PENGENDALIAN Saprolegnia sp. PADA TELUR GURAMI
(Osphronemus gouramy) MENGGUNAKAN ISOLAT
BAKTERI KITINOLITIK
TESIS
Oleh
RITA ROSMALA DEWI
097030001
PROGRAM STUDI MAGISTER BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS SUMATERA UTARA
PENGENDALIAN Saprolegnia sp. PADA TELUR GURAMI
(Osphronemus gouramy) MENGGUNAKAN ISOLAT BAKTERI
KITINOLITIK
TESIS
Diajukan Sebagai Salah Satu Syarat untuk Memperoleh Gelar
Magister Sains (M.Si) dalam Program Studi Biologi pada
Sekolah Pascasarjana Universitas Sumatera Utara
RITA ROSMALA DEWI
097030001
PROGRAM STUDI MAGISTER BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS SUMATERA UTARA
ABSTRAK
Penelitian ini bertujuan untuk mengetahui potensi 24 bakteri kitinolitik yang diisolasi dari kolam budidaya ikan gurami (Osphronemus gouramy) untuk menghambat pertumbuhan isolat Saprolegnia sp. yang diisolasi dari telur gurami. Uji antagonisme dilakukan pada media garam minimum dengan 2% koloidal kitin untuk mengetahui kemampuan isolat bakteri menghambat Saprolegnia sp. Sepuluh isolat bakteri menunjukkan daya hambat dengan diameter 10–19 mm. Abnormalitas hifa diamati secara mikroskopis meliputi lisis (40%), lisis pada ujung hifa (30%), hifa bengkok (23,33%) dan hifa terpuntir (6,67%). Uji patogenesa isolat Saprolegnia sp. dilakukan untuk mengetahui kemampuan isolat tersebut menginfeksi telur gurami. Hasil penelitian menunjukkan bahwa isolat Saprolegnia sp. patogen terhadap telur. Uji patogenesa dan perlekatan bakteri potensial dilakukan untuk mengeliminasi isolat yang bersifat patogen dan tidak dapat melakukan perlekatan terhadap mukosa sel telur. Isolat PB01 bersifat patogen bagi telur sehingga tidak dapat digunakan sebagai pengendali hayati Saprolegnia sp. Semua isolat memiliki kemampuan melekat pada sel telur ditunjukkan dengan uji reisolasi dan pengamatan Scanning Electron Microscope (SEM). Uji tantang isolat bakteri potensial menghambat infeksi Saprolegnia sp. pada telur dilakukan pada bak percobaan dan kondisi terkontrol. Hasil penelitian menunjukkan bahwa 6 isolat bakteri yaitu PB05, PB08, PB13, PB14, PB15, dan PB17 mampu menurunkan tingkat mortalitas serta meningkatkan daya tetas telur yang yang diinfeksi Saprolegnia sp. sehingga potensial untuk dijadikan agen pengendali hayati Saprolegnia sp. pada telur gurami.
ABSTRACT
A study on potential antagonist activity of 24 chitinolytic bacteria isolated from Osphronemus gouramy pond culture to inhibit the Saprolegnia sp. causal agent of saprolegniasis in gouramy egg was carried out. The ability of bacterial isolates to inhibit the Saprolegnia sp. in vitro was evaluated by antagonism assay on minimum salt medium agar with 2% colloidal chitin as C source. Hyphal abnormality as a result of antagonistic assay was examined in this study. The result showed that 10 isolates, PB3A, PB01, PB02, PB05, PB08, PB10, PB13, PB14, PB15 dan PB17 showed relatively high inhibition zone. The diameter of inhibition zone ranged from 10-19 mm. The abnormality of miselium was lysis (40%), a lised hyphae tips (30%), bent hyphae (23,33%) and twisted hyphae (6,67%). Saprolegnia sp. was examined for its ability causing infection in egg. The result showed that this isolates caused infection on gouramy egg. Pathogenecity and attachment of bacteria assay was done to eliminate pathogen isolate and to know the ability of isolates to adhere on the egg mucosa. The result showed that PB01 was pathogen and all potential bacteria had ability to adhere to egg surface. It was proved by reisolation assay and examination on Scanning Electron Microscope (SEM). Potential chitinolytic bacteria were assayed to Saprolegnia sp. in vivo using gouramy egg. Potential bacteria (PB05, PB08, PB13, PB14, PB15, and PB17) could reduce mortality rate and increase hatching rate on infected egg. Based on this study, we concluded that PB05, PB08, PB13, PB14, PB15, and PB17 had the ability to inhibit the Saprolegnia sp. these isolates could be utilized as potential biological control candidates against Saprolegniasis.
PRAKATA
Puji syukur penulis panjatkan kepada Allah SWT atas karunia-Nya sehingga tesis ini berhasil diselesaikan. Tema penelitian yang dilaksanakan sejak bulan Juli 2010 ini adalah pengendalian hayati Saprolegniasis pada telur gurami dengan judul Pengendalian Infeksi Saprolegnia sp. pada telur gurami (Osphronemus gouramy) menggunakan bakteri kitinolitik.
Terima kasih disampaikan kepada Bapak Prof. Dr. Dwi Suryanto, M.Sc. dan Bapak Dr. Ir. Yunasfi, MS. atas semua bimbingan dan arahannya dalam penelitian dan penulisan tesis ini. Bapak Prof. Dr. Erman Munir, M.Sc. dan Dr. Ir. Edy Batara Mulya Siregar, MS. sebagai penguji yang telah memberikan saran dalam penyusunan tesis ini. Penghargaan juga penulis sampaikan kepada Bapak Prof. Dr. Syafruddin Ilyas, M. Biomed selaku Ketua Pasca Sarjana Biologi. Ibu Dra. Nunuk Priyani, M.Sc. selaku Kepala Laboratorium Mikrobiologi. Staf pengajar Pasca Sarjana Biologi. Ibu Ir. Heny Budi Utari, M.Si dan Bapak Yuri Sutanto, Ph.D. selaku pimpinan Divisi Aquatic Health Centre (AHC) PT. Central Proteinaprima (CPP) yang telah memberikan saran dan motivasi. Ami Lifda Sinulingga, ST., Yudanto Tri Anggoro, S.Pi., Edrizal, S.Pi. dan Supriyadi, S.Pi dari AHC PT. CPP atas bantuannya dalam penelitian penulis. Kepada Bapak Surono dan Mardi sebagai praktisi budidaya gurami, Penulis mengucapkan terima kasih atas bantuan tempat pengambilan sampel untuk penelitian ini.
Ungkapan terima kasih tak lupa penulis ucapkan kepada semua rekan seperjuangan di Pascasarjana Biologi Angkatan 2009 terutama untuk Yuni terima kasih atas bantuannya selama kuliah hingga penelitian, Ibu Yos, Ibu Nafiah, Ibu Bunga atas bantuannya dalam penelitian ini. Asisten Laboratorium Mikrobiologi Asril, Mirza spesial untuk Yanti.
Akhirnya terima kasih penulis sampaikan kepada suami tercinta Arif Nuryawan, S.Hut. M.Si. atas perhatian, doa, pengorbanan dan kasih sayangnya. Anakku tersayang almarhum Airlangga Rafi Nurilmi yang telah memberikan pelajaran berharga buat penulis dan Adinda Aisyah Nursalsabila yang telah membuat penulis bersemangat dalam menyelesaikan studi. Kepada Ayahanda Drs. Sutarman, Ibunda Dra. Sri Murwani, Endah, Bambang, Putri, Bapak Mertua H. Suhartono, Ibu Hj. Faridah, Mbak Ati, Mas Bagus dan Nofar, penulis sampaikan terima kasih, semoga Allah SWT berkenan memberikan pahala yang akan terus mengalir.
Semoga tesis ini bermanfaat.
RIWAYAT HIDUP
Penulis dilahirkan di Kabupaten Karanganyar, Jawa Tengah 23 Maret 1979 sebagai anak pertama dari pasangan yang berbahagia ayahanda Drs. Sutarman dan Ibunda Dra. Sri Murwani.
Pendidikan Sekolah Menengah diselesaikan di SMA Negeri 1 Palembang pada tahun 1996. Sarjana ditempuh di Fakultas Kedokteran Hewan (FKH), Institut Pertanian Bogor (IPB), lulus pada tahun 2000. Program Pendidikan Dokter Hewan (PPDH) ditempuh di FKH-IPB, lulus pada tahun 2002. Pada tahun 2002 - 2007 penulis bekerja sebagai Patologist Satwa Aquatik di Animal Health Laboratory (AHL) PT. Charoen Pokphand Indonesia (CPI) di Jakarta Utara. Pada tahun 2007- 2009 penulis bekerja sebagai Laboratory and Technical Service di Aquatic Health Centre (AHC) PT. Central Proteinaprima (CPP) di Medan. Pada tahun 2009 – 2010 penulis merupakan Konsultan Profesional AHC-PT.CPP di Medan hingga pada akhirnya mengundurkan diri sementara waktu untuk menyelesaikan tugas akhir.
Pada tahun 2009 penulis melanjutkan studi pada sekolah Pasca Sarjana Universitas Sumatera Utara, Program Studi Biologi konsentrasi Mikrobiologi.
