DNA FRAGMENTATION ON HCT-116 COLON CANCER CELL IN THE
PRESENCE OF SHORT CHAIN FATTY ACID (SCFA) EXTRACT
Nurul Kurnia Sari and Maggy T Suhartono
Department of Food Science and Technology, Faculty of Agricultural Engineering, Bogor Agricultural University, IPB Darmaga Campus,
PO Box 220, Bogor, West Java, Indonesia
Phone: +62 858 8385 3225, e-mail: [email protected]
ABSTRACT
Type 3 resistant starch (RS3) is retrogaded starch that resists digestion in the small intestine of healthy people and fermented in the large intestine. The main fermentation products are the short chain fatty acids (SCFA) acetate, propionate, and butyrate. Butyrate is the major energy source for the colonocytes and have anti-carcinogenic effects mainly by affecting proliferation, differentiation, and apoptosis of colonocytes. The aim of this study was to analyze apoptotic DNA fragments of HCT-116 human colon cancer cell incubated with SCFA derived from fermentation of RS3 of sweet potato (Ipomoea batatas) by C. Butyricum BCC B2571 using DNA ladder assay. The DNA bands detected from attached cells treated with SCFA containing 1.00 and 1.25 mM butyrate were different from the control, appeared as longer smear. There were separate DNA bands in the floating cells in the treated medium (SCFA containing 0.625, 1.00, and 1.25 mM butyrate) at ±100 bp. These bands were not presence in the control. DNA fragmentation was not clear in the attached cells, but was observed as a separate bands in the floating cells in the medium which contained the dead cells. This research still needs optimazion of DNA extraction to get the best result of DNA fragmentation assay.
1
I.
INTRODUCTION
A.
BACKGROUND
Sweet potato (Ipomoea batatas) is an alternative food rich in starch. In Indonesia, economical value of sweet potato is low, compared with other starch sources. However, the sweet potatoes which are rich in starch made it a potential source of resistant starch. At present, various studies about resistant starch had been conducted. Recent studies showed that resistant starch consumption may reduce colorectal cancer risk.
Resistant starch has been defined as total amount of starch, and the products of starch degradation that resists digestion in the small intestine of healthy people. There are four types of resistant starch. Type 1 resistant starch (RS1) is starch that escapes digestion in the small intestine due to physical protection by the food matrix, such as grains. Type 2 resistant starch (RS2) is raw starch granules (ungelatinized) with compact structure which limits accessibility of digestive enzymes. Type 3 resistant starch (RS3) is retrogaded starch in which parts of the starch chains can crystallize into components that are less digestible. Most often, this occurs by cooking and cooling starch-containing foods. And the last, type 4 resistant starch (RS4) is not found naturally in foods. Starch that has been chemically modified to introduce bonds that are not digestible by human enzymes.
Starches that resist small intestinal breakdown are fermented by the resident bacteria in the
large intestine, producing a variety of end products, the most significant of which is the short chain
fatty acids (SCFA). SCFA, especially butyrate maintenance of colonic health and barrier function, butyrate has drawn most attention as this fatty acid is the major energy source for the colonocytes (Roediger 1990). Furthermore, butyrate has been shown to have anti-carcinogenic effects mainly by affecting proliferation, differentiation and apoptosis of colonocytes (Scheppach, et. al 1995).
There are several bacterial species which produce butyric acid. It is an anaerobic process. The genera Clostridium, Butyribacterium, and Butyrivibrio are the mostly studied microorganism. For commercial purposes, Clostridium species are preferred for butyric acid production. C. butyricum, C. tyobutyricum, C. thermobutyricum, C. beijerinckii, and C. populeti are of interest for the production of butyrate. Purwani, et. al (2012) observed that RS3 derived from sago and rice starch could be well utilized as substrate by Clostridium butyricum, and revealed that good proportion of acetate: propionat: butyrate in that fermentation.
Apoptosis occurs normally during development and aging and as a homeostatic mechanism to maintain cell populations in tissues and the cell is an active participant in its own demise (“cellular suicide”). Apoptosis is a form of programmed cell death characterized by cytoplasmic condensation, plasma membrane blebbing and nuclear pycnosis, leading to nuclear DNA breakdown into multiples of ~200 bp oligonucleosomal size fragments (Chen and Ioannou 1996).
