cocok maka aktivitas enzim terhadap substrat tidak dapat terjadi.
4.2.3. Penentuan pH Optimum terhadap Aktivitas Crude Enzim Lipase dari Kecambah Biji Kelapa Sawit pada Hidrolisis RBDPO
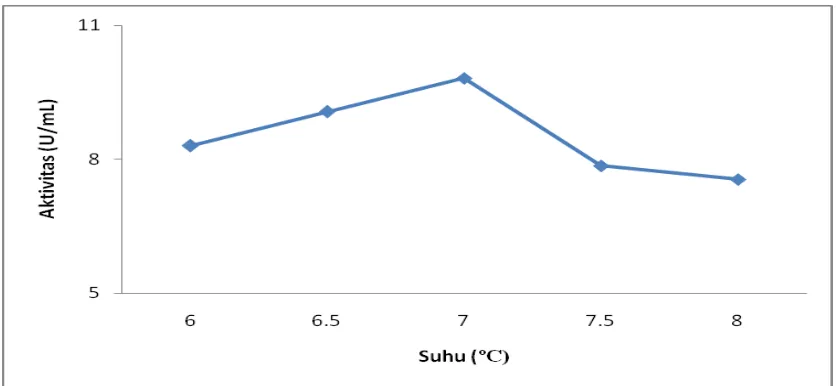
Pengaruh suhu terhadap aktivitas crude enzim lipase dari kecambah biji kelapa sawit
Gambar 4.8. Penentuan pH optimum terhadap aktivitas crude enzim lipase pada
hidrolisis RBDPO
Pada gambar diatas dapat dilihat bahwa aktivitas crude enzim lipase bertambah
dengan naiknya pH. Pada gambar dapat dilihat bahwa pH optimum untuk aktivitas
crude enzim lipase dalam menghidrolisis RBDPO terdapat pada pH 7,0 dengan
aktivitas sebesar 9,815 U/mL. Aktivitas ekstrak kasar enzim lipase mulai menurun
pada pH 7,5 dengan aktivitas 7,852 U/mL dan semakin menurun pada pH 8,0 dengan
aktivitas sebesar 7,550 U/mL.
Pada pH tinggi atau rendah memungkinkan terjadinya denaturasi dan ini akan
mengakibatkan menurunnya aktivitas enzim. Karena enzim merupakan protein, maka
perubahan pH akan menyebabkan ionisasi pada molekul protein berubah. Perubahan
ini akan mengakibatkan struktur tiga dimensi protein berubah sehingga aktivitas
BAB 5
KESIMPULAN DAN SARAN
5.1. Kesimpulan
Dari hasil penelitian yang telah dilakukan mengenai isolasi crude enzim lipase dari
kecambah biji kelapa sawit serta uji aktivitas ekstrak kasar enzim lipase, dapat
disimpulkan sebagai berikut :
a. Isolasi ekstrak kasar enzim lipase dilakukan menggunakan metode ekstraksi
dengan penambahan garam (NH4)2SO4 untuk memisahkan protein (enzim)
dengan partikel non enzim dan metode sentrifugasi untuk memisahkan enzim
dari garam (NH4)2SO4.
b. Suhu dan pH optimum aktivitas ekstrak kasar enzim lipase dari kecambah biji
kelapa sawit (Elaeis guineensis Jacq) terhadap hidrolisis RBDPO (Refined
Bleached Deodorized Palm Oil) adalah 40
o
C dan 7,0 dengan nilai aktivitas
tertinggi yaitu 9,664 U/mL dan 9,815 U/mL.
5.2. Saran
Sebaiknya dilakukan penelitian lebih lanjut yaitu dengan memurnikan dan
menentukan kadar protein dari ekstrak kasar enzim lipase dari kecambah biji kelapa
DAFTAR PUSTAKA
Abidin,Z. 1991. Dasar Pengetahuan Ilmu Tanaman. Bandung: Angkasa.
Abigor,R.D. 2002. Partial Purification and Properties of Lipase from Germinating Seeds Of Jatropha curcas L.J. Am Oil. Soc, 79:hal. 1123-1126.
