RESPONS CENDAWAN MIKORIZA ARBUSKULA
PADA
AQUILARIA SPP
.
INDRIAT Y
PROGRAM STUDI BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
INDRIATY. Respons Cendawan Mikoriza Arb uskula pada Aquilaria spp. Dibimbing oleh AGUSTIN WYDIA GUNAWAN dan MAMAN TURJAMAN.
Aquilaria malaccensis dan A. microcarpa merupakan tanaman penghasil gaharu yang pengadaannya dalam bentuk bibit perlu ditingkatkan. Cendawan mikoriza arbuskula (CMA) diharapkan mampu meningkatkan pertumbuhan bibit. Penelitian ini bertujuan mengetahui respons CMA pada bibit A. malaccensis dan A. microcarpa. Inokulum CMA yang digunakan ialah Glomus
sp. ACA, Glomus sp. ZEA, Glomus clarum, Entrophospora sp., dan Gigaspora sp. Kolonisasi CMA pada akar A. malaccensis dan A. microcarpa terjadi sebelum minggu ke-7 setelah inokulasi. Inokulasi CMA efektif meningkatkan tinggi bibit , diameter batang, dan bobot kering total A. malaccensis dan A. microcarpa.
ABSTRACT
INDRIATY. Resp onses Arbuscular Mycorrhizal Fungi in Aquilaria spp.Supervised by AGUSTIN WYDIA GUNAWAN and MAMAN TURJAMAN.
Aquilaria malaccensis and A. microcarpa are the main source of gaharu which their seedlings production needs to be increased. One of the potential methods that can optimize the growth of seedlings is arbuscular mycorrhizal fungi (AMF). The objectived of this research is to study response of AMF in A. malaccensis and A. microcarpa seedlings. The AMF inoculants which used in this research were Glomus sp. ACA, Glomus sp. ZEA, Glomus clarum, Entrophospora sp., and
Gigaspora sp. After AMF inoculation, colonization in roots of A. malaccensis and A. microcarpa
RESPONS CENDAWAN MIKORIZA ARBUSKULA PADA
AQUILARIA SPP.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains pada Fakultas MIPA
Institut Pertanian Bogor
INDRIATY
PROGRAM STUDI BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul Skripsi : RESPONS CENDAWAN MIKORIZA ARBUSKULA PADA AQUILARIA SPP.
Nama : Indriaty
NIM : G34101020
Menyetujui,
Pembimbing I Pembimbing II
Ir. Agustin Wydia Gunawan, M.S. Ir. Maman Turjaman, DEA. NIP. 130367078 NIP. 710020697
Mengetahui,
Dekan Fakultas MIPA
Dr. Ir. Yonny Koesmaryono , M.S. NIP. 131473999
PRAKATA
Puji dan syukur penulis panj atkan ke hadirat Allah SWT yang telah melimpahkan rahmat, kesehatan, serta kekuatan sehingga penulis dapat menyelesaikan karya ilmiah dengan judul Respons Cendawan Mikoriza Arbuskula pada Aquilaria spp.
Pada kesempatan ini, penulis ingin mengucapkan terima kasih kepada Ibu Ir. Agustin Wydia Gunawan, M.S. dan Bapak Ir. Maman Turjaman, DEA. yang telah membantu memberikan bimbingan dan saran selama penelitian dan penyusunan karya ilmiah ini. Penulis mengucapkan terima kasih kepada ibu Dra. Triadiati, M .Si. selaku dosen penguji yang banyak memberi masukan atas karya ilmiah ini, kepada Kepala Laboratorium Mikrobiologi Tanah Puslitbang Hutan, dan Konservasi Alam, Bogor Bapak Dr. Ir. Erdi Santoso atas perizinan penelitian ini. Ungkapan terima kasih juga disampaikan kepada staf Laboratorium Mikrobiologi Hutan dan teman-teman dari Universitas Padjajaran (Wulan, Nopi, Gamar, Nina, Rini, dan Lidia) atas bantuan dan kerjasama yang telah terjalin dengan baik selama penelitian, kepada temanku Dian, Nani, Ani, Feni, Iir, Dedek, Nopi, Ritma, dan mbak Rida atas informasi, pengetahuan, serta dukun gannya.
Penulis tidak lupa untuk mengucapkan terima kasih kepada keluarga tercinta Mama, Ayah, Desi, Asep, Ika, dan I ki atas doa, cinta, dan kasih sayang yang tak pernah terputus.
Akhirnya penulis berharap agar karya ilmiah ini bermanfaat bagi perkembangan ilmu pengetahuan .
Bogor, Februari 2006
RIWAYAT HIDUP
Penulis dilahirkan di Banda Aceh pada tanggal 19 Maret 1983 sebagai anak sul ung dari lima bersaudara, putri dari pasangan Mahmud Saleh dan Khadijah Zeha.
