MODEL HABITAT TRENGGILING DI TAMAN NASIONAL
GUNUNG HALIMUN SALAK
AFROH MANSHUR
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Model Habitat Trenggiling di Taman Nasional Gunung Halimun Salak adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
AFROH MANSHUR. Model Habitat Trenggiling di Taman Nasional Gunung Halimun Salak. Dibimbing oleh AGUS PRIYONO KARTONO dan BURHANUDDIN MASYUD.
Trenggiling (Manis javanica) merupakan salah satu mamalia terancam punah berdasarkan kategori International Union for Conservation of Nature. Hal ini disebabkan oleh 3 hal, yakni: (1) keterbatasan dalam proses evolusi, (2) kemampuan reproduksi yang rendah, serta (3) tingginya perdagangan ilegal terhadap jenis ini. Upaya konservasi M. javanica masih sangat terbatas mengingat kajian ekologi jenis ini belum banyak diungkap. Salah satu aspek kajian yang penting diketahui adalah habitat M. javanica. Hal ini karena habitat merupakan dasar dalam ekosistem fungsional sehingga informasi terkait hal ini berguna dalam mendukung strategi pelestarian satwa dan upaya mengelola populasinya. Oleh karena itu, tujuan penelitian ini adalah untuk mengetahui: (1) karakteristik habitat M. javanica , (2) komponen lingkungan yang berpengaruh terhadap kehadiran M. javanica, (3) pola seleksi habitat M. javanica dan (4) model kehadirannya berdasarkan komponen lingkungan tersebut.
Identifikasi kehadiran M. javanica diketahui dengan metode eksplorasi dengan penelusuran jejak. Guna efektivitas waktu dan biaya dalam pengamatan, maka digunakan teknik open grid. Lokasi yang diketahui merupakan habitat M. javanica kemudian diukur seluruh komponen habitatnya dengan plot pengamatan berbentuk plot tunggal seluas 1 ha. Adapun komponen tersebut secara umum terdiri dari: komposisi vegetasi, pakan, kompetisi, predasi, tutupan vegetasi, suhu, kelembaban udara, ketinggian tempat, tekstur tanah dan kelerengan lahan.
Hasil penelitian menunjukkan bahwa M. javanica menggunakan 9 komponen habitat dengan karakteristik khusus berdasarkan uji runs. Penggunaan areal sebagai habitat oleh jenis ini dipengaruhi oleh kehadiran kompetitornya, yakni jika terdapat kompetitor maka M. javanica tidak akan menggunakan habitat tersebut meskipun memiliki karakteristik yang sesuai. Kehadiran M. javanica pada suatu lokasi ditentukan oleh 8 komponen habitat yang terdiri dari 3 komponen biotik dan 5 komponen abiotik. Sifat selektif M. javanica diketahui dari 3 fungsi habitat, yakni: (1) habitat makan yang berkorelasi positif dengan jumlah jenis tumbuhan sarang mangsanya; (2) habitat istirahat yang berkorelasi negatif dengan persentase fraksi debu, ketinggian tempat dan keberadaan kompetitornya serta (3) habitat reproduksi dan mengasuh anak yang berkorelasi positif dengan suhu udaranya. Adapun model terbaik yang terbentuk dalam pendugaan kehadiran M. javanica disuatu lokasi lebih dipengaruhi oleh komponen biotiknya dengan model regresi y= 1.467 - 0.093 *ZJumlah jenis tumbuhan sebagai sarang mangsa - 0.091Z*Dominasi tumbuhan sarang mangsa pada tingkat pohon -
0.089*ZJarak terhadap kompetitor (r =0.472, sig.= 0.076).
SUMMARY
AFROH MANSHUR. Habitat models of Pangolin (Manis javanica) in Mount Halimun Salak National Park. Supervised by AGUS PRIYONO KARTONO and BURHANUDDIN MASYUD.
Pangolin (Manis javanica) is one of the threatened mammals based on the category of the International Union for Conservation of Nature. This is caused by 3 reason, i.e.: (1) the limitations in the evolutionary process, (2) a low reproductive ability, and (3) high illegal trade in this species. M. javanica’s conservation efforts is still limited considering the ecological study of this species hasn’t been revealed. One of the important aspects which should be known is M. javanica’s habitat. This is because the habitat is basic in the ecosystem functional so that this information related to this useful in supporting wildlife conservation strategies and efforts to manage the population.Therefore, the objectives of this study was to determine: (1) the characteristics of M. javanica habitat, (2) environmental components that influence the presence of M. javanica, (3) habitat selection patterns of M. javanica and (4) a model based on the presence of the environmental component.
Identification of the presence of M. javanica known by the exploration methods based by traces. To the effectiveness time and cost of the observation, then used open grid method. The location is to be the M. javanica’s habitat then
measured of all components habitat with a single plot covering an area of 1 ha. The components are generally consists of: the vegetation composition, feed, competition, predation, vegetation cover, temperature, humidity, altitude, soil texture and slope steepness.
The results showed that M. javanica using the 9 components habitats with special characteristics based on runs test. The area that used by this species is
affected by the presence of it’s competitors, which is if there are competitors then
M. javanica will not use these habitats despite having the appropriate characteristics. The presence of M. javanica at a location determined by the 8 habitat components, consists of 3 biotic components and 5 abiotic components. selectivity M. javanica can detect by 3 functions of the habitat, are: (1) eating habitat that positively correlated with the number of plant species used as his prey digs; (2) resting habitat that negatively correlated with the percentage fraction of dust; altitude and the presence of his competitors, and (3) reproduction and parenting habitat that positively correlated with environment temperature. The best model is formed to estimation the presence of M. javanica in a location more influenced by biotik component with regression model are y = 1467-0093 * ZNumber of plant species that used as his prey den - 0.091ZDominance of plant species used as his prey den on tree level - 0089 * ZDistance from his competitors (r = 0.472, sig. = 0.076).
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Konservasi Biodiversitas Tropika
MODEL HABITAT TRENGGILING (
Manis javanica
)
DI TAMAN NASIONAL GUNUNG HALIMUN SALAK
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Penelitian ini berjudul Model Habitat Trenggiling di Taman Nasional Gunung Halimun Salak yang dilaksanakan pada bulan Februari hingga Juni 2014. Adapun karya ilmiah yang memuat salah satu sub bagian dari tulisan ini diterbitkan pada Jurnal Media Konservasi (ISSN: 0215-1677) Volume 10 No 1 Bulan April 2015 dengan judul Karakteristik Habitat Trenggiling Jawa (Manis javanica) di Taman Nasional Gunung Halimun Salak.
Terima kasih penulis ucapkan kepada Bapak Dr Ir Agus Priyono Kartono, MSi dan Bapak Dr Ir Burhanuddin Masyud, MS selaku pembimbing serta Prof.(RIS) Dr. Ir. Ibnu Maryanto M.Si. selaku penguji luar pada sidang tesis. Penghargaan penulis juga disampaikan kepada Romi Prasetyo, S. Hut yang telah membantu dalam analisis data serta pemetaan dan kepada Balai Taman Nasional Gunung Halimun Salak yang telah memberikan saran mengenai lokasi penelitian. Ungkapan terima kasih juga disampaikan kepada istri, bapak, ibu, serta seluruh keluarga, atas segala doa dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN vii
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 3
Manfaat Penelitian 3
Kerangka Pemikiran 3
Ruang Lingkup Penelitian 5
2 METODE 5
Lokasi dan Waktu Penelitian 5
Objek dan Alat Penelitian 6
Metode Pengumpulan Data 6
Prosedur Analisis Data 11
3 HASIL DAN PEMBAHASAN 14
Hasil 14
Pembahasan 24
4 SIMPULAN DAN SARAN 36
Simpulan 36
Saran 37
DAFTAR PUSTAKA 37
LAMPIRAN 47
DAFTAR TABEL
1. Variabel karakteristik habitat yang diukur dan asumsi pengaruhnya
terhadap hidup strategi hidup M. javanica 8
2. Variabel komponen habitat yang diukur dalam pendugaan kebutuhan
hidup M. javanica 10
3. Karakteristik habitat yang digunakan M. javanica sesuai dengan
kebutuhan hidupnya 12
4. Hasil uji statistik non parametrik terhadap komponen habitat M. javanica 18 5. Hasil uji analisis faktor pada komponen biotik dan abiotik habitat yang
digunakan oleh M. javanica 19
6. Ringkasan nilai akar ciri, persentase variasi data dan korelasi fungsi
habitat dan komponen lingkungan 20
7. Vektor ciri dan bobot setiap variabel penyusun PCA 23 8. Hasil analisis regresi terhadap ketiga faktor baru yang terbentuk untuk
menduga kehadiran M. javanica 23
9. Hasil perbandingan nilai temuan dengan nilai prediksi dari model regresi
yang terbentuk 24
DAFTAR GAMBAR
1. Kerangka pikir penelitian model pendugaan habitat M. javanica 4
2. Peta lokasi penelitian M. javanica di TNGHS 5
3. Sketsa plot ukur pengambilan data variabel habitat M. javanica 7 4. Dua pola berjalan pada M. javanica ketika bersifat terestrial 14 5. Pola berjalan M. javanica ketika bersifat arboreal 15 6. Cetakan yang terbentuk pada bidang pijakan daun dan kayu 15 7. Posisi ekor M. javanica ketika berjalan sebagai penyebab arah rebah yang
seragam pada tumbuhan bawah yang dilaluinya 16
8. Tipe sarang aktif M. javanica yang teridentifikasi di TNGHS 17 9. Diagram ordinasi seleksi habitat dan komponen habitat M. javanica
berdasarkan sumbu 1 dan sumbu 2 21
10.Diagram ordinasi seleksi habitat dan komponen habitat M. javanica
berdasarkan sumbu 1 dan sumbu 3 22
DAFTAR LAMPIRAN
1. Daftar jenis tumbuhan kayu yang teridentifikasi di lokasi penelitian 47 2. Hasil uji normalitas data pada komponen habitat M. javanica 51 3. Hasil uji korelasi pearson komponen biotik untuk mengetahui autokorelasi
antar variabel. 53
4. Hasil uji korelasi pearson komponen abiotik untuk mengetahui adanya
autokorelasi antar variabel 54
5. Hasil uji toleransi dan VIF data komponen biotik untuk mengetahui
adanya multikolinearitas antar variabel 54
6. Hasil uji toleransi dan VIF data komponen biotik untuk mengetahui
adanya multikolinearitas antar variabel 55
7. Hasil uji reduksi faktor untuk mengetahui komponen habitat biotik yang
berpengaruh terhadap kehadiran M. javanica 55
8. Analisis komponen PCA untuk mengetahui komponen baru pada habitat biotik yang berpengaruh terhadap kehadiran M. javanica 57 9. Hasil uji reduksi faktor untuk mengetahui komponen habitat abiotik yang
berpengaruh terhadap kehadiran M. javanica 58
10.Hasil analisis regresi terhadap variabel komponen biotik habitat untuk
menduga kehadiran kehadiran M. javanica 62
11.Hasil analisis regresi terhadap variabel komponen abiotik habitat untuk
1
1
PENDAHULUAN
Latar Belakang
Manis javanica merupakan satu-satunya Pholidota yang terdapat di Indonesia, yakni di Pulau Sumatera, Jawa dan Kalimantan (Corbet & Hill 1992). Jenis lain yang telah punah dan pernah teridentifikasi di Pulau Jawa dan Kalimantan adalah M. palaeojavanica (Dubois 1926; Cranbrook 2010). Laju penurunan populasi pada genus Manis mencapai 9.12% selama 45 tahun terakhir (Wu et al. 2004a). Hal ini menunjukkan tingginya tingkat ancaman kepunahan pada jenis ini. Terdapat empat faktor utama yang menyebabkan tingginya laju penurunan populasi pada genus Manis, yakni: perdagangan ilegal, pakan terbatas dan spesifik, daya reproduksi yang rendah dan kemampuan anti predator yang rendah (Challender 2011; Wu et al. 2004b). Kondisi ini menyebabkan status konservasi M. javanica yang meningkat pada tahun 2013 menjadi kritis (critically endangered) berdasarkan International Union for Conservation of Nature (IUCN).