Arbianti, R. 2008. Pemanfaatan Biji Wijen sebagai Sumber Enzim Lipase untuk Reaksi Esterifikasi Gliserol-Asam Laurat pada Pembuatan Agen Pengemulsi. Jakarta:: Universitas Indonesia.
Belter,P.A.,Cussler,E.L.,1988. Bioseparation Downstream Processing for Biotechnology.New York: John Wiley and Sons.
Boatman,S.G. and Crumble,W.M.1958. Fat Metabolism In The West African Oil : Part 2 Fatty Acid Metabolism in The Developing Seedling in Some Enzymic Activities in the Germination Oil Palm Seedling. ed Cheang, O.K. Plant Physiol.
Boyer, R. 2006. Concepts In Biochemistry.Third Edition.New York: John Wiley and Sons
Campbell, N.A. 2002. Biologi. Jilid I. Edisi Kelima. Jakarta: Erlangga.
Chusnul,H. 2008. Optimasi Produksi Lipase Kecambah Biji Kacang Tanah (Arachis Hypogaea L) sebagai Biokatalis dengan Metode Response Surface Methodology. Yogyakarta: Universitas Gadjahmada.
Cole,A.S. and Eastoe,J.E. 1977. Biochemistry and Oral Biology. England: The Pitman Press.
Corley,R.H.V. 1976. Germination and Seed Growth.In RHV Corley, JJ Hardon, B Wood. Eds. Amsterdam: Development in Crop Science Oil Palm Research Elsevier Publishing Co.
Darwis, A. dan Sukara, E. 1990. Isolasi, Purifikasi dan Karakterisasi Enzim. Dalam Sri, W, M, et al. Produksi, Karakterisasi, dan Isolasi Lipase dari Aspergillus Niger: hal. A08-2 – A08-3.Yogyakarta: Seminar Nasional Teknik Kimia.
Dundas, D.G.A. 1977. Lipase from Vicia faba Minor Food Chem. England: Applied Science Publisher, Ltd. 3:171-178.
Ejedegba, B.O. 2007. Characteristics of Lipase Isolated from Coconut (Cocos nucifera Linn) Seed Under Different Treatments. Nigeria: African Journal of Technology hal 723-727.
Hussey,G. 1958. An Analysis of The Factors Controling The Germination of The Seed of Oil Palm . in Some Enzymic Activities in the Germination Oil Palm Seedling. ed Cheang, O.K. Plant Physiol.
Gaman,P.M. and Sherington,B. 1992. Pengantar Ilmu Pangan Nutrisi dan Mikrobiologi.Edisi Kedua. Yogyakarta: Gadjahmada University Press.
Gandhi, N.N. 1997. Application of Lipase. JAOCS. AOCS Press. Vol 74.
Ghosh, P.K. 1996. Microbial Lipase: Production and Application. Sci. Prog. 79-119-157
Handayani, R. 2005. Transesterifikasi Ester Asam Lemak Melalui Pemanfaatan Teknologi Lipase. LIPI. Bogor.
Johnson, W.H. and Laubengayer, R.A. 1995. Biology. New York:Revised Edition. Holt, Rinehart and Winston.
Kusumo, D.P. 2008. Sintesis dan Karakterisasi Minyak Kaya DAG (MK-DAG)BerbahanBaku RBDPO dengan Metode Gliserolisis Enzimatis. Bogor:: Fakultas Teknologi Pertanian Institut Pertanian Bogor.
Lakitan, B. 2011. Dasar-Dasar Fisiologi Tumbuhan. Raja Grafindo Persada. Jakarta.
Lee, J.M. 1992. Biochemical Engineering. New Jersey: Prentice Hall
Lehninger, A.L. 1997. Dasar-Dasar Biokimia. Jilid I.Jakarta: Erlangga.
Macrae, A.R. 1983. Extracellular Microbial Lipase In Microbial Enzyme and Biotechnology.England: Ed. Fogarty W.M Applied Science Publisher Ltd.
Martoharsono, S. 1994. Biokimia. Jilid I. Yogyakarta: Gadjahmada University Press .
Moentamaria,D. 2009. Kajian Awal Pembuatan Biokatalisator Lipase Teramobil Dari Mucor Miehei Untuk Pengolahan Minyak Randu Menjadi Biodiesel.Malang:: Jurusan Teknik Kimia Politeknik Negeri Malang.