2
in the appearance of “DNA laddering” when the DNA is analyzed by agarose gel electrophoresis (Arends, et. al 1990). Principle of this assay is apoptotic DNA binds quickly to glass fiber fleece in the presence of a chaotropic salt, guanidine hydrochloride (guanidine HCl). After cellular impurities are washed off the fleece, the DNA is released from the fleece with a low salt buffer (Wyllie, et. al 1998).
B.
OBJECTIVE
3
II.
LITERATURE REVIEW
A.
RESISTANT STARCH (RS)
Resistant starch (RS) has been defined as total amount of starch, and the products of starch degradation that resists digestion in the small intestine of healthy people then enter the large bowel. It occurs for various reasons including chemical structure, cooking of food, chemical modification, and food mastication (Topping and Clifton 2001).There are four types of resistant starch. Starch found in plant cell walls that is inaccessible to amylase activity is one type of resistant starch, designated type 1 resistant starch (RS1). Food sources of RS1 include partially milled grains and seeds. Resistant starch also may be formed during food processing. Ungelatinized granules of starch are typically resistant to enzymatic digestion and are designated type 2 resistant starch (RS2). This type of starch can be found in potatoes and unripe (green) bananas. Cooking and cooling starchy foods by moist heat or extrude starchy foods, for example, generates retrogade starch called type 3 resistant starch (RS3). Chemical modifications of starch, such as formation of starch esters, or cross-bonded starches, also result in resistant starch, called type 4 resistant starch (RS4). RS3 and RS4 may be partially fermented by colonic bacteria (Gropper, et. al 2009). Interestingly, if RS2 granules are heated to over 100ºC, the granularity is lost and starch is gelatinized or the granules swell thereby increasing the availability of the starch to amylase. However, when the starch cools, there is some recrystallization of the starch. This is called retrogradation, which is resistant to α–amylase hydrolysis (Medeiros and Wildman 2012).
Starch that is trapped within whole plant cells or within the food matrix, and some starch granules that have not been fully gelatinized, are hydrolysed only very slowly by a-amylase and therefore may escape complete digestion in the small intestine. The energy yield to the body from this source is less than that provided if starch is digested and absorbed in the small intestine (Englyst, et. al 1996). The glycemic effects of starch is therefore lowered, which decreases insulin secretion. In addition to glycemic management resistant starches may play role in weight management as well (Medeiros and Wildman 2012).
The material included in the definition of RS reaches the human large intestine and thus becomes a substrate for microbial fermentation. The end-products are H2and CO2, CH4 in about half the population, and short-chain fatty acids. SCFA consist primarily of butyrate, propionate and acetate (Englyst, et. al 1996). SCFA contribute to normal large bowel function and prevent pathology through their actions in the lumen and on the colonic musculature and vasculature, through their metabolism by colonocytes (Topping and Clifton 2001).
B.
SHORT CHAIN FATTY ACID (SCFA)
4
varying amounts depending on the diet and the composition of the intestinal microbiota (Marleen,
et. al 2003). SCFA constitute approximately two-thirds of the colonic anion concentration (70-130 mmol/l), mainly as acetate, propionate, and butyrate (Mortensen and Clausen 1996). Various population data show that SCFA production is in order of acetate > propionate > butyrate in a molar ratio of approximately 60:20:20 or 3:1:1, respectively in the proximal and distal colon (Cummings, et. al 1987; Topping and Clifton 2001).
The major source of this in the human colon is thought to be plant cellwall polysaccharides such as cellulose, pectins, and hemicelluloses, currently referred to in human nutrition as dietary fibre. Starch would also be a suitable substrate if it were to reach the colon in significant quantities. Plant cell-wall polysaccharides are composed of hexose (glucose and galactose), pentose (xylose and arabinose), and uronic acid monomers which are fermented by gut micro-organisms along a variety of pathways. The important feature to remember of this metabolism is that it is anaerobic. Many lines of evidence therefore support the idea that fermentation occurs in the colon but, because of the difficulties in obtaining access to the human large bowel during normal digestion, most of the evidence for fermentation has, of necessity, been indirect (Cummings, et. al 1987).
There are several bacterial species which produce butyric acid. Most butyric acid-producing bacteria form acetic acid in addition to butyric acid as their major fermentation products.