DAFTAR ISI
Halaman
DAFTAR TABEL ... vii
DAFTAR GAMBAR ... vii
DAFTAR LAMPIRAN ... viii
PENDAHULUAN ... 1
BAHAN DAN METODE ... 1
HASIL ... 2
PEMBAHASAN ... 6
SIMPULAN ... 8
DAFTAR PUSTAKA ... 8
DAFTAR TABEL
Halaman 1 Persentase kolonisasi CMA pada akar A. malaccensis ... 2
2 Tinggi bibit dan diameter batang bibit A. malaccensis pada minggu ke-25 setelah
inokulasi CMA ... 3
3 Bobot kering total dan nisbah pucuk/akar bibit A. malaccensis pada minggu ke-25
setelah inokulasi CMA... 3
4 Kada r P dan N jaringan tajuk bibit A. malaccensis pada minggu ke-25 setelah
inokulasi CMA ... 4
5 Uji beda nilai tengah dua populasi berpasangan antarperlakuan CMA pada pertumbuhan bibit A. malaccensis... 4
6 Persentase kolonisasi CMA pada akar A. microcarpa... 5
7 Tinggi bibit dan diameter batang bibit A. microcarpa pada minggu ke-25 setelah
inokulasi CMA ... 5
8 Bobot kering total dan nisbah pucuk/akar bibit A. microcarpa pada minggu ke -25
setelah inokulasi CMA... 5
9 Kadar P dan N jaringan tajuk bibit A. microcarpa pada minggu ke-25 setelah
inokulasi CMA... 5
10 Uji beda nilai tengah dua populasi berpasangan antarperlakuan CMA pada pertumbuhan bibit A. microcarpa... 6
DAFTAR GAMBAR
Halaman
1
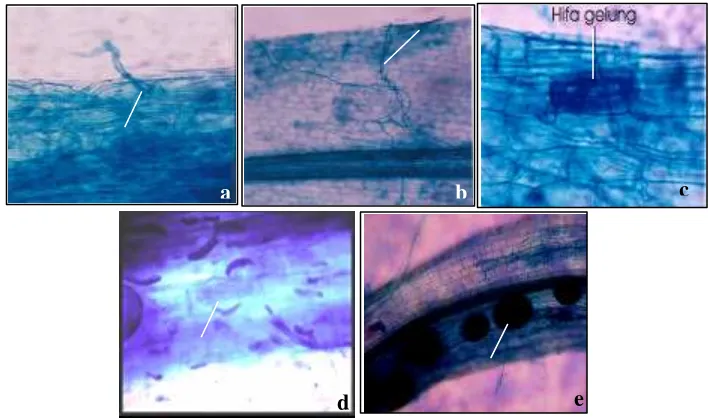
Struktur kolonisasi CMA pada akar bibit: a apresorium, b hifa internal,c hifa gelung, d arbuskula, dan e vesikula ... 3
2 Pertumbuhan Aquilaria spp . pada umur 25 minggu setelah inokulasi: a, b bibit
A. malaccensis,c, d bibit A. microcarpa, 1 kontrol, 2 perlakuan Glomus sp. ACA, 3 perlakuan Glomus sp. ZEA, 4 perlakuan G. clarum, 5 perlakuan Entrophospora sp.
DAFTAR LAMPIRAN
Halaman
1 Pewarnaan akar (Koske & Gemma 1989) dengan modifikasi... 11
2 Uji beda nilai tengah dua populasi berpasangan tinggi bibit A. malaccensis... 12
3 Uji beda nilai tengah dua populasi berpasangan diameter batang A. malaccensis... 12
4 Uji beda nil ai tengah dua populasi berpasangan bobot kering total A. malaccensis ... 12
5 Uji beda nilai tengah dua populasi berpasangan nisbah pucuk/akar A. malaccensis ... 12
6 Uji beda nilai tengah dua populasi berpasangan kadar P jaringan tajuk A. malaccensis ... 13
7 Uji beda nilai tengah dua populasi berpasangan kadar N jaringan tajuk A. malaccensis ... 13
8 Uji beda nilai tengah dua populasi berpasangan tinggi bibit A. microcarpa ... 13
9 Uji beda nilai tengah dua populasi berpasangan diameter batang A. microcarpa... 13
10 Uji beda nilai tengah dua populasi berpasangan bobot kering total A. microcarpa... 14
11 Uji beda nilai tengah dua populasi berpasangan nisbah pucuk/ akar A .microcarpa... 14
12 Uji beda nilai tengah dua populasi berpasangan kadar P jaringan tajuk A. microcarpa... 14
1
PENDAHULUAN
Hutan tropik Indonesia menyimpan sumber daya alam yang besar jumlahnya.
Aquilaria malaccensis dan A. microcarpa dari famili Thymelaceae merupakan sumber daya alam penghasil gaharu bernilai tinggi (Hou 1960). Gaharu dapat dimanfaatkan sebagai bahan dasar parfum (wewangian), bahan kosmetik, obat -obatan, dan dupa (Barden et al. 2000).
Saat ini status keberadaan A. malaccensis
dan A. microcarpa dalam keadaan terancam. Penebangan hutan dan perubahan hutan tanpa henti di Riau, Kalimantan Barat, dan Kalimantan Timur membuat populasinya menjadi sangat rendah (1 pohon ha-1) (Soehartono & Newton 2000). Perdagangan gaharu yang meningkat setiap tahunnya juga menjadi ancaman bagi keberadaan spesies sehingga keduanya dimasukkan dalam
Appendix II dari Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES ) (CITES 2004).
Akhir -akhir ini penelitian mengenai A. malaccensis dan A. microcarpa menjadi salah satu topik konservasi. Salah satu kegiatan konservasi ialah mengusahakan A. malaccensis dan A. microcarpa dalam bentuk bibit yang tumbuh optimal.Bibit yang tumbuh optimal dan siap tanam memiliki tinggi 5-20 cm dan diameter batang 0.5-2.5 cm, bebas hama penyakit, serta tahan terhadap kekeringan.
Cendawan mikoriza arbuskula (CMA) merupakan cendawan yang memiliki kemampuan bersimbiosis secara mutualisme dengan tanaman. Sejumlah besar spesies tanaman di alam secara alami bersimbiosis dengan CMA. Gonystylus bancanus dari famili Thymelaceae diketahui dapat bersimbiosis dengan CMA (Mui n 2003). Sehingga s angat dimungkinkan Aquilaria
yang berasal dari satu famili dengan
Gonystylus juga bersimbiosis dengan CMA. Beberapa penelitian membuktikan bahwa cendawan mikoriza arbuskula berperan dalam penyerapan hara khususnya fosfor (P) yang tidak tersedia bagi tanaman sehingga menjadi tersedia. Fosfor dan nitrogen (N) merupakan hara makro yang penting untuk pertumbuhan tanaman. Penyerapan P selalu diiringi dengan penyerapan N (Smith & Read 1997). Penelitian ini bertujuan mengetahui respons CMA terhadap pertumbuhan A. malaccensis
dan A. microcarpa.