DAFTAR ISI
1.2. Pembatasan dan Perumusan Masalah ... 4
1.3. Kerangka Pemikiran ... 4
2.3. Pengendalian Saprolegnia sp. di Pembenihan ... 11
2.4. Bakteri Kitinolitik sebagai Pengendali hayati Saprolegnia sp... 12
III. METODE PENELITIAN 3.1. Tempat dan Waktu ... 16
3.2. Bahan dan Alat ... 16
3.3. Isolasi dan Identifikasi Saprolegnia sp. ... 17
3.4. Isolasi dan Identifikasi Bakteri Kitinolitik ... 17
3.5. Penampakan Morfologi dan Biokimia Bakteri Kitinolitik ... 18
3.6. Uji Penghambatan Pertumbuhan Saprolegnia sp. ... 18
3.7. Uji Pengukuran N-asetilglukosamin ... 18
3.8. Uji Produksi Glukanase oleh Isolat Bakteri Kitinolitik ... 19
3.9. Uji Patogenitas Saprolegnia sp. ... 19
3.10. Uji Patogenitas dan Perlekatan Mikroba pada Telur ... 19
3.11. Tahap Evaluasi Efek Bakteri Kitinolitik Secara In Vivo ... 20
3.1.1. Rancangan Percobaan ... 21
IV. HASIL DAN PEMBAHASAN
4.1. Hasil Isolasi dan Karakterisasi Bakteri Kitinolitik ... 23
4.2. Hasil Isolasi dan Karakterisasi Saprolegnia sp. ... 25
4.3. Hasil Antagonisme secara In Vitro ... 28
4.4. Hasil Patogenitas Saprolegnia sp. ... 39
4.5. Hasil Patogenitas dan Perlekatan Bakteri Kitinolitik ... 41
4.6. Hasil Antagonisme sevara In Vivo ... 47
V. KESIMPULAN DAN SARAN 5.1. Kesimpulan ... 52
5.2. Saran ... 52
DAFTAR PUSTAKA LAMPIRAN
Halaman
1 Ringkasan penelitian penggunaan probiotik sebagai pengendali
hayati infeksi Saprolegnia sp. di bidang akuakultur ... 14 2 Hasil pengamatan morfologi dan uji biokimia isolat bakteri
kitinolitik ... 24 3 Hasil uji antagonisme in vitro bakteri kitinolitik terhadap
Saprolegnia sp. ... 30 4 Kategori kemampuan daya hambat isolat bakteri kitinolitik
terhadap Saprolegnia sp. ... 32 5 Hasil pengamatan mikroskopis abnormalitas hifa Saprolegnia
sp. yang disebabkan oleh isolat bakteri kitinolitik pada uji
antagonisme ... 33 6 Hasil pengukuran diameter zona bening pada uji awal glukanase 37 7 Hasil isolasi ulang bakteri kitinolitik pada telur gurami pada
media MGMC ...
44
DAFTAR GAMBAR
Halaman
1 Kerangka memikiran penelitian ... 5 2 Kolam budidaya gurami ... 8 3 Infeksi Oomycetes Saprolegnia sp. pada telur gurami di pembenihan
tradisional ... 8 4 Pengamatan preparat basah sampel kulit yang mengalami lesi akibat
infeksi Saprolegnia sp. ... 10 5 Wadah kaca untuk percobaan ... 21 6 Telur gurami yang terinfeksi jamur air Saprolegnia sp. ... 26 7 Preparat basah telur terinfeksi Saprolegnia sp. hifa bercabang,
caenocytic dan tidak bersekat ... 27 8 Struktur Saprolegnia sp. yang ditumbuhkan pada media SDA... 28 9 Zona hambat hari ke-4 beberapa isolat bakteri kitinolitik terhadap
isolat Saprolegnia sp. ... 29 10 Perubahan morfologi hifa Saprolegnia sp. setelah diuji antagonisme
dengan bakteri kitinolitik ... 34 11 Abnormalitas hifa Saprolegnia sp. pada uji antagonis in vitro ... 35 12 Perubahan lapisan bagian luar telur umur 4 hari dan perubahan
epidermal dan dermal telur umur 7 hari yang berhubungan dengan
lesi yang disebabkan oleh Saprolegnia sp. ... 40 13 Gambaran histologi telur dan larva ikan ... 42 14 Gambaran Permukaan sel telur gurami dengan Scanning Electron
Microscope (SEM) ... 45 15 Histogram tingkat kematian telur gurami pada uji tantang isolat
bakteri terhadap Saprolegnia sp. secara in vivo ... 47 16 Histogram tingkat daya tetas telur gurami pada uji tantang isolat
DAFTAR LAMPIRAN
Halaman
1 Alur Kerja Pengambilan Sampel ... 60
2 Komposisi Medium ... 61
3 Alur Kerja Uji Penghambatan Saprolegnia sp. In Vitro ... 62
4 Alur Kerja Pengukuran N-asetilglukosamin ... 63
5 Preparasi Saprolegnia sp. untuk Uji In Vivo ... 64
6 Alur Kerja Patogenitas Saprolegnia sp. ... 65
7 Alur Kerja Preparasi Isolat Bakteri ... 66
8 Alur kerja patogenitas Isolat Bakteri... 67
9 Alur kerja Perlekatan Isolat Bakteri ... 68
10 Cakupan Kegiatan Penelitian ... 69
11 Isolat Biakan Murni Bakteri Kitinolitik dan Beberapa Hasil Uji Biokimia ... 70
12 Hasil Pengukuran Kadar N-asetilglukosamin pada Uji Antagonisme Isolat Bakteri Kitinolitik Terhadap Saprolegnia sp.. 71
ABSTRAK
Penelitian ini bertujuan untuk mengetahui potensi 24 bakteri kitinolitik yang diisolasi dari kolam budidaya ikan gurami (Osphronemus gouramy) untuk menghambat pertumbuhan isolat Saprolegnia sp. yang diisolasi dari telur gurami. Uji antagonisme dilakukan pada media garam minimum dengan 2% koloidal kitin untuk mengetahui kemampuan isolat bakteri menghambat Saprolegnia sp. Sepuluh isolat bakteri menunjukkan daya hambat dengan diameter 10–19 mm. Abnormalitas hifa diamati secara mikroskopis meliputi lisis (40%), lisis pada ujung hifa (30%), hifa bengkok (23,33%) dan hifa terpuntir (6,67%). Uji patogenesa isolat Saprolegnia sp. dilakukan untuk mengetahui kemampuan isolat tersebut menginfeksi telur gurami. Hasil penelitian menunjukkan bahwa isolat Saprolegnia sp. patogen terhadap telur. Uji patogenesa dan perlekatan bakteri potensial dilakukan untuk mengeliminasi isolat yang bersifat patogen dan tidak dapat melakukan perlekatan terhadap mukosa sel telur. Isolat PB01 bersifat patogen bagi telur sehingga tidak dapat digunakan sebagai pengendali hayati Saprolegnia sp. Semua isolat memiliki kemampuan melekat pada sel telur ditunjukkan dengan uji reisolasi dan pengamatan Scanning Electron Microscope (SEM). Uji tantang isolat bakteri potensial menghambat infeksi Saprolegnia sp. pada telur dilakukan pada bak percobaan dan kondisi terkontrol. Hasil penelitian menunjukkan bahwa 6 isolat bakteri yaitu PB05, PB08, PB13, PB14, PB15, dan PB17 mampu menurunkan tingkat mortalitas serta meningkatkan daya tetas telur yang yang diinfeksi Saprolegnia sp. sehingga potensial untuk dijadikan agen pengendali hayati Saprolegnia sp. pada telur gurami.
ABSTRACT
A study on potential antagonist activity of 24 chitinolytic bacteria isolated from Osphronemus gouramy pond culture to inhibit the Saprolegnia sp. causal agent of saprolegniasis in gouramy egg was carried out. The ability of bacterial isolates to inhibit the Saprolegnia sp. in vitro was evaluated by antagonism assay on minimum salt medium agar with 2% colloidal chitin as C source. Hyphal abnormality as a result of antagonistic assay was examined in this study. The result showed that 10 isolates, PB3A, PB01, PB02, PB05, PB08, PB10, PB13, PB14, PB15 dan PB17 showed relatively high inhibition zone. The diameter of inhibition zone ranged from 10-19 mm. The abnormality of miselium was lysis (40%), a lised hyphae tips (30%), bent hyphae (23,33%) and twisted hyphae (6,67%). Saprolegnia sp. was examined for its ability causing infection in egg. The result showed that this isolates caused infection on gouramy egg. Pathogenecity and attachment of bacteria assay was done to eliminate pathogen isolate and to know the ability of isolates to adhere on the egg mucosa. The result showed that PB01 was pathogen and all potential bacteria had ability to adhere to egg surface. It was proved by reisolation assay and examination on Scanning Electron Microscope (SEM). Potential chitinolytic bacteria were assayed to Saprolegnia sp. in vivo using gouramy egg. Potential bacteria (PB05, PB08, PB13, PB14, PB15, and PB17) could reduce mortality rate and increase hatching rate on infected egg. Based on this study, we concluded that PB05, PB08, PB13, PB14, PB15, and PB17 had the ability to inhibit the Saprolegnia sp. these isolates could be utilized as potential biological control candidates against Saprolegniasis.
BAB I PENDAHULUAN
1.1. Latar Belakang
Produksi perikanan dunia mengalami peningkatan hingga 11% selama 10 tahun terakhir (Van West 2006). Data FAO (2010) menyebutkan bahwa produksi perikanan di Indonesia meningkat dari 996.659 ton pada tahun 2003 menjadi 1.045.051 ton pada tahun 2004. Hal ini menunjukkan bahwa perikanan menjadi sektor yang berkembang pesat dalam perekonomian pangan. Kebutuhan protein hewani masyarakat yang semakin meningkat menyebabkan peningkatan produksi di bidang perikanan. Sejumlah lapangan kerja di sektor perikanan memberikan kontribusi dalam penyerapan tenaga kerja di Indonesia. Jumlah tenaga kerja di bidang perikanan meningkat dari 2.384 208 orang pada tahun 2003 menjadi 2.459.355 orang pada tahun 2004 (FAO 2010).