Clostridium butyricum, C. tyobutyricum, C. thermobutyricum, C. beijerinckii, and C. populeti are of interest for the production of butyrate. There are some benefits using Clostridium butyricum.
These bacteria needs simple growth medium, produces high metabolite, and easy to isolate. Therefore, Clostrium butyricum also has butyrogenic activity, produce more butyric acid than acetic acid and propionic acid. C. butyricum had the highest specific growth rates compared with other genera. On the other hand, Clostridium spp. are generally less dominant in the human colon and hence the role of Clostridium in the production of butyrate in humans is unclear (Wang 1999).
SCFA will be absorbed and contribute about 3 kilocalories per gram (Medeiros and Wildman 2012). SCFA are rapidly absorbed and have shown to have distinct bioactivity depending on their chain length. With regard to maintenance of colonic health and barrier function, butyrate has drawn most attention as this fatty acid is the major energy source for the colonocytes (Roedriger 1990). Furthermore, butyrate has been shown to have anti-carcinogenic effects mainly by affecting proliferation, differentiation, and gene expression of colonocytes (Scheppach, et. al 1995). Butyrate concentration didn‟t show a significant correlation with pH or
propionic concentration (McIntyre, et. al 1993). The production of propionate is by two main, but circuitous, routes, firstly, involving fixation of CO2, to form succinate which is subsecquently decarboxylated (the 'decarboxylic acid pathway') or secondlly, from lactate and acrylate (the 'acrylate pathway'). Acetate is usually formed by the oxidative decarboxylation of pyruvate and butyrate by reluction of acetoacetate formed from acetate (Cummings, et. al 1981).
5
1. Cells of the ceco-colonic epithelium that use butyrate as a major substrate for the maintenance of energy producing path-ways;
2. Liver cells that metabolize residual butyrate with propionate used for gluconeogenesis; 50% to 70% of acetate is also taken up by the liver;
3. Muscle cells that generate energy from the oxidation of residual acetate.
In ruminants and other herbivores, SCFA are absorbed and transported via the portal vein to the liver, and the fraction not absorbed is distributed to the other body organs and tissues for metabolism. In herbivores, peripheral venous SCFA concentrations are high due to comparatively low visceral extraction and high rates of absorption into the circulation. However, human peripheral venous blood concentrations are normally low, and only acetate is present in measurable amounts. This profile reflects the lower SCFA production rates and greater visceral extraction in omnivores, meaning that human peripheral venous SCFA are not representative of those in the portal circulation. Human experimentation has been confined largely to fecal measurements, which are also limited as 95% of SCFA are produced and absorbed within the colon (Topping and Clifton 2001).
Purwani, et. al (2012) study revealed that type 3 resistant starch (RS3) derived from sago and rice starch that treated with pullulanase could be well utilized as substrate by Clostridium butyricum and showed good proportion of acetate: propionate: butyrate in the bacterial culture filtrate. The fermentation of resistant starch type 3 (RS3) derived from sago starch treated with pullulanase (RSSP) by C. butyricum produced 17.65 mM of butyrate, and fermentation of RS3 derived from rice starch treated with pullulanase (RSRP) by C. butyricum produced 21.76 mM, higher concentration than fermentation of RSSP. This study implied that difference of starch source resulted difference of SCFA concentration.
C.
