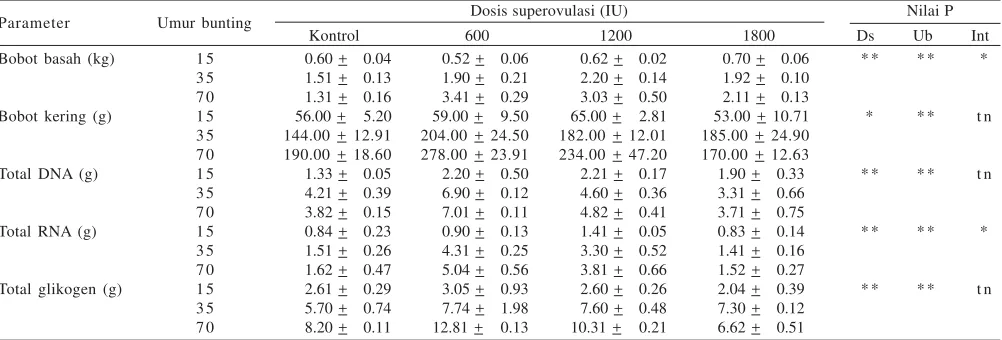
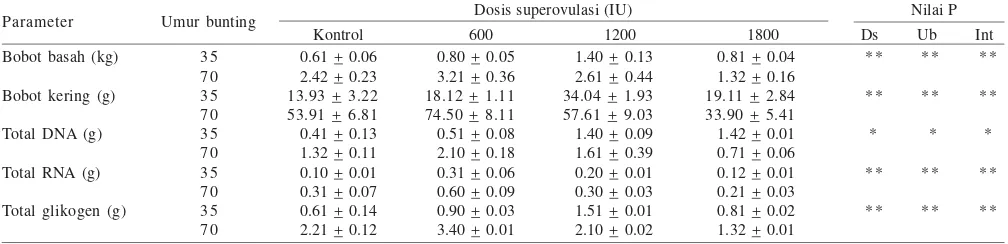
Growth and Development of the Uterus and Placenta of Superovulated Gilts
Teks penuh
Gambar
Dokumen terkait
Penelitian ini dilakukan dengan tujuan untuk mengatahui: 1) efektivitas media film dalam meningkatkan minat dan hasil belajar Ekonomi siswa, 2) perbedaan peningkatan minat
harga bahan dan upah kerja sesuai dengan kondisi setempat;.. b) Spesifikasi dan cara pengerjaan setiap jenis pekerjaan disesuaikan dengan standar. spesifikasi teknis pekerjaan
Pramuka tentang mekanisme Corporate Governance , manajemen laba dan kinerja keuangan (2007). Kesimpulan dari hasil penelitian yang telah dilakukan sebagai.. berikut: 1)
Adapun perangkat teori yang relevan untuk dijadikan landasan dalam penelitian ini adalah teori-teori tentang ikhtiar-ikhtiar yang yang dilakukan oleh Perbankan Syariah, yang
DECLARE THAT THE PARTICULARS SHOWN IN THIS FORM AND ACCOMPANYING DOCUMENTS ARE TRUE AND CORRECT IN EVERY DETAIL AND DISCLOSE A FULL AND COMPLETE STATEMENT OF TOTAL INCOME DERIVED
Berdasarkan analisis dengan menggunakan metode COBIT 4.1 yang dilakukan pada divisi administrasi RSIA Hamami menghasilkan nilai maturity pada level 2 yaitu dimana
Kuatnya pengaruh angin muson dapat dilihat dalam bentuk aliran massa air pada lapisan permukaan ke arah tenggara di sepanjang pantai barat daya Sumatera dan ke arah
Dengan demikain terwujudlah apa yang diinginkan Mas Yuma dan Mas Briliant melalui tindak tutur meminta yang disampaikan kepada lawan tuturnya secara implisit dan lawan