BAHAN DAN METODE
Waktu dan Tempat. Penelitian ini dilaksanakan pada bulan Februari sampai dengan Oktober 2005 di Laboratorium Mikrobiologi Hutan dan Rumah Kaca, Pusat Penelitian dan Pengembangan (Puslitbang) Hutan dan Konservasi Alam, Bogor serta Laboratorium Mikologi, Institut Pertanian Bogor (IPB).
Bahan. Bibit terdiri atas A. malaccensis
(dari Bangka) dan A. microcarpa (dari Kalimantan Timur) berumur satu bulan dengan tinggi rata-rata 2.6 cm, diameter 0.9 mm, dan memiliki 2-3 helai daun. Inokulum CMA yang digunakan berasal dari Laboratorium Mikrobiologi Hutan, Puslitbang Hutan dan Konservasi Alam, Bogor. Inokulum tersebut berupa zeolit yang bercampur dengan spora, hifa, dan akar terkolonisasi CMA. Ada lima jenis inokulum CMA uji, yaitu Glomus sp. ACA, Glomus sp. ZEA, Glomus clarum, Entrophospora sp., dan
Gigaspora sp. Media tanam berupa tanah latosol pH 5.6 dengan kandungan N total 0.12% (sangat rendah) dan P tersedia 5.6 ppm (sangat rendah) (Rosma rkam & Yowono 2002). Tanah terlebih dahulu disterilkan dengan autoklaf yang bertekanan 15 psi pada suhu 121 oCselama satu jam. Tanah diambil dari lahan Puslitbang Hutan dan Konservasi Alam, Bogor.
Pelaksanaan Inokulasi Cendawan Mikoriza Arbuskula dan Pemeliharaan Bibit. Pot plastik berukuran 6 cm x 6 cm x 15 cm diisi 300 g media tanah steril, kemudian dibuat sumur dan 10 g inokulum CMA dimasukkan ke dalamnya. Selanjutnya, bibit ditanam dan sumur ditutup tanah kembali. Masing-masing unit perlakuan dalam sat u pot plastik merupakan kombinasi satu jenis
Aquilaria dan satu jenis CMA, sehingga diperoleh 10 kombinasi perlakuan CMA. Sebagai kontrol, media tanah dalam pot plastik tidak diberi inokulum CMA. Pemeliharaan meliputi penyiraman setiap hari, pembersihan gulma, dan pembersihan serangga pengganggu.
Rancangan Perlakuan. Setiap
2
jaringan tajuk bibit . Data pertumbuhan bibit, kadar P dan N bibit diuji secara statistika menggunakan uji beda nilai tengah dua populasi berpasangan pada taraf 1%.
Pengamatan. Pengamatan persentase kolonisasi dan struktur CMA pada akar sekunder dilakukan pada minggu ke -7, 11, 15, dan 25 setelah inokulasi. Akar dicuci dengan air lalu dipotong-potong sepanjang satu sentimeter. Contoh akar diwarnai menggunakan biru tripan mengikuti metode Koske & Gemma (1989) dengan modifikasi (Lampiran 1). Pemeriksaan terhadap contoh akar dilakukan menggunakan mikroskop cahaya pada perbesaran 100-400 kali. Struktur kolonisasi CMA yang diamati meliputi apresorium, hifa internal, hifa gelung, arbuskula, dan vesikula. Persentase kolonisasi CMA dihitung dengan rumus:
Oby ek akar ber-CMA Total obyek akar yang diamati
Peubah pertumbuhan yang diamati antara lain tinggi bibit, diameter batang, bobot kering total, nisbah pucuk/akar, kadar P jaringan tajuk bibit, dan kadar N jaringan tajuk bibit. Tinggi dan diameter batang diukur pada minggu ke-25 setelah inokulasi. Bibit umur 25 minggu setelah inokulasi dikeringkan dalam oven bersuhu 60 ºC sampai diperoleh bobot kering total konstan (± 4 hari). Analisis kadar P jaringan tajuk bibit mengikuti metode pengabuan kering dan kadar N total jaringan tajuk bibit menggunakan metode Kjeldahl. Analisis P dan N dilaksanakan di Laboratorium Tanah, Fakultas Pertanian, IPB.
HASIL
Kolonisasi Cendawan Mikoriza Arbuskula pada Akar A. malaccensis. Kolonisasi CMA pada akar A. malaccensis
dimulai sebelum minggu ke-7 setelah inokulasi. Simbiosis yang terjadi berkembang terus dan pada minggu ke-25 mencapai lebih
besar dari 50% dan bahkan pada perlakuan
Glomus sp. ACA dan Entrophospora sp. meningkat hingga 84% dan 93%. Bibit A. malaccensis kontrol pada minggu ke-25 dikolonisasi CMA sebesar 6% (Tabel 1).
Struktur CMA yang diamati meliputi apresorium, hifa internal, hifa gelung, arbuskula, dan vesikula (Gambar 1). Apresorium dapat diamati pada setiap minggu pengamatan, namun pada minggu ke-25 struktur ini hampir tidak teramati. Hifa internal tampak pada seluruh minggu pengamatan. Hifa gelung tampak jelas mengisi ruang sel korteks akar. Struktur arbuskula termasuk struktur yang sulit diamati. Kedua struktur ini tampak pada minggu ke-11, 15, dan 25. Vesikula juga tampak pada minggu ke-11, 15, dan 25 pada seluruh perlakuan termasuk yang mendapat perlakuan Gigaspora sp.
Respons Cendawan Mikoriza Arbuskula pada Pertumbuhan Bibit A. malaccensis. Inokulum CMA secara nyata meningkatkan tinggi bibit A. malaccensis
berkisar dari 52% sampai 87% dan diameter batang berkisar dari 41% sampai 59% dibandingkan dengan kontrol (Tabel 2).