APOPTOSIS
6
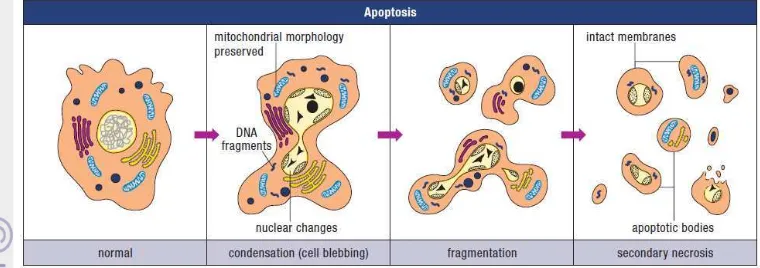
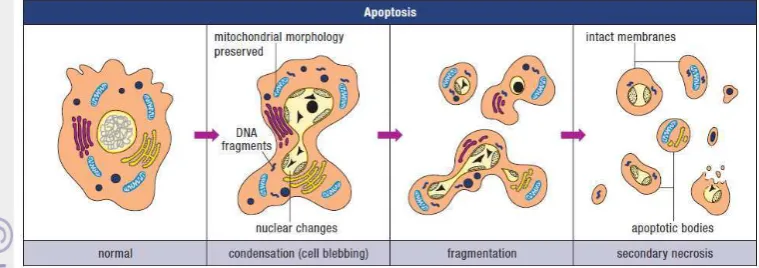
Figure 1 Illustration of the morphological features of apoptosis (Wyllie, et. al 1998)
Apoptosis is characterised by an initial shrinkage of the cell, which in a tissue means breaking cell-to-cell contacts with neighbours and rounding up. The consequence of this stage tends to be that the volume of the cell becomes smaller, and the cytoplasmic internal membranes, ribosomes, mitochondria, and other cytoplasmic organelles are more concentrated in the cytoplasm, which then consequently looks darker. The organelles remain intact and healthy looking very late into the process of death, suggesting that the cell continues metabolic activity for some considerable time. This indeed can be demonstrated in many cell systems by using various inhibitors of metabolic processes (RNA and protein synthesis inhibitors), which delay the progression of a cell through the apoptotic sequence. Apoptosis characteristically involves single isolated cells and not clusters. The DNA or chromatin material in the nucleus condenses very extreme (Potten and Wilson 2004). Pyknosis is the result of chromatin condensation and this is the most characteristic feature of apoptosis. On histologic examination with hematoxylin and eosin stain, apoptosis involves single cells or small clusters of cells. The apoptotic cell appears as a round or oval mass with dark eosinophilic cytoplasm and dense purple nuclear chromatin fragments. Electron microscopy can better define the subcellular changes. Early during the chromatin condensation phase, the electron-dense nuclear material characteristically aggregates peripherally under the nuclear membrane although there can also be uniformly dense nuclei.
7
do not release their cellular constituents into the surrounding interstitial tissue; (2) they are quickly phagocytosed by surrounding cells thus likely preventing secondary necrosis; and (3) the engulfing cells do not produce anti-inflammatory cytokines (Savill and Fadok 2000; Kurosaka, et. al 2003).
The biochemical hallmark of apoptosis is the fragmentation of the genomic DNA, an irreversible event that commits the cell to die and occurs before changes in plasma membrane permeability (prelytic DNA fragmentation). The fragmentation of chromosomal DNA is a hallmark of apoptosis and may facilitate apoptosis by terminating DNA replication and gene transcription. In many systems, this DNA fragmentation has been shown to result from activation of an endogenous Ca2+ and Mg2+ dependent nuclear endonuclease. This enzyme selectively cleaves DNA at sites located between nucleosomal units (linker DNA) generating mono- and oligonucleosomal DNA fragments (Arends et. al, 1990; Wyllie, et. al 1998).
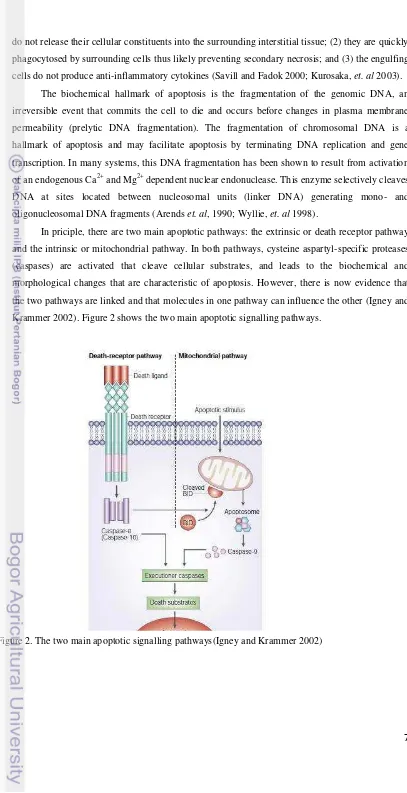
In priciple, there are two main apoptotic pathways: the extrinsic or death receptor pathway and the intrinsic or mitochondrial pathway. In both pathways, cysteine aspartyl-specific proteases (caspases) are activated that cleave cellular substrates, and leads to the biochemical and morphological changes that are characteristic of apoptosis. However, there is now evidence that the two pathways are linked and that molecules in one pathway can influence the other (Igney and Krammer 2002). Figure 2 shows the two main apoptotic signalling pathways.
