STRUKTUR KOMUNITAS CENDAWAN SAPROB BERSPORA
ASAL SERASAH MERANTI (SHOREA spp.) DI HUTAN
PENELITIAN DRAMAGA, BOGOR
ISRAWATI HARAHAP
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Struktur Komunitas Cendawan Saprob Berspora Asal Serasah Meranti (Shorea spp.) di Hutan Penelitian Dramaga, Bogor adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juni 2013
Israwati Harahap
RINGKASAN
ISRAWATI HARAHAP. Struktur Komunitas Cendawan Saprob Berspora Asal Serasah Meranti (Shorea spp.) di Hutan Penelitian Dramaga, Bogor. Dibimbing oleh GAYUH RAHAYU dan IMAN HIDAYAT.
Hutan hujan tropis Indonesia didominasi oleh Dipterocarpaceae. Salah satu spesies dari Dipterocarpaceae yang memiliki nilai ekonomi tinggi adalah meranti (Shorea spp). Indonesia merupakan daerah endemik meranti dan beberapa spesies meranti sudah mulai dibudidayakan. Di hutan alaminya, meranti memiliki peranan yang penting dalam menjaga keseimbangan ekosistem karena menghasilkan serasah yang melimpah sebagai sumber nutrisi bagi kelangsungan hidup mikroorganisme termasuk cendawan saprob. Beberapa cendawan saprob merupakan dekomposer kayu dan daun serta berperan penting dalam siklus nutrien pada areal pertanaman meranti, karena mampu mendegradasi lignoselulosa menjadi senyawa yang lebih sederhana.
Komunitas cendawan saprob pada serasah meranti di Indonesia belum pernah dilaporkan. Oleh karena itu, penelitian ini dilakukan agar informasi tentang ragam dan populasi cendawan yang mengkolonisasi serasah meranti diketahui. Penelitian ini memiliki dua tujuan utama yaitu (1) mengetahui struktur komunitas cendawan saprob pada serasah daun dan ranting meranti yang meliputi keanekaragaman, kekayaan dan kemerataan spesies cendawan; (2) mengetahui spesies-spesies cendawan saprob yang melimpah dan dominan pada serasah meranti.
Sampel berupa serasah daun dan ranting meranti dikumpulkan dari Hutan Penelitian Dramaga. Sampel diambil dari 13 titik dalam area seluas 50 m x 50 m. Sampel diletakkan dalam kantong plastik zip lock disegel dan diberi label. Sampel kemudian dibawa ke laboratorium dan diberi kertas tisu yang telah dilembapkan. Sampel diperiksa di bawah mikroskop untuk melihat adanya tubuh buah cendawan dan mengamati ciri mikroskopis. Cendawan diidentifikasi berdasarkan ciri morfologi. Spora diisolasi menggunakan teknik isolasi spora tunggal dan dipelihara sebagai biakan spora tunggal. Biakan diidentifikasi secara morfologi dan secara molekuler berdasarkan daerah ITS ribosomal DNAnya.
Sebanyak 260 sampel serasah meranti, 54.7% serasah daun dan 45.3% serasah ranting menunjukkan adanya spora atau tubuh buah cendawan. Sebanyak 29 spesies cendawan terdiri dari 7 askomiset (24.1%) dan 22 spesies cendawan anamorf yang terdiri dari 12 soelomiset (41.4%) dan 10 hifomiset (34.5%) telah diidentifikasi. Askomiset terdiri dari Annulohypoxylon purpureonitens, Diatrype chlorosarca, Didymosphaeria epidermidis, Lophiostoma sp., Lophodermium sp.,
Pemphidium sp., Valsa sp., Soelomiset terdiri dari Coniella musaiaensis,
Coryneum betulinum, Hendersoniopsis thelebola, Lasiodiplodia theobromae,
Lasmeniella guaranitica, Leptodothiorella sp., Massariothea themedae,Pestalotia guepinii, Pestalotiopsis sp., Pseudolachnea hispidula, Septoriella sp., dan Soelomiset sp. 1, sedangkan hifomiset terdiri dari Beltraniella portoricensis,
Cryptophialoidea fasciculata, Hermatomyces sphaericus, Kiliophora ubiensis,
portoricensis, Cryptophialoidea fasciculata, Kiliophora ubiensis dan Hifomiset sp. 1. Beltraniella portoricensis dan Pemphidium sp. termasuk kategori cendawan yang sangat sering ditemukan pada serasah daun meranti, sedangkan
Cryptophialoidea fasciculata dan Lasiodiplodia theobromae adalah spesies yang sering ditemukan pada serasah ranting meranti.
Komunitas cendawan bersifat spesifik substrat. Komunitas cendawan pada serasah ranting lebih beragam dibandingkan pada serasah daun. Rasio C/N pada substrat serasah diduga berkaitan erat dengan struktur komunitas cendawan tersebut.
SUMMARY
ISRAWATI HARAHAP. Community structure of sporulating fungi on decaying litter of Shorea spp at Dramaga Research Forest, Bogor. Supervised by GAYUH RAHAYU and IMAN HIDAYAT
Indonesian tropical rain forests are dominated by the Dipterocarpaceae.
One of the most valuable Dipterocarpaceae is Shorea. Indonesia is a centre of endemic of Dipterocarpaceae and this tree is now being cultivated. In natural forests, Shorea spp. has an important role in maintaining the balance of the ecosystem because they produce abundant lignocellulosic rich substrates as a source of nutrients for the survival of microorganisms including saprobic fungi. Fungi have been known as the major wood and leaf decomposer organisms and they play an important role in the nutrient cycle in Shorea spp. plantation because they degrade lignocellulosic materials into a more simple compound.
A few study on the community structure of litter degrading fungi on Shorea
spp. has been done, but none from Indonesia. With highly diversity of Shorea spp. in Indonesia, survey on the fungal diversity on Shorea spp. litter is necessary in order to enrich the information regarding fungal community inhabiting Shorea spp. litter. Therefore, this study has two main objectives, as follows: (1) unreaveling the community structure of sporulating fungi on leaf and branch litter of Shorea
that includes diversity, species richness and evenness of fungi; (2) to know the most abundant and dominant species of fungi in Shorea spp. litter.
Leaf and branch litter samples were collected in 50 m x 50 m area at Dramaga Research Forest. Samples were placed in a zip lock plastic bag, sealed and labeled. Samples were then brought to the laboratory and moistened by placing wet tissue into these plastic bags. Samples were examined under microscopes to find fruiting bodies, and the microscopic features of the fungi were observed and noted. The fungi were mainly identified morphologically and those that can be isolated were identified further using molecular approach on the bases of the sequence of their ITS region.
Of 260 samples, about 54.7% leaf litters and 45.3% branch litters were infested by the fungi and detected either in the form of spores or fruiting bodies. About 29 fungal species composed of 7 ascomycetes (24.1%) and 22 anamorphs that comprised of 12 coelomycetes (41.4%) and 10 hyphomycetes (34.5%) were identified. The ascomycetes are Annulohypoxylon purpureonitens, Diatrype chlorosarca, Didymosphaeria epidermidis, Lophiostoma sp., Lophodermium sp.,
Pemphidium sp., Valsa sp., The coelomycetes are Coniella musaiaensis,
Coryneum betulinum, Hendersoniopsis thelebola, Lasiodiplodia theobromae,
Lasmeniella guaranitica, Leptodothiorella sp., Massariothea themedae,Pestalotia guepinii, Pestalotiopsis sp., Pseudolachnea hispidula, Septoriella sp., Coelomycete sp 1., and the hyphomycetes are Beltraniella portoricensis,
Cryptophialoidea fasciculata, Hermatomyces sphaericus, Kiliophora ubiensis,
frequent species found on leaf litter, while those of branch litter were
Cryptophialoidea fasciculata and Lasiodiplodia theobromae.
The fungal community was subtrate specific. The community on decaying branch litter was more diverse than those on leaf litter. C/N ratio of the substrate were closely related to the fungal community structure.
© Hak Cipta Milik IPB, Tahun 2013
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Mikrobiologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2013
STRUKTUR KOMUNITAS CENDAWAN SAPROB BERSPORA
ASAL SERASAH MERANTI (SHOREA spp.) DI HUTAN
PENELITIAN DRAMAGA, BOGOR
Judul Tesis : Struktur Komunitas Cendawan Saprob Berspora Asal Serasah Meranti (Shorea spp.) di Hutan Penelitian Dramaga, Bogor Nama : Israwati Harahap
NIM : G351110071
Disetujui oleh Komisi Pembimbing
Dr Ir Gayuh Rahayu Ketua
Dr Iman Hidayat Anggota
Diketahui oleh
Ketua Program Studi Mikrobiologi
Prof Dr Anja Meryandini
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
Tanggal Ujian: 9 Juli 2013
PRAKATA
Alhamdulillahirobbilalamin, puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga penulis dapat menyelesaikan penelitan dengan judul Struktur Komunitas Cendawan Saprob Berspora Asal Serasah Meranti (Shorea spp.) di Hutan Penelitian Dramaga, Bogor.
Terima kasih penulis ucapkan kepada Dr Ir Gayuh Rahayu selaku ketua komisi pembimbing, Dr Iman Hidayat sebagai anggota komisi pembimbing, atas segala bimbingan, arahan, waktu, tenaga, pikiran, serta nasehat sehingga penulis dapat menyelesaikan penulisan tesis ini. Ucapan terima kasih penulis sampaikan kepada Prof Dr Mien A. Rifai sebagai penguji luar komisi dan Prof Dr Anja Meryandini sebagai ketua program studi mikrobiologi yang telah banyak memberikan masukan dan saran demi kesempurnaan tesis ini. Disamping itu, penulis mengucapkan terima kasih kepada Ibu Yeni, Ibu Mia, Alin, Reva, Mbak Nur, Mas Dian, dan seluruh staf pegawai LIPI-Cibinong yang telah banyak membantu penulis dalam penelitiannya.