Inokulum CMA secara nyata juga meningkatkan bobot kering total bibit A. malaccensis berkisar dari 267% sampai 333% dan nisbah pucuk/akar berkisar dari 42% sampai 74% dibandingkan dengan kontrol (Tabel 3).
Kadar P jaringan tajuk bibit A. malaccensis nyata meningkat pada pemberian inokulum Glomus sp. ZEA dan
Entrophospora sp. dibandingkan dengan kontrol. Sedangkan, kadar N jaringan tajuk bibit secara nyata meningkat hanya pada perlakuan Gigaspora sp. (Tabel 4). Hasil perbandingan satu persatu antarperlakuan CMA disajikan pada Tabel 5, dan lebih jelasnya dapat dilihat pada Lampiran 2 hingga 7.
Tabel 1 Persentase kolonisasi CMA pada akar A. malaccensis
3
Gambar 1 Struktur kolonisasi CMA pada akar bibit: a apresorium, b hifa internal, c hifa gelung, d arbuskula, dan e vesikula.
Tabel 2 T inggi bibit dan diameter batang bibit A. malaccensis pada minggu ke-25 setelah inokulasi CMA
** Beda sangat nyat a pada uji beda nilai tengah dua populasi berpasangan setiap inokulum CMA dan kontrol pada taraf uji 1%; dan #dibandingkan dengan kontrol .
Tabel 3 Bobot kering total dan nisbah pucuk/akar bibit A. malaccensis pada minggu ke-25 setelah inokulasi CM A
* Beda nyata, ** beda sangat nyata pada uji beda nilai tengah dua populasi berpasangan setiap inokulum CMA dan kontrol pada taraf uji 1%; dan #dibandingkan dengan kontrol .
Tinggi bibit Diameter batang Perlakuan
cm %# cm %#
Kontrol 10.6 ± 1.3 0 1.7 ± 0.1 0
Glomus sp. ACA 16.1 ± 0.7** 52 2.7 ± 0.2** 59
Glomus sp. ZEA 17.8 ± 1.7** 68 2.6 ± 0.2** 53
G. clarum 19.8 ± 1.7** 87 2.7 ± 0.2** 59
Entrophospora sp. 18.9 ± 0.8** 78 2.6 ± 0.1** 53
Gigaspora sp. 18.4 ± 1.0** 74 2.4 ± 0.2** 41
Bobot kering total Perlakuan
g %#
Nisbah pucuk/akar %#
Kontrol 0.3 ± 0.04 0 1.9 ± 0.2 0
Glomus sp. ACA 1.2 ± 0.09** 300 3.3 ± 0.3** 74
Glomus sp. ZEA 1.1 ± 0.1** 267 3.1 ± 0.4* 63
G. clarum 1.3 ± 0.1** 333 3 ± 0.2 ** 58
Entrophospora sp. 1.2 ± 0.2 * 300 2.7 ± 0.2* 42
Gigaspora sp. 1.2 ± 0.1** 300 2.8 ± 0.3* 47
a
b
c
4
Tabel 4 Kadar P dan N jaringan tajuk bibit A. malaccensis pada minggu ke-25 setelah inokulasi CMA
* Beda nyata, ** beda sangat nyata, tn tidak nyata pada uji beda nilai tengah dua populasi berpasangan setiap inokulum CMA dan kontrol pada taraf uji 1%; dan #dibandingkan dengan kontrol .
Tabel 5 Uji beda nilai tengah dua populasi berpasangan antarperlakuan CMA pada pertumbuhan bibit A. malaccensis
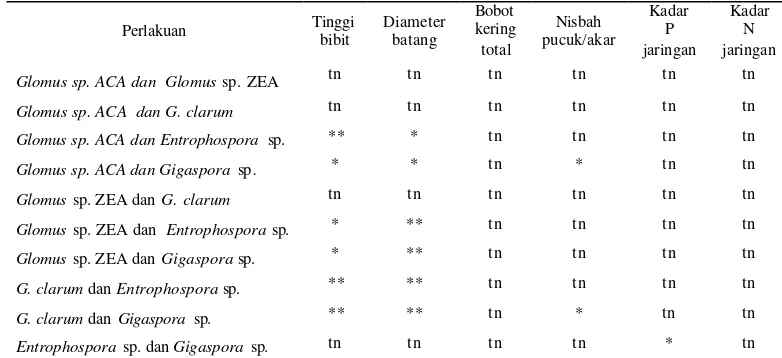
*Beda nyata, ** beda sangat nyata, tn:tidak nyata pada uji beda nilai tengah dua populasi berpasangan antarperlakuan CMA pada taraf uji 1%.
Kolonisasi Cendawan Mikoriza Arbuskula pada Akar A. microcarpa . Kolonisasi CMA yang terjadi pada akar bibit
A. microcarpa sama halnya pada bibit A. malaccensis , dimulai sebelum minggu ke-7 setelah inokulasi dan hingga minggu ke-25 mencapai lebih besar dari 70% (Tabel 6). Apresorium dan arbuskula terlihat pada akar bibit A. microcarpa pada minggu ke-7 setelah inokulasi. Pada minggu ke -11 dan 15 kelima struktur CMA yang diamati tampak pada hampir seluruh perlakuan CMA.
Respons Cendawan Mikoriza
Arbuskula pada Pertumbuhan Bibit A.
microcarpa. Seperti halnya pada bibit A. malac censis, inokulum CMA secara nyata
meningkatkan tinggi dan diameter batang bibit
A. microcarpa dibandingkan dengan kontrol (Tabel 7).
Inokulasi CMA secara nyata juga meningkatkan bobot kering total bibit A. microcarpa dibandingkan dengan kontrol, namun ha nya Entrophospora sp. dan
Gigaspora sp. yang meningk atkan nisbah pucuk/akar (Tabel 8). Perlakuan CMA tidak meningkatkan kadar P dan N jaringan tanaman A. microcarpa (Tabel 9). Hasil perbandingan satu persatu antarperlakuan CMA disajikan pada Tabel 10, dan lebih jelasnya dapat dilihat pada Lampiran 8 hingga 13.