8
Apoptotic cells exhibit several biochemical modifications such as protein cleavage, protein cross-linking, DNA breakdown, and phagocytic recognition that together result in the distinctive structural pathology described previously (Hengartner 2000). Caspases are widely expressed in an inactive proenzyme form in most cells and once activated can often activate other procaspases, allowing initiation of a protease cascade. Some procaspases can also aggregate and autoactivate. This proteolytic cascade, in which one caspase can activate other caspases, amplifies the apoptotic signaling pathway and thus leads to rapid cell death (Elmore 2007).
Although DNA fragmentation into oligonucleosomal ladders is characteristic of apoptosis, recent evidence indicates that not all cells undergo such extensive DNA fragmentation (Cohen, et. al1. 1992 in Chen and Ioannou 1996). In fact, fragmentation of DNA into kilobase-size fragments appears to be an early event in apoptosis, preceding the complete digestion of DNA into multiples of nucleosomal size fragments (Cohen and Sun 1994 in Chen and Ioannou 1996).
D.
DNA LADDER ASSAY
One of the most easily measured features of apoptotic cells is the break-up of the genomic DNA by cellular nucleases. These DNA fragments can be extracted from apoptotic cells and result in the appearance of “DNA laddering” when the DNA is analyzed by agarose gel electrophoresis (Arends, Morris, and Wyllie 1990). Principle of this assay is apoptotic DNA binds quickly to glass fiber fleece in the presence of a chaotropic salt, guanidine hydrochloride (guanidine HCl). After cellular impurities are washed off the fleece, the DNA is released from the fleece with a low salt buffer (Wyllie, et. al 1998).
DNA ladder is asolution of DNA molecules of different lengths used in agarose gel electrophoresis. it is applied to an agarose gel as a reference to estimate the size of unknown DNA molecules. In addition it can be used to approximate the mass off a band by comparison to a special mass ladder. Different DNA ladders are commercially available depending on expected DNA length. The 1 kb ladder with fragment ranging from about 0.5 kbp to 10 or 12 kbp and the 100 bp ladder with fragment ranging from 100 bp to just above 1000 bp are the most frequent. DNA ladders are often produced by a suitable restriction digest of plasmid.
DNA electrophoresis is an analytical technique used to separate DNA fragments by size. An electric field forces the fragments to migrate through a gel. DNA molecules normally migrate from negative to positive potential due to the net negative potential due to the net negative charge of the phosphate backbone of the DNA chain. At the scale of the length of DNA molecules, the gel looks much like a random, intricate network. Longer molecules migrate more slowly because they are more easily „trapped‟ in the network. After the separation is completed, the fractions of DNA fragments of different length are often visualized using a fluorescent dye spesific for DNA, such as ethidium bromide. The gel shows bands corresponding to different DNA molecules populations with different molecular weight. Fragment size is usually reported in “nucleotides”, “base pairs” or
9
separated. Fragment size determination is typically done by comparison to commercially available DNA ladders containing linear DNA fragments of known length.
The types of gel most commonly used for DNA electrophoresis are agarose (for relatively long DNA molecules) and polyacrylamide (for high resolution of short DNA molecules, for example in DNA sequencing). Electrophoresis techniques used in the assessment of DNA damage include alkaline gel electrophoresis and pulsed field gel elctrophoresis. The measurement and analysis are mostly done with a specialized gel analysis software.
10
III.
MATERIALS AND METHODS
A.
MATERIALS AND EQUIPMENTS
1. MATERIALS
The sample used for this research was the fermentation product of type III resistant starch (RS3) derived from Ipomoea batatas var. Jago provided by Indonesian Center of Agricultural Post Harvest Research and Development, Bogor in the form of cell free supernatant consisted of 52.95 mM acetic acid, 66.93 mM propionic acid, and 92.41 mM butyric acid. HCT-116 colon cancer cells was provided by Primate Research Center, Bogor Agricultural University. Materials used in this research were Dulbecco‟s Modified Eagle‟s Medium (DMEM) Gipco USA, Fetal Bovine Serum (FBS) Gipco USA, antibiotics (penicillin and streptomycin) Invitrogen USA, Phosphate Buffered Saline (PBS) Gipco USA, Trypsin 0.25% Difco USA, Qiagen DNA extraction kit (lysis buffer, Aw 1, Aw 2, and AE buffer), agarose powder Promega, ethidium bromide Gipco USA, gel loading buffer, double-distilled water, and TAE buffer 50x Gipco USA (0.484 gram Tris-Base, 0.1142 mL acetic acid, and 0.2 mL 0.5M EDTA pH 8.0).