Ungkapan terima kasih juga disampaikan kepada Floreta Fiska Yuliarni, Vivi Oktavianis, Ibu Nani Radiastuti, Ibu Tatik Hartanti, Kemala S. Nagur, Lismayana Hansur, Rahayu Fitriani Wangsa Putri, teman-teman di Mikrotropisian 2013, Lisdayanti, Reikha Rahmasari, Novita Karunia Sari, Uswatun Hasanah, Latifatul Hayati, Siti Komariyah, dan adik-adik di Wisma Bintang atas bantuan, doa dan motivasinya.
Ucapan terima kasih yang teristimewa penulis sampaikan kepada Ayahanda M.M Harahap dan Ibunda Erni Helmida, atas doa yang tidak pernah putus, nasehat serta motivasi yang selalu diberikan kepada penulis, serta kepada Abang Hamdani Harahap, Kakak Kiki Maryuni Harahap & Abang Fadly Herman, Rachmat Harahap, Muhammad Abduh Harahap atas doa dan motivasi yang diberikan kepada penulis.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juni 2013
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 2
2 METODE 3
Waktu dan Tempat Penelitian 3 Bahan dan Alat 3 Pengumpulan Sampel dan Deskripsi Lokasi 3 Pengamatan Cendawan Secara Langsung dan Isolasi Cendawan 4
Penyimpanan Isolat 4
Identifikasi Cendawan 4
Ekstraksi DNA, Amplifikasi PCR dan Sekuensing 5
Analisis Filogenetik 5
Analisis Data Ekologi 5
3 HASIL DAN PEMBAHASAN 7
Struktur Komunitas Cendawan Saprob Berspora Asal Serasah Meranti 7
4 SIMPULAN DAN SARAN 37
Simpulan 37 Saran 37
DAFTAR PUSTAKA 37
DAFTAR TABEL
1 Parameter lingkungan pada lokasi pengambilan sampel 7 2 Frekuensi keberadaan dari cendawan saprob berspora penghuni serasah
meranti 34
DAFTAR GAMBAR
1 Titik sampling serasah daun dan ranting meranti 3 2 Kurva spesies area dari jumlah spesies cendawan yang diperoleh
terhadap jumlah sampel yang diobservasi 8
3 Komposisi kelompok cendawan yang menginfestasi serasah daun dan
ranting meranti 8
4 Stroma Annulohypoxylon purpureonitens di permukaan ranting, askus
dan askosporanya 9
5 Koloni Annulohypoxylon purpureonitens pada media PDA 10 6 Kedudukan Annulohypoxylon purpureonitens IPBCC.13.1077 dalam
pohon filogenetik 10
7 Stroma Diatrype chlorosarca yang terbenam pada ranting, askus
dengan askosporanya 11
8 Askoma Didymosphaeria epidermidis yang terbenam pada ranting,
askus dan askosporanya 11
9 Askoma Lophiostoma sp. terbenam dalam ranting, askus, cincin apikal,
dan askospora 12
10 Askoma Lophodermium sp. di permukaan daun, askus, dan
askosporanya 13
11 Askoma Pemphidium sp. di permukaan daun, askus, parafisis dan
askospora 14
12 Stroma Valsa sp. di atas permukaaan ranting, askus dan askospora 14
13 Konidia Soelomiset sp.1 15
14 Konidia Coniella musaiaensis 15
15 Konidia Coryneum betulinum 16
16 Konidia Hendersoniopsis thelebola 17
17 Konidia Lasiodiplodia theobromae 17
18 Konidia Lasmeniella guaranitica 18
19 Konidia Leptodothiorella sp. 19
20 Konidia Massariothea themedae 20
21 Konidia Pestalotia guepinii 20
22 Konidia Pestalotiopsis sp. 21
23 Konidia Pseudolachnea hispidula dan setula 22
24 Konidia Septoriella sp. 22 25 Konidia Beltraniella portoricensis, sel konidiogen, dan seta 23 26 Koloni Beltraniella portoricensis pada media PDA 24 27 Kedudukan Beltraniella portoricensis IPBCC.13.1078 dalam pohon
filogenetik 24
30 Kedudukan Cryptophialoidea fasciculata IPBCC.13.1079 dalam pohon
filogenetik 26
31 Konidia Hermatomyces sphaericus 26
32 Konidia Hifomiset sp.1 27
33 Koloni Hifomiset sp.1 pada media PDA 27
34 Kedudukan Hifomiset sp.1 IPBCC.13.1081 dalam pohon filogenetik 27 35 Konidia Kiliophora ubiensis, sel konidiogen dan seta 28 36 Koloni Kiliophora ubiensis pada media PDA 28 37 Kedudukan Kiliophora ubiensis IPBCC.13.1080 dalam pohon
filogenetik 29
38 Konidia Minimidochium setosum dan seta 30
39 Konidia Monodisma fragilis 30
40 Konidia Nodulisporium sp., dan konidiofor 31 41 Konidia Stilbella fimetaria dan konidiofor 31 42 Konidia Virgatospora echinovibrosa, konidiofor dan papila 32 43 Distribusi kelompok cendawan berdasarkan jumlah spesies cendawan 33 44 Distribusi kelompok cendawan berdasarkan kelimpahan setiap spesies
cendawan 33
45 Keanekaragaman cendawan berdasarkan indeks Shannon-Wiener pada
tigabelas titik pengambilan sampel 35
46 Analisis koresponden yang memperlihatkan hubungan antara spesies
1 PENDAHULUAN
Latar Belakang
Meranti (Shorea spp.) merupakan anggota Dipterocarpaceae yang tumbuh dominan di hutan hujan tropis Asia Tenggara terutama di Malaysia dan Indonesia. Indonesia memiliki sekitar 143 spesies Shorea yang dapat dikelompokkan ke dalam empat nama perdagangan yaitu meranti merah, meranti kuning, meranti putih dan balau. Pohon ini tersebar di pulau Sumatera, Bangka-Belitung, Kalimantan dan beberapa lokasi di pulau Jawa yang memiliki curah hujan di atas 2000 mm per tahun (Departemen Kehutanan 2007). Kayu meranti dapat digunakan untuk berbagai keperluan seperti kayu lapis, kayu gergajian dan bahan bangunan. Dalam hutan alaminya, meranti menghasilkan serasah yang melimpah sebagai sumber nutrisi bagi kelangsungan hidup mikroorganisme saprob termasuk cendawan.
Cendawan saprob sangat besar peranannya dalam menjaga keseimbangan ekosistem. Cendawan memperoleh nutrisi dari lingkungannya dengan cara dekomposisi serasah melalui proses enzimatik (Thorn et al. 1996). Pada berbagai ekosistem, nutrisi itu juga menjadi sumber makanan bagi berbagai mahluk hidup lain yang menempati ekosistem tersebut. Pada umumnya cendawan mampu mendekomposisi selulosa dan lignin yang merupakan komponen terbesar dari serasah yang sulit didekomposisi. Selain itu, cendawan berperan dalam daur ulang nutrien dan pembentukan humus (Dwivedi & Shukla 1977; Sinha 1982; Osono et al. 2009). Proses dekomposisi dan daur ulang nutrien merupakan aktivitas dari banyak mikrob dalam suatu komunitas.
Studi mengenai struktur komunitas cendawan pendegradasi serasah Shorea
spp. tidak banyak dilaporkan. Padahal struktur komunitas cendawan saprob perlu diketahui, sebelum proses-proses yang melibatkan cendawan saprob tersebut dimanipulasi untuk kepentingan manusia. Struktur komunitas merupakan suatu konsep yang mempelajari susunan atau komposisi spesies dan kelimpahannya dalam suatu komunitas (Schowalter 1996). Ada beberapa pendekatan yang dapat digunakan untuk menggambarkan struktur komunitas yaitu keanekaragaman spesies, interaksi spesies dan organisasi fungsional (Schowalter 1996). Dalam penelitian ini, pendekatan yang digunakan adalah keanekaragaman spesiesnya.
Beberapa studi tentang struktur komunitas cendawan saprob dari daerah penyebaran Shorea spp. seperti India dan Thailand pernah dilakukan. Soni et al.
(2011) mempelajari komunitas cendawan pada Shorea robusta di India dalam kaitannya dengan perubahan musim. Beberapa jenis cendawan kosmopolit seperti
Aspergillus flavus, Aspergillus niger dan Rhizopus stolonifer terlibat dalam dekomposisi serasah sepanjang tahun, sementara Aspergillus fumigatus,
Cladosporium cladosporioides, Cladosporium oxysporum, Curvularia indica, dan
Curvularia lunata terlibat hanya dalam tiga musim. Beberapa cendawan ektomikoriza yaitu Astraeus hygrometricus, Boletus fallax, Calvatia elata,
Mycena roseus, Periconia minutissima, Russula emetica, Scleroderma bovista,
Pithomyces cortarum jarang dan hanya ditemukan dalam satu musim. Studi struktur komunitas cendawan pada serasah meranti dilaporkan oleh Osono et al. (2009) di Thailand. Mereka mengisolasi delapan puluh spesies cendawan dari serasah daun Shorea obtusa dan mengamati suksesi cendawan tersebut selama proses dekomposisi. Trichoderma asperellum dan Aspergillus sp. merupakan cendawan yang dominan ditemukan selama proses dekomposisi, sedangkan
Nigrospora sp., Cladosporium oxysporum dan Talaromyces sp. menurun selama proses dekomposisi dan satu spesies dalam Amphisphaeriaceae yang belum teridentifikasi mengalami peningkatan selama dekomposisi.