Produksi perikanan dunia tahun 1999 didominasi oleh ikan air tawar (58%), diikuti oleh ikan air laut (36%) dan air payau (6%) (Van West 2006). Di Indonesia, ikan air tawar mendominasi produksi ikan yaitu sebesar 46% (FAO 2010). Gurami (Osphronemus gouramy) merupakan satu diantara beberapa spesies asli Indonesia yang banyak dibudidayakan. Produksi gurami sebesar 7,68% dari total produksi ikan air tawar (FAO 2010). Teknik budidaya gurami merupakan hal penting dalam peningkatan produksinya. Petani dapat meningkatkan produktivitas dengan ekstensifikasi, yaitu memperluas daerah budidaya atau meningkatkan produktivitas dengan intensifikasi lahan budidaya. Penyakit merupakan salah satu kendala dalam budidaya ikan yang menyebabkan penurunan produksi ikan. Infeksi oleh Saprolegnia sp. merupakan satu diantara beberapa permasalahan yang dijumpai baik pada ikan maupun telurnya (Bruno & Wood 1999). Tingkat keberhasilan penetasan telur gurami akan menurun jika terinfeksi oleh Saprolegnia sp.
peradangan yang disebabkan oleh Saprolegnia. Pada lesi terdapat miselium Saprolegnia dengan bentukan seperti kapas yang tumbuh pada permukaan tubuh, insang (Noga 2000) atau telur ikan ketika di dalam air (Khoo 2000). Warna lesi bervariasi yaitu kemerahan, cokelat atau kehijauan. Hal ini disebabkan partikel sedimen bahan organik ataupun alga yang menempel pada miselium Saprolegnia (Khoo 2000). Jumlah telur dalam satu meter kubik pada pembenihan mempengaruhi infeksi Saprolegnia pada telur (Celada et al. 2004). Inkubasi telur ikan dengan kepadatan tinggi menjadi faktor predisposisi terjadinya infeksi Saprolegnia pada telur (Rach et al. 1997). Apabila infeksi Saprolegnia sudah terdapat pada salah satu telur maka segera menyebar ke telur yang sehat. Infeksi Saprolegnia pada telur menyebabkan tingkat hidup (survival rate) telur ikan menjadi menurun. Telur ikan yang mati dan bahan organik pada sistem budidaya merupakan substrat yang sesuai bagi pertumbuhan Saprolegnia (Rach et al. 1998). Infeksi Saprolegnia pada telur merupakan kendala utama pada usaha budidaya perikanan sehingga diperlukan usaha untuk pengendaliannya dengan menggunakan bahan anti Saprolegnia yang efektif dalam pembenihan telur.
Pengendalian infeksi Saprolegnia yang efektif hingga tahun 2002 adalah menggunakan malachite green (Van West 2006). Namun penggunaannya telah dilarang oleh Dewan Farmasi Jepang pada tahun 2003 (Khomvilai et al. 2006), Food and Drug Administration (FDA) pada tahun 1991 (Schereier et al. 1996) dan seluruh dunia (Van West 2006). Hal ini disebabkan karena malachite green bersifat karsinogenik dan toksik (Van West 2006), mutagenik dan teratogenik (Srivastava et al. 2004) bagi makhluk hidup. Nowak & de Guingand (1997) melaporkan bahwa malachite green dapat menyebabkan residu pada jaringan ikan salmon.
korosif dan dosis efektifnya tinggi yaitu 1000 ppm. Penggunaan hidrogen peroksida tidak diperbolehkan di Amerika (Schreier et al. 1996). Natrium klorida relatif aman dalam penggunaannya namun dalam aplikasinya membutuhkan biaya besar karena dosis efektifnya tinggi yaitu 30.000 ppm (Khomvilai et al. 2006).
Sejak tahun 1990, Eropa telah membatasi penggunaan produk yang mempengaruhi lingkungan dalam bidang akuakultur dengan membatasi penggunaan kemoterapi (Mousavi et al. 2009). Sehingga saat ini banyak penelitian yang mencari kandidat bahan anti Saprolegnia yang berasal dari alam atau bahan biologi. Pengendalian infeksi Saprolegnia dengan menggunakan bakteri Aeromonas telah dilaporkan oleh Lategan et al. (2004a). Pengendalian hayati patogen oleh beberapa mikroorganisme dari laut juga telah dilaporkan (El Kassas & Khairy 2009). Pengendalian hayati menjadi alternatif yang dipilih karena lebih ramah lingkungan dan tidak menimbulkan efek toksik (Gohel et al. 2006).
Enzim kitinase yang berasal dari organisme telah banyak dilaporkan memiliki kemampuan sebagai kontrol biologi Saprolegnia. Enzim kitinase telah dilaporkan mampu mengendalikan infeksi patogen pada tanaman (Gohel et al. 2006). Sistem kitinolitik terdiri atas endokitinase, kitobiase dan eksokitinase yang bekerja secara sinergis dalam mendegradasi kitin menjadi N asetilglukosamin bebas. Selain kemampuannya mendegradasi dinding sel Saprolegnia secara langsung, kitinase melepaskan oligo-N-asetil glukosamin yang berfungsi mengaktifkan mekanisme pertahanan tubuh tanaman (Gohel et al. 2006). Kitosan sebagai turunan kitin dilaporkan dapat menghambat infeksi Saprolegnia sp. (Muzareli et al. 2001) dan sebagai antibakterial pada patogen ikan (Li et al. 2008). Berdasarkan kemampuannya dalam mendegradasi kitin sebagai penyusun dinding sel Saprolegnia maka mikroba penghasil enzim kitinase seperti bakteri kitinolitik dapat menjadi kandidat pengendali Saprolegnia.
et al. 2001; Kamil et al. 2007). Bakteri Aeromonas dan Pseudomonas fluorescens dilaporkan memiliki kemampuan sebagai pengendali Saprolegnia sp. (Lategan et al. 2004a; Hatai & Willoughby 1988). Aplikasi bakteri kitinolitik yang diisolasi dari perairan budidaya sebagai pengendali Saprolegnia sp. pada telur gurami belum banyak dilaporkan. Sehingga perlu dilakukan penelitian untuk mengetahui kemampuan isolat bakteri kitinolitik sebagai pengendali hayati Saprolegnia sp. yang menginfeksi telur gurami.
1.2. Pembatasan dan Perumusan Masalah
Penelitian tentang pengendalian hayati Saprolegnia sp. menggunakan bakteri kitinolitik perairan dibatasi aplikasinya pada telur gurami didasarkan pada pertimbangan bahwa infeksi Saprolegnia sp. merupakan permasalahan utama di pembenihan gurami. Saprolegnia di pembenihan pada saat ini dikendalikan dengan menggunakan bahan kimia seperti malachite green dan formalin (Van West 2006).
Pengendalian hayati infeksi Saprolegnia sp. menjadi alternatif dalam meningkatkan tingkat keberhasilan pembenihan ikan gurami. Penggunaan bakteri Aeromonas strain A 199 dan P. fluorescens sebagai pengendali hayati Saprolegnia sp. telah dilaporkan oleh Lategan et al. (2004a) serta Hatai & Willoughby (1989). Bakteri kitinolitik memiliki kemampuan mendegradasi kitin sebagai salah satu komponen utama Saprolegnia.
Berdasarkan uraian di atas dapat dirumuskan beberapa permasalahan, yaitu 1. Apakah bakteri di perairan air tawar di kolam budidaya gurami memiliki
kemampuan kitinolitik ?
2. Apakah bakteri kitinolitik memiliki potensi sebagai pengendali hayati infeksi Saprolegnia sp. pada telur gurami ?
1.3. Kerangka Pemikiran
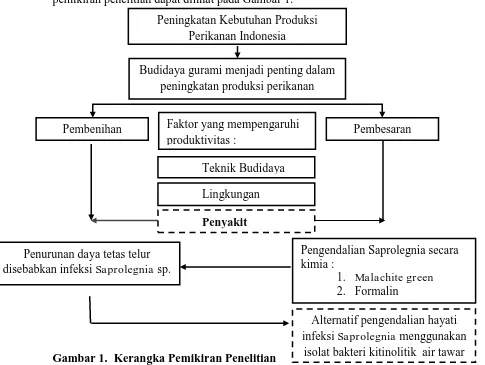
dan manusia. Sehingga diperlukan alternatif pengendalian infeksi Saprolegnia dengan menggunakan bahan alami. Bakteri yang memiliki kemampuan mendegradasi kitin seperti bakteri kitinolitik merupakan satu dari beberapa kandidat yang dapat digunakan untuk mengendalikan infeksi Saprolegnia. Bakteri kitinolitik tersebar di alam, beberapa spesies bakteri tersebut habitatnya di perairan. Kandidat bakteri yang memiliki kemampuan kitinolitik di perairan yang dapat digunakan sebagai pengendali Saprolegnia sp. belum banyak dilaporkan. Berdasarkan hal tersebut, telah dilakukan penelitian tentang kemampuan bakteri kitinolitik yang berasal perairan air tawar untuk mengendalikan infeksi Saprolegnia sp. pada telur gurami. Sehingga diperoleh alternatif pengendalian Saprolegnia di pembenihan gurami secara alami. Kerangka pemikiran penelitian dapat dilihat pada Gambar 1.
Gambar 1. Kerangka Pemikiran Penelitian
= Hubungan antar faktor – faktor yang berperan dalam budidaya = Faktor – faktor yang diamati dalam penelitian
Peningkatan Kebutuhan Produksi Perikanan Indonesia
Budidaya gurami menjadi penting dalam peningkatan produksi perikanan
Pembenihan Faktor yang mempengaruhi Pembesaran produktivitas :
isolat bakteri kitinolitik air tawar Teknik Budidaya
Lingkungan
1.4. Tujuan Penelitian
Penelitian ini bertujuan untuk :
1. Mendapatkan jenis bakteri air tawar yang berpotensi dan yang memiliki kemampuan kitinolitik
2. Mengevaluasi kemampuan bakteri kitinolitik air tawar sebagai pengendali infeksi Saprolegnia air Saprolegnia sp. secara in vitro
3. Mengevaluasi kemampuan bakteri kitinolitik dalam mengendalikan infeksi Saprolegnia air Saprolegnia sp. pada telur gurami secara in vivo
1.5. Manfaat Penelitian
Penelitian ini diharapkan dapat memberikan informasi jenis bakteri air tawar yang memiliki kemampuan kitinolitik dan dapat digunakan sebagai pengendali hayati Saprolegnia sp. pada telur gurami baik secara in vitro maupun secara in vivo. Sehingga dapat memberikan kontribusi bagi pengendalian penyakit dalam budidaya gurami.
1.6. Hipotesis
BAB II
TINJAUAN PUSTAKA
2.1. Budidaya Gurami (Osphronemus gouramy)
Gurami merupakan ikan air tawar yang berasal dari Indonesia (Welcomme 1988). Gurami merupakan spesies ikan yang berukuran besar, memiliki sifat herbivora dan dapat hidup pada kelarutan oksigen rendah di dalam air (Welcomme 1988). Gurami disukai karena dagingnya yang tebal dan rasanya yang enak. Gurami telah dibudidayakan secara luas oleh masyarakat Indonesia dan menyebar ke negara lain. Teknik budidaya gurami menyebar dari Indonesia ke India tahun 1916, Filipina tahun 1927, Srilanka tahun 1939 dan Maritius tahun 1951 (Welcomme 1988).