2. EQUIPMENT
The equipments used in this research were biosafety cabinet, dissposable gloves, dissposable pipettes (1 mL, 2 mL, 5 mL, 10 mL, 25 mL, and 50 mL), micropipette, T25 and T75 tissue culture flask, incubator (37oC, 5% CO2), refrigerator, water-bath, inverted microscope, centrifuge, centrifuge tube, glass bottle, gel doc, cuvette, and DNA electrophoresis.
B.
METHODS
1. Cell Culture
HCT-116 colon cancer cells were warmed up from cryogenic state using waterbath with 37 ºC. Then, the cells were pelleted using centrifuge 750 g and media was discarded. HCT-116 colon cancer cells have been cultured in Dulbecco‟s Modified Eagle‟s Medium (DMEM) supplemented with 10% Fetal Bovine Serum (FBS) and 1% antibiotics (penicillin 100 unit/mL; streptomycin 100µg/mL). Cells (1 x 105) were seeded into T25 tissue culture flask. The cells were incubated at 37oC in 5% CO2 incubator.
11
hemocytometer to seed the cell into flasks in an acurate amount. 1 × 105 cells were seeded into T25 flask every subculture for maintaining.
2. Incubation with SCFA
Analysis was done using eight T25 flasks which contained 1 x 105 cells as the starting cell number. The flasks was incubated for 2 x 24 hr or to reach 50% confluency. Next, incubation was continued for another 4 x 24 hr in the absence (control) or presence of various concentration of cell free supernatant contained SCFA (0.625 mM, 1.00 mM, and 1.25 mM butyrate in the cell free supernatant). For the treatment of the experimental cultures, the cell free supernatant contained SCFA was diluted with DMEM. At the end of 4 x 24 hr incubation period, the cells were observed using inverted microscope with magnification (4x10). Then, the medium contained of dead cells was collected into centrifuge tube and the attached cells were washed with phosphate buffered saline (PBS). The medium contained of dead cells that floating in the medium and made the medium became turbid. After that, the attached cells were trypsinized with trypsin 0.25% to free the cells from adhesion to the flask followed by incubation in 37 ºC for 5-10 minutes and pipetted into centrifuge tube. Next, the attached cells in the trypsin and the medium were pelleted with centrifugation 750 g for 5 minutes. After that, the supernatant was discarded. The pellet were resuspended using 10 mL of PBS steril and centrifuged as before. Then, the supernatant was discarded. The pellet were resuspended using 1 mL PBS steril, then pipetted into eppendorf. Next, attached and dead cells were centrifuged 1000 g for 5 menit. The supernatant was discarded. And the last, the cuvettes were stored at -20oC.
3. DNA Extraction (Qiagen DNA Extraction Kit)
The cells pellet were treated for 10 minutes with 200 μl lysis buffer. Then, the sample was mixed with ethanol 95% and pipetted into filter tube (silica gel). The filter tube was centrifuged 8,000 g for 1 minute and the flow through was discarded. After that, the treated and untreated cells were washed with wash buffer (Aw 1) and centrifuged as before. Next, the treated and untreated cells were washed with wash buffer (Aw 2) and centrifuged 8,000 rpm for 3 minutes. Then, the sample was mixed with 100 μL elution buffer (AE buffer) and centrifuged 8,000 rpm for 1 minute.
4. DNA Ladder Assay
12
IV.
RESULTS AND DISCUSSION
A.