Cendawan saprob merupakan bagian dari mikroflora serasah hutan. Salah satu aspek dalam teknik silvikultur intensif yang berkaitan dengan mikroflora serasah adalah pemanfaatan mikroba asal areal pertanaman pohon hutan untuk menunjang pertumbuhan bibit meranti.
Keanekaragaman cendawan pendekomposisi serasah daun pada daerah tropis diketahui sangat tinggi (Polishook et al. 1996; Paulus et al. 2003; Santana
et al. 2005). Namun, perubahan-perubahan ekosistem dapat menyebabkan kepunahan pada spesies-spesies cendawan pendekomposisi, oleh sebab itu cendawan-cendawan ini perlu dilestarikan. Upaya pelestarian ini hanya dapat dilakukan jika setiap komponen penyusun komunitas dikenal dan diketahui baik spesies maupun perannya di alam. Struktur komunitas cendawan saprob pada serasah meranti di Indonesia belum pernah dilaporkan, padahal Indonesia kaya dengan spesies meranti, oleh sebab itu biodiversitas dari cendawan saprob sangat penting dipelajari dan diharapkan keanekaragaman yang tinggi juga memberikan peluang untuk ditemukannya spesies langka dan spesies baru.
Tujuan Penelitian
Penelitian ini bertujuan untuk:
1. Mengetahui struktur komunitas cendawan saprob berspora pada serasah daun dan ranting meranti yang meliputi keanekaragaman, kekayaan dan kemerataan spesies cendawan.
2. Mengetahui spesies cendawan saprob yang melimpah dan dominan pada serasah daun dan ranting meranti.
Manfaat Penelitian
2 METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan September 2012 sampai Februari 2013 di laboratorium Mikologi LIPI, Cibinong.
Bahan dan Alat
Bahan yang digunakan dalam penelitian ini adalah serasah daun dan ranting meranti, KOH 3%, media agar air, media Potato Dextrose Agar (PDA), Potato Dextrose Broth (PDB) dan DNA PhythopureTM Kit Extraction (GE Healthcare, UK). Alat-alat yang digunakan adalah GPS (Garmin, USA), hand lens, pH meter, plastik zip lock, dissecting stereo microscope SZX7 (Olympus), compound microscope CX41, mikropipet (Eppendorf), mesin PCR (Takara), minisentrifus (BioExpress), elektroforesis (Mupid-exu) serta alat-alat gelas.
Pengumpulan Sampel dan Deskripsi Lokasi
Serasah daun dan ranting meranti dikoleksi dari areal pertanaman meranti di hutan Penelitian Dramaga, Pusat Penelitian dan Pengembangan Hutan Konservasi dan Rehabilitasi yang terletak di Desa Situ Gede dan Desa Bubulak, Kecamatan Bogor Barat, Kabupaten Bogor, Provinsi Jawa Barat. Sampel serasah hanya diambil satu kali pada bulan September. Sampel serasah daun dan ranting meranti diambil dari lantai hutan pertanaman tegakan meranti (Shorea javanica, Shorea leprosula dan Shorea selanica) seluas ± 50 m x 50 m. Areal sampling dibagi dalam 13 titik sampling secara diagonal (Gambar 1). Jarak antara titik sampling adalah 3 m. Dari setiap titik diambil 10 sampel serasah daun dan 10 sampel serasah ranting meranti. Ukuran panjang ranting ± 10 cm.
50 m
50 m
Serasah daun dan ranting meranti dari masing-masing titik pengambilan sampel dimasukkan ke dalam kantong plastik zip lock yang berbeda dan diberi kertas tisu yang sebelumnya telah dilembabkan dengan air steril. Kantong plastik
zip lock disegel dan diberi label dengan informasi yang berisi nama sampel, titik (GPS) pengambilan sampel, kolektor dan tanggal. Sampel kemudian dibawa ke laboratorium untuk diamati.
Pada saat pengambilan sampel, pH tanah dan kadar air tanah diukur. Kadar air serasah, rasio C/N serasah juga dianalisis. Besarnya jumlah sampel dianalisis untuk mengetahui tingkat kecukupan sampel dalam menggambarkan komunitas cendawan saprob berspora. Titik asimtot adalah titik kecukupan sampel.
Pengamatan Cendawan Secara Langsung dan Isolasi Cendawan
Cendawan saprob pada serasah daun dan ranting diamati di bawah mikroskop Olympus SZX7. Serasah yang mengandung tubuh buah cendawan dipisahkan berdasarkan kelompok cendawannya. Tubuh buah askomiset dan soelomiset diambil menggunakan pisau spesimen. Jika tubuh buah sudah kering, tubuh buah direhidrasi terlebih dahulu dengan cara diberi satu atau dua tetes air steril atau kalium hidroksida (KOH) 3%. Tubuh buah cendawan yang sudah lembab diletakkan pada gelas preparat untuk dibuat preparat semi permanennya dan selanjutnya diamati dengan menggunakan mikroskop Olympus CX41. Pada hifomiset, struktur reproduksi diambil dengan pinset dan dibuat preparat semi permanen dengan air steril.
Preparat semi permanen dibuat bentuk permanennya dengan menggunakan medium shear’s. Morfologi cendawan diamati secara mikroskopis. Data pengamatan mikroskopis ini digunakan untuk identifikasi spesimen.
Selain dibuat preparat permanennya cendawan juga diisolasi. Metode isolasi yang dilakukan yaitu isolasi spora tunggal menurut Choi et al. (1999) dengan menggunakan media agar air. Biakan dimurnikan pada media Potato Dextrose Agar (PDA).
Penyimpanan Isolat
Cendawan yang berhasil diisolasi, disimpan pada PDA agar tabung yang diberi parafin. Biakan disimpan di IPB Culture Collection (IPBCC).
Identifikasi Cendawan
Identifikasi cendawan dilakukan berdasarkan ciri morfologi. Untuk konfirmasi identifikasi secara morfologi, analisis molekuler dilakukan pada semua cendawan yang berhasil diisolasi. Identifikasi morfologi dilakukan secara langsung dari tubuh buah pada serasah daun dan ranting yang sebelumnya telah dibuat preparat semi permanennya.
(1993) dan Sutton (1980). Hifomiset: Ellis (1971, 1976), Carmichael et al. (1980) dan Seifert et al. (2011). Beberapa database pada website dapat dijadikan sebagai acuan cendawan yang sudah pernah dilaporkan seperti: Index Fungorum (Http://www.indexfungorum.org/Names/Names.asp). Mycobank (Http://www. mycobank.org) dan USDA fungus host database (Http://nt.ars-grin.gov/fungaldatabases/fungushost/fungushost.cfm). Acuan lainnya adalah jurnal-jurnal publikasi terbaru yang sesuai dengan spesies cendawan yang ditemukan.
Ekstraksi DNA, Amplifikasi PCR dan Sekuensing
Kultur cendawan saprob dianalisis secara molekuler. Biomassa miselium pada media Potato Dextrose Broth (PDB) berumur 3-5 hari dijadikan sumber DNA. Ekstraksi DNA menggunakan DNA PhythopureTM Kit Extraction (GE Healthcare, UK).
DNA diamplikasi dengan primer ITS5 (5’-GGAAGTAAAAGTCG TAACAAGG -3’) dan ITS4 (5’-TCCTCCGCTTATTGATATGC-3’) (White et al. 1990) dalam mesin PCR. Setiap tabung berisi 25 μl terdiri dari 10 μl nuclease free water, 12.5 μl Go taq green mastermix TM, ITS5 & ITS4 masing-masing 0.5 μl, DMSO 0.5 μl dan DNA template 1 μl. Amplifikasi PCR dilakukan dengan kondisi PCR yaitu predanaturasi 95oC selama 90 detik, selanjutnya diikuti oleh 35 siklus, denaturasi (95°C, 30 detik), annealing (55°C, 30 detik) dan extension (72°C, 90 detik) dan final extension 72oC selama 5 menit Hasil amplifikasi kemudian dielektroforesis pada gel agarose dengan konsentrasi 1 % dan direndam dalam etidium bromida selama ± 30 menit selanjutnya divisualisasikan dibawah sinar UV (Bio-Rad UV Transilluminator 2000). Hasil PCR dikirim ke 1st Base (Malaysia) untuk disekuensing.
Analisis Filogenetik
Sekuen ITS termasuk 5.8S rDNA yang diperoleh akan dibandingkan dengan sekuen dari database the National Center for Biotechnology Information (NCBI) (www.ncbi.nlm.nih.gov) melalui program BLAST (Basic Local Alignment Search Tool). Sekuens homolog akan diambil untuk di-align dengan sekuen yang sedang dianalisa menggunakan program MEGA 5 (Tamura et al. 2011). Pohon filogenetiknya dibangun dengan metoda Neighbour joining menggunakan program MEGA 5 dengan 1000 x ulangan bootstrap.
Analisis Data Ekologi
Total jumlah spesies cendawan pada sampel serasah dicatat dan dihitung. Frekuensi keberadaan (FK) setiap spesies cendawan digunakan untuk menetapkan dominansi cendawan pada habitatnya.