5
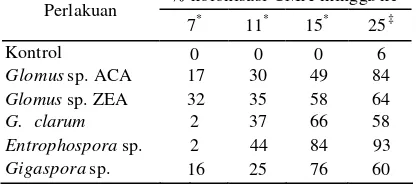
Tabel 6 Persentas e kolonisasi CMA pada akar A. microcarpa
% kolonisasi CMA minggu ke - Perlakuan
7* 11* 15* 25‡
Kontrol 0 0 0 0
Glomus sp. ACA 6 30 33 89
Glomus sp. ZEA 25 46 85 76
G. clarum 4 6 53 71
Entrophospora sp. 7 31 74 93
Gigaspora sp. 23 84 65 74
*
Rata-rata dari 2 ulangan, dan ‡ rata-rata dari 5 ulangan.
Tabel 7 T inggi bibit dan diameter batang bibit A. microcarpa pada minggu ke-25 setelah inokulasi CMA
* Beda nyata, ** beda sangat nyata pada uji beda nilai tengah dua populasi berpasangan setiap inokulum C MA dan kontrol pada taraf uji 1 %; dan # dibandingkan dengan kontrol .
Tabel 8 Bobot kering total dan nisbah pucuk/akar bibit A. microcarpa pada minggu ke-25 setelah inokulasi CMA
Bobot kering total Perlakuan
g %# Nisbah pucuk/akar %
#
Kontrol 0.3 ± 0.03 0 2.4 ± 0.2 0
Glomus sp. ACA 1.2 ± 0.3 b* 300 2.6 ± 0.2 t n 8
Glomus sp. ZEA 1.6 ± 0.2 c** 433 2.4 ± 0.3 tn 0
G. clarum 1.4 ± 0.3 b* 367 2.5 ± 0.2 tn 4
Entrophospora sp. 1.4 ± 0.08 c** 367 3.1 ± 0.2* 29
Gigaspora sp. 2 ± 0.3 c** 567 3.3 ± 0.1 ** 38
* Beda nyata, ** beda sangat nyata, tn tidak nyata pada uji beda nilai tengah dua populasi berpasangan setiap inokulum CMA dan kontrol pada taraf uji 1%; dan # dibandingkan dengan kontrol .
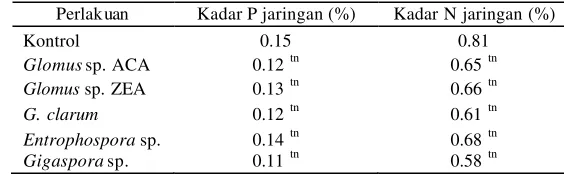
Tabel 9 K adar P dan N jaringan tajuk bibit A. microcarpa pada minggu ke-25 setelah inokulasi CMA
Perlakuan Kadar P jaringan (%) Kadar N jaringan (%)
Kontrol 0.15 0.81
Glomus sp. ACA 0.12 tn 0.65 tn
Glomus sp. ZEA 0.13 tn 0.66 tn
G. clarum 0.12 tn 0.61 tn
Entrophospora sp. 0.14 tn 0.68 tn
Gigaspora sp. 0.11 tn 0.58 tn
tn
Tidak nyata pada uji beda nilai tengah dua populasi berpasangan setiap inokulum CMA dan kontrol pada taraf uji 1%. Tinggi bibit Diameter batang Perlakuan
cm %# cm %#
Kontrol 9.5 ± 0.7 0 1.7 ± 0.1 0
Glomus sp. ACA 13.5 ± 1.4* 42 2.5 ± 0.2** 47
Glomus sp. ZEA 14.7 ± 1.4** 55 2.4 ± 0.2** 41
G. clarum 12.8 ± 1.3* 35 2.3 ± 0.3* 35
Entrop hospora sp. 19.4 ±1.1** 104 3.2 ± 0.2** 88
6
Tabel 10 Uji beda nilai tengah dua populasi berpasangan antarperlakuan CMA pada pertumbuhan bibit A. microcarpa
*Beda nyata, ** beda sangat nyata, tn:tidak nyata pada u ji beda nilai tengah dua populasi berpasangan antarperlakuan CMA pada taraf uji 1%.
PEMBAHASAN
Kolonisasi CMA pada akar A. malaccensis dan A. microcarpa terjadi sebelum minggu ke-7 setelah inokulasi. Hasil pengamatan dapat dilihat bahwa asosiasi CMA dengan akar bibit terjadi ketika hifa kontak dengan permukaan epidermis akar membentuk apresorium. Awal kolonisasi CMA pada akar bibit kelap a sawit dilaporkan terjadi pada minggu ke-7 setelah inokulasi (Widiastuti et al. 2005), sedangkan pada akar bawang daun (Allium porum) terjadi pada hari kedua setelah inokulasi (Brundrett et al. 1985). Kolonisasi CMA pada tanaman tahunan memerlukan waktu lebih lama dibandingkan dengan tanaman semusim. Keadaan ini kemungkinan disebabkan pertumbuhan akar tanaman tahunan lebih lambat dibandingkan dengan tanaman semusim.