Perdagangan satwa merupakan salah satu faktor eksternal yang perlu menjadi perhatian penting dalam upaya konservasi M. javanica. Hal ini dikarenakan teknik budidaya satwa ini belum sepenuhnya dikuasai oleh para praktisi. Kasus perdagangan ilegal M. javanica di Indonesia merupakan yang terbesar kedua setelah Vietnam dengan jumlah temuan hingga 15.7 ton dalam kurun waktu 2008-2011 (Challender 2011). Besarnya jumlah perdagangan ilegal ini disebabkan oleh tingginya permintaan pasar dan nilai ekonomi M. javanica. Hal ini dikarenakan beragamnya manfaat M. javanica terutama sebagai bahan obat-obatan yang mampu menyembuhkan berbagai penyakit seperti seperti: asma, reumatik, kanker, memperlancar sistem peredaran darah, dan afrodisiak (Douglas 1957; Huang et al. 2004). Khusus pada sisik M. javanica diketahui memiliki kekuatan magis (de Heusch et al. 1993) bahkan sisik M. javanica juga diketahui memiliki kandungan Tramdol HCL (Farida 2012; Maryanto et al. 2013) yang merupakan kimia utama menyusun anestesi seperti shabu-shabu.
2
ekologi habitat M. javanica perlu diketahui secara rinci dalam rangka konservasi jenis ini. Terlebih lagi M. javanica juga dikategorikan sebagai 100 mamalia EDGE (Evolutionarily Distinct and Globally Endangered) oleh ZSL (Zoological Society of London). Hal ini mengindikasikan bahwa spesifikasi habitat merupakan strategi utama M. javanica dalam bertahan hidup.
Berbagai tinjauan studi terkait M. javanica yang telah dilaporkan diantaranya terfokus pada analisis anatomi dan morfometri (Wu et al. 2004c; Nisa’ et al. 2010; Zhou et al. 2012), histologi (Pongchairerk et al. 2008; Meyer et al. 2010) serta pembuktian kemampuannya sebagai obat melalui analisis struktur dan kandungan bioaktif di dalam tubuhnya (Huang et al. 2004). Penelitian ekologi M. javanica yang dilaporkan hanya terkait dengan luas wilayah jelajah dan perilaku hariannya (Lim & Ng 2007). Kondisi ini berbeda dengan ordo Pholidota lain yang telah banyak dilakukan kajian ekologinya, sebagai contoh adalah M. pentadactyla yang sudah diketahui pola penggunaan habitat (Wu et al. 2003), perilaku harian, pakan (Wu et al. 2004a) dan perilaku reproduksinya termasuk perilaku mengasuh anak (Heath & Vanderlip 1988; Chao et al. 1993; Wu 1998). Kajian ekologi juga dilakukan pada spesies M. temminckii terkait luas wilayah jelajah (Heath & Coulson 1997a), aktivitas harian, pakan dan perilaku makannya (Jacobsen et al. 1991; Richer et al. 1997; Swart et al. 1999). Berdasarkan tinjauan ini, maka dapat disimpulkan bahwa informasi habitat yang merupakan kunci dalam pemanfaatan M. javanica yang sesuai dengan peraturan perundangan di Indonesia belum diketahui.
Salah satu lokasi yang merupakan bagian habitat M. javanica di Jawa Barat dan Banten adalah kawasan hutan di Taman Nasional Gunung Halimun Salak (TNGHS) yang terletak di Kabupaten Bogor, Sukabumi dan Lebak. Belum adanya informasi dan pengetahuan mengenai ekologi M. javanica di kawasan ini menyebabkan konservasi pada jenis ini hanya mengandalkan efektivitas pengelolaan secara umum pada kawasan di TNGHS meskipun secara individual jenis ini juga dilindungi berdasarkan peraturan perundangan di Indonesia maupun konvensi internasional. Oleh karena itu, mengacu pada lokasi dan kondisi di atas, informasi detail mengenai penggunaan habitat M. javanica perlu diteliti.
Perumusan Masalah
Geldmann et al. (2013) menunjukkan bahwa kawasan konservasi yang ada belum mampu melindungi seluruh spesies di dalamnya meskipun cukup efektif dalam penurunan laju kerusakan habitat di kawasan tersebut. Penelitian sebelumnya dari Catullo et al. (2008) menunjukkan bahwa hanya sebanyak 32.7% jenis mamalia di kawasan Asia Tenggara yang memiliki kesesuaian habitat di dalam kawasan konservasi. Hal ini dikarenakan dalam penetapan kawasan konservasi, pada umumnya memiliki tujuan yang beragam, tidak terkecuali di TNGHS. Informasi yang sedikit terkait ekologi habitat M. javanica menyebabkan belum adanya lembaga konservasi yang berhasil dalam upaya penangkaran jenis ini. Beberapa aspek penting ekologi habitat yang perlu di jawab dalam kaitannya konservasi jenis ini baik secara in situ maupun ex situ adalah:
1. Bagaimana karakteristik habitat yang digunakan oleh M. javanica?
3 3. Bagaimana M. javanica menyeleksi habitat yang akan digunakan sesuai dengan
kebutuhan hidupnya?
4. Bagaimana hubungan antara M. javanica di alam dan komponen habitatnya? Tujuan Penelitian
Tujuan penelitian ini adalah:
1. Mengidentifikasi karakteristik habitat M. javanica berdasarkan pemanfaatan ruang.
2. Mengidentifikasi komponen habitat yang mempengaruhi kehadiran M. javanica pada suatu lokasi.
3. Mengidentifikasi pola seleksi habitat oleh M. javanica.
4. Menentukan model kehadiran M. javanica di alam berdasarkan komponen habitatnya.
Manfaat Penelitian
Secara teoritis hasil penelitian ini diharapkan mampu memberikan sumbangan bagi ilmu pengetahuan terutama informasi mengenai ekologi habitat M. javanica di Indonesia. Secara teknis praktis, informasi dari hasil penelitian ini dapat menjadi rujukan dalam dasar penentuan strategi konservasi M. javanica baik di in situ maupun ex situ. Pengetahuan terkait habitat M. javanica dapat digunakan sebagai dasar penentuan metode inventarisasi populasi dan parameter demografi lainnya yang cocok bagi jenis ini. Informasi tersebut selanjutnya dapat digunakan sebagai dasar penentuan kebijakan pemanfaatan langsung di alam sesuai dengan peraturan perundangan yang berlaku di Indonesia. Pada strategi konservasi ex situ, informasi terkait karakteristik habitat dan komponen habitatnya yang berpengaruh terhadap kehadiran M. javanica dengan dapat digunakan sebagai dasar manipulasi kondisi lingkungan di lembaga konservasi sehingga peluang keberhasilan dalam pemeliharaan M. javanica akan lebih tinggi.