Mohammed, M.A. 2000. Distribution of Lipase in Graminae. Partial and Purification and Characterization of Esterase from Avena fature. Bioresource Technology. 73:227-234.
Muliad, D. 2009. Direktorat Produk Pertanian dan Kehutanan.
Nelson, L.A. 1996. Lipase Catalyse Production of Biodiesel. JAOC. 73 (8):1191-1195.
Permana, M. 2012. Optimasi Isolasi Lipase Indigenous Biji Kakao (Theobroma cacao L). Bali : Universitas Udayana.
Poedjiadi, A. 1994. Dasar-Dasar Biokimia. Jakarta:Universitas Indonesia Press .
Rees, A.R.1962. High Temparature pre-Treatment and Germination of The Seed of The Oil Palm in Some Enzymic Activities in the Germination Oil Palm Seedling.Plant ed Cheang, O.K. Plant Physiol.
Sennayake, S.P.J.N. dan F.Shahidi. 2000. Lipid Component of Borage (Borago officinalisL) Seed and Their Change During Germination. J Am. Oil. Chem.
Soc.77:55-61.
Siregar, N.A. 2011. Penentuan pH dan Suhu Optimum untuk Aktivitas Ekstrak Kasar Enzim Lipase dari Kecambah Biji Jarak Kepyar (Ricinus communis L) terhadap Hidrolisis Minyak Wijen. Medan: Universitas Sumatera Utara.
Sri, W.M. 2011. Produksi, Karakterisasi, dan Isolasi Lipase dari Aspergillus niger. Yogyakarta: Seminar Nasional Teknik Kimia.
Stumpf, P.K. 1983. Some Enzyme Activity in the Germinating Oil Palm. California: Plant Physiol. Hal 1029 .
Sutopo, L. 2002. Teknologi Benih. Fakultas Pertanian.Malang:Universitas Brawijaya.
Swadaya, P. 2001. Kelapa Sawit, Usaha Budidaya, Pemanfaatan Hasil dan Aspek Pemasaran. Jakarta:Penebar Swadaya.
Tranggono, B.S. 1989. Petunjuk Laboratorium Biokimia Pangan. Yogyakarta: Pusat Antar Universitas Pangan dan Gizi.
Williamson,K.L & L.F.Fieser. (1992). Organic Experiment 7th
Winarno, 1983. Enzim Pangan.PT Jakarta: Gramedia.
Edition. United States
of America: D C Health and Company.
Lampiran 1. Perhitungan Pembuatan Buffer Fosfat 0,05 M
Dipakai Rumus :
pH = pKa + log
Untuk pH = 6,0
6,0 = 7,2 + log
log = - 1,2
=
% = x 100%
= x 100%
= 5,93 %
% = x 100%
= x 100%
= 94,07 %
g = % x M x BM
= 0,0593 x 0,05 x 142
= 0,421 g/L
g = % x M x BM
= 0,9407 x 0,05 x 138
Lampiran 2. Perhitungan Kadar ALB (Asam Lemak Bebas) Hasil hidrolisis
RBDPO Oleh Crude Enzim Lipase
Lampiran 3. Perhitungan Aktivitas Ekstrak Kasar Enzim Lipase
Aktivitas crude enzim lipase dihitung berdasarkan metode Linfield (1984) yaitu :
Aktivitas lipase (U/mL)=
60
1000 )
(A−B ×NKOH×
Dengan : A = mL KOH untuk titrasi substrat, B = mL KOH untuk titrasi blanko,
1000= konversi dari mmol ke µ mol dan 60 = waktu reaksi (1 jam = 60 menit) dan 1
Unit aktivitas enzim adalah jumlah yang menyebabkan pengubahan 1 µ mol (10-6mol
substrat per menit.
Maka aktivitas crude enzim lipase pada suhu 40ºC dengan A = 6,7 mL, B = 0,8 mL,
Lampiran 4. Kelapa Sawit dan Hasil Penelitian
4.1 Tumbuhan Kelapa Sawit 4.2 Biji Kelapa Sawit
4.3 Kecambah Biji Sawit 1 hari 4.4 Kecambah Biji Sawit 14 hari