Pada akar A. malaccensis struktur arbuskula terlihat pada minggu ke-11 setelah inokulasi sedangkan pada akar A. microcarpa
pada minggu ke-7 setelah inokulasi. Hadirnya struktur arbuskula menandakan mulai terjadinya simbiosis fungsional. Pada kelapa sawit struktur arbuskula tampak pada minggu ke-15 setelah inokulasi (Widiastuti 2004), sedangkan pada bawang daun pada hari ke-4 setelah inokulasi (Brundrett et al. 1985). Struktur arbuskula berperan dalam transfer nutrisi dua arah (Smith & Read 1997). Transfer nutrisi dua arah antara cendawan dan inang disebabkan adanya aktivitas H+/ATPase
yang terdapat pada membran periarbuskula (Smith & Read 1997). Struktur arbuskula sukar diamati karena umurnya yang relatif singkat sekitar 7 hari, lalu meluruh (Brundrett
et al. 1985).
Kecepatan kolonisasi CMA pada akar bibit berbeda-beda (Tabel 1 dan 6). Keberhasilan kolonisasi CMA bergantung pada interaksi antara tanah, cendawan, dan tanaman inang (Smith & Read 1997). Media tanah untuk percobaan memiliki kadar P dan N yang sangat rendah sehingga memungkinkan bibit segera kontak dengan CMA. Selain itu kesesuaian, kebergantungan bibit dengan CMA, serta jarak antara propagul infektif CMA dengan akar juga ikut berpengaruh.
Kontaminasi terjadi pada akar bibit kontrol dengan terlihatnya hifa CMA (Tabel 1) dan perlakuan Gigaspora sp. dengan terlihatnya struktur vesikula. Vesikula seharusnya tidak ditemukan pada perlakuan
Gigaspora sp. Menurut Smith dan Read (1997) ada dua genus CMA yaitu Gigaspora
dan Scuttelospora yang tidak membentuk vesikula. Kontaminasi berpeluang besar terjadi saat tanaman berada di area terbuka seperti halnya di rumah kaca saat penelitian berlangsung. Kontaminasi terjadi melalui spora yang terbawa air pada saat hujan atau penyiraman dan melalui angin.
Kadar P dan N jaringan tajuk kedua tanaman hampir tidak memperlihatkan peningkatan yang nyata (Tabel 4 dan 9). Hal ini menunjukkan bahwa inokulum CMA tidak
7
berpengaruh dalam meningkatkan metabolisme P dan N tanaman. Hanya perlakuan Gigaspora sp. yang mampu meningkatkan P dan N tanaman A. microcarpa dibandingkan dengan CMA lainnya. Metabolisme P di dalam jaringan tanaman meliputi pembentukan ATP, asam nukleat, fosfolipida, dan fitin sedangkan metabolisme N meliputi pembentukan asam amino, enzim, protein, dan amida (Sa lisbury & Ross 1995). Kadar P dan N jaringan tajuk bibit yang rendah juga kemungkinan disebabkan karena hara P dan N yang diserap oleh hifa eksternal masih ditahan di daerah akar. Triwahyuni dan Rohayati (2000 ) menyatakan tanaman jati yang diinokulasi dengan beberapa jenis CMA memiliki kadar P jaringan tajuk tanaman yang lebih rendah dibandingkan dengan tanaman yang tidak diinokulasi CMA. Sementara itu, Douds dan Schenck (1990) melaporkan bahwa tanaman
Paspalum notatum yang diinokulasi dengan CMA memiliki kadar N jaringan akar yang lebih tinggi dibandingkan dengan kadar N jaringan tajuknya.
Inokulum CMA meningkatkan bobot kering total bibit A. malaccensis mencapai 300% dan A. microcarpa mencapai lebih dari lebih dari 500% dibandingkan dengan kontrol (Tabel 3 dan 8). Sekitar 85-90% produksi asimilat tanaman yang diperlihatkan dari bobot kering bibit berasal dari hasil fotosintesis (Kolek dan Kojinka 1992). Hal ini dapat diartikan bahwa aktivitas fotosintesis pada bibit yang diinokulasi CMA tergolong tinggi. Tingginya aktivitas fotosintesis karena unsur hara P dan N yang diperlukan untuk prosesnya cukup tersedia di jaringan tanaman. M eskipun kadar hara P dan N tanah rendah namun penyerapan hara oleh bibit berlangsung efisien. Hal ini terjadi karena adanya bantuan dari hifa eksternal CMA. Perangkat absorsi hara selain akar tanaman menjadi bertambah dengan kehadiran hifa eksternal CMA.
Hifa eksternal CMA mampu menjelajah sejumlah besar volume tanah serta memperbaiki difusi P anorganik yang lambat di tanah (Schachtman et al. 1998). Hifa eksternal CMA menghasilkan asam organik yang dapat melarutkan P dari bentuk yang tidak tersedia (P-Al, P-Fe dan P-Ca) menjadi tersedia, dan melarutkan P yang terjerap dalam struktur tanah (Yassir 2005). Sastrahidayat et al. (1999) melaporkan bahwa akar tanaman yang terkolonisasi CMA menghasilkan enzim fosfat ase yang membantu mengkatalis hidrolisis kompleks fosfor yang
tidak larut dalam tanah, sehingga terjadilah peningkatan P yang tersedia pada daerah tersebut. Akar terkolonisasi CMA juga meningkatkan aktivitas glutamin sintase (Smith & Read 1997). Glutamin sintase tersebut berperan pada penggabungan N anorganik ke dalam komponen organik dan sebagai prekusor terbentuknya asam amino (Sharma & Johry 2002). Enzim tersebut aktif di hifa internal CMA (Gianinazzi-Pearson et al. 1997).
Peran CMA meningkatkan fotosintesis juga terlihat dari pertumbuhan tinggi bibit dan diameter batang bibit A. malaccensis dan A. microcarpa yang lebih besar dibandingkan dengan kontrol (Gambar 2). Hal ini karena asimilat yang dihasilkan dari aktivitas fotosintesis terus digunakan untuk pembelahan sel dan perkembangan jaringan meristem tanaman. Besarnya diameter batang memungkinkan tanaman mengangkut air dan garam mineral lebih banyak guna keperluan fotosintesis. Gejala defisiensi pada bibit A. malaccensis dan A. microcarpa terkolonisasi CMA tidak terjadi walaupun kadar hara P dan N sangat rendah di tanah.