Kerangka Pemikiran
4
Perilaku satwa merupakan dasar dalam penelitian ekologi habitat karena merepresentasikan pola pemanfaatan habitat oleh suatu spesies (Hall et al. 1997). Karakteristik habitat merupakan ciri dari komponen habitat yang dapat menunjukkan fungsi habitat bagi satwa yang memanfaatkannya (Morrison et al. 2006). Oleh karena itu, pendekatan yang digunakan dalam mengidentifikasi karakteristik habitat M. javanica sebaiknya didasarkan pada perilakunya. Skala informasi yang lebih detail mengenai karakteristik habitat dapat digunakan sebagai dasar dalam mempelajari adanya seleksi habitat oleh suatu spesies (Morrison et al. 2006). Seleksi habitat didefinisikan sebagai proses satwaliar memilih komponen habitat yang digunakan sesuai dengan perilaku alami dan pengalamannya (Johnson 1980). Dalam konteks pemilihan habitat, satwa cenderung dinamis akibat adanya
5 kebutuhan dalam pencarian habitat optimal (Stamps & Davis 2006). Oleh karena itu, diperlukan informasi detail mengenai komponen habitat yang benar-benar digunakan oleh M. javanica untuk menunjang keberlangsungan hidupnya. Pada akhirnya ketika informasi ini diperoleh, maka akan diperoleh suatu pola terkait kehadiran M. javanica di alam berdasarkan komponen habitat tersebut.
Ruang Lingkup Penelitian
Ditinjau dari aspek kewilayahan, ruang lingkup penelitian berada di kawasan Taman Nasional Gunung Halimun Salak (TNGHS). Oleh karenanya, hasil penelitian ini hanya mampu menunjukkan dugaan pola pemanfaatan habitat M. javanica pada kondisi ekosistem yang serupa dengan lokasi penelitian. Ruang lingkup penelitian secara substansial tertuju pada pemahaman karakteristik penggunaan habitat, penentuan komponen habitat yang mempengaruhi kehadiran M. javanica, pola seleksi habitat, serta pada perumusan model kehadiran M. javanica berdasarkan komponen habitat yang diukur.
2
METODE
Lokasi dan Waktu Penelitian
Penelitian dilaksanakan di tiga Resort TNGHS, yaitu Gunung Butak, Gunung Botol (SPTN Wilayah II Bogor) dan Gunung Kendeng (SPTN Wilayah III Sukabumi). Gambar 2 menunjukkan letak wilayah pengamatan M. javanica yang termasuk dalam areal pengelolaan Balai Taman Nasional Gunung Halimun Salak.
6
Penelitian dilaksanakan selama empat bulan, yakni survey pendahuluan yang dilaksanakan selama seminggu pada akhir bulan Februari 2014 dan pengambilan data lapangan yang dilakukan pada tanggal 20 Maret - 7 Juni 2014.
Objek dan Alat Penelitian
Objek yang digunakan dalam penelitian ini adalah M. javanica dan struktur komunitas habitat alaminya. Alat yang digunakan antara lain: alat tulis, headlamp dengan mode infrared, timbangan, meteran, jangka sorong, busur, thermometer dry-wet, GPS model Garmin 76CSXi, Suunto clinometers, kamera, lensa mode fish eye, tabung spesimen, detergen non busa dan akuades. Terkait analisis data, digunakan komputer dengan softwere ArcGIS 10.3, Global Mapper ver 13.0, Canoco 4.56, HemiView Canopy Analysis 2.1 SR1 dan IBM SPSS Statistic 20.0.0.
Metode Pengumpulan Data Penentuan Plot Pengukuran
Penentuan petak pengukuran mengacu pada kehadiran M. javanica pada suatu lokasi. Metode yang digunakan untuk mengetahui kehadiran jenis ini adalah metode eksploratif. Efektivitas metode tersebut dapat dilakukan menggunakan teknik open grid pada Explorer Search and Rescue (ESAR). Metode ini dilakukan dengan membagi dua kelompok pengamat dengan titik awal berupa jejak terjauh yang dijumpai. Masing-masing kelompok pengamat kemudian berjalan memotong kontur dengan jalur pengamatan berbentuk semi radial pada lokasi penelitian hingga diketahui arah pergerakan yang menunjukkan ke satu titik lokasi yang diyakini merupakan sarang M. javanica. Kehadiran M. javanica pada suatu habitat berdasarkan hasil penelitian setidaknya dapat dilihat dari 3 hal, yakni: jejak kaki, jejak ekor dan bekas lokasi makan.
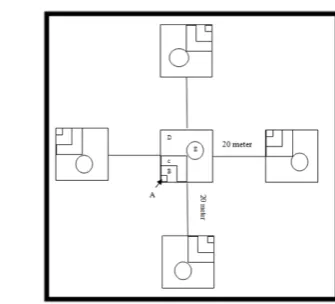
Guna keseragaman dalam pendugaan pemanfaatan habitat oleh M. javanica, digunakan asumsi dalam penelitian ini, yaitu: (1) lokasi makan dicirikan dengan adanya sarang pakan yang rusak akibat cakaran M. javanica; (2) lokasi istirahat dicirikan dengan keberadaan sarang aktif yang dibuktikan dengan kehadiran individu M. javanica; (3) lokasi interaksi sosial dicirikan dengan keberadaan individu lain baik sejenis maupun tidak dalam radius 100 m (Taylor & Knight 2003); dan (4) lokasi reproduksi dan mengasuh anak dicirikan dengan keberadaan dua individu M. javanica dewasa berbeda jenis kelamin atau keberadaan anak dan induk betinanya dalam habitat dan waktu yang sama. Asumsi ini digunakan sebagai dasar dalam penentuan fungsi habitat. Pencatatan lokasi dengan GPS dan pembuatan plot pengukuran komponen habitatnya seluas 1 hektar dilakukan setelah fungsi habitat teridentifikasi. Sketsa lokasi pengamatan dibuat dengan mengacu pada Deuser & Shugart (1978) dengan beberapa modifikasi (Gambar 3).
Karakteristik Habitat
7
Komposisi Vegetasi
Data komposisi vegetasi diperoleh dengan metode plot tunggal. Plot ukur awal berada di titik perjumpaan dengan M. javanica kemudian dibuat 4 plot lain mengikuti penjuru mata angin (Gambar 3). Notasi-notasi yang digunakan sesuai pada Gambar 3 adalah : A = inventarisasi untuk tingkat semai (4 m2), B =
inventarisasi untuk tingkat pancang (25 m2), C = inventarisasi untuk tingkat tiang
(100 m2), D = inventarisasi untuk tingkat pohon (400 m2). Data yang dikumpulkan
adalah nama jenis, jumlah individu, diameter tumbuhan pada tingkat tiang dan pohon serta identifikasi jenis tumbuhan yang digunakan oleh semut (Hymnoptera) atau rayap (Isoptera) sebagai sarangnya. Nilai tersebut digunakan untuk memperoleh nilai kerapatan tumbuhan pada tiap tingkat jenis, dominansi tumbuhan dan jumlah jenis tumbuhan yang dimanfaatakan sebagai sarang pakan.
Pakan
Pakan M. javanica yang diukur hanyalah pakan yang teridentifikasi di dalam plot ukur (1 ha). Pakan yang teridentifkasi kemudian dilakukan pencatatan lokasi berdasarkan sarang koloni terbesarnya dengan GPS untuk selanjutnya dianalisis jaraknya terhadap lokasi M. javanica dengan Global Mapper ver 13.0. Adapun variabel yang dicatat meliputi jumlah koloni dan jenis ordo (Isoptera atau Hymnoptera).
Kompetitor dan Predator
Keberadaan satwa kompetitor dan predator dilakukan menggunakan metode eksplorasi dan pengamatan tidak langsung berdasarkan feses, jejak kaki dan cakaran (Wilson et al. 1996). Kompetitor M. javanica setidaknya dapat dikategorikan pada dua hal, yakni kesamaan pola pemanfaatan ruang dan kesamaan jenis pakan. Jenis satwa yang memiliki kesamaan pemanfaatan ruang adalah: Leporidae, Tupaiidae (Nakagawa et al. 2007), Muridae (Buckle et al. 1997), Hystricidae (Morin et al. 2005), dan Paradoxurus hermaproditus (Mudappa et al. 2010). Jenis satwaliar yang memiliki kesamaan pemanfaatan pakan antara lain: Agamidae (Abensperg-Traun & Steven 1997), Scincidae (James 1991), Tropiduridae (Faria & Araujo 2004), Lorisidae (Weins et al. 2006), dan Suidae (Herrero et al. 2006). Predator potensial bagi M. javanica di Jawa Barat adalah Panthera pardus (Kawanishi & Sunquist 2004) dan Python reticulatus (Shine et al. 1999). Khusus untuk kompetitor mamalia
8
kecil digunakan metode jebakan menggunakan model Tomahawk life trap (Pearson & Ruggiero 2003). Jebakan ditempatkansecara acak sebanyak sepuluh buah dan diambil kembali setelah 24 jam. Metode jebakan juga digunakan untuk kompetitor jenis kadal dengan menggunakan papan berbentuk segi empat yang diberi lem perekat (Ribeiro-Júnior et al. 2006). Setiap titik perjumpaan dengan satwa kompetitor dan predator dilakukan penandaan menggunakan GPS untuk selanjutnya dianalisis jarak temuan terdekat dengan lokasi kehadiran M. javanica. Jarak dengan Aktivitas Manusia
Analisis jarak kehadiran M. javanica dengan aktivitas manusia menggunakan bantuan ArcGIS 10.3. Definisi gangguan antropogenik yang dimaksud adalah gangguan indikatif sebagai ancaman bagi satwaliar yang dinilai berdasarkan jarak habitat dari lokasi budidaya dan atau pusat beraktivitas manusia seperti jalan, sawah, ladang maupun perkebunan. Data mengenai tutupan lahan dianalisis berdasarkan Citra Landsat Path/Row: 122/65 liputan Tanggal 8 Agustus 2013.