Berdasarkan frekuensi keberadaan (FK) pada setiap spesies cendawan, maka cendawan dikelompokkan menjadi sangat sering ditemukan (FK >10%), sering ditemukan (5%< FK ≤10%), kadang-kadang ditemukan (1%< FK ≤5%) dan jarang ditemukan (FK ≤1%).
Keanekaragaman spesies dihitung menggunakan indeks keanekaragaman spesies (kekayaan spesies dan kemerataan spesies). Indeks keanekaragaman Shannon digunakan untuk menghitung kelimpahan spesies yang ada (H’). Indeks kekayaan spesies (DMg) digunakan untuk menghitung kekayaan spesies dalam komunitas. Indeks kemerataan digunakan untuk menghitung distribusi setiap spesies pad substrat serasah daun dan serasah ranting meranti. Kisaran kemerataan dari 0 sampai 1. Komunitas dengan nilai kemerataan sama dengan 1 artinya bahwa setiap spesies dalam komunitas memiliki frekuensi keberadaan yang sama (Magurran 1988).
Indeks Keanekaragaman Spesies
Indeks keanekaragaman spesies cendawan dihitung menggunakan indeks Shannon-Wiener (H’) dengan menggunakan rumus sebagai berikut:
H= - pi. ln pi
pi kelimpahan relatif ∑ individu dalam spesies ke i∑ individu semua spesies
Indeks Kekayaan Spesies
Indeks kekayaan spesies cendawan (DMg) dihitung menggunakan rumus sebagai berikut:
DMg = S N ;
N= Jumlah individu dalam komunitas S= Jumlah spesies dalam komunitas
Indeks Kemerataan Spesies
Indeks kemerataan spesies cendawan (E) dihitung menggunakan rumus sebagai berikut:
E= H’/ln S
H’= Nilai indeks Shannon-Wiener S = Jumlah spesies
Pada cendawan, individu menunjukkan koloni sehingga satu individu merupakan satu koloni.
3 HASIL DAN PEMBAHASAN
Struktur Komunitas Cendawan Saprob Berspora Asal Serasah Meranti
Sebanyak 260 sampel serasah meranti yang terdiri dari 130 serasah daun dan 130 serasah ranting diamati keberadaan cendawan saprob bersporanya. Dari seluruh sampel serasah, hanya 71 (54.7%) sampel serasah daun dan 59 (45.3%) sampel serasah ranting yang terinfestasi cendawan saprob. Persentase serasah daun yang diinfestasi oleh cendawan lebih tinggi daripada persentase serasah ranting. Hal ini diduga berkaitan dengan kadar air subtrat dan kadar air tanah. Serasah daun memiliki kadar air (3.98%) yang sedikit lebih tinggi dibandingkan kadar air (3.39%) serasah ranting (Tabel 1) dan kadar air kedua substrat ini relatif sangat rendah. Tumpukan daun yang lebih luas daripada ranting di permukaan tanah akan membentuk kondisi lingkungan mikro yang menunjang kehidupan cendawan di permukaan daun yang menghadap ke tanah. Kadar air dan kelembapan udara merupakan salah satu faktor yang mempengaruhi pertumbuhan cendawan terutama dalam proses perkecambahan spora, pertumbuhan miselium dan pembentukan spora (Yoder & Wood 1973). Beberapa cendawan seperti
Aspergillus flavus, Aspergillus candidus dan Aspergillus glaucus tumbuh pada subtrat dengan kadar air sekitar 9.3-11.3% (Paderes et al. 1996). Oleh sebab itu kemungkinan lain adalah cendawan saprob yang ditemukan pada serasah tersebut merupakan cendawan yang berada dalam keadaan dorman.
Tabel 1 Parameter lingkungan pada lokasi pengambilan sampel
Parameter Nilai
pH tanah 6 Intensitas cahaya 120 Kadar air tanah 38.34% Kadar air
serasah ranting 3.39% serasah daun 3.98% Rasio C/N
serasah ranting 67.12 serasah daun 55.97
Gam
Kelompok cendawan anamorf lebih dominan dibandingkan teleomorf (Gambar 3) seperti yang ditemukan oleh Osono et al. (2009) dan Soni et al. (2011). Pinruan et al. (2007) juga menemukan pertumbuhan anamorf yang lebih cepat dalam menginfestasi daun palem karena mampu menggunakan pati dan gula pada serasah daun palem tersebut.
Anggota-anggota komunitas cendawan pada serasah meranti memiliki pertelaan sebagai berikut:
1. Annulohypoxylon purpureonitens Hsieh, Ju & Rogers, Mycologia 97: 844-865 (2005) (Xylariaceae)
Annulohypoxylon purpureonitens memiliki stroma yang terdapat di permukaan serasah ranting (Gambar 4a), tubuh buah semiglobos, menyerupai bantalan berwarna hitam karbon dengan ostiol. Askus silindris dengan 8 askospora (Gambar 4b), askospora uniseriat, satu sel, berwarna coklat (Gambar 4c) dan memiliki celah kecambah. Miselium pada media PDA (Gambar 5) berwarna putih.
Sebanyak 49 spesies Annulohypoxylon telah tercatat dari berbagai wilayah di dunia (Index Fungorum 2013). Annulohypoxylon purpureonitens pertama kali dilaporkan oleh Hsieh et al. pada tahun 2005 (Index Fungorum 2013). Keberadaan Annulohypoxylon purpureonitens di Indonesia belum pernah dilaporkan sebelumnya, oleh sebab itu tulisan ini merupakan laporan yang pertama. Selain pada serasah meranti, Annulohypoxylon tersebar pada beberapa inang diantaranya Alnus glutinosa, Betula pendula, Betula pubescens, Betula pubescens subsp. carpatica, Carpinus betulus, Corylus avellana, Fagus orientalis, Fraximus excelsior, Quercus acutissima, Quercus variabilis, Salix sp. dan Sorbus aucuparia subsp. glabrata (USDA fungus host database 2006).
Gambar 4 Stroma Annulohypoxylon purpureonitens di permukaan ranting (a), askus (b) dan askosporanya (c)
a
c
b
a
b
dan Broome pada tahun 1873 (Index Fungorum 2013). Adanya Diatrype chlorosarca pada serasah meranti merupakan catatan baru bagi spesies ini di Indonesia. Diatrype chlorosarca tersebar pada beberapa inang yaitu
Archontophoenix alexandrae, Archontophoenix sp., Parashorea plicata,
Trachycarpus fortunei, dan Trachycarpus sp. (USDA fungus host database 2006).
Gambar 7 Stroma Diatrype chlorosarca yang terbenam pada ranting (a), askus (b) dengan askospora.
3. Didymosphaeria epidermidis Fuckel, Jb. nassau. Ver. Naturk. 23-24: 140 (1870) (Didymosphaeriaceae)
Didymosphaeria epidermidis memiliki askoma yang terbenam dalam serasah ranting (Gambar 8a). Askus silindris dengan 8 askospora (Gambar 8b). Askospora uniseriat, elipsoid, 2 sel dan berwarna coklat (Gambar 8c).
Gambar 8 Askoma Didymosphaeria epidermidis yang terbenam pada ranting (a), askus (b) dan askosporanya (c)
Didymosphaeria epidermidis pertama kali dilaporkan oleh Fuckel pada tahun 1870 dengan ciri-ciri yaitu memiliki askus silindris dengan 8 askospora, 2 sel, tersusun secara uniseriat. Sebanyak 511 spesies Didymosphaeria telah tercatat dari berbagai wilayah di dunia (Index Fungorum 2013). Didymosphaeria epidermidis yang menginfestasi serasah meranti belum pernah dilaporkan
a
b
c a
b
sebelumnya dan spesies ini ditemukan untuk pertama kalinya di Indonesia. Selain pada meranti, spesies ini tersebar pada beberapa inang yaitu Aster ericoides var.
platyphyllus, Berberis sp., Berberis vulgaris, Calicotome villosa, Calicotome spinosa, Dalbergia melanoxylon, Erigeron canadensis, Eucalyptus globulus,
Philadelphus caucasius, Ribes uva-crispa, Salix caprea, Salix purpurea dan beberapa inang lain (USDA fungus host database 2006).
4. Lophiostoma sp. (Lophiostomataceae)
Lophiostoma sp. memiliki askoma yang juga terbenam dalam ranting (Gambar 9a). Askoma akan berada di atas substrat setelah terjadi pelapukan jaringan epidermis inang. Askus berbentuk gada, bitunikat dengan 8 askospora (Gambar 9b) dan memiliki cincin apikal (Gambar 9c). Askospora sedikit fusiform, 2-seriat, eusepta dan hialin (Gambar 9d).
Sebanyak 441 spesies Lophiostoma telah tercatat dari berbagai wilayah di dunia (Index Fungorum 2013). Lophiostoma tersebar pada banyak inang diantaranya Abies sp., Acer campestre, Acer cinerascens, Acer negundo, Acer platanoides, Acer saccharum, Achillea millefolium var. alpicola, Aconitum napellus, Aconitium septentrionale, Aegiceras corniculatum, Agropyron sp., Alnus glutinosa, Alnus incana, Amaranthus graecizans, Amelanchier alnifolia,
Andropogon sp., Anthriscus silvestris, Arbutus unedo, Archontophoenix alexandrae, Cocos nucifera, Phragmites australis dan pada beberapa inang yang lain (USDA fungus host database 2006). Lophiostoma yang menginfestasi meranti baru pertama kali dilaporkan di Indonesia.