Mulida (1999) melaporkan tanaman A. malaccensis yang ditumbuhkan pada media tanah latosol hingga umur 34 minggu memiliki tinggi rata-rata 14.2 cm dan diameter batang rata-rata 2.19 mm. Dalam penelitian ini penggunaan inokulum CMA menghasilkan bibit A. malaccensis dengan tinggi berkisar dari 16.1 cm hingga 19.8 cm dan diameter batang rata-rata 2.6 mm pada umur 25 minggu setelah inokulasi dan pada A. microcarpa
menghasilkan tinggi berkisar dari 12.8 hingga 19.4 cm dan diameter batang berkisar dari 2.3 mm hingga 3.5 mm (Tabel 2 dan 7). Hal tersebut menunjukkan bahwa bibit siap tanam bisa diperoleh lebih cepat dibandingkan dengan tanaman yang tidak diinokulasi CMA.
8
proses simbiosis untuk meningkatkan pertumbuhan tanaman.
Berdasarkan pada hasil pengamatan kedua tanaman diperoleh bahwa seluruh perlakuan CMA uji pada A. malaccensis tidak berbeda dalam meningkatkan tinggi bibit, diameter batang, bobot kering total , dan nisbah pucuk /akar (Tabel 5). Pada A. microcarpa jenis inokulum Gigaspora sp.
dan Enthropospora sp. menghasilkan tinggi bibit, diameter batang, bobot kering total, dan nisbah pucuk/akar paling baik (Tabel 10) dibandingkan dengan jenis CMA lainnya. Cendawan mikoriza arbuskula yang telah berkolonisasi pada bibit selanjutnya akan membantu bibit dalam penyerapan hara saat di lapangan.
Gambar 2 Pertumbuhan Aquilaria spp. pada umur 25 minggu setelah inokulasi: a, b bibit A. malaccensis, c, d bibit A. microcarpa, 1 kontrol, 2 perlakuan Glomus sp. ACA, 3 perlakuan Glomus sp. ZEA, 4 perlakuan G. clarum, 5 perlakuan Entrophospora sp., dan 6 perlakuan Gigaspora sp.
SIMPULAN
Pemberian inokulum Glomus sp. ACA,
Glomus sp. ZEA, G. clarum, Entrophospora
sp., dan Gigaspora sp. dapat meningkatkan tinggi bibit, diameter batang, dan bobot kering total bibit A. malaccensis dan A. microcarpa
pada minggu ke -25 setelah inokulasi.
DAFTAR PUSTAKA
Barden A, Anak N A, Mulliken T, Song M. 2000. Heart of the matter agarwood use and trade and cites implementation for
Aquilaria malaccensis. Cambridge: TRAFFIC International.
Brundrett M C, Piche Y, Piterson R L. 1985. A new method observing the morphology of vesicular-arbuscular mycorrhizae. Can J Bot 63:184 -193.
[CITES 2004]. 2004. Convention on International Trade in Endangered Species of Wild Fauna and Flora. http://www.cites.org/common/cop/13/raw _props/ID -Aquilaria-Gyrinops.pdf. [1 Mei 2005].
9
Gianinazzi-Pearson V, Dumas -Gaudot E, Gianinazzi S. 1997. Proteins and protein activities in endomycorrhizal symbioses. Di dalam: Varma A, Hock B, editor.
Mycorrhiza Structure, Function, Molecular Biology and Biotechnology. Ed ke-2. Berlin: Springer. hlm 255-272.
Hou D. 1960. Thymeleaceae. Di dalam: Van Steenis, editor. Flora malesiana. Volume ke-6. Netherlands: Wolter-Noordhoff.
Kolek J, Kojinka V. 1992. Physiology of the Plant Root System. Netherland: Kluwer Ac Pub.
Koske R E, Gemma J N. 1989. A Modified procedure for staining roots to detect VA mycorrhizas. Mycol Res 92:486-505.
Muin A. 2003. Pertumbuhan anakan ramin (Gonystylus bancanus (Miq.) Kurz) dengan inokulasi cendawan mikoriza arbuskula pada berbagai intensitas cahaya dan dosis fosfat alam [disertasi]. Bogor: Sekolah Pasca Sarjana, Institut Pertanian Bogor.
Mulida L. 1999. Pengaruh intensitas cahaya dan persaingan terhadap pertumbuhan semai gaharu (Aquilaria malaccensis
LAMK) pada tanah latosol dan podsolik merah kuning [skripsi]. Fakultas Kehutanan, Institut Pertanian Bogor.
Rosmarkam A, Yuwono M Y. 2002. Ilmu Kesuburan Tanah. Yogyakarta: Kanisius.
Salisbury F B, Ross C W. 1995. Fisiologi
Tumbuhan Jilid 2. Lukman R D,
Sumaryono, penerjemah. Bandung: Penerbit ITB. Terjemahan dari: Plant Physiology.
Sastrahidayat I R, Wakidah K, Syekhfani. 1999. Pengaruh mikoriza arbuskula terhadap peningkatan enzim fosfatase, beberapa asam organik dan pertumbuhan kapas (Gossypium hirsutum L.) pada vertisol dan alfisol. Agrivita 21:60-64.
Schachtmant D P, Reid R J, Anyling S M. 1998. Phosphorus uptake by plants: from soil to cell. Plant Physiol 116:447-453.
Sharma A K, Johry B N. 2002. Arbuscular Mycorrhizae Interact ion in Plants, Rhizosphere and Soils. Enfield: Science.
Smith S E, Read D J. 1997. Mycorrhizal Symbiosis. Ed ke-2. London: Academic Pr.
Soehartono T, Newton A C. 2000. Conserva-tion and sustainable use tropical trees in the genus Aquilaria I. Status and distribution in Indonesia. Biol Cons
96:83-94.