Tabel 1 Variabel karakteristik habitat yang diukur dan asumsi pengaruhnya terhadap hidup strategi hidup M. javanica
Variabel
Karakteristik Asumsi Korelasi Sitasi
Komposisi vegetasi 1. Strategi anti predator. Hochman & Kotler (2007); Jensen et al. (2005) 2. Keanekaragaman jenis
pakan.
Gillison et al. (2003)
3. Efisiensi energi. Hof et al. (2012) Jenis pakan Kandungan gizi pakan dan
kemampuan mencernanya.
Kuantitas pakan 1. Efisiensi energi. 2. Kompetisi satwa.
Predator Resiko predasi Lima & Dill (1990) Suhu dan
Ketinggian Tempat 1. Suhu dan kandungan oksigen lingkungan
Korner (2007)
2. Keanekaragaman hayati Jones (2000) Tekstur tanah 1. Efisiensi energi McNab (1984)
2. Kemudahan membuat sarang
Farida (2010)
9 Suhu dan Kelembaban
Lokasi pengukuran terletak pada notasi E sesuai dengan Gambar 3. Pengukuran suhu udara (°C) dan kelembaban relatifnya (%) diukur dengan jarak 15 cm dari atas tanah (Rodríguez-Calcerrada et al. 2009), dilakukan tiap 15 menit sebanyak dua ulangan. Nilai yang digunakan adalah nilai rata-rata dari hasil dua ulangan pengukuran tersebut.
Ketinggian Tempat
Data ketinggian tempat diperoleh dengan merata-ratakan hasil pengukuran di lapangan dengan altimeter pada lokasi penemuan satwa (notasi E pada Gambar 3) dan nilai ketinggian tempat pada atribut peta Citra Aster GDEM S07E106 resolusi 30 m.
Tekstur Tanah
Sampel tanah diambil pada kedalaman 20 cm (Rodríguez-Calcerrada et al. 2009) pada notasi E sesuai Gambar 3. Masing-masing plot contoh terdiri dari dua sampel tanah. Fraksi tanah dianalisis menggunakan jar test (EPA 1991). Tekstur tanah dapat diketahui dengan memasukkan nilai rata-rata dari persentase masing-masing sampel fraksi tanah ke dalam segitiga tekstur tanah menurut Posadas et al. (2001). Hasil tekstur ini kemudian diklasifikasikan berdasarkan Hanafiah (2007). Kelerengan
Data kelerengan lahan (%) diperoleh dengan menggunakan clinometer yang dilakukan sebanyak lima ulangan pada masing-masing notasi E pada plot ukur sesuai Gambar 3. Data yang digunakan dalam analisis adalah nilai rata-rata kelerengan.
Komponen Utama, Seleksi dan Model Habitat
Komponen habitat yang dipilih sebagai variabel harus mampu merepresentasikan fungsi habitat bagi M. javanica. Sebanyak 34 komponen habitat biotik dan abiotik yang dipilih untuk menentukan komponen utama habitat dan pola seleksi habitat M. javanica disajikan dalam Tabel 2.
Komposisi Biotik dan Abiotik
Metode pengambilan data berupa komposisi vegetasi, pakan, kompetitor, predator dan jarak habitat dengan pusat aktivitas manusia, suhu, kelembaban serta ketinggian tempat. telah dijelaskan pada bagian karakteristik habitat. Khusus pada komposisi vegetasi pengukuran data juga dilakukan terhadap diameter tumbuhan pada tingkat tiang dan pohon. Data yang digunakan dalam analisis data ditunjukkan pada Tabel 2.
Tutupan Tajuk
10
Tekstur Tanah
Pengukuran tekstur tanah mengikuti metode pengukuran tekstur tanah pada analisis karakteristik habitat. Perbedaannya hanya pada data yang digunakan
Tabel 2 Variabel komponen habitat yang diukur dalam pendugaan kebutuhan hidup M. javanica
Sasaran Penelitian Variabel Sitasi
A. Komponen Biotik menjadi sarang pakan (x11)
Deperno et al. (2003) d. Dominansi pada tingkat pertumbuhan
tiang dan pohon (x12-x13) Wall & Shine
(2013) e. Dominansi tumbuhan yang menjadi
sarang pakan pada tingkat tiang dan pohon (x14-x15)
2. Pakan a. Jumlah koloni total (x16)
b. Jumlah koloni semut (x17) Wu et al. (2005)
c. Jumlah koloni rayap (x18)
d. Jarak rata-rata sumber pakan dengan
sarang aktif (x19) Hof & Bright
(2010) e. Jarak sumber pakan terdekat dengan
sarang aktif (x20)
3. Kompetitor a. Jumlah jenis kompetitor (x21) Loehle (2012)
b. Jarak terdekat dengan kompetitor (x22)
4. Predator a. Jarak terdekat dengan predator (x23) Lima & Dill
(1990) 5. Gangguan
Antropogenik
a. Jarak terdekat antara habitat dengan pusat aktivitas manusia (x24)
Taylor & Knight
3. Kelembaban Kelembaban relatif (x27) Avila-Flores &
Medellin (2004) 4. Ketinggian
tempat
Ketinggian tempat (x28) Korner (2007)
11 sebagai analisis data. Pada analisis seleksi habitat dan komponen utama habitat, data yang digunakan adalah data persentase masing-masing fraksi tanah.
Kelerengan Lahan
Metode pengukuran kelerengan lahan yang digunakan sama dengan pengukuran kelerengan pada pembahasan karakteristik habitat di atas. Perbedaannya hanya pada data yang digunakan sebagai analisis data. Pada analisis seleksi habitat dan komponen utama habitat, data yang digunakan adalah data kelerengan rata-rata, kelerengan terendah dan kelerengan tertinggi pada masing-masing plot pengukuran.
Prosedur Analisis Data Karakeristik Habitat
Seluruh komponen yang diukur dikategorisasi menjadi 5 kelas kecuali pada kelas tekstur tanah yang hanya 3 kelas. Nilai pengelompokkan data didasarkan pada sebaran nilai pengukuran variabel tersebut atau berdasarkan berbagai tinjuan studi khususnya di kawasan TNGHS. Adapun klasifikasi data kategorik di TNGHS, antara lain: kerapatan pohon (Nugroho 2012), kelas jumlah jenis tumbuhan yang digunakan sebagai sarang pakan M. javanica (Ke et al. 1999; Wu et al. 2005; Li et al. 2011), jarak sarang M. javanica terhadap kompetitor,predator dan lahan yang didominasi manusia (Taylor & Knight 2003), kelembaban relatif (Hidayati et al. 2012), ketinggian tempat (Lieberman et al. 1996; Nagy & Grabherr 2009), kelerengan lahan (SK Mentan Nomor 837/Kpts/Um/11/1980) dan tekstur tanah (Hanafiah 2007). Adapun kelas kategorik data tersebut disajikan dalam Tabel 3.
Seluruh uji tersebut merupakan uji non parametrik dan analisisnya menggunakan bantuan softwere IBM SPSS Statistic 20.0.0. Terdapat tiga analisis karakteristik habitat, yaitu:
1. Analisis komponen habitat yang digunakan oleh M. javanica dengan karaktristik khusus menggunakan uji run. Karakteristik khusus digunakan jika nilai uji signifikansi < 0.05.
2. Analisis komponen habitat dengan karakteristik tertentu yang menunjukkan kekhasan fungsi habitat bagi M. javanica menggunakan uji beda Kruskal – Wallis (H-test). Variabel yang berepengaruh diketahui jika signifikansi < 0.05. 3. Analisis komponen habitat yang berpengaruh terhadap penggunaan suatu fungsi
habitat tertentu dengan uji chi-square (χ2). Variabel tersebut diketahui dengan
ketentuan hasil uji signifikansi < 0.5.
Komponen Habitat yang Mempengaruhi Kehadiran M. javanica
Penentuan komponen habitat yang berpengaruh terhadap kehadiran M. javanica dilakukan dengan menggunakan Analisis faktor (Johnson & Wichern 2007) dengan menggunakan software IBM SPSS Statistic 20.0.0. Komponen habitat dibagi menjadi 2 kelompok, yaitu komponen biotik (23 variabel) dan komponen abiotik (11 variabel). Tahapan dalam analisis faktor dijelaskan oleh Santoso (2002) pada masing-masing komponen biotik dan abiotiknya meliputi : 1. Tranformasi angular square root seluruh data yang memiliki satuan persen (%)
ke dalam nilai sebenarnya dengan rumus: ArcSin((x)1/2).
12
3. Uji normalitas data seluruh variabel, variabel dianggap memiliki sebaran data normal apabila nilai sig. pada hasil uji > 0.05.
4. Uji autokorelasi dengan pengujian terhadap nilai uji r-Pearsonnya < 0.8. Jika terdapat autokorelasi antar komponen habitat yang diukur, maka dipilih satu komponen habitat yang berpengaruh langsung terhadap kebutuhan hidup M. javanica berdasarkan landasan teoritis yang ada.
5. Uji multikolinearitas data dengan uji VIF (Variance Inflation Factor) dengan ketentuan variabel dianggap tidak ada multikolinearitas diantaranya jika 0.1 < VIF < 10.
6. Uji nilai KMO (Kaiser-Meyer-Olkin Measure of Sampling Adequacy) dan Bartlett-test sebagai prasyarat dilakukannya analisis komponen utama ̶ analisis komponen utama dapat dilakukan jika nilai KMO ≥ 0.5.