Gambar 9 Askoma Lophiostoma sp. terbenam dalam ranting (a), askus (b), cincin apikal (c), dan askospora (d)
a b b
5. Lophodermium sp. (Rhytismataceae)
Lophodermium sp. memiliki askoma subkutikular atau subepidermal, berbentuk elipsoid-fusiform atau elipsoid-oblong, berwarna coklat muda sampai hitam, memiliki sel seperti bibir, askoma sering mengelompok dan dikelilingi atau tidak dengan garis daerah hitam yang tipis (Gambar 10a). Askus silindris, unitunikat dengan 8 askospora (Gambar 10b). Askospora berbentuk filliform, 1-2 sel, lurus, hialin dan memiliki dinding sel yang halus (Gambar 10c).
Sebanyak 342 spesies Lophodermium telah tercatat dari berbagai wilayah di dunia (Index Fungorum 2013). Lophodermium asal Indonesia telah dilaporkan sebelumnya sebanyak 5 spesies yang tersebar pada beberapa inang yaitu Calamus
sp, Mangifera indica, Scirpus sp, dan Elettaria sp. (USDA fungus host database 2006). Lophodermium yang menginfestasi Shorea baru pertama kali dilaporkan.
Gambar 10 Askoma Lophodermium sp. di permukaan daun (a), askus (b), dan askosporanya (c)
6. Pemphidium sp.(Xyalariaceae)
Pemphidium sp. memiliki askoma yang terbenam dibawah pseudostroma, terlihat sebagai piringan berwarna hitam, mengkilap, soliter atau mengelompok (Gambar 11a). Askus silindris, unitunikat dengan 8 askospora (Gambar 11b), memiliki parafisis (Gambar 11c). Askospora silindris, 3-4 seriat, 1 sel dan hialin (Gambar 11d).
Dari 17 spesies Pemphidium telah tercatat dari berbagai wilayah di dunia (Index Fungorum 2013), tidak satupun spesies pernah dilaporkan sebelumnya di Indonesia, oleh sebab itu laporan ini merupakan yang pertama. Selain pada meranti, spesies ini tersebar pada beberapa inang yaitu Berlinia sp., Calamus
c a
australis, Calamus caryotoides, Calamus moti, Dipterocarpus sp., Jessenia bataua, Mauritia flexuosa, Mauritia sp., Maximiliana regia dan Phenakospermum guianense (USDA fungus host database 2006).
Gambar 11 Askoma Pemphidium sp. di permukaan daun (a), askus (b), parafisis (c) dan askospora (d)
7. Valsa sp. (Valsaceae )
Valsa sp. memiliki stroma erumpent yang mengandung banyak askoma. Askoma terbenam dalam substrat, berwarna hitam dengan leher ostiol panjang (Gambar 12a), mengelompok atau di dalam lingkaran dalam stroma. Askus berbentuk gada (Gambar 12b) dengan 8 askospora. Askospora alantoid, biseriat, 1-sel dan hialin (Gambar 12c).
Sebanyak 865 spesies Valsa yang telah tercatat di dunia (Index Fungorum 2013). Dua spesies Valsa yaitu Valsa eugeniae dan Valsa myrtagena, telah dilaporkan menginfestasi Eucalyptus grandis di Indonesia. Valsa yang menginfestasi Shorea merupakan catatan pertama. Selain itu, Valsa tersebar pada banyak inang diantaranya Abies alba, Abies balsamea, Acer saccharum, Betula alba, Cocos nucifera, Eucalyptus globulus, Fagus sylvatica dan pada beberapa inang lain (USDA fungus host database 2006).
Gambar 12 Stroma Valsa sp. di atas permukaaan ranting (a), askus (b) dan askospora (c)
a
b
d
c
a
8. Soelomiset sp. 1
Soelomiset sp. 1 memiliki konidia oval, tidak bersepta, hialin, dindingnya halus (Gambar 13) dan konidiofor tidak teramati.
Gambar 13 Konidia Soelomiset sp.1
9. Coniella musaiaensis Sutton (Schizoparmaceae)
Coniella musaiaensis memiliki konidia fusiform, tidak bersepta, berwarna coklat tua, dengan ukuran 7-10 μm x 3-5 μm (Gambar 14), konidiofor tidak teramati. Coniella musaiaensis pertama kali dilaporkan oleh Sutton pada tahun 1969 dengan ciri-ciri konidia fusiform berukuran 11-16 µm x 3.5-5 µm. Coniella
memiliki ciri-ciri miselium terbenam, hialin, bercabang dan bersepta. Konidioma dalam bentuk piknidia, terpisah, bulat, berwarna coklat muda, terbenam dalam substrat memiliki satu lokus dan berdinding tebal. Konidiofor tidak ada. Sel konidiogen enteroblastik, fialid. Konidia berbentuk fusiform, lurus atau falkat, berwarna kuning langsat sampai coklat, tidak bersepta, halus, dan dindingnya tipis atau tebal (Sutton 1980).
Sebanyak 26 spesies Coniella telah tercatat di dunia (Index Fungorum 2013). Coniella musaiaensis pertama kali dilaporkan ada di Indonesia. Selain pada meranti, spesies ini terdapat pada Bauhinia malabarica (USDA fungus host database 2006).
Gambar 14 Konidia Coniella musaiaensis
10. Coryneum cf. betulinum Schulzer
Coryneum ini memiliki konidia fusiform, 4-5 distoseptat, berwarna coklat muda dengan ukuran 18-23 μm x 8-10 μm (Gambar 15). Ukuran konidia ini kurang dari pertelaan yang pertama kali dilihat oleh Schulzer pada tahun 1882 berukuran 31-36 µm x 14-17 µm, lurus, 4-5 distoseptat, meskipun bentuknya sama, konidiofor tidak teramati.
Coryneum memiliki ciri-ciri miselium terbenam, bercabang, memiliki septa, dan berwarna coklat muda. Konidioma dalam bentuk aservulus, konidiofor silindris, septat, bercabang pada bagian dasar, berwarna hialin sampai coklat pucat, dibentuk dari pseudoparenkim atas. Konidia berbentuk fusiform atau globos, mengerucut pada bagian dasar, sel pada bagian ujung sering pucat (Sutton 1980).
Sebanyak 228 spesies Coryneum telah tercatat dari berbagai wilayah di dunia (Index Fungorum 2013). Catatan tentang Coryneum di Indonesia belum ada.
Coryneum penginfestasi serasah meranti juga belum pernah dilaporkan sebelumnya. Selain pada meranti, spesies ini tersebar pada beberapa inang yaitu
Betula pendula, Betula rubra, Castanopsis fissa dan Fagus sylvatica (USDA fungus host database 2006).
Gambar 15 Konidia Coryneum cf. betulinum
11. Hendersoniopsis thelebola Höhnel, Annls mycol. 16: 124 (1918)
Hendersoniopsis thelebola mempunyai konidia fusiform, bersepta, dengan ukuran 24-27 μm x 5-7 μm (Gambar 16).
Hendersoniopsis thelebola pertama kali dilaporkan oleh Höhn pada tahun 1918 dengan ciri-ciri konidioma berukuran 1200 µm x 950 µm, makrokonidiofor berukuran 40 µm x 4.5-6 µm, makrokonidia berukuran 31-53 x 10-12 µm, mikrokonidiofor berukuran 21 x 1.5-2 µm dan mikrokonidia berukuran 6.5-8 x 1.5-2 µm (Sutton 1980).
pada bagian dasar, meruncing sampai bagian ujung, dibentuk dari dinding sel pada bagian dalam lokus. Mikrokonidia berbentuk fusiform, falkat, hialin, tidak bersepta dan halus (Sutton 1980).
Gambar 16 Konidia Hendersoniopsis thelebola
Sebanyak 1 spesies Hendersoniopsis telah dilaporkan di dunia (Index Fungorum 2013). Hendersoniopsis thelebola yang ditemukan pada penelitian ini akan menambahkan informasi mengenai daerah distribusi dari Hendersoniopsis thelebola dan merupakan laporan pertama di Indonesia. Selain pada meranti, spesies ini tersebar hanya pada beberapa inang yaitu Alnus glutinosa, Alnus hirsuta, Alnus incana dan Alnus viridis (USDA fungus host database 2006).
12. Lasiodiplodia theobromae Griffon & Maubl. (Botryosphaeriaceae)
Lasiodiplodia theobromae memiliki konidia elipsoid, pada bagian ujungnya mengerucut, dengan satu septa pada bagian tengahnya, berwarna coklat, dinding selnya tebal dan berukuran17-23 μm x 8-11 μm (Gambar 17). Konidiofor tidak teramati.
Sebanyak 28 spesies Lasiodiplodia telah tercatat dari berbagai wilayah di dunia (Index Fungorum 2013). Lasiodiplodia theobromae pertama kali dilaporkan oleh Griffon dan Maubl. pada tahun 1909 (Index Fungorum 2013). Spesies ini tersebar pada banyak inang diantaranya Acacia crassicarpa, Allium sativum, Aloe vera, Areca catechu, Dipterocarpus sp., Eucalyptus urophylla, Pandanus utilis,
Zea mays dan pada banyak inang lain (USDA fungus host database 2006).
13. Lasmeniella guaranitica Petr. & Syd.
Lasmeniella guaranitica memiliki konidia bulat, tidak bersepta, berwarna coklat tua, dindingnya tebal, halus, berukuran 6-8 μm x 3-5 dan memiliki pori pada bagian tengah (Gambar 18).
Lasmeniella guaranitica pertama kali dilaporkan oleh Petr. dan Syd. pada tahun 1927 dengan ciri konidioma mencapai ukuran 1200 µm, epidermal. Sel konidiogen berukuran 13-14 µm x 1-1.5, konidia 7-9 µm, coklat muda dengan bintik jernih pada bagian tengah nya (Sutton 1980).