Ikan gurami tergolong ikan air tawar yang pertumbuhannya lambat. Di perairan alam, gurami hidup di sungai, rawa air tawar yang berada 50–600 meter di atas permukaan laut. Tempat ideal untuk budidaya gurami berada pada ketinggian 50–400 meter di atas permukaan laut dengan suhu optimal bagi pertumbuhan gurami adalah 24–28°C (Murtidjo 2001). Salah satu lokasi di Sumatera Utara yang menjadi pusat budidaya gurami adalah Perbaungan. Perbaungan merupakan kecamatan di Kabupaten Serdang Bedagai. Wilayah Perbaungan yang berada pada ketinggian 0 – 500 meter di atas permukaan laut merupakan daerah ideal untuk budidaya gurami (Pemerintah Kabupaten Serdang Bedagai 2006). Kolam budidaya gurami di daerah Perbaungan ditampilkan pada Gambar 2. Keberhasilan budidaya gurami sangat dipengaruhi oleh kualitas bibit, kondisi cuaca, kondisi kualitas air dan penyakit.
Gambar 2. Kolam budidaya gurami
Penetasan yang dilakukan oleh petani di daerah Perbaungan di lakukan pada bak beton berukuran 1 x 1 x 1 meter dan diberi pelindung agar tidak terkena cahaya matahari langsung. Tingkat keberhasilan penetasan telur sangat dipengaruhi oleh kualitas induk. Telur yang berasal dari induk yang masih muda biasanya tingkat penetasan telurnya rendah. Hal ini disebabkan telur masih muda sehingga sisa kuning telur yang tidak menetas menjadi media yang baik untuk Saprolegnia (Rach et al. 1997). Infeksi Saprolegnia dengan cepat menyebar kepada telur yang sehat dan menyebabkan penurunan tingkat keberhasilan penetasan telur gurami. Bentuk infeksi Oomycetes pada telur gurami di pembenihan tradisional ditampilkan pada Gambar 3.
Gambar 3. Bentuk infeksi Saprolegnia sp. pada telur gurami di pembenihan tradisioan. (A) Telur normal, (B) Telur terinfeksi Saprolegnia sp.
A
2.2. Saprolegnia sp.
Saprolegnia yang termasuk dalam Oomycetes merupakan patogen utama pada ikan air tawar (Noga 2000). Oomycetes dibagi menjadi 4 ordo dan tiga diantaranya dapat menginfeksi ikan yaitu Saprolegniales, Leptomitales dan Peronosporales (Noga 1993). Infeksi Saprolegnia pada ikan paling sering disebabkan oleh Saprolegnia dari famili Saprolegniaceae (Noga 2000). Klasifikasi Saprolegnia didasarkan pada siklus hidup, morfologi hifa dan unit reproduksi seperti tipe spora yang dihasilkannya (Khoo 2000).
Secara konvensional, Oomycetes dimasukkan ke dalam kingdom Saprolegnia berfilamen. Namun analisis secara molekular dan biokimia menyebutkan bahwa Oomycetes secara taksonomi sedikit memiliki kesamaan dengan jamur berfilamen namun dekat kekerabatannya dengan alga cokelat (heterokont) dalam Stramenophiles yaitu salah satu eukaryot (Kamoun 2003). Karakteristik Saprolegnia ini yang membedakannya dengan yang lain adalah Oomycetes menghasilkan heterokont zoospora yaitu spora motil biflagelata (Roberts 2001). Zoospora adalah alat reproduksi aseksual utama yang dihasilkan oleh zoosporangium. Reproduksi seksual dengan peleburan dua gamet membentuk dinding tebal yaitu oospora yang merupakan asal penamaan kelas Oomycetes (Roberts 2001). Secara ultrastruktur, Oomycetes memiliki krista mitokondria dengan bentuk tubular berbeda dengan Saprolegnia lainnya yang memiliki bentuk platelike cristae atau bentuk pipih (Kamoun 2003). Keunikan Oomycetes juga ditemukan pada komposisi dinding selnya. Penyusun utama dinding sel Oomycetes adalah polimer β-1,3 glukan dan selulosa namun dengan sedikit kitin. Walaupun kitin penyusun dinding sel yang minor namun merupakan komponen penting dalam dinding sel disebabkan kitin sintase menghambat polyoxin D yang menyebabkan terhambatnya pertumbuhan dinding sel Oomycetes (Kamoun 2003).
sampel kulit ikan yang mengalami lesi akibat Saprolegnia sp. dapat dilihat pada Gambar 4.
Gambar 4. Pengamatan preparat basah sampel kulit yang mengalami lesi akibat infeksi Saprolegnia sp. (Khoo 2000)
Oomycetes secara alami tersebar di perairan air tawar dan mewakili kelompok patogen yang menginfeksi ikan dan telur (Mastan 2008). Saprolegnia tidak dapat mensintesis nutrisi karena bersifat heterotrof yaitu membutuhkan bahan organik untuk pertumbuhan dan perkembangannya. Saprolegnia dikategorikan sebagai saprofit yang menggunakan bahan organik ataupun sebagai parasit yang menginfeksi mahluk hidup agar dapat bertahan hidup (Khoo 2000).
metode ini memerlukan ketelitian dan dapat menyebabkan kerusakan pada telur sehat (Carral 2004). Pada tahap ini diperlukan bahan yang bersifat fungistatik untuk menghambat pertumbuhan Saprolegnia dari telur yang mati yang terinfeksi dan menghambat penyebaran Saprolegnia.
2.3 Pengendalian Oomycetes di Pembenihan
bahwa bakteri Aeromonas strain A 199 memiliki kemampuan menurunkan kejadian saprolegniosis pada ikan.
2.4. Bakteri Kitinolitik sebagai Pengendali Hayati Saprolegnia sp.
Kitin adalah adalah homopolimer β 1-4 N-asetilglukosamin yang tersebar luas di alam ini dan ditemukan pada kutikula serangga, kerapas krustasea dan dinding sel Saprolegnia (Watanabe et al. 1999; Gohel et al. 2006). Enzim yang dapat melakukan degradasi kitin adalah kitinase atau enzim kitinolitik. Kitinase terdiri atas famili glycosyl hydrolase 18 dan 19 berdasarkan klasifikasi dari Henrissat & Bairoch (1993), serta 20 (Chernin & Chet 2002). Klasifikasi ini didasarkan pada persamaan sekuen asam amino dari domain katalitik enzim tersebut (Henrissat dan bairoch 1993). Walaupun dalam famili yang sama, kitinase menunjukkan perbedaan dalam hal spesifikasi substrat, reaksi dan pH optimum (Chernin & Chet 2002). Sifat hidrolisis dari kitinase menjadi dasar untuk dapat digunakan sebagai pengendali hayati Saprolegnia dengan memanfaatkan organisme yang menghasilkan enzim ini.
Organisme yang dapat mendegradasi kitin tersebar luas di alam, termasuk organisme yang tidak memiliki kitin seperti bakteri, virus, tumbuhan tingkat tinggi dan hewan yang memiliki peran penting dalam fisiologi dan ekologi. Mikroba mendegradasi kitin dengan mensekresikan enzim yang memiliki spesifitas tertentu untuk mengubah atau menghidrolisis kitin (Matsumoto 2006). Kitinase yang diproduksi oleh beberapa mikroba memiliki perbedaan dalam hal berat molekul, suhu optimum dan tingkat stabilitasnya serta memiliki rentang pH yang luas (Chernin & Chat 2002).
air laut (Han et al. 2009) dan danau yang memiliki hipersalin dan alkalin (Lecleir & Hollibaugh 2006).
Keberadaan bakteri kitinolitik di lingkungan akuatik berperan penting sebagai pendegradasi sisa kitin dari eksosekeleton karapas krustasea, diatom, protozoa, nematoda (Brzezinska & Donderski 2006) yang telah mati sehingga berperan dalam siklus C dan N dalam ekosistem perairan (Chernin & Chat 2002). Bakteri ini dapat mengubah kitin menjadi bahan organik sehingga dapat digunakan sebagai sumber nitrogen dan karbon (Han et al. 2009). Bakteri kitinolitik menyebabkan shell diseases syndrome yaitu kondisi degradasi pada eksoskeleton krustasea sehingga terbentuk lesi spot hitam (Vogan dan Rowley 2002).
Bakteri kitinolitik merupakan kandidat bakteri yang dapat digunakan dalam pengendali hayati jamur. Gohel (2006) menyebutkan beberapa bakteri yang digunakan sebagai biokontrol jamur patogen pada tanaman antara lain Pseudomonas syringae sebagai biokontrol patogen Botrytis cinerea, Penicillium spp., Mucor pyroformis dan Geotrichum candidum. Jamur patogen pada tanaman yaitu Fusarium semitectum, Ganoderma boninense and Penicillium citrinum dapat dihambat oleh isolat bakteri kitinolitik (Suryanto & Munir 2006).
Tabel 1. Ringkasan penelitian penggunaan probiotik sebagai pengendali hayati infeksi Saprolegnia sp. dibidang akuakultur
Hewan Percobaan
Probiotik
potensial Patogen Metode uji Referensi
Aeromonas
Saprolegnia sp. In vivo Lategan et al. (2004a)
BAB III
METODE PENELITIAN
3.1. Waktu dan Tempat
Penelitian ini dilaksanakan dari bulan Juli 2010 hingga Juli 2011. Pengambilan sampel dilakukan di bak pembenihan dan kolam budidaya gurami yang berlokasi di Kecamatan Perbaungan, Kabupaten Serdang Bedagai, Propinsi Sumatera Utara. Penelitian dilaksanakan di Laboratorium Mikrobiologi Departemen Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam (MIPA), Universitas Sumatera Utara, Laboratorium Ilmu Dasar, Universitas Sumatera Utara, Laboratorium Patologi Anatomi dan Laboratorium Biokimia Fakultas Kedokteran, Universitas Sumatera Utara, Laboratorium Kultur Jaringan Fakultas Pertanian, Universitas Sumatera Utara, serta Lembaga Ilmu Pengetahuan Indonesia (LIPI) Cibinong, Jawa Barat.