Tabel 3 Karakteristik habitat yang digunakan M. javanica sesuai dengan kebutuhan hidupnya
Komponen Habitata Kelas Kategori Komponen Habitat
Kode Satuan I II III IV V
a Kse = kerapatan semai, Kpa = kerapatan pancang, Kti = kerapatan tiang, Kpo = kerapatan pohon,
Jjsp = Jumlah jenis tumbuhan sebagai sarang prey pada tingkat tiang dan pohon, Ksp = Kerapatan tumbuhan sebagai sarang prey pada tingkat tiang dan pohon, Jkt = jumlah koloni pakan total, Jks = Jumlah koloni pakan ordo Hymnoptera, Jkr = jumlah koloni pakan ordo Isoptera, Jpr = jarak rata-rata lokasi pakan terhadap sarang M. javanica, Jpt = jarak lokasi pakan terdekat terhadap sarang M. javanica, Jjk = jumlah jenis kompetitor, Jtk = jarak sarang M. javanica terhadap kompetitor, Jtp = Jarak sarang M. javanica terhadap predator, Jam = jarak sarang M. javanica
terhadap pusat aktivitas manusia, Shu = suhu rata-rata, Klr = kelembaban relatif, Ktm = ketinggian tempat, Ker = kelerengan lahan. b Kelas I = Sangat Jarang (Sangat Rendah), Kelas II
13 7. Reduksi faktor, variabel dikatakan komponen utama yang berpengaruh terhadap kehadiran M. javanica jika nilai Bartlett-test < 0.05 atau Measure Sampling Adequacy (MSA) ≤ 0.5, jika terdapat beberapa variabel bebas yang memiliki
nilai MSA < 0.5 maka variabel yang memiliki nilai MSA terkecil harus direduksi dan analisis komponen utama diulang hingga seluruh variabel tersisa tidak ada yang memiliki nilai MSA < 0.5.
Komponen habitat yang berpengaruh terhadap kehadiran M. javanica adalah komponen yang memiliki nilai sesuai dengan persyaratan di atas. Adapun faktor yang paling berpengaruh ditunjukkan dengan nilai komunikalitas yang menjelaskan besaran persentase variabel yang mampu menjelaskan faktor yang terbentuk. Seleksi Habitat
Metode analisis yang digunakan untuk menguji sifat selektif M. javanica adalah metode ordinal dalam Analisis korespondensi kanonik / CCA (Rencher 2002) dengan softwere Canoco 4.56. Asumsi yang digunakan dalam analisis CCA adalah (Rencher 2002): (1) adanya sifat linier antara variabel dependen dan variabel independen; (2) data yang diukur harus memiliki sebaran normal dan (3) tidak ada multikolinearitas diantara variabel ukurnya baik dependen maupun variabel independen. Oleh karena itu variabel yang digunakan adalah hasil variabel sisa berdasarkan hasil analisis PCA. Berdasarkan variabel tersebut, maka bentuk umum fungsi kanoniknya adalah:
+ + + = + + + ⋯ + �
Adapun keterangan notasi di atas adalah: y1= keberadaan habitat pakan, y2=
keberadaan habitat istirahat, y3= keberadaan habitat sosial, y4= keberadaan habitat
yang menunjang perilaku reproduksi dan mengasuh anak dan x1,2,…,n= faktor
lingkungan yang berpengaruh terhadap kehadiran M. javanica.
Hasil analisis CCA dilakukan dengan menggunakan diagram ordinasi (Lepš & Šmilauer 2003). Sifat selektif M. javanica pada masing-masing fungsi habitat ditunjukkan jika axis fungsi habitat berdekatan dengan salah satu atau lebih komponen habitat yang diukur dan menjauhi titik pusat. Penilaian terhadap dominansi variabel komponen habitat ditunjukkan dengan pendek dan panjangnya panah. Semakin pendek panah komponen-x, maka komponen tersebut merupakan variabel yang tidak dominan, dan berlaku sebaliknya. Validasi hasil analisis dilakukan dengan membandingkan hasil penghitungan oleh 2 set komputer yang
berbeda (Lepš & Šmilauer 2003). Jika perbedaan hasil kedua sampel tidak besar, bisa dikatakan hasil analisis korelasi kanonikal adalah valid (Lepš & Šmilauer
2003).
Model Kehadiran M. javanica
Variabel pembentuk model kehadiran M. javanica berasal dari pengelompokan faktor lingkungan yang berpengaruh terhadap kehadiran M. javanica hasil analisis PCA. Banyaknya model yang terbentuk bergantung pada banyaknya PC yang terbentuk. Model kehadiran M. javanica yang dibentuk adalah regresi linier berganda menggunakan IBM SPSS Statistic 20.0.0 dan divalidasi dengan Minitab 16.2.1 dengan rumus umum:
y = β0 + β1 F1+ …..+ βn Fn+εn ….(1)
14
Keterangan dari notasi di atas adalah: y merupakan jumlah individu M. javanica, β0 adalah konstanta korelasi, βn adalah koefisien pengaruh regresi pada F ke-n, dan εn merupakan bias contoh. Sedangkan τn merupakan besaran koefisien dimensi penyusun faktor ke- n penyusun Fn. Sehingga penggabungan dari kedua
rumus tersebut menjadi model regresi sebagai berikut.
y = β0 + β1 (τ1 x1+ τ1 x2 +….+ εn)+…+ βn Fn+εn …
Model regresi yang dipilih adalah model regresi terbaik dari PC yang terbentuk. Model terbaik dapat dilihat dari nilai koefisien korelasi (r), koefisien determinasi (R2), dan nilai signifikansi pada uji ANOVA. Hasil regresi yang terpilih
kemudian dianalisis lebih lanjut dengan uji t-parsial untuk mengetahui kelayakan dan signifikansi model dan masing-masing variabel dalam menduga kehadiran M. javanica.
3
HASIL DAN PEMBAHASAN
Hasil Karakteristik Habitat M. javanica
Sebanyak 15 lokasi habitat M. javanica yang teridentifikasi selama 78 hari efektif (20 Maret - 7 Juni 2014) dengan jumlah temuan sebanyak 18 individu. Kondisi ini menunjukkan bahwa efektivitas perjumpaan dengan satwa sangat rendah (19.23%). Hasil penelitian juga menunjukkan bahwa M. javanica cenderung bersifat soliter (83.33%). Berdasarkan cara hidupnya, M. javanica diketahui merupakan satwa yang cenderung bersifat terestrial meskipun jenis ini juga dapat bersifat arboreal atau fossorial. Terdapat dua pola berjalan yang teridentifkasi pada M. javanica, yaitu pola quadrupedal dan pola bipedal (Gambar 4). Cara berjalan yang unik teridentifikasi pada saat M. javanica bersifat arboreal, yakni dengan cara menjepit bidang pijaknya dengan ruas pertama jari-jarinya. Berbeda ketika turun, M. javanica akan memanfaatkan ekor prehensilnya dengan melilitkan ekornya ke bidang pijakannya (Gambar 5). Hal ini menunjukkan bahwa M. javanica hanya dapat memanjat jenis pohon yang ditumbuhi epifit atau liana. Adapun jenis-jenis liana dan epifit tersebut adalah : areuy canar (Smilax macrocarpa), areuy kecemang
a. Pola bipedal b. Pola quadrupedal
15
(Ilex triflora), areuy korong anjing (Acalypha hispida), areuy konyal (Ficus recurvua), areuy leuksa (Ficus septica), areuy meong tanan (Dracaena angustifolia), lolo (Anadendrum microstachyum), kadaka (Asplenium nidus), paku kadal (Cyclosorus aridus), paku sarengseng (Pandanus caricosus), ramo giling (Schefflera aromatica) dan rotan seel (Daemonorops melanochaetes). Pola semacam ini berhubungan dengan strategi dalam pengalihan beban tubuh untuk menekan kecelakaan ketika berjalan di atas pohon.
Cetakan jejak yang terbentuk umumnya hanya berupa ujung kuku dan telapak kakinya. Tidak jarang pada beberapa lokasi seperti di dedaunan dan batang pohon hanya goresan ujung kuku yang dapat teridentifikasi. Hal ini disebabkan oleh tertutupnya ruas buku kaki M. javanica ketika berjalan sehingga ruas buku kaki tersebut tidak tercetak (Gambar 6). Penanda ini dapat diketahui dengan mudah terutama pada musim penghujan. Hal ini karena adanya aliran permukaan (run off) air hujan yang memiliki fungsi menghapus jejak pada hari sebelumnya sehingga jejak baru dengan mudah dapat dikenali dan pada saat yang bersamaan aliran permukaan akan membuat bidang pijakan lebih gembur akibat besarnya kadar air dalam tanah sehingga cetakan kaki mudah terbentuk.
Gambar 5 Pola berjalan M. javanica ketika bersifat arboreal.
16
Penciri kedua adalah jejak ekor yang merupakan penguat dari penciri pertama. Ekor M. javanica akan bergesekan dengan bidang pijaknya ketika berjalan. Hal ini mengakibatkan tumbuhan bawah yang dilewatinya membentuk arah rebah yang seragam. Pada serasah atau tanah, jejak yang ditinggalkan berupa pola garis searah. Kombinasi dua penanda inilah yang membedakan jenis ini dengan jenis lain yang memiliki cara berjalan sama seperti: landak (Hystrix sp.) dan berang-berang (Aonyx sp.) (Gambar 7). Penanda khas oleh M. javanica juga dapat diketahui dari penciri ketiga, yaitu bekas lokasi makan yang yang umumnya diketahui dengan adanya lubang-lubang bekas cakaran M. javanica pada sarang pakan. Ketiga penanda di atas digunakan untuk menemukan keberadaan sarang aktif M. javanica.