Spesies Lasmeniella memiliki ciri miselium terbenam, bercabang, hialin, bersepta. Konidioma eustromatik, sub epidermal, sel konidiogen holoblastik, hialin sampai coklat pucat, membengkak pada bagian dasar, silindris sampai meruncing ke atas. Konidia berwarna coklat, berdinding tebal, halus, tidak bersepta, datar, meruncing pada bagian dasar dan sering dengan pori pada bagian tengah (Sutton 1980).
Gambar 18 Konidia Lasmeniella guaranitica
Sebanyak 13 spesies Lasmeniella telah tercatat dari berbagai wilayah di dunia (Index Fungorum 2013). Lasmeniella guaranitica yang menginfestasi
14. Leptodothiorella sp. (Botryosphaeriaceae)
Leptodothiorella sp. memiliki konidia silindris dan sedikit membengkak pada bagian ujung, hialin, tidak bersepta dan berukuran 7-8 μm x 2-3 μm (Gambar 19).
Leptodotiorella memiliki ciri miselium terbenam, bercabang, bersepta, berwarna coklat sampai coklat muda. Konidioma berbentuk pikinidium, globos sampai subglobos, coklat gelap sampai hitam, terpisah atau bergabung, immersed, dindingnya tebal, dindingnya hialin sampai coklat pucat. Sel konidiogen enteroblastik, fialid. Konidia hialin, tidak bersepta, halus, dengan bentuk silindris, atau sedikit membengkak pada bagian ujung dan terdapat guttulate (Sutton 1980).
Sebanyak 16 spesies Leptodothiorella telah tercatat di dunia (Index Fungorum 2013). Leptodothiorella yang berasal dari Indonesia belum pernah dilaporkan sebelumnya. Selain pada meranti, Leptodothiorella tersebar pada beberapa inang yaitu Cedrela toona, Eurycles amboinensis, Ficus pleurocarpa,
Murraya exotica, Musa sp., Thymus pulegioides, Vitis vinifera dan Yucca aloifolia
(USDA fungus host database 2006).
Gambar 19 Konidia Leptodothiorella sp.
15. Massariothea themedae Sydow, Annls mycol. 37: 249 (1939)
Massariothea themedae memiliki konidia fusiform, 7 distoseptat dengan ukuran 80 μm x 27 μm dan berwarna coklat muda (Gambar 20).
Massariothea themedae pertama kali dilaporkan oleh Sydow pada tahun 1939 dengan ciri konidioma datar berukuran 600 µm. Konidia fusiform, berlekuk, 6-7 distosepta, berukuran 31.5-44 x 10.5-12 µm (Sutton 1980).
Spesies Massariothea memiliki ciri miselium di atas permukaan substrat atau terbenam, bercabang, bersepta dan berwarna coklat. Konidiama eustromatik, terpisah atau bergabung, berbentuk globos sampai rostrate, berwarna coklat gelap sampai hitam, terbenam sampai superfisial, memiliki satu sampai banyak lokus, berdinding tebal dengan warna coklat tua. Sel konidiogen enteroblastik, fialid, dengan bentuk doliform sampai lageniform, hialin, halus. Konidia berwarna coklat, 3-7 distoseptat, pada bagian dasar mengerucut, ujungnya tumpul, halus dan berbentuk silindris (Sutton 1980).
pernah dilaporkan sebelumnya, oleh sebab itu laporan ini merupakan laporan yang pertama. Selain pada meranti, Massariothea tersebar pada beberapa inang yaitu
Heteropogon triticeus, Phragmites communis, Sorghum plumosum dan Shorgum vulgare (USDA fungus host database 2006).
Gambar 20 Konidia Massariothea themedae
16. Pestalotia guepinii Desm. (Amphisphaeriaceae)
Pestalotia guepinii memiliki konidia fusiform, 6 sel dengan empat sel sentral berwarna coklat, sedangkan sel pada bagian ujung hialin, berukuran 23-31
μm x 8-9 μm dan memiliki setula yang simpel (Gambar 21).
Pestalotia guepinii pertama kali ditemukan oleh Desm. pada tahun 1840 dengan ciri konidioma berukuran 200 µm, konidiofor berukuran 10-15 x 1-3 µm, konidia berukuran 21-27 x 6.5-8.5 µm, halus, memiliki 2-5 setula pada pada bagian ujung konidia, sebagian besar berjumlah 3 tidak bercabang, pada bagian ujung tumpul, setula berukuran 4-12 µm (Sutton 1980).
Spesies Pestalotia memiliki ciri miselium terbenam, bercabang, septat, coklat. Konidioma eustromatik, hitam atau coklat tua, terbenam kemudian menjadi erumpent. Konidiofor hialin, bercabang secara teratur, septat, halus, dibentuk dari sel pada bagian atas pseudoparenkim, sel konidiogen holoblastik,
indeterminate, menyatu, silindris, hialin, halus. Bentuk konidia fusiform, lurus atau sedikit berliku, 6 sel dengan 4 sel sentral berwarna coklat, sel pada bagian ujung hialin dan memiliki setula yang simpel atau bercabang (Sutton 1980).
17. Pestalotiopsissp. (Amphisphaeriaceae)
Pestalotiopsis sp. memiliki konidia fusiform, 5 sel dengan sel bagian dasarnya hialin dan bagian tengahnya berwarna coklat dengan ukuran 10-11 μm x 3-5 μm, dan memiliki setula yang simpel (Gambar 22).
Spesies Pestalotiopsis memiliki ciri miselium terbenam, bercabang, septat, hialin sampai coklat muda. Konidiofor hialin, bercabang dan bersekat pada bagian dasar. Konidia fusiform, 5 sel, pada bagian dasar sel berwarna hialin, sedangkan bagian tengah berwarna coklat dan memiliki setula yang simpel atau bercabang (Sutton 1980).
Sebanyak 250 spesies Pestalotiopsis telah tercatat dari berbagai wilayah di dunia (Index Fungorum 2013). Di Indonesia, Pestalotiopsis tersebar pada banyak inang diantaranya Ficus elastica, Myrica javanica, Cocos nucifera dan Elaeis guineensis (USDA fungus host database 2006). Pestalotiopsis yang menginfestasi meranti belum pernah dilaporkan sebelumnya dan ini merupakan laporan yang pertama.
Gambar 22 Konidia Pestalotiopsis sp.
18. Pseudolachnea hispidula Sutton
Pseudolachnea hispidula memiliki konidia fusiform sampai silindris, simpel, hialin, satu septa, berukuran 90-100 μm x 17-23 μm (Gambar 23a) dan pada bagian ujung memiliki satu setula (Gambar 23b).
Pseudolachnea hispidula pertama kali dilaporkan oleh Sutton pada tahun 1977 dengan ciri konidioma berbentuk mangkok dengan ukuran 260-850 µm, seta berukuran 400 x 3-5 µm, konidiofor berukuran 20-30 x 1.5-2 µm, konidia bersepta satu pada bagian tengah, eggutulate, pada bagian ujung dan dasar memiliki satu setula, tidak bercabang berukuran 2-3 µm (Sutton 1980).
Gambar 23 Konidia Pseudolachnea hispidula (a) dan setula (b)
Sebanyak 20 spesies Pseudolachnea telah tercatat dari berbagai wilayah di dunia (Index Fungorum 2013). Pseudolachnea asal Indonesia belum pernah dilaporkan sebelumnya, oleh sebab itu laporan ini merupakan laporan yang pertama. Selain pada meranti, spesies ini tersebar pada beberapa inang diantaranya Abies sp., Calamagrostis arundinaceae, Morus alba, Phytolacca sp.,
Robinia pseudoacacia, Salix sp. dan Syringa vulgaris (USDA fungus host database 2006).
19. Septoriella sp.
Septoriella sp. mempunyai konidia silindris, 8 eusepta dengan ukuran 9-14
μm x 71-88 μm, hialin, pada bagian dasarnya mengerucut dan bagian ujungnya tumpul, memiliki dinding yang tipis dan halus (Gambar 24).
Spesies Septoriella memiliki ciri miselium terbenam, hialin sampai coklat pucat, bercabang dan bersepta. Konidioma berbentuk piknidium, coklat gelap, terbenam pada substrat, sub epidermal, satu lokus, memiliki dinding yang tebal dan tidak terdapat konidiofor. Sel konidiogen holoblastik, silindris tetapi membengkak pada bagian ujungnya, hialin, halus, dibentuk dari sel bagian dalam dinding lokus. Konidia berwarna coklat pucat, 8 eusepta, ujungnya tumpul dan bagian dasar mengerucut, dindingnya halus, silindris, eguttulate, lurus atau beberapa bengkok tidak beraturan (Sutton 1980).
Gambar 24 Konidia Septoriella sp. a
Sebanyak 23 spesies Septoriella telah tercatat di dunia (Index Fungorum 2013). Di Indonesia, Septoriella belum pernah dilaporkan sebelumnya, oleh sebab itu laporan ini merupakan laporan yang pertama. Selain itu, Septoriella yang menginfestasi meranti juga belum pernah dilaporkan sebelumnya. Selain pada meranti, spesies ini tersebar pada banyak inang diantaranya Arundo donax,
Bambusa sp., Euchlaena luxurians, Ficus alba, Ficus sp., Heteropogon contortus,
Pinus montana, Phragmites australis dan pada beberapa inang lain (USDA fungus host database 2006).