3.2. Bahan dan Alat
3.3. Isolasi dan Identifikasi Saprolegnia sp.
Telur gurami yang terinfeksi Saprolegnia diambil dan disimpan pada botol steril dengan suhu penyimpanan 10-14°C (Mousavi et al 2009). Teknik pengambilan sampel Saprolegnia disajikan pada Lampiran 1. Telur diletakkan pada cawan Petri steril. Hifa Saprolegnia diambil kemudian diamati morfologi Saprolegnia yang tumbuh di telur di bawah mikroskop cahaya. Untuk mendapatkan koloni murni Saprolegnia ditanam pada media SDA (Bruno & Wood 1999) yang telah diberi antibiotika klorampenikol. Kemudian diinkubasi pada suhu 30°C selama 48 jam. Identifikasi morfologi berdasarkan zoosporangium dan organ seksual. Isolat yang tumbuh diidentifikasi menggunakan metoda Hughes (1994), Beakes et al. (1994) dan Rajan (2000).
3.4. Isolasi dan Identifikasi Bakteri Kitinolitik
Sampel air diambil dari kolam ikan yang tidak pernah terinfeksi Saprolegnia atau sampel air dari ikan yang sehat. Sampel air diambil pada bagian permukaan kolam yaitu 20–30 cm dari permukaan (Brzezinska & Donderski 2006) untuk mendapatkan sampel air yang mewakili. Sampel air diambil dengan menggunakan botol steril kemudian disimpan dalam kotak pendingin yang disesuaikan suhunya yaitu kurang dari 7°C (Brzezinska & Donderski 2006). Pengambilan sampel air disajikan pada Lampiran 1.
3.5. Penampakan Morfologi dan Biokimia Bakteri Kitinolitik
Biakan murni bakteri kitinolitik dikarakterisasi sifat morfologi dan biokimianya. Pengamatan ciri-ciri morfologi koloni bakteri antara lain bentuk dan warna koloni, motilitas, bentuk sel dan sifat gram. (Cappuccino & Sherman 1996). Uji motilitas menggunakan media semi padat Sulfide indol motility (SIM). Sifat biokimia yang diuji meliputi penggunaan sitrat dengan media Simmons Citrate Agar (SCA), uji amilase dengan media Starch Agar (SA), uji karbohidrat menggunakan media Triple Sugar Indol Agar (TSIA). Uji katalase menggunakan larutan 3% H2O2 (Cappuccino & Sherman 1996; Allen et al. 1983).
3.6. Uji Penghambatan Pertumbuhan Saprolegnia sp. secara In Vitro
Kemampuan bakteri kitinolitik menghambat pertumbuhan Oomycetes diuji dengan asai antagonisme in vitro. Biakan Saprolegnia sp. diinokulasikan pada agar MGMC dengan jarak 3,5 cm dari kertas cakram tempat inokulan bakteri. Bakteri kitinolitik sebanyak 10 µl (setara dengan 108 sel/ml) diinokulasikan pada kertas cakram. Biakan diinkubasi pada suhu 30°C. Zona hambat terhadap miselium yang tumbuh diamati mulai hari kedua hingga hari ketujuh. Diameter zona hambat dihitung dengan mengukur selisih radial pertumbuhan Saprolegnia normal dengan radial pertumbuhan Saprolegnia yang terhambat oleh isolat bakteri. Alur kerja uji antagonisme in vitro dapat dilihat pada Lampiran 3.
3.7. Uji Pengukuran Kadar N-asetilglukosamin
ditambahkan hifa kering tanpa bakteri kitinolitik. Inkubasi pada shaker water bath 150 rpm selama 24 jam. Sampel dihitung gula reduksinya yaitu kadar GlcNAc dengan metode Jeaniaux 1966 dalam Irawati 2008. Metode pengukuran kadar GlcNAc pada uji antagonisme ini disajikan pada Lampiran 4.
3.8. Uji Awal Produksi Glukanase oleh Isolat Bakteri Kitinolitik
Isolat Candida albicans (isolat laboratorium mikrobiologi FMIPA USU) dengan konsentrasi 108 disebar pada media Muller Hinton Agar (MHA) dengan menggunakan cotton swab. Sebanyak 10µl (setara dengan 108) bakteri kitinolitik yang telah diinokulasikan pada kertas cakram diletakkan pada keempat sisi cawan Petri yang telah diinokulasikan C. albicans. Biakan diinkubasi pada suhu 30°C selama 24 jam. Pengamatan yang dilakukan adalah mengukur zona hambat yang terbentuk yaitu zona bening di sekitar koloni bakteri.
3.9. Uji Patogenitas Saprolegnia sp.
Isolat Saprolegnia sp. diuji patogenitasnya terhadap sel telur gurami untuk mengetahui patogenitasnya terhadap sel telur. Isolat Saprolegnia yang digunakan adalah pada tahap zoospora sehingga perlu untuk melakukan preparasi Saprolegnia sp. Alur kerja preparasi Saprolegnia sp. tersaji pada Lampiran 5. Telur gurami sehat sebanyak 25 buah ditempatkan pada wadah kaca percobaan yang memiliki volume 400 ml air steril. Aplikasi perlakuan dengan pemberian isolat zoospora Saprolegnia sp. sebanyak 0,4 ml (104 sel/ml) ke dalam wadah kaca selama masa perkembangan telur hingga menjadi larva. Kontrol tidak diberikan isolat zoospora Saprolegnia sp. Pengamatan yang dilakukan adalah menghitung tingkat penetasan, tingkat kematian dan tingkat infeksi. Alur kerja uji patogenitas tersaji pada Lampiran 6.
3.10. Uji Patogenitas dan Perlekatan Mikroba pada Telur
gurami sehat sebanyak 25 buah ditempatkan pada wadah kaca percobaan yang memiliki volume 400 ml air steril. Aplikasi perlakuan dengan memberikan isolat bakteri kitinolitik sebanyak 0,4 ml (105 sel/ml) ke dalam wadah kaca selama masa perkembangan telur hingga menjadi larva. Kontrol tidak diberikan bakteri. Pengamatan yang dilakukan adalah menghitung tingkat penetasan dan tingkat kematian (mortality rate).
Uji kemampuan bakteri berikatan dengan jaringan sel telur dilakukan dengan memberikan isolat bakteri kitinolitik pada telur gurami. Telur gurami sehat sebanyak 25 buah ditempatkan pada tempat kaca yang memiliki volume 400 ml air steril. Kontrol tidak diberikan bakteri kitinolitik. Perlakuan yaitu dengan memberikan isolat bakteri kitinolitik sebesar 0,4 ml (105 sel/ml) ke dalam wadah kaca. Pengambilan sel telur dilakukan setelah 48 jam. Sel telur yang diambil dibilas dengan menggunakan akuadest steril tiga kali kemudian digerus dan dikultur dalam media MGMC. Bakteri yang tumbuh diamati pembentukan zona bening apakah merupakan bakteri kitinolitik yang ditambahkan. Telur yang telah diinokulasikan bakteri selama 48 jam dan tahap larva diambil dan difiksasi menggunakan formaline buffer saline untuk uji histologi setelah untuk mengetahui perubahan struktur sel dan respons sel terhadap bakteri. Alur kerja uji patogenitas dan perlekatan tersaji pada Lampiran 8.
3.11. Tahap Evaluasi Efek Bakteri Kitinolitik Secara in Vivo
Pada tahap ini dilakukan evaluasi pengaruh bakteri kitinolitik terhadap infeksi Saprolegnia sp. pada telur. Parameter yang akan diamati adalah jumlah telur yang terinfeksi (infection rate), tingkat penetasan (hatching rate) dan tingkat kematian (mortality rate) telur gurami.
Telur gurami umur 3 hari berasal dari pembenihan tradisional di Yogyakarta. Telur gurami merupakan hasil fertilisasi alami induk betina dewasa dan jantan dewasa. Hanya telur yang sehat yang akan ditetaskan. Telur akan menetas sempurna menjadi larva rata-rata pada hari ke 10 hingga 11.
(dengan diameter 5,5 mm) dan ditempatkan di cawan petri yang terdapat 20 ml Glucose yeast extract agar (GY agar) kemudian diinkubasi 25°C selama 24 -28 jam. Miselium yang dipotong dibilas dengan akuadest steril selama 3 kali kemudian dipindahkan ke dalam 20 ml akuades steril dan diinkubasi pada suhu 25°C selama 24 jam untuk produksi zoospora. Zoospora diamati di bawah mikroskop kemudian jumlah zoospore dihitung dengan menggunakan haemocytometer. Jumlah zoospora yang diinfeksi yaitu 1 x 104 zoospora (Hanjavanit et al. 2008).
Isolat bakteri kitinolitik ditumbuhkan pada media MGMC pada suhu ruang selama 24 jam. Kemudian sebanyak 0,4 ml kultur bakteri kitinolitik yang setara dengan 105 sel/ml ditambahkan kedalam 400 ml air di wadah kaca (Lategan et al 2004a). Penelitian pendahuluan dilakukan untuk mengetahui konsentrasi bakteri ini pada air di wadah percobaan dan kemampuan bakteri ini dapat hidup setelah 24 jam dengan metode di atas. Gambar wadah kaca percobaan disajikan pada Gambar 5
.
Gambar 5. Wadah kaca untuk percobaan
3.11.1.Rancangan Percobaan
zoospora Saprolegnia sp. Kelompok perlakuan yaitu terdiri atas pemberian kandidat bakteri kitinolitik dengan dosis pemberian bakteri yaitu 105 sel/ml dan setelah 24 jam dilakukan uji tantang dengan pemberian isolat zoospora Saprolegnia sp. Setiap perlakuan dilakukan ulangan 3 kali. Tingkat kematian dan tingkat penetasan telur ikan diamati setelah diberikan zoospora. Alur kerja uji tantang secara in vivo dan cakupan kegiatan penelitian disajikan pada Lampiran 9 dan Lampiran 10.