Terdapat 4 tipe sarang M. javanica (Gambar 8), yaitu: (1) tanah (23.08%), (2) ceruk batu (15.38%), (3) pohon (53.84%) dan (4) serasah tumbuhan (7.69%). Pemilihan substrat pohon yang lebih disukai oleh M. javanica disebabkan oleh investasi energi yang sedikit karena M. javanica tidak perlu menggali atau membuat sarangnya, sehingga energi yang dikeluarkan lebih sedikit. Lokasi yang digunakan sebagai sarang M. javanica dapat diidentifikasi dengan mudah dengan ciri-ciri: (1) adanya gundukan tanah yang cenderung menutupi lubang sarangnya khususnya pada tipe sarang tanah dan ceruk batu serta (2) keberadaan jejak kaki yang membentuk pola siklik di sekitar sarangnya. Hal ini diduga terkait dengan pengaturan suhu di dalam sarangnya dan strategi toritorial pada jenis ini. Adapun sarang M. javanica yang ditemukan memiliki ukuran rata-rata lebar sebesar 24.37 ± 11.44 cm (minimal= 11.10 cm, maksimal= 56.10 cm), tinggi sebesar 27.71 ± 10.80 cm (minimal= 10.96 cm, maksimal = 54.80 cm) dan panjang atau kedalaman sarang sebesar 89.62 ± 80.04 cm (minimal= 14.90 cm, maksimal= 362.90 cm).
Berdasarkan ketinggian tempatnya, lokasi kehadiran M. javanica di TNGHS termasuk dalam tipe hutan dataran rendah perbukitan dan tipe pegunungan bawah dengan lokasi habitat berkisar dari 895 m dpl hingga 1,170 m dpl. Sebanyak 145 jenis tumbuhan yang teridentifikasi pada areal yang dimanfaatkan oleh M. javanica sebagai habitatnya (Lampiran 1). Jika dilihat dari struktur tegakan hutannya, M. javanica cenderung menggunakan hutan sekunder tua sebagai habitatnya dengan tegakan dominan berupa manii (Maesopsis eminii), puspa (Schima wallichii), ki endog (Xanthophyllum excelsum), ki jebug (Kibessia azurea) dan rasamala (Altingia excelsa). Hasil desk study menunjukkan bahwa sebanyak 12 jenis
17
tumbuhan pada tingkat tiang dan pohon yang digunakan oleh mangsa M. javanica sebagai sarangnya, yaitu : cecengkehan (Urophyllum arboreum), hamerang (Ficus padana), jaranak (Castanopsis acuminatissus), ki ara (Ficus calophylla), ki endog (Xanthophyllum excelsum), ki ronyok (Alchornea rugosa), mara bodas (Macaranga triloba), pasang bataruwa (Lithocarpus indutus), pasang kayang (Lithocarpus daphnoideus), puspa (Schima wallichii), rasamala (Altingia excelsa) dan saninten (Castanopsis javanica).
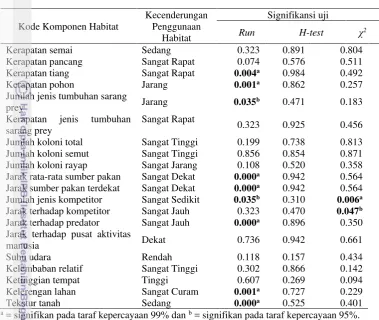
Hasil uji run menunjukkan bahwa M. javanica menggunakan habitat dengan karakteristik khusus (Tabel 4). Terdapat 9 komponen habitat yang memiliki karakteristik khusus (sig. = < 0.05) yang dikelompokkan menjadi 6 ciri utama, yaitu: (1) kerapatan tutupan tajuk atas dengan kategori tinggi yang dicirikan dengan nilai kerapatan yang sangat tinggi pada tumbuhan tingkat tiang (498.67±150.14 individu / ha) dan jarang pada tumbuhan tingkat pohon (160.33±113.25 individu / ha), (2) jumlah jenis tumbuhan yang digunakan sebagai sarang pakan dengan kategori cenderung jarang (5.13 ± 1.41 jenis / ha), (3) keberadaan pakan yang sangat dekat di sekitar sarang M. javanica (118.66 ± 156.46 m), (4) kecenderungan habitat tidak terdapat kompetitor (426.86 ± 366.25 m) maupun predator M. javanica (2,492.90 ± 1,484.24 m), (5) kelerengan lahan sangat curam (72.23 ± 17.93%) dan (6) tekstur tanah pada lokasi yang cenderung dikategorikan sedang (tekstur lempung). Kondisi di atas menunjukkan bahwa M. javanica memiliki strategi habitat yang cenderung kompleks dengan memperhatikan aspek kelimpahan sumberdaya pakan dan strategi keamanan dari kompetisi maupun predasi. Upaya penghindaran diri dari predatornya dilakukan secara langsung dengan memilih
Sarang di tanah Sarang di ceruk batu
18
habitat yang tidak dimanfaatkan oleh predatornya maupun tidak langsung dengan memilih kondisi abiotik habitat yang mampu mendukung upaya strategi anti predatornya.
Uji Krusal-Wallis (H test) menunjukkan bahwa nilai signifikansi terendah adalah komponen suhu dengan nilai sig. = 0.157. Uji tersebut bermakna bahwa tidak ada komponen habitat dengan karakteristik tertentu yang menunjukkan kekhasan penggunaan habitat oleh M. javanica. Kondisi ini berarti bahwa M. javanica mampu memanfaatkan seluruh komponen habitat yang sesuai secara maksimal untuk berbagai fungsi. Hal ini juga menjelaskan bahwa M. javanica memiliki pola seleksi habitat yang ketat karena seluruh lokasi yang digunakan untuk berbagai fungsi harus memenuhi kriteria komponen habitat yang dipersyaratkan.
Hasil uji Chi-square (χ2) menunjukkan bahwa keberadaan pesaing dalam hal pemanfaatan pakan atau sarang merupakan faktor yang mempengaruhi penggunan satu lokasi oleh M. javanica untuk berbagai fungsi habitat (jumlah jenis kompetitr sig. = 0.006 dan jarak sarang M. javanica terhadap keberadaan kompetitor sig.= 0.047). Hal ini bermakna bahwa apabila terdapat kompetitor di suatu lokasi, maka M. javanica cenderung tidak akan menggunakan lokasi tersebut sebagai habitatnya meskipun lokasi tersebut memiliki karakteristik habitat yang sesuai dengan kebutuhan M. javanica.
Kerapatan pancang Sangat Rapat 0.074 0.576 0.511
Kerapatan tiang Sangat Rapat 0.004a 0.984 0.492
Kerapatan pohon Jarang 0.001a 0.862 0.257
Jumlah jenis tumbuhan sarang
prey Jarang 0.035b 0.471 0.183
Kerapatan jenis tumbuhan sarang prey
Sangat Rapat 0.323 0.925 0.456
Jumlah koloni total Sangat Tinggi 0.199 0.738 0.813
Jumlah koloni semut Sangat Tinggi 0.856 0.854 0.871
Jumlah koloni rayap Sangat Jarang 0.108 0.520 0.358
Jarak rata-rata sumber pakan Sangat Dekat 0.000a 0.942 0.564
Jarak sumber pakan terdekat Sangat Dekat 0.000a 0.942 0.564
Jumlah jenis kompetitor Sangat Sedikit 0.035b 0.310 0.006a
Jarak terhadap kompetitor Sangat Jauh 0.323 0.470 0.047b
Jarak terhadap predator Sangat Jauh 0.000a 0.896 0.350
Jarak terhadap pusat aktivitas
manusia Dekat 0.736 0.942 0.661
Suhu udara Rendah 0.118 0.157 0.434
Kelembaban relatif Sangat Tinggi 0.302 0.866 0.142
Ketinggian tempat Tinggi 0.607 0.269 0.094
Kelerengan lahan Sangat Curam 0.001a 0.727 0.229
Tekstur tanah Sedang 0.000a 0.525 0.401
19 Komponen Habitat yang Mempengaruhi Kehadiran M. javanica
Terdapat 8 komponen utama habitat yang diduga berpengaruh terhadap kehadiran M. javanica pada suatu lokasi yang terdiri dari 3 komponen biotik dan 5 komponen abiotik (Tabel 5). Komponen biotik terdiri dari jumlah jenis tumbuhan yang berpotensi sebagai sarang oleh pakan M. javanica (Jjsp), dominansi tumbuhan yang digunakan sebagai sarang oleh mangsa M. javanica pada tingkat pohon (Dtpo) dan jarak terdekat antara habitat M. javanica terhadap kompetitor yang teridentifikasi (Jtk). Adapun komponen abiotik yang mempengaruhi kehadiran M. javanica pada suatu habitat adalah : suhu udara (Shu), kelembaban relatif (Klr), ketinggian tempat (Ktm), rataan besaran fraksi debu pada tanah (Ftd), dan rataan kelerengan lahan (Ker). Nilai KMO dan Barlett-test pada komponen biotik secara berturut-turut adalah 0.689 dan 0.031 sedangkan pada komponen abiotik adalah 0.031 dan 0.183. Besaran nilai Barlett-test pada komponen abiotik (sig. 0.183 > 0.05) yang tidak sesuai dengan asumsi analisis faktor dapat diabaikan karena variabel komponen habitat yang terbentuk sudah memenuhi kaidah uji MSA (Measure Sampling Adequacy) > 0.5 Hal ini bermakna bahwa variabel yang terpilih di atas telah terbukti berpengaruh signifikan terhadap kehadiran M. javanica tanpa harus melihat nilai Barlett-test nya.