Triwahyuni C, Rohayati. 2000. Studi efektivi- tas jenis endomikoriza pada pembibitan jati (Tectona grandis Linn F.). Di dalam:
Pemanfaatan Cendawan Mikoriza sebagai Agen Bioteknologi Ramah Lingkungan dalam Meningkatkan Produktivitas Lahan di Bidang Kehutananan, Perkebunan, dan Pertanian di Era Milenium Baru. Prosiding Seminar Nasional Mikoriza I. Bogor , 15-16 Nov 1999. Bogor: Asosiasi Mikoriza Indonesia. hlm 154-161.
Widiastuti H. 2004. Biologi interaksi cendawan mikoriza arbuskula kelapa sawit pada tanah masam sebagai dasar pengembangan teknologi aplikasi dini [disertasi]. Bogor: Sekolah Pasca Sarjana, Institut Pertanian Bogor.
Widiastuti H, Sukarno N, Darusman L K, Goenadi D H, Smith S, Guhardja E. 2005. Tingkat kedinian infeksi Acaulospora tuberculata dan Gigaspora margarita
pada bibit kelapa sawit. J Mikrobiol Indones 10:42-44.
10
11
Lampiran 1 Pewarnaan akar ( Koske & Gemma 1989) dengan modifikasi
Modifikasi dilakukan pada beberapa hal yaitu: perlakuan dengan KOH 10% dan perendaman akar dengan KOH kembali selama 24 jam, tanpa perlakuan H2O2, perlakuan dengan HCl 2%, dan
penggunaan larutan asam laktat pada perlakuan staining dan destaining tanpa pemanasan. Akar direndam dalam KOH 10% dan dipanaskan dalam penangas bersuhu 90 oC selama 15-30 menit. Selanjutnya akar direndam dalam larutan KOH selama 24 jam, lalu dibilas dengan air. Akar kemudian direndam dalam HCl 2% selama 24 jam. Akar diwarnai dengan larutan staining
(gliserol: asam laktat: akuades dengan perbandingan 2: 2: 1)yang mengandung biru tripan 0.05%, selama 24 jam. Selanjutnya larutan pewarna dibuang dan akar disimpan dalam larutan destaining
(gliserol: asam laktat : akuades dengan perbandingan 2: 2: 1) tanpa biru tripan selama 24 jam.
Metode pewarnaan akar Koske dan Gemma (1989). Akar direndam dalam KOH 2.5% dan dipanaskan dalam penangas air bersuhu 90 oC selama 10 -30 menit lalu akar dicuci dengan air. Selanjutnya akar direndam dalam larutan H2O2 alkalin (NH4OH 20% 3 ml: H2O2 3% 27 ml)
selama 10-45 menit lalu dibilas dengan air. Akar kemudian direndam dalam HCl 1% selama 24 jam. Akar diwarnai dengan asam gliserol (gliserol 500 ml: akuades 450 ml: HCl 1% 50 ml) yang mengandung biru tipan 0.5 gram, kemudian dipanaskan dengan penangas air pada suhu 90 oC
12
Lampiran 2 Uji beda nilai tengah dua populasi berpasangan tinggi bibit A. malaccensis
Perlakuan Glomus sp.
Angka menunjukkan nilai peluang nyata, * beda nyata, dan tn: tidak nyata pada taraf 1%.
Lampiran 3 Uji beda nilai tengah dua populasi berpasangan diameter batang A. malaccensis
Perlakuan Glomus sp.
Angka menunjukkan nilai peluang nyata dan tn: tidak nyata pada taraf 1%.
Lampiran 4 Uji beda nilai tengah dua populasi berpasangan bobot kering total A. malaccensis
Perlakuan Glomus sp.
Angka menunjukkan nilai peluang nyata dan tn: tidak nyata pada taraf 1%.
Lampiran 5 Uji beda nilai tengah dua populasi berpasangan nisbah pucuk /akar A. malaccensis
Perlakuan Glomus sp.
13
Lampiran 6 Uji beda nilai tengah dua populasi berpasangan kadar P jaringan tajuk A. malaccensis
Perlakuan Glomus sp.
Angka menunjukkan nilai peluang nyata, * beda nyata, ** beda sangat nyata, dan tn: tidak nyata pada taraf 1%.
Lampiran 7 Uji beda nilai tengah dua populasi berpasangan kadar N jaringan tajuk A. malaccensis
Perlakuan Glomus sp.
Angka menunjukkan nilai peluang nyata, * beda nyata, dan tn: tidak nyata pada taraf 1%.
Lampiran 8 Uji beda nilai tengah dua populasi berpasangan tinggi bibit A. microcarpa
Perlakuan Glomus sp.
Angka menunjukkan nilai peluang nyata, * beda nyata, ** beda sangat nyata, dan tn: tidak nyata pada taraf 1%.
Lampiran 9 Uji beda nilai tengah dua populasi berpasangan diameter batang A. microcarpa
Perlakuan Glomus sp.
14
Lampiran 10 Uji beda nilai tengah dua populasi berpasangan bobot kering total A. microcarpa
Perlakuan Glomus sp.
Angka menunjukkan nilai peluang nyata dan tn: tidak nyata pada taraf 1%.
Lampiran 11 Uji beda nilai tengah dua populasi berpasangan nisbah pucuk/akar A. microcarpa
Perlakuan Glomus sp.
Angka menunjukk an nilai peluang nyata, * beda nyata, dan t n: tidak nyata pada taraf 1%.
Lampiran 12 Uji beda nilai tengah dua populasi berpasangan kadar P jaringan taj uk
A. microcarpa
Angka menunjukkan nilai peluang nyata, * beda nyata, dan tn: tidak nyata pada taraf 1%.
Lampiran 13 Uji beda nilai tengah dua populasi berpasangan kadar N jaringan tajuk
A. microcarpa