Persentase infeksi, tingkat penetasan dan tingkat kematian telur dihitung menggunakan rumus (Hanjavanit et al 2008):
3.12. Analisis data
BAB 1V
HASIL DAN PEMBAHASAN
4.1. Isolasi dan Karakterisasi Bakteri Kitinolitik
Bakteri kitinolitik pada kolam air tawar pembesaran gurami di Perbaungan
Sumatera Utara telah diisolasi pada penelitian ini. Sebanyak 24 isolat bakteri menunjukkan kemampuan kitinolitik yaitu dengan membentuk zona bening pada daerah sekitar koloni. Penelitian sebelumnya telah melaporkan keberadaan bakteri kitinolitik di danau air tawar. Bakteri kitinolitik telah diisolasi pada dua danau yang berbeda yaitu danau eutrophic dan mesotrophic (Brzezinska & Donderski 2006). Donderski & Brzezinska (2001) melaporkan bahwa beberapa bakteri kitinolitik telah diisolasi dari danau air tawar dan genus Aeromonas sp. mendominasi bakteri kitinolitik tersebut. Isolat bakteri kitinolitik pada media kitin disajikan pada Lampiran 11.
Isolat bakteri kitinolitik memiliki kemampuan dalam menghidrolisis kitin pada media sehingga daerah yang berada di sekitar koloni isolat bakteri menjadi zona bening. Enzim yang menghidrolisis kitin adalah enzim kitinase yang dihasilkan oleh bakteri apabila ada yang menginduksinya. Glukosamin yang merupakan residu dari kitin yang telah deasitilasi dapat menginduksi kitinase (Sahai & Monacha 1993). Kitin yang dipreparasi dari hidrolisis parsial HCl 10 N menghasilkan koloidal kitin mampu menginduksi kitinase kompleks seperti N-asetilglukosamidase, endokhitinase dan khitobiosidase pada Aeromonas caviae (Inbar & Chet 1991). Media MGMC yang digunakan pada penelitian ini terdiri atas koloidal kitin yang dapat menginduksi
kitinase isolat bakteri untuk menghidrolisis kitin pada media.
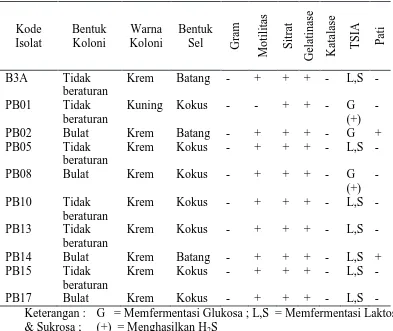
Pengamatan morfologi dan uji biokimia sederhana dilakukan pada 10 isolat
sebanyak 4 isolat dan tidak beraturan sebanyak 6 isolat. Warna koloni didominasi warna krem kecuali isolat PB01 berwarna kuning. Bentuk sel batang terdapat pada 3 isolat dan 7 isolat lainnya berbentuk kokus. Pewarnaan gram menunjukkan semua isolat merupakan bakteri gram negatif. Pewarnaan gram bertujuan untuk menentukan kelompok bakteri gram positif atau negatif. Bakteri gram negatif berwarna merah karena kompleks zat warna kristal violet yodium dilarutkan oleh larutan pemucat sehingga mengambil warna merah saat diteteskan safranin (Lay 1994). Morfologi dan hasil uji biokimia isolat bakteri potensial disajikan pada Tabel 2.
Tabel 2. Hasil pengamatan morfologi dan uji biokimia isolat bakteri kitinolitik
Kode
Keterangan : G = Memfermentasi Glukosa ; L,S = Memfermentasi Laktosa & Sukrosa ; (+) = Menghasilkan H2S
satu-satunya sumber karbon dan energi. Hal ini ditunjukkan dengan perubahan warna hijau menjadi biru pada media Simmon’s citrate agar. Bila mikroba mampu menggunakan sitrat maka asam akan dihilangkan dari media biakan sehingga meningkatkan pH dan mengubah warna media dari hijau menjadi biru (Lay 2004). Uji hidrolisis gelatin merupakan salah satu cara untuk pencirian suatu mikroorganisme. Kemampuan menghidrolisis gelatin dihubungkan juga dengan sifat patogen suatu mikroba. Semua isolat menunjukkan kemampuan menghidrolisis gelatin yaitu ditunjukkan dengan mencairnya gelatin setelah diinkubasi selama 30 menit dalam lemari es (Lay 1994). Uji katalase untuk mengetahui bakteri yang memiliki enzim katalase yang dapat menguraikan hidrogen peroksida menjadi air dan oksigen. Uji TSIA untuk melihat kemampuan mikroba menguraikan asam amino yang mengandung sulfur. Isolat PB01 dan PB08 menghasilkan senyawa FeS pada media TSIA.
4.2. Isolasi dan Karakterisasi Saprolegnia sp.
Isolat Saprolegnia sp. diperoleh dari telur gurami berusia 48 jam yang terinfeksi Saprolegnia. Saprolegnia telah lama diketahui bersimbiosis dengan ikan baik sebagai saprofit maupun parasit. Infeksi Saprolegnia pada telur ditandai dengan adanya bentukan seperti kapas pada telur dengan warna telur yang sudah kuning pucat. Telur ini ditanam pada media SDA dan diinkubasi pada suhu 30 °C selama 48
Gambar 6. Telur gurami yang terinfeksi Saprolegnia sp. dan koloninya pada media GYA. (A) Telur terinfeksi Saprolegniasis (B) koloni Saprolegnia sp. pada media GYA
Identifikasi spesies dari Saprolegnia sp. berdasarkan pada morfologi
Saprolegnia baik bentuk vegetatif maupun alat reproduksi seksual maupun aseksual (Hughes 1994; Beakes et al. 1994). Identifikasi spesies pada Saprolegnia adalah hal yang sulit (Hughes 1994) dan hanya dapat dibedakan berdasarkan analisis taksonomi struktur seksual Saprolegnia dikombinasi dengan identifikasi morfologi Saprolegnia yang terbatas. Isolat Saprolegnia yang diambil dari lesi pada ikan atau telur apabila di
kultur tidak menghasilkan struktur seksual sehingga tidak dapat diidentifikasi spesiesnya (Hughes 1994). Metode menggunakan analisis DNA Saprolegnia merupakan salah satu cara yang dapat dilakukan untuk mengidentifikasi isolat Saprolegnia sp. hingga spesies.
Hasil pengamatan terhadap isolat Saprolegnia yang diperoleh dari isolasi pada telur gurami adalah bentuk vegetatif dan reproduksi aseksual. Morfologi Saprolegnia dengan preparat basah dari telur yang terinfeksi adalah terdapat hifa Saprolegnia yang bercabang, caenocytic, tidak bersekat dan pada ujung hifa terdapat zoospongarium yang berwarna lebih gelap dan lebih bergranular yang akan menghasilkan zoospora. Hal serupa mengenai morfologi Saprolegnia sp. pada pengamatan preparat basah juga telah digambarkan oleh Khoo (2000) & Rajan (2000). Pengamatan preparat basah telur terinfeksi Saprolegnia sp. disajikan pada Gambar 7.
Gambar 7. Preparat basah telur terinfeksi Saprolegnia sp. Hifa Saprolegnia bercabang, cenocytic dan tidak bersekat. (A) Bagian ujung hifa terdapat zoospongarium yang terlihat lebih gelap (panah) (Perbesaran 100x), (B) zoospongarium dengan zoosporangi matang pada bagian dalam kantung yang akan dilepaskan menjadi zoospora (Perbesaran 400x)
Alat reproduksi aseksual Saprolegnia sp. yaitu sel uniselular dan memiliki flagela yang disebut zoospora yang diproduksi pada zoospongarium yang terdapat pada ujung hifa. (Beakes 1996; Rajan 2000). Zoospora dilepaskan dari zoosporangiun menjadi zoospora primer kemudian pecah menjadi zoospora sekunder yang akan berenang bebas. Zoospora sekunder akan melalui beberapa siklus sebelum akhirnya germinasi menjadi hifa. Zoospora yang diamati di bawah mikroskop motil dengan ukuran bulat pipih.
Alat reproduksi seksual tidak ditemukan pada isolat Saprolegnia dari telur gurami. Isolat Saprolegnia tanpa alat reproduksi seksual pertama kali ditemukan oleh Coker pada tahun 1923 dan diberi nama Saprolegnia parasitica (Hughes 1994). Para ahli sekarang menyimpulkan bahwa setiap isolat tanpa alat reproduksi seksual dinamakan Saprolegnia sp. (Hughes 1994). Sebagian besar jenis Saprolegnia yang tumbuh pada jaringan ikan sebagai parasit atau perthrotoph tidak menghasilkan alat reproduksi seksual (Hughes 1994). Saprolegnia yang diambil dari lesi jaringan ikan
biasanya tidak dapat ditemukan alat reproduksi seksualnya (Noga 1993).
Gemmae atau klamidospora ditemukan juga pada isolat sampel Saprolegnia yang ditumbuhkan pada media SDA. Klamidospora merupakan aplanospora aseksual
A A
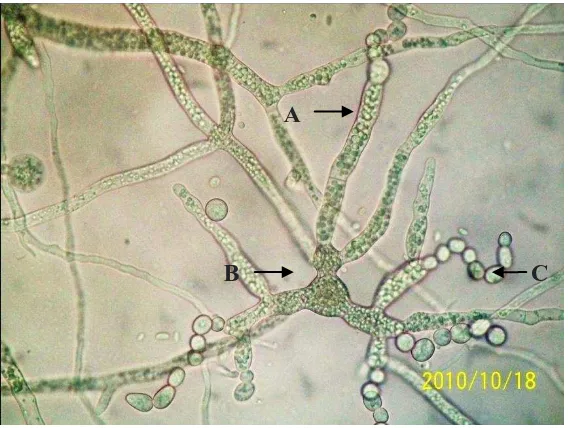
yang terbentuk dari modifikasi hifa yang dapat diproduksi oleh Saprolegnia. Klamidospora dihasilkan oleh Saprolegnia yang diisolasi dari ikan dan sebagian famili yang lain karena kondisi ketersediaan oksigen berkurang pada masa pertumbuhannya (Hughes 1994). Klamidospora berbentuk tidak teratur dan terkadang membentuk rantai seperti ditunjukkan pada Gambar 8. Klamidospora kemudian akan bertunas menghasilkan hifa atau hifa yang disertai zoospongarium pada ujung hifa.