20. Beltraniellaportoricensis Piroz. & Patil (Hyponectriaceae)
Beltraniella portoricensis memiliki konidia amerospora, obovoid, hialin dengan ukuran 14-20 μm x 3-4 μm (Gambar 25a). Sel konidiogen simpodial, coklat muda (Gambar 25b). Seta berwarna coklat tua, tidak bercabang (Gambar 25c). Beltraniella portoricensis pada media PDA (Gambar 26).
Beltraniella portoricensis pertama kali dilaporkan oleh Piroz. dan Patil pada tahun 1970 (Index Fungorum 2013) dengan ciri seta lurus, konidiofor kadang-kadang muncul dari bagian dasar seta, simpel atau bercabang, sel konidiogen apikal, konidia lageniform sampai simbiform (navikular) berukuran 18-27 x 5-8.5 µm, pada konidia terdapat berkas hialin (Shirouzu et al. 2010).
Sebanyak 21 spesies Beltraniella telah tercatat dari berbagai wilayah di dunia (Index Fungorum 2013). Beltraniella portoricensis yang menginfestasi serasah meranti asal Indonesia belum pernah dilaporkan sebelumnya dan ini merupakan laporan yang pertama. Informasi ini berguna untuk mengetahui penyebaran dari Beltraniella portoricensis. Selain pada meranti, Beltraniella portoricensis banyak terdapat pada serasah daun dan tersebar pada banyak inang diantaranya Acacia aulacocarpa, Agathis loranthifolia, Areca catechu, Mangifera indica, Lithocarous edulis, Pinus khasya, Quercus acuta dan pada beberapa inang lain (USDA fungus host database 2006).
Gambar 25 Konidia Beltraniella portoricensis (a), sel konidiogen (b) dan seta (c)
a
c
Dipterocarpus sp., Eucalyptus urophylla, Licuala longicalycata dan Ocotea nemodaphne (USDA fungus host database 2006).
Gambar 28 Konidia Cryptophialoidea fasciculata (a), sel konidiogen (b), seta (c)
Gambar 29 Koloni Cryptophialoidea fasciculata pada media PDA, permukaan atas (A) dan permukaan bawah (B)
Cryptophialoidea fasciculata IPBCC.13.1079 berada satu clade dengan
Lecanicillium saksenae dengan nilai bootstrap sebesar 68% (Gambar 30). Hal ini menegaskan bahwa isolat tersebut merupakan Lecanicillium saksenae. Tetapi berdasarkan pengamatan secara morfologi, Cryptophialoidea fasciculata memiliki seta tidak bercabang, simpel, dan berwarna coklat. Sel konidiogen monofialid, berbentuk lageniform, berukuran 6-8 μm x 2-3.5 μm, fialid tersusun dalam ikatan/ bundel. Konidia berbentuk falkat, hialin, tidak bersepta dan berukuran 20-31 μm x 1-2 μm. Sedangkan Lecanicillium saksenae memiliki hifa aerial, fialid, ramping ke arah apeks. Konidia dimorfik yaitu makrokonidia berbentuk falkat dengan ukuran 6-13 x 1.5-2 μm dan mikrokonidia berbentuk elipsoid sampai fusoid dengan ukuran 2.5-5 x 1.5-2 μm (Sukarno et al. 2009).
Adanya perbedaan hasil antara analisis molekuler dan pengamatan morfologi pada isolat ini, diduga karena tidak adanya data sekuen nukleotida dari
Cryptophialoidea fasciculata di NCBI/Genbank, sehingga ketika dilakukan analisis filogenetik, sekuen dari Cryptophialoidea fasciculata akan me-alignment
A B
c
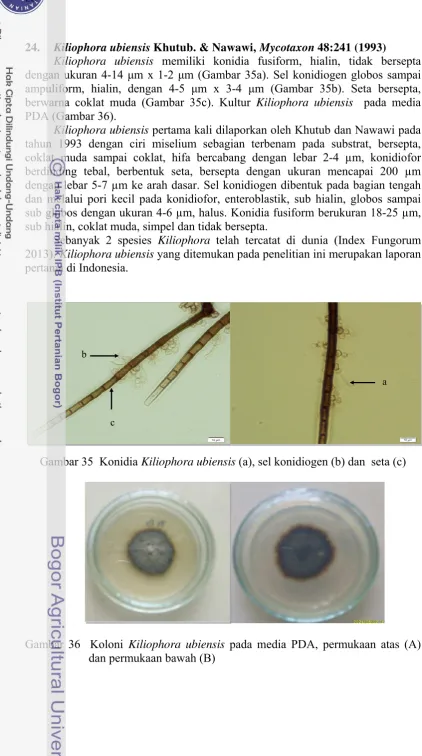
24. Kiliophora ubiensis Khutub. & Nawawi, Mycotaxon 48:241 (1993)
Kiliophora ubiensis memiliki konidia fusiform, hialin, tidak bersepta dengan ukuran 4-14 μm x 1-2 μm (Gambar 35a). Sel konidiogen globos sampai ampuliform, hialin, dengan 4-5 μm x 3-4 μm (Gambar 35b). Seta bersepta, berwarna coklat muda (Gambar 35c). Kultur Kiliophora ubiensis pada media PDA (Gambar 36).
Kiliophora ubiensis pertama kali dilaporkan oleh Khutub dan Nawawi pada tahun 1993 dengan ciri miselium sebagian terbenam pada substrat, bersepta, coklat muda sampai coklat, hifa bercabang dengan lebar 2-4 µm, konidiofor berdinding tebal, berbentuk seta, bersepta dengan ukuran mencapai 200 µm dengan lebar 5-7 µm ke arah dasar. Sel konidiogen dibentuk pada bagian tengah dan melalui pori kecil pada konidiofor, enteroblastik, sub hialin, globos sampai sub globos dengan ukuran 4-6 µm, halus. Konidia fusiform berukuran 18-25 µm, sub hialin, coklat muda, simpel dan tidak bersepta.
Sebanyak 2 spesies Kiliophora telah tercatat di dunia (Index Fungorum 2013). Kiliophora ubiensis yang ditemukan pada penelitian ini merupakan laporan pertama di Indonesia.
Gambar 35 Konidia Kiliophora ubiensis (a), sel konidiogen (b) dan seta (c)
Gambar 36 Koloni Kiliophora ubiensis pada media PDA, permukaan atas (A) dan permukaan bawah (B)
b
c
Gambar 38 Konidia Minimidochium setosum (a) dan seta (b)
26. Monodisma fragilis Alcorn, Trans. Br. Mycol. Soc. 65: 140 (1975)
Monodisma fragilis memiliki konidia fusiform, pragmospora dengan 7 septa, hialin, dan berukuran 58 x 9 μm (Gambar 39).
Monodisma fragilis pertama kali dilaporkan oleh Alcorn pada tahun 1975 dengan ciri hifa berukuran 2-5 µm, panjang konidiofor 700 µm, lebar 4.5-7.5 µm, konidia 3-13 distosepta dengan ukuran 27-80 x 5-10 µm.
Gambar 39 Konidia Monodisma fragilis
Sebanyak 1 spesies Monodisma telah tercatat di dunia (Index Fungorum 2013). Monodisma fragilis sebelumnya telah dilaporkan dari Polandia yang menginfestasi Carex gracilis dan dari Papua Nugini pada Saccharum officinarum
(USDA fungus host database 2006). Di Indonesia, Monodisma belum pernah dilaporkan sebelumnya, oleh sebab itu laporan ini merupakan laporan yang pertama.
27. Nodulisporium sp. (Xylariaceae)
Nodulisporium sp. memiliki konidia terdapat pada ujung, amerospora, hialin sampai subhialin kemudian berpigmen berukuran 2-3 μm x 1-2 μm (Gambar 40a). Konidiofor tegak lurus, bercabang, berpigmen (Gambar 40b).
Sebanyak 44 spesies Nodulisporium telah tercatat dari berbagai wilayah di dunia (Index Fungorum 2013). Nodulisporium tersebar pada banyak inang diantaranya Abies beshanzuensis, Acca sellowiana, Acer rubrum, Mangifera indica, Pinus strobus, Oryza sativa, Thuja plicata dan pada beberapa inang lain (USDA fungus host database 2006).
Gambar 40 Konidia Nodulisporium sp. (a) dan konidiofor (b)
28. Stilbella fimetaria Lindau
Stilbella fimetaria memiliki konidia satu sel, tidak bersepta, hialin, berukuran 5-7 μm x 2-3 μm (Gambar 41a). Konidiofor berupa sinema (Gambar 41b).
Gambar 41 Konidia Stilbella fimetaria (a), konidiofor (b)
Sebanyak 94 spesies Stilbella telah tercatat di dunia (Index Fungorum 2013). Stilbella fimetaria pertama kali dilaporkan oleh Lindau tahun 1905 (Index Fungorum 2013). Spesies ini tersebar pada banyak inang diantaranya Acer circinatum, Acer rubrum, Allium cepa, Bambusa sp., Eucalyptus sp., Solanum tuberosum, Theobroma cacao dan pada banyak inang lainnya (USDA fungus host database 2006).
a b
a
29. Virgatospora echinofibrosa Finley, Mycologia 59: 538 (1967)
Virgatospora echinofibrosa memiliki konidia berbentuk fusiform, bersepta, berwarna hijau, 4 sel berukuran 36-44 μm x 9-11μm (Gambar 42a). Konidiofor berupa sinema (Gambar 42b) dan memiliki papila (Gambar 42c).