Berdasarkan hasil uji komunikalitas diperoleh nilai terkecil sebesar 0.637 pada variabel Jtk. Hal ini berarti sebanyak 63.7% dari variabel Jtk mampu menjelaskan kehadiran M. javanica di suatu lokasi. Besaran persentase tersebut dapat diterima karena faktor tersebut mampu menjelaskan > 50 % variasi data. Lebih lanjut Tabel 5 menunjukkan bahwa 3 komponen habitat yang paling berpengaruh terhadap kehadiran M. javanica pada suatu lokasi berturut-turut adalah kelerengan lahan (82.20%), suhu udara (73.60%) dan jumlah jenis tumbuhan yang digunakan sebagai sarang pakan M. javanica (70.70%) yang ditunjukkan berdasarkan nilai uji komunikalitas tertinggi diantara variabel komponen habitat yang terpilih. Hal ini menunjukkan adanya strategi efektivitas dan efisiensi energi oleh satwa ini yang dibuktikan dengan pemilihan lokasi yang memiliki kekayaan pakan dan mampu memberikan keamanan terhadap predator serta pada saat bersamaan lokasi tersebut mampu memberikan kondisi optimal bagi berlangsungnya pola termoregulasi.
Tabel 5 Hasil uji analisis faktor pada komponen biotik dan abiotik habitat yang digunakan oleh M. javanica
20
Seleksi habitat M. javanica
Hasil analisis menunjukkan bahwa pada masing-masing sumbu memberikan pengaruh yang cenderung lemah terhadap model yang terbentuk (eigenvalues < 0.5) meskipun secara keseluruhan variasi fungsi habitat dapat dijelaskan oleh fungsi komponen habitatnya dengan nilai akar ciri (eigenvalues) sebesar 0.549 dan nilai total inertia sebesar 0.812 (Tabel 6). Kondisi ini dapat diartikan bahwa M. javanica cenderung selektif dalam memanfaatkan suatu habitat berdasarkan komponen habitatnya. Berdasarkan nilai korelasi antara fungsi habitat dan komponen lingkungannya, sumbu 1 memiliki nilai korelasi tertinggi sebesar 95.5% dan mampu menjelaskan sebanyak 62.80% dari total variasi data. Berdasarkan Tabel 6, dapat disimpulkan bahwa gambar ordinasi terbaik yang terbentuk adalah kombinasi dari sumbu 1 dan sumbu 2 dengan nilai akar ciri total sebesar 0.469 dan kemampuan menjelaskan variasi data sebesar 85.40%. Berbeda dengan 2 kombinasi lainnya yang cenderung lebih lemah, yakni (1) kombinasi sumbu 1 dan 3 yang memiliki nilai akar ciri 0.425 dengan kemampuan menjelaskan data sebesar 77.4% dan (2) kombinasi sumbu 2 dan 3 yang hanya memiliki akar ciri sebesar 0.204 dengan kemampuan menjelaskan data hanya sebesar 37.2%.
Gambar 9 menjelaskan bentuk model seleksi habitat terbaik oleh M. javanica berdasarkan nilai terbesar pada akar ciri dan kemampuan penjelasan variasi datanya. Berdasarkan gambar ordinasi tersebut, habitat makan M. javanica dipengaruhi oleh jumlah jenis tumbuhan yang digunakan sebagai sarang mangsa M. javanica. Kondisi ini menunjukkan adanya M. javanica diduga membutuhkan makanan dengan jenis yang bervariasi. Kondisi berbeda ditunjukkan oleh pemilihan habitat istirahat oleh M. javanica yang setidaknya dipengaruhi oleh 3 hal, yakni kondisi tanah sarangnya, lokasi sarang terhadap keberadaan kompetitornya dan faktor ketinggian tempat lokasi sarang tersebut. Pemilihan habitat reproduksi dan mengasuh anak lebih ditentukan oleh suhu udara. Berbeda dengan pemilihan habitat sosial yang menunjukkan bahwa tidak terdapat sifat selektif terhadap komponen habitat yang diukur dalam penelitian ini. Hal ini berarti dalam menjalankan fungsi sosialnya, M. javanica tidak dibatasi oleh komponen habitat tertentu.
21
yang dibuktikan dengan komponen fraksi debu yang memiliki panjang garis terbesar. Kondisi ini menunjukkan bahwa M. javanica hanya memilih areal-areal yang memiliki fraksi debu yang rendah atau bertekstur tanah cenderung liat atau lempung (tekstur sedang) sesuai dengan karakteristik habitat yang teridentifikasi sebelumnya. Adapun komponen habitat lain yang juga dipilih secara selektif oleh M. javanica adalah dominansi tumbuhan yang berpotensi sebagai sarang mangsa pada tingkat pohon (Dtpo) dan Suhu udara (Shu). Kondisi ini menunjukkan pengaruh komponen pakan dan abiotik lingkungan yang kuat dalam terhadap sifat selektif M. javanica. Komponen kelerengan lahan dan kelembaban relatif lingkungannya yang cenderung seragam diketahui tidak selektif dilakukan oleh M. javanica. Hal ini bermakna bahwa komponen tersebut cenderung umum diperhatikan oleh M. javanica untuk memilih lokasi sebagai habitatnya.
Gambar 10 menunjukkan bentuk model seleksi habitat lain yang dilakukan oleh M. javanica berdasarkan sumbu 1 dan sumbu 3 yang tidak dapat dijelaskan oleh model sebelumnya. Berdasarkan model ini, M. javanica mampu memanfaatkan berbagai lokasi sebagai habitat makan dan sosialnya tanpa prasyarat khusus. Berbeda dengan habitat istirahat yang dipengaruhi oleh karakteristik ketinggian tempat, suhu dan keberadaan kompetitornya yang sepenuhnya menunjukkan adanya kebutuhan habitat yang mendukung pola termoregulasinya dan perilaku menghindar dari kompetisi. Komponen fraksi tanah yang lebih diperhatikan oleh M. javanica ketika hendak memanfaatkan suatu habitat dalam proses reproduksi dan mengasuh anaknya menunjukkan adanya kebutuhan M. javanica terhadap lokasi yang mampu menjaga kondisi habitat mikro agar tetap pada kisaran normal sebagai pilihan sarangnya.
Secara umum, sifat selektif tertinggi pada model ini ditunjukkan pada komponen habitat kelembaban relatif lingkungannya dan sifat selektif terendah ditunjukkan oleh besaran dominansi tumbuhan yang berpotensi sebagai sarang mangsa pada tingkat pohon dan nilai kelerengan lahannya. Hal ini menunjukkan bahwa karakteristik kelimpahan pakan dan kemiringan lahan merupakan komponen yang wajib diperhatikan oleh M. javanica pada setiap lokasi yang dimanfaatkan Gambar 9 Diagram ordinasi seleksi habitat dan komponen habitat M. javanica
22
sebagai habitatnya meskipun bukan merupakan prasayarat utama dalam pemilihan lokasi untuk fungsi habitat tertentu. Berdasarkan ke dua model tersebut dapat ditarik beberapa kesimpulan, yakni : (1) sifat selektif terhadap habitat makan cenderung dipengaruhi oleh adanya kelimpahan pakannya; (2) Habitat istirahat dan reproduksi serta mengasuh anak yang lebih dipengaruhi oleh kondisi habitat mikro yang tetap pada kisaran normal dan mampu menjamin pola termorgulasi dengan baik; (3) Faktor lain yang diperhatikan dalam pemilihan habitat istirahat adalah kecenderungan satwa untuk menghindari diri dari kompetisi terutama dalam memperebutkan sarang dan pakan; (4) M. javanica mampu memilih habitat sosialnya secara bebas tanpa harus membutuhkan karakteristik habitat yang khusus. Model Kehadiran M. javanica
Kehadiran M. javanica dipengaruhi oleh kedua komponen habitatnya baik faktor biotik maupun faktor abiotik. Hasil analisis reduksi faktor menunjukkan bahwa terdapat tiga pengelompokan variabel baru (F) sebagai pembangun model kehadiran M. javanica. Dari ketiga komponen baru yang terbentuk, 2 diantaranya merupakan komponen abiotik. Hasil akar ciri menunjukkan bahwa pada komponen biotik, faktor baru yang terbentuk mampu menjelaskan sebesar F1 = 67.27 % dari
data sedangkan pada komponen abiotik mampu menjelaskan sebesar 70.03 % dengan nilai akar ciri masing-masing faktor baru secara berturut-turut adalah F2=
43.90% dan F3=21.93%. Kondisi ini menunjukkan bahwa secara umum, model
dengan variabel bebas komponen habitat abiotik lebih mampu merepresentasikan variasi data jika dibandingkan dengan penggunaan komponen habitat biotiknya sebagai variabel bebas.
Berdasarkan faktor baru yang terbentuk, maka dapat diketahui bahwa komponen penyusun pada masing-masing faktor baru yang terbentuk termasuk valid dikarenakan nilai vektor ciri > 0.5 (Tabel 7). Jika didasarkan pada koefisien dimensi penyusun faktor, maka dapat disimpulkan bahwa komponen rataan kelerengan lahan (Ker) merupakan komponen yang memiliki korelasi terkuat terhadap faktor baru yang terbentuk (0.766) sedangkan komponen suhu udara (Shu)
23
merupakan komponen yang memiliki korelasi terlemah terhadap faktor baru yang terbentuk dengan nilai |-0.364|.
Berdasarkan faktor baru yang terbentuk, maka dapat diketahui bahwa komponen biotik memiliki pengaruh yang lebih besar (sig. = 0.076) terhadap kehadiran M. javanica jika dibandingkan dengan komponen abiotiknya (sig. = 0.237) meskipun koefisien korelasi yang terbentuk cenderung sama (rbiotik = 0.472;
rabiotik = 0.462) (Tabel 8). Hal ini bermakna pada taraf 90%, dipercaya bahwa
kehadiran M. javanica dikarenakan nilai jumlah jenis tumbuhan yang digunakan sebagai sarang mangsa M. javanica, ukuran dominansi tumbuhan sarang mangsa pada tingkat pohon dan jarak terhadap kompetitor. Hal ini menunjukkan bahwa model terbaik yang dapat digunakan untuk menduga kehadiran M. javanica adalah y = 1.467 – 0.224 * [(0.417*ZJjsp) + (0.407*ZDtpo) + (0.396)*ZJtk)] atau jika dijabarkan akan menjadi y= 1.467 - 0.093 *ZJjsp - 0.091*ZDtpo - 0.089*ZJtk. Kondisi ini bermakna bahwa jumlah individu M. javanica yang hadir pada suatu lokasi cenderung berkorelasi negatif dengan ketiga faktor tersebut.