Gambar 8. Struktur Saprolegnia sp. yang ditumbuhkan pada media SDA. (A) Hifa dengan banyak inti sel, globule minyak dan glikogen, (B) gemmae atau klamidospora dengan bentuk iregular keluar tunas terbentuk hifa baru dan (C) katenulasi klamidospora seperti rantai
4.3. Hasil Uji Antagonisme In Vitro
Bakteri kitinolitik yang telah diisolasi kemudian diuji antagonisme dengan isolat Saprolegnia sp. Hal ini dilakukan untuk mengetahui kemampuan isolat bakteri ini dalam menghambat pertumbuhan Saprolegnia sp. Bakteri yang diuji adalah
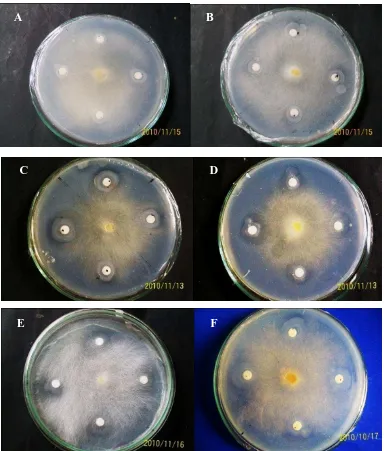
sebanyak 24 isolat yang memiliki kemampuan menghidrolisis kitin pada media. Pengamatan yang dilakukan adalah pengukuran zona hambat pertumbuhan Saprolegnia oleh bakteri selama 7 hari masa inkubasi. Bentuk zona hambat tersebut adalah berupa cerukan penipisan elevasi seperti terlihat pada Gambar 9
C B
Gambar 9. Zona hambat hari ke-4 beberapa isolat bakteri kitinolitik terhadap isolat Saprolegnia sp. Isolat PB04 menunjukkan tidak memiliki daya hambat terhadap isolat Saprolegnia sp. (A). Isolat PB06 menunjukkan daya hambat yang rendah terhadap isolat Saprolegnia sp. (B). Isolat PB13 dan PB17 memiliki daya hambat yang baik terhadap isolat Saprolegnia sp. (C;D). Isolat PB10 dan PB08 memiliki kemampuan yang baik dalam menghambat Saprolegnia sp. (E; F)
Hasil uji antagonisme 24 bakteri air terhadap Saprolegnia sp. menunjukkan bahwa bakteri yang memiliki kemampuan menghambat Saprolegnia dengan baik
C
A B
D
adalah 10 bakteri yaitu isolat bakteri dengan kode PB3A, PB01, PB02, PB05, PB08, PB10, PB13, PB14, PB15 dan PB17. Diameter daya hambat isolat bakteri tersaji pada Tabel 3.
Tabel 3. Hasil uji antagonisme in vitro bakteri kitinolitik terhadap isolat Saprolegnia sp.
Kode Bakteri Zona Hambat Isolat Saprolegnia sp. Hari ke (mm)
4 5 6 7
Isolat bakteri kode PB17 memiliki daya hambat terhadap pertumbuhan Saprolegnia sp yang paling tinggi yaitu dengan diameter zona hambat sebesar 18,46 mm. Efektivitas paling rendah dalam menghambat Saprolegnia sp ditunjukkan oleh isolat bakteri kode PB06 dengan diameter zona hambat sebesar 0,15 mm.
Saprolegnia paling tinggi. Prapagdee et al. melaporkan bahwa filtrat kultur isolat bakteri pada fase stasioner menunjukkan kemampuan daya hambat yang lebih besar terhadap C. gloeosporioides dibandingkan kultur isolat bakteri pada fase eksponensial. Pada hari kelima hingga tujuh tidak menunjukkan peningkatan bahkan ada yang mengalami penurunan diameter zona hambat. Pada beberapa bakteri yang tidak potensial maka Saprolegnia akan terus tumbuh sehingga pada hari kelima rata-rata koloni Saprolegnia menutupi cakram isolat bakteri. Hal ini disebabkan interaksi antara kemampuan bakteri kitinolitik dalam menghasilkan enzim hidrolitik, umur biakan bakteri, jumlah enzim yang dihasilkan, komposisi medium dan waktu inkubasi. Penurunan zona hambat pada setelah hari keenam menunjukkan bahwa kemampuan bakteri semakin menurun karena isolat bakteri sudah masuk fase
kematian karena sumber nitrisi pada media terbatas, kitin sebagai inducer kitinase
dalam media berkurang sehingga sekresi kitinase berkurang dan perkembangan
Saprolegnia sp. yang tumbuh vertikal sehingga dapat melewati daerah zona hambat.
Kemampuan bakteri dalam menghambat Saprolegnia dibagi menjadi 4 kategori yaitu daya hambat kuat yaitu dengan diameter > 20 mm, daya hambat sedang
Tabel 4. Kategori pengelompokan kemampuan daya hambat isolat bakteri kitinolitik
Beberapa penelitian sebelumnya telah melakukan uji antagonisme secara in
vitro bakteri sebagai biologi kontrol Saprolegnia sp. Hatai dan Willoughby (1988) melaporkan bahwa bakteri yang diambil dari lesi ikan salmon yaitu Pseudomonas
fluorescens memiliki kemampuan dalam menghambat pertumbuhan Saprolegnia S. parasitica secara in vitro. Bakteri lain yang ditemukan yaitu Aeromonas dan Enterobacteriaceae tidak menunjukkan daya hambat terhadap pertumbuhan S. parasitica yang ditunjukkan hifa Saprolegnia yang menutupi koloni bakteri tersebut. Bakteri yang termasuk dalam strain Aeromonas tidak patogen (Non Pathogenic Aeromonas Strain (NPAS)) telah diketahui memiliki kemampuan menghambat pertumbuhan Saprolegnia spp. secara in vitro (Osman et al. 2008). Beberapa derivat kitosan seperti MP, NCM dan NPHM telah dilaporkan memberikan penghambatan pertumbuhan terhadap Saprolegnia Saprolegnia parasitica secara in vitro. Hal ini ditunjukkan dengan hasil pengukuran pertumbuhan S. parasitica pada media YM dengan penambahan kitosan NCM dan NPHM yaitu pertumbuhan S. parasitica terhambat dibandingkan dengan kontrol bahkan pada MP, S parasitica tidak menunjukkan pertumbuhan (Muzarelli et al. 2001).
abnormalitas. Pengamatan ini dilakukan untuk mengetahui pengaruh bakteri terhadap struktur morfologi Saprolegnia sp. Abnormalitas pada hifa Saprolegnia adalah perubahan morfologi yang terjadi pada hifa yang seharusnya tumbuh normal. Perubahan morfologi yang terjadi merupakan penyebab terhambatnya pertumbuhan Saprolegnia. Beberapa perubahan morfologi Saprolegnia sp. disajikan pada Tabel 5. Tabel 5. Hasil pengamatan mikroskopis abnormalitas hifa Saprolegnia sp. yang
disebabkan oleh isolat bakteri kitinolitik pada uji antagonisme
Kode
a
Gambar 10. Perubahan morfologi hifa Saprolegnia sp. setelah diuji antagonisme dengan bakteri kitinolitik. Hifa normal (A) Lisis pada ujung hifa (B ; C) Hifa Bengkok (D) Lisis pada dinding sel hifa (E ; F) lisis ujung hifa (G1) dan Hifa terputus disertai lisis (G2 : H) (Perbesaran 400x).
E
A B
C D
E F
Nekrosis pada hifa merupakan abnormalitas yang paling banyak dijumpai yaitu 40% kemudian nekrosis ujung hifa yaitu 30%, hifa bengkok 23,33% dan persentase paling rendah ditemukan pada abnormalitas hifa terputus yaitu 6,67%. Persentase masing-masing abnormalitas dapat dilihat pada Gambar 11 di bawah ini
Gambar 11. Abnormalitas hifa Saprolegnia sp. pada uji antagonis in vitro
Mekanisme penghambatan Saprolegnia sp. oleh isolat bakteri kitinolitik dipelajari pada penelitian ini. Enzim kitinase yang disekresikan oleh isolat bakteri kitinolitik pada uji antagonis terhadap Saprolegnia sp. kemungkinan menjadi salah satu mekanisme dalam penghambatan terhadap Saprolegnia sp. namun bukan merupakan mekanisme utama penghambatan tersebut. Hasil uji kadar GlcNAc bebas menunjukkan hanya pada isolat PB17 terdapat kadar GlcNAc bebas dengan kadar yang rendah yaitu 0,008 µg/ml sedangkan kedua bakteri potensial lainnya PB08 dan PB15 tidak ditemukan. N asetilglukosamin merupakan gula yang terlepas apabila enzim kitinase mendegradasi kitin yang terdapat pada dinding sel Saprolegnia sp. Hasil pengukuran kadar N asetilglukosamin hasil degradasi dinding hifa Saprolegnia sp oleh isolat bakteri kitinolitik potensial disajikan pada Lampiran 12.
Saprolegnia sp. merupakan kelompok Oomycetes dengan komposisi polisakarida pada dinding sel didominasi oleh β-(1-3) dan β-(1-6)-glukan dan selulosa. Kitin pada dinding sel Oomycetes merupakan komponen minor dan tidak lebih dari 4 % dari total polisakarida pada dinding sel (Compos-Takaki et al. 1982). Namun demikian kitin merupakan komponen yang penting pada dinding sel hifa
oomycetes. Banyak penelitian melaporkan bahwa gen kitin sintase terdapat pada beberapa spesies Oomycetes. Mort-Bontemps et al. (1997) melaporkan bahwa biosintesis kitin pada Saprolegnia monaica diperantarai oleh kitin sintase yang dikode oleh gen yang sama dengan Saprolegnia yang berkitin sehingga kitin bukan menjadi perbedaan antara Oomycetes dan jamur berkitin.
Mekanisme penghambatan Saprolegnia oleh glukanase yang dikeluarkan oleh isolat bakteri kitinolitik dipelajari pada penelitian ini. Hasil uji menunjukkan bahwa delapan isolat menunjukkan kemampuan menghasilkan glukanase ditandai dengan adanya zona bening di sekitar koloni bakteri yaitu PB3A, PB01, PB02, PB05, PB13, PB14, PB15 dan PB17 dalam uji antagonisme dengan C. albicans. Isolat PB17 menunjukkan diameter zona bening paling besar yaitu 6,75 mm dan yang paling rendah adalah PB3A yaitu 1,5 mm. Dinding sel C. albicans didominasi oleh karbohidrat glukan yaitu 47-60% glukan (Chaffin et al. 1998). Enzim glukanase yang dikeluarkan isolat bakteri diduga menyebabkan terhambatnya pertumbuhan C. albicans pada daerah sekitar koloni bakteri. Hal ini menyebabkan terbentuknya zona bening disekitar bakteri yang menunjukkan zona hambat terhadap pertumbuhan C. albicans oleh bakteri. Hasil pengamatan uji glukanase tersaji pada Tabel 6.