Virgatospora echinofibrosa pertama kali ditemukan oleh Finley pada tahun 1967 dengan ciri sinema, konidia bersepta, obovoid, alantoid sampai fusiform berukuran 39-50 x 9-15 µm dan pada bagian dasar mengerucut.
Sebanyak 2 spesies Virgatospora telah tercatat di dunia (Index Fungorum 2013). Selain pada serasah meranti, spesies ini juga terdapat pada ranting Inga goldmanii (USDA fungus host database 2006).
Gambar 42 Konidia Virgatospora echinovibrosa (a), konidiofor (b) dan papila (c)
Jumlah spesies cendawan yang ditemukan pada penelitian ini lebih sedikit dari jumlah spesies yang dilaporkan oleh Maria dan Sridhar (2003) dari serasah mangrove, oleh Pinruan et al. (2007) dari serasah palem dan oleh Osono et al. (2009) dari serasah Shorea obtusa (80 spesies). Perbedaan jumlah spesies ini diduga disebabkan oleh perbedaan pengambilan sampel (musim, frekuensi, jumlah dan metode pengambilan sampel). Maria dan Sridhar (2003) menemukan 91 spesies cendawan selama musim hujan-musim panas pada serasah 5 jenis mangrove, Pinruan et al. (2007) menemukan sebanyak 147 spesies cendawan dengan pengambilan sampel selama beberapa bulan. Osono et al. (2009) menggunakan dua metode yaitu metode disinfeksi permukaan serasah daun dan metode pencucian daun, sehingga peluang untuk mengisolasi cendawan yang tidak bersporulasi pada sampel serasah sangat besar. Sebaliknya, penelitian ini hanya menggunakan metode observasi langsung serasah pada satu kali pengambilan sampel. Isolasi dilakukan terbatas pada spora yang ditemukan pada serasah. Metode ini memiliki kelebihan yaitu dapat mengetahui dengan tepat cendawan yang menginfestasi serasah dan mampu membentuk spora pada serasah.
Faktor lain yang mempengaruhi jumlah spesies yang ditemukan kemungkinan adalah adanya metabolit sekunder. Shorea memiliki senyawa fenolik, seperti oligostilbenoid, flavonoid, fenil propanoid dan turunan asam fenolik yang dapat menghambat kolonisasi cendawan. Senyawa oligostilben
a
(Shirouzu et al. 2009). Cryptophialoidea fasciculata dan Lasiodiplodia theobromae penghuni serasah ranting memiliki FK berturut-turut sebesar 5.7% dan 6.5%, sehingga tergolong ke dalam spesies yang sering ditemukan pada serasah ranting. Cendawan lainnya termasuk kedalam kategori kadang-kadang dan jarang ditemukan (Tabel 2).
Tabel 2 Frekuensi keberadaan dari cendawan saprob berspora penghuni serasah meranti
Nama spesies cendawan Ranting Daun Total % FK
Askomiset
Annulohypoxylon purpureonitens (Aap) 2 0 2 0.7
Diatrype chlorosarca (Adc) 5 0 5 1.9
Didymosphaeria epidermidis (Ade) 3 0 3 1.1
Lophiostoma sp. (Als) 2 0 2 0.7
Lophodermium sp. (Ald) 0 2 2 0.7
Pemphidium sp. (Apd) 0 30 30 11.5
Valsa sp. (Avs) 1 0 1 0.3
Cendawan anamorf Soelomiset
Coniella musaiaensis (Ccm) 3 0 3 1.1
Coryneum betulinum (Ccb) 1 0 1 0.3
Hendersoniopsis thelebola (Cht) 1 0 1 0.3
Lasiodiplodia theobromae (Clt) 17 0 17 6.5
Lasmeniella guaranitica (Clg) 1 0 1 0.3
Leptodothiorella sp. (Cld) 1 0 1 0.3
Massariothea themedae (Cmt) 2 0 2 0.7
Pestalotia guepinii (Cpg) 2 0 2 0.7
Pestalotiopsis sp. (Cps) 0 1 1 0.3
Pseudolachnea hispidula (Cph) 1 0 1 0.3
Septoriella sp. (Cst) 6 0 6 2.3
Soelomiset sp. 1 (Csp) 1 0 1 0.3 Hifomiset
Beltraniellaportoricensis (Hbp) 0 44 44 16.9
Cryptophialoidea fasciculata (Hcf) 15 0 15 5.7
Hermatomyces sphaericus (Hhs) 2 0 2 0.7
Kiliophora ubiensis (Hku) 0 2 2 0.7
Minimidochium setosum (Hms) 1 0 1 0.3
Monodisma fragilis (Hmf) 4 0 4 1.5
Nodulisporium sp. (Hns) 2 0 2 0.7
Stilbella fimetaria (Hsf) 0 8 8 3.0
Virgatospora echinofibrosa (Hve) 4 0 4 1.5
Hifomiset sp. 1 (Hh1) 0 1 1 0.3 Jumlah individu cendawan 77 88 165
Jumlah sampel serasah 130 130 260
daun. Karena indeks kemerataan kurang dari 1 maka pada masing-masing substrat terdapat spesies cendawan saprob yang dominan.
Analisis koresponden dari komunitas cendawan pada serasah ranting dan daun memperlihatkan bahwa spesies cendawan seperti Annulohypoxylon purpureonitens, Diatrype chlorosarca, Didymosphaeria epidermidis,
Lophiostoma sp., Valsa sp., Soelomiset sp.1, Coniella musaiaensis, Coryneum betulinum, Hendersoniopsis thelebola, Lasiodiplodia theobromae, Lasmeniella guaranitica, Leptodothiorella sp., Massariothea themedae, Pestalotia guepinii,
Pseudolachnea hispidula, Septoriella sp., Cryptophialoidea fasciculata,
Hermatomyces sphaericus, Minimidochium setosum, Monodisma fragilis,
Nodulisporium sp., dan Virgatospora echinofibrosa merupakan cendawan yang umum ditemukan pada serasah ranting. Pada serasah daun meranti, cendawan yang ditemukan yaitu Kiliophora ubiensis, Lophodermium sp., Pemphidium sp.,
Pestalotiopsis sp., Beltraniellaportoricensis, Stilbella fimetaria dan Hifomiset sp. 1 (Gambar 46).
Gambar 46 Analisis koresponden yang memperlihatkan hubungan antara spesies cendawan (akronim) dengan organ tanaman meranti. (Untuk akronim lihat tabel 2)
Analisis koresponden memperlihatkan hubungan antara spesies cendawan dengan organ tanaman. Spesies tertentu hanya ditemukan pada serasah ranting meranti atau serasah daun meranti. Perbedaan komposisi komunitas cendawan pada tipe organ yang berbeda pada meranti, mengindikasikan bahwa cendawan pada meranti bersifat substrat spesifik. Informasi mengenai sifat-sifat organ dari tanaman meranti terbatas, sehingga sulit menentukan faktor-faktor yang dapat mempengaruhi perbedaan komunitas pada setiap jenis organ.
4 SIMPULAN DAN SARAN
Simpulan
Sebanyak 29 jenis cendawan saprob berspora telah ditemukan pada serasah meranti yaitu Annulohypoxylon purpureonitens, Diatrype chlorosarca,
Didymosphaeria epidermidis, Lophiostoma sp., Valsa sp., Soelomiset sp.1,
Coniella musaiaensis, Coryneum betulinum, Hendersoniopsis thelebola,
Lasiodiplodia theobromae, Lasmeniella guaranitica, Leptodothiorella sp.,
Massariothea themedae, Pestalotia guepinii, Pseudolachnea hispidula,
Septoriella sp., Cryptophialoidea fasciculata, Hermatomyces sphaericus,
Minimidochium setosum, Monodisma fragilis, Nodulisporium sp., Virgatospora echinofibrosa pada serasah ranting dan Kiliophora ubiensis, Lophodermium sp.,
Pemphidium sp., Pestalotiopsis sp., Beltraniella portoricensis, Stilbella fimetaria, Hifomiset sp. 1 pada serasah daun.
Keanekaragaman cendawan pada serasah ranting lebih tinggi dibandingkan serasah daun dengan nilai indeks Shannon-Weiner pada serasah ranting yaitu 2.608 dan pada serasah daun sebesar 1.205. Distribusi setiap spesies dalam komunitas cendawan tidak merata pada substrat ranting dan daun.
Beltraniella portoricensis dan Pemphidium sp. merupakan cendawan yang paling sering ditemukan pada serasah daun meranti dengan FK sebesar 16.9% dan 11.5%. Cryptophialoidea fasciculata dan Lasiodiplodia theobromae memiliki FK sebesar 5.7% dan 6.5% yang tergolong ke dalam spesies yang sering ditemukan pada serasah ranting meranti, sedangkan spesies cendawan yang lain termasuk ke dalam kategori kadang-kadang dan jarang ditemukan.
Saran
Berdasarkan hasil penelitian, banyak hal yang dapat mempengaruhi dominansi suatu spesies cendawan terhadap spesies cendawan yang lain. Oleh karena itu, faktor yang mempengaruhi perbedaan struktur komunitas cendawan pada serasah ranting dan serasah daun sangat sulit ditentukan. Penelitian selanjutnya harus mempertimbangkan semua faktor yang mempengaruhi frekuensi keberadaan cendawan pada meranti terutama pada inang. Untuk mengetahui komunitas cendawan pada serasah meranti secara lengkap, pengamatan pada beberapa musim selama satu tahun diperlukan.
DAFTAR PUSTAKA
Alcorn JL. 1975. Monodisma, a new genus of Hyphomycetes. Trans. Br. Mycol. Soc. 65: 140.