Hasil analisis terhadap uji t faktor abiotik menunjukkan bahwa komponen suhu udara, ketinggian tempat dan tekstur tanah (dalam hal ini diwakili oleh besaran fraksi debu) lebih berpengaruh terhadap peluang kehadiran M. javanica jika dibandingkan dengan kelerengan lahan dan kelembaban lingkungan. Lebih lanjut berdasarkan model tersebut, diperoleh fakta bahwa secara statistik kedua model yang terbentuk belum secara baik menunjukkan adanya pengaruh yang nyata terhadap kehadiran M. javanica di suatu lokasi. Hasil ini juga dikonfirmasi berdasarkan nilai uji residual (Tabel 9) yang terstandar data yang secara umum
Tabel 8 Hasil analisis regresi terhadap ketiga faktor baru yang terbentuk untuk menduga kehadiran M. javanica
24
menunjukkan nilai yang menjauhi 0. Hal ini bermakna bahwa model yang terbentuk baik dari komponen biotik maupun abiotik habitat sama-sama belum mampu memprediksi kehadiran M. javanica pada suatu lokasi dengan baik.
Pembahasan Pengaruh Cuaca
Hasil penelitian ini mampu menunjukkan bahwa penggunaan metode eksploratif dengan teknik open grid dapat digunakan dalam mengidentifikasi kehadiran mamalia kecil pada suatu habitat terlebih bagi jenis-jenis yang tidak diketahui secara pasti sebaran dan preferensi habitatnya. Sama halnya dengan penelitian dengan metode non invasif lainnya, ketergantungannya terhadap cuaca merupakan hal yang paling berpengaruh pada metode ini. Pada mamalia kecil yang berambut sedikit, waktu aktif satwa umumnya dipengaruhi kondisi tutupan vegetasi dan proses metabolisme di dalam tubuhnya (Klir et al. 1990). Kondisi ini menunjukkan bahwa M. javanica akan cenderung melakukan hibernasi pada musim penghujan dikarenakan kondisi tutupan vegetasi tidak mampu menjaga suhu tubuhnya agar tetap pada kisaran normal. Perilaku serupa juga dilaporkan oleh Wang (2000) dan Wu et al. (2004d) pada M. pentadactyla yang menunjukkan bahwa adanya suhu ekstrim mengakibatkan waktu aktif satwa ini berkurang hingga 72% dan tidak lebih dari 3 jam per hari.
Kondisi tersebut menunjukan bahwa adanya hujan mengakibatkan M. javanica hanya memiliki waktu aktif yang cenderung singkat. Hal ini berimplikasi terhadap wilayah jelajah yang sempit dan tentu menyebabkan jumlah jejak dan peluang perjumpaan dengannya akan menjadi semakin kecil. M. javanica cenderung memiliki waktu aktif yang lebih lama pada musim kemarau meskipun cetakan jejak yang ditimbulkan tidak mudah untuk teridentifikasi oleh pengamat.
Tabel 9 Hasil perbandingan nilai temuan dengan nilai prediksi dari model regresi yang terbentuk
Lokasi Residual Standart residual
Biotik Abiotik Biotik Abiotik
25 Hal ini bermakna bahwa adanya musim penghujan juga memiliki keuntungan dalam upaya identifikasi kehadiran M. javanica, yakni presisi yang tinggi dalam menentukan arah pergerakan M. javanica sehingga efektivitas pengamatan dengan metode eksploratif akan lebih tinggi. Pada mamalia kecil seperti M. javanica, hasil cetakan jejak pada tanah yang ditinggalkan sangatlah kecil (< 5 cm) sehingga jejak ini mudah hilang tersapu oleh aliran permukaan akibat air hujan.
Ekologi Perilaku M. javanica
Segala bentuk pemanfaatan terhadap sumberdaya oleh suatu jenis tercermin dalam perilaku hidupnya (Hall et al. 1997). Ekspresi yang tercermin dari perilaku satwa tidak terlepas dari bentuk morfologi tubuhnya (Fenton & Bogdanowicz 2002). Hal ini kemudian memunculkan adanya berbagai dugaan strategi hidup yang dilakukan oleh satwa yang tercermin dari perilakunya. Pola berjalan quadrupedal dan bipedal pada saat M. javanica bertujuan sebagai strategi pembagian beban tubuhnya dikarenakan otot kaki bagian depan lebih kuat sedangkan beban tubuh terbesar berada di bagian pinggulnya (Chin & Tsao 2012). Tujuan serupa juga dimaksudkan ketika M. javanica bersifat arboreal, namun demikian pembagian beban tubuhnya dilakukan dengan memanfaatkan kuku kaki bagian depan dan ekornya.
Pola quadrupedal pada dasarnya merupakan perilaku berjalan yang alami oleh M. javanica. Pada pola quadrupedal, penyangga beban lebih dititikberatkan pada kaki bagian depan (Chin & Tsao 2012). Pembagian beban tubuh yang tidak seragam mengakibatkan satwa lebih cepat lelah. Hal ini berimplikasi terhadap waktu istirahat yang cenderung lebih lama dan ekuivalen dengan wilayah jelajahnya yang cenderung sempit. Akibatnya, peluang M. javanica untuk memperoleh sumberdaya seperti makanan akan berkurang. Oleh karenanya, M. javanica juga mampu berjalan dengan pola bipedal. Pola berjalan bipedal ini dibantu oleh ekornya yang berfungsi sebagai penyeimbang tubuh. Implikasi pola bipedal ini adalah M. javanica berjalan lebih lambat terutama jika digunakan untuk mencari makan (Chin & Tsao 2012) atau berjalan lebih cepat terutama pada saat menghindari diri dari predator (Rose & Gaudin 2010). Dengan demikian, baik pola quadrupedal maupun bipedal sama-sama menunjukkan adanya strategi efektivitas waktu ketika mencari makan dan mengurangi peluang kematian akibat predator. Sebagai perbandingan, adanya aktivitas berjalan dan mengawasi lingkungan ketika makan pada mamalia herbivora dapat mengurangi resiko kematian hingga 35 % jika dibandingkan dengan satwa yang hanya berkonsentrasi untuk makan (Laundrea et al. 2001).
26
mekanik (menjaga keseimbangan dan membantu pergerakan serta alat pertahanan diri), (2) fungsi perilaku (alat komunikasi) dan (3) fungsi psikologi (membantu termoregulasi, penyimpan cadangan lemak tubuh dan mengatur keseimbangan kadar air dalam tubuh). Pada M. javanica setidaknya fungsi termoregulasi dan alat pertahanan diri oleh ekor teridentifikasi dalam penelitian ini dengan dibuktikan adanya pola melingkar pada M. javanica pada saat tidur maupun ketika tertangkap oleh predatornya (Gambar 11).
Adanya efektivitas sumberdaya dan efisiensi energi juga ditunjukkan oleh M. javanica ketika mencari sarangnya. M. javanica cenderung menggunakan lubang pada pohon dikarenakan jenis ini tidak perlu menggali atau membuat sarangnya. Rudd (1966) menyatakan bahwa pemilihan habitat istirahat oleh suatu satwa harus mampu mengurangi biaya energi yang dikeluarkan dan pada saat yang bersamaan sarang tersebut dapat memberikan iklim mikro yang dapat menghangatkan tubuh satwa tersebut. Oleh karena itu, jenis pohon yang digunakan M. javanica dalam bersarang umumnya terdapat epifit atau liana yang memiliki fungsi ganda, yaitu : (1) memudahkan M. javanica untuk memanjat pohon tersebut dan (2) mampu menjaga iklim mikro di dalam lubang sarang agar tetap pada kisaran normal dengan cara menghindarkan lubang dari paparan lingkungan sekitarnya secara langsung.
Pemilihan sarang pada pohon oleh M. javanica juga ditunjang dengan adanya ciri umum pohon yang umumnya memiliki tekstur kayu keras dan mampu menjaga kelembaban di dalam sarangnya agar tetap tinggi dan pada kisaran normal. Panshin & Ramdan (2002) mengemukakan bahwa kayu memiliki sifat higroskopis. Hal ini berarti sarang pada kayu umumnya menyediakan pilihan kelembaban ruang yang lebih tinggi jika dibandingkan dengan lingkungan luar. Hal ini juga membuktikan bahwa pemilihan sarang M. javanica berkaitan dengan kebutuhannya terhadap kondisi suhu dan kelembaban relatif yang tetap pada kisaran normal. Kondisi ini sesuai dengan tujuan utama satwa beristirahat, yakni yakni mengembalikan kondisi suhu tubuh kembali normal setelah berbagai aktivitas yang dilakukannya atau lebih dikenal dengan proses termoregulasi (Hulbert & Else 2000).
Adanya kondisi ini sesuai dengan hasil penelitian M. javanica di Singapura yang juga cenderung memanfaatkan sarang pada pohon dengan diameter > 50 cm (Lim & Ng 2007). Kebutuhan terkait suhu dan kelembaban ini pula yang menunjukkan adanya pola berbeda pada M. pentadactyla di Cina dengan kecenderungan pemilihan pada sarang tanah (77.18%) (Wu et al. 2004d). Tumbuhan yang terdapat pada wilayah beriklim sub tropis cenderung memiliki