Tabel 6. Hasil pengukuran diameter zona bening pada uji awal glukanase Kode Bakteri Diameter Zona Bening (mm)
PB3A 1,5
2000). Strain Paenibacillus sp. 300 dan Streptomyces sp. 385 menghasilkan kitinase dan β-1,3-glukanase dalam media kultur (Singh et al. 1999).
Pada penelitian ini perubahan morfologi hifa yang paling mendominasi yaitu lisis pada dinding sel (Gambar 10E dan 10F) disertai koagulasi sitoplasma yaitu sebesar 40%. Berdasarkan perubahan tersebut bisa disimpulkan bahwa kemungkinan glukanase yang dikeluarkan oleh isolat bakteri potensial berperan dalam mekanisme menghambat pertumbuhan Saprolegnia sp. secara in vitro. Hal ini sesuai dengan laporan oleh Diby et al. (2005) yaitu glukanase memiliki mekanisme melisiskan dinding sel Saprolegnia dan menyebabkan koagulasi sitoplasma Phytophtora capsici setelah diuji antagonis dengan Pseudomonas fluorescens. Glukanase dan kitinase yang dihasilkan oleh Pichia membranafaciens dan Candida guillermondii merupakan enzim hidrolitik yang berperan dalam penghambatan terhadap Rhizopus stolonifer dan terdapat mekanisme yang sinergis diantara keduanya telah dilaporkan juga oleh Qing et al. (2002).
Isolat bakteri potensial juga kemungkinan memiliki mekanisme yang lain dalam melakukan penghambatan Saprolegnia sp. selain dengan menghasilkan enzim
hidrolitik. Probiotik Aeromonas media A199 menghasilkan substansi penghambat ekstraseluler terhadap S. parasitica yaitu Indol (T1) (Lategan et al. 2006). Pada penelitian ini sitoplasma hifa juga mengalami perubahan pada sitoplasma yang
menyebabkan hifa terpuntir atau terputus (Gambar 10G dan 10H) sehingga
kemungkinan terdapat substansi indol yang dikeluarkan oleh isolat bakteri.
Mekanisme lain yang dapat terjadi adalah kompetisi isolat bakteri terhadap logam
tertentu. P. fluorescens melakukan penghambatan terhadap S. parasitica dengan melakukan kompetisi terhadap besi melalui sistem penangkapan besi siderophore (Hatai & Willoughby 1988).
kemampuan dalam menghambat patogen dari akar gandum yaitu Gaeumannomyces dan Pythium (Gurusiddaiah et al. 1986).
4.4. Hasil Patogenitas Saprolegnia sp.
Patogenitas isolat Saprolegnia sp. dilakukan untuk mengetahui kemampuan isolat ini menyebabkan penyakit saprolegniasis pada telur serta dalam tahapannya hingga menjadi larva. Hasil patogenitas menunjukkan bahwa tingkat infeksi isolat
Saprolegnia sp. yang diinokulasikan pada telur kelompok perlakuan adalah sebesar 15% dan berbeda nyata dengan kelompok kontrol (P<0,05). Tingkat kematian telur gurami yang diinokulasikan Saprolegnia sp. pada kelompok perlakuan adalah sebesar 33,33% dan berbeda nyata dengan kelompok kontrol 6,67 % (P<0,05). Daya tetas telur kelompok perlakuan adalah sebesar 76,66% dan berbeda nyata dengan kelompok kontrol 93,3% (P<0,05)
Tingkat infeksi, tingkat kematian dan daya tetas pada kelompok perlakuan menunjukkan perbedaan nyata dengan kelompok kontrol. Hal ini menunjukkan
bahwa isolat Saprolegnia sp. yang diambil dari lesi pada telur di pembenihan bersifat patogen terhadap sel telur. Hanjavanit et al. (2008) telah melaporkan bahwa isolat beberapa spesies Saprolegnia dapat diinfeksikan secara skala laboratorium dan bersifat patogen terhadap telur ikan lele. Walaupun beberapa laporan menyebutkan bahwa infeksi buatan pada telur ikan membutuhkan beberapa stres lingkungan agar gejala klinis jelas terlihat. Pada penelitian ini gejala klinis yang tampak terdapat pada telur yang mati sedangkan pada tahapan telur yang masih hidup atau sakit gejala klinis tidak terlihat nyata seperti telur yang mati.
Saprolegniasis merupakan penyakit dengan gejala yang umum terlihat pada permukaan kulit seperti bentukan kapas putih pada ikan ataupun telur (Bruno & Wood 1999). Gejala klinis yang tampak pada tiap tahapan telur menjadi larva berbeda. Telur gurami yang terinfeksi isolat Saprolegnia sp. memiliki gejala klinis yaitu telur mengalami perubahan warna menjadi lebih pucat dan dalam 24 jam akan tampak hifa menyelimuti permukaan telur. Tahapan awal perubahan gejala klinis
tidak terlalu terlihat pada telur yang belum memiliki ekor dan mata (umur 4 hari). Pada tahap telur yang memiliki ekor dan mata (umur 7 hari), gejala klinis yang terlihat adalah kemerahan pada bagian abdomen, tidak berenang aktif sehingga hanya mengapung di pinggir wadah, sisa kuning telur lebih banyak dibandingkan normal dan pada tubuh seperti sirip atau kulit terdapat bentukan putih. Bruno & Wood (1999) menyebutkan bahwa gejala klinis infeksi awal adalah lesi kulit berwarna putih atau abu-abu yang kemudian dapat dengan berkembang cepat menyebabkan kerusakan pada kulit dan otot sehingga ikan akan lemah dan kehilangan keseimbangan. Berdasarkan hal ini menunjukkan pada telur umur 7 hari adalah tahapan awal infeksi. Gambaran perubahan jaringan telur gurami disajikan pada Gambar 12.
Gambar 12. Perubahan lapisan bagian luar telur umur 4 hari dan perubahan epidermal dan dermal telur umur 7 hari yang berhubungan dengan lesi yang disebabkan oleh Saprolegnia sp. (A) Kontrol, (B) Lapisan paling luar telur terdapat perubahan berupa peradangan dan degenerasi vacuolar (B1). (C) Kontrol (D) Perubahan pada epidermal dan dermal menunjukkan degenerasi vakuolar pada bagian epidermal (D1) disertai peradangan pada bagian otot (D2)
A B
C D
D1
Perubahan histologi menunjukkan bahwa infeksi Saprolegnia sp. menyebabkan peradangan dan degenerasi vakuolar pada bagian lapisan luar telur yang terdapat hifa Saprolegnia. Perubahan pada lapisan telur ini disebabkan Saprolegnia mengeluarkan enzim untuk mendegradasi komponen lapisan pada telur. Pada telur umur 7 hari maka lapisan dermal dan epidermal telur menunjukkan perubahan peradangan dan degenerasi vakuolar. Perubahan yang sama dilaporkan oleh Giesker et al. (2006) yaitu infeksi Saprolegnia pada ikan salmon Onchorhyncus mykiss ditemukan hemorrhagi dan peradangan mononuklear pada bagian tepi lesi, degenerasi vakuolar pada sel epitel dan degenerasi pada jaringan pengikat dan otot. Gejala klinis dan perubahan jaringan telur gurami serta tahapan telur menjadi larva menunjukkan bahwa isolat Saprolegnia sp. mampu menyebabkan infeksi sehingga bersifat patogen bagi telur.
Telur terinfeksi diambil sampelnya dan diisolasi pada media GYA kemudian dilakukan pengamatan morfologi. Hasil uji reisolasi pada telur yang terinfeksi menunjukkan bahwa telur tersebut yang terinfeksi oleh isolat yang sama dengan isolat
Saprolegnia sp. yang diinfeksikan. Hal ini menunjukkan telur yang terinfeksi pada penelitian ini disebabkan karena isolat Saprolegnia sp.
4.5. Hasil Patogenitas dan Perlekatan Bakteri Kitinolitik
Uji patogenitas dilakukan untuk mengetahui apakah isolat bakteri tersebut bersifat patogen terhadap telur gurami. Uji patogenitas merupakan salah satu tahapan penting dalam menentukan mikroba yang bisa digunakan sebagai probiotik. Mikroba yang digunakan sebagai kontrol biologi dalam akuakultur harus memiliki beberapa persyaratan satu diantaranya adalah mikroba tersebut harus tidak menyebabkan penyakit atau patogen terhadap inang (Verschuere et al. 2000).
Patogenitas bakteri PB01 terhadap telur dan larva yang sudah menetas dipelajari pada penelitian ini dengan pengamatan jaringan pada telur hari ke empat dan larva ikan. Gambaran histologi telur ikan normal yaitu telur terdiri dari dua lapisan yaitu lapisan terluar berwarna merah terang dan lapisan bagian dalam berwarna merah muda (Hanjavanit et al. 2008). Pada telur ikan gurami yang dinokulasi isolat bakteri PB01 terdapat proliferasi sel radang dan kongesti pada lapisan luar telur. Hal ini menunjukkan adanya peradangan pada mukosa telur. Gambaran histologi larva ikan yang diinkulasikan PB01 menunjukkan perubahan proliferasi sel radang pada kulit, edema dan proliferasi sel radang pada otot, disertai degenerasi hidropis pada hati dan ginjal. Perubahan patologi telur dan larva disajikan pada Gambar 13.
Gambar 13. Gambaran histologi telur dan larva ikan. (A) Telur kontrol menunjukkan histologi normal (B) Telur yang diinokulasikan bakteri PB01 menunjukkan akumulasi darah (kongesti) pada lapisan luar telur (B1). (C) Kulit dan otot larva ikan kontrol. (D) Larva ikan diinokulasikan bakteri PB01 menunjukkan peradangan disertai edema pada otot dan epithelial lifting pada kulit (D1:D2)
A B
C D
B1
D1