INHIBITION TO HCT-116 COLON CANCER CELL
After 4 x 24 hr incubation in the presence and absence of the cell free supernatant contained SCFA, observation was conducted using inverted microscope with magnification (4x10). Cellular debris in the media could be observed on HCT-116 cells that were treated with SCFA containing 0.625, 1.00, and 1.25 mM butyrate (figure 3). This research showed that the attached cells of control became multi layer. The treated cells with 1.00 mM butyrate showed higher confluency than the treated cells with 1.25 mM butyrate, but lower than the treated cells with 0.625 mM butyrate. This results implied that higher concentration of butyrate showed higher inhibition to HCT-116 colon cancer cell. The inhibition effect was implied at butyrate concentration as low as 0.625 mM and was lower than those reported by Purwani, et. al (2010) that the inhibition effect was observed at butyrate concentration as low as 2.6 mM. SCFA derived from fermentation of RS3 of sago decreased the viability, the total cell number, and increased the inhibition growth of HCT-116 colon cancer cell in a dose dependent manner (p>0.05).
Other research concluded that butyrate inhibited Caco-2 cell proliferation at concentration as low as 0.1 mM (Ruemmele, et. al 1999). This research implied inhibition at higher concentration of butyrate. This difference might caused by cancer cell difference that was used in each research, therefore difference in effect may occur.
(1)
(2)
(3)
(4)
13
B.
DETECTION OF APOPTOSIS USING DNA LADDER ASSAY
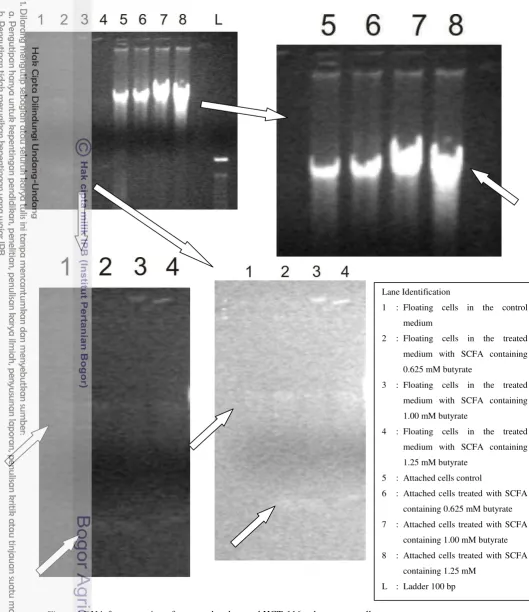
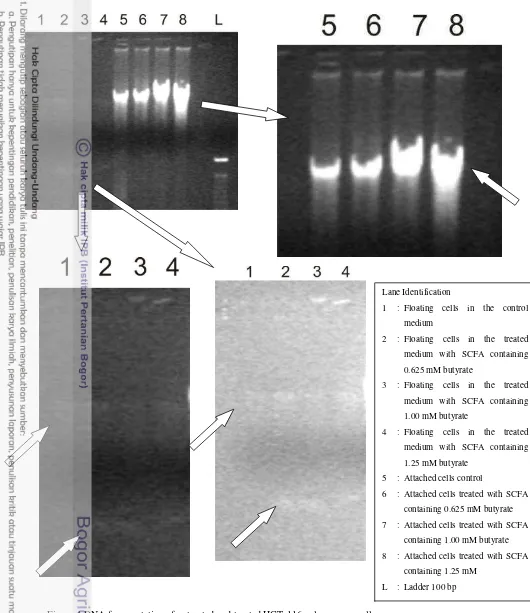
DNA fragmentation of HCT-116 colon cancer cell was observed with DNA ladder assay using electrophoresis gel. Observartion data showed different DNA bands between control and cells treated with SCFA (figure 4). DNA bands from attached cells treated with SCFA containing 0.625 mM butyrate was not significantly different from the control. However, the attached cells treated with SCFA containing 1.00 and 1.25 mM butyrate had DNA bands that were significantly different from the control: appeared as longer smear.
This research also observed DNA fragmentation of the floating cells in the medium. There were different result of DNA bands of the floating cells in the control medium and in the treated medium. There were thin DNA bands in the floating cells in the treated medium (SCFA containing 0.625, 1.00, and 1.25 mM butyrate) at ±100 bp (compared with commercial ladder), lower density than normal DNA bands. These bands were not presence in the control. This result showed that DNA of floating cells in the treated medium was fragmented and had lower density than control, the bands observed at 180-200 bp.
The appearance of the nucleosomal DNA ladder in agarose gels has become the hallmark of programmed cell death via apoptosis. This research implied that SCFA derived from fermentation of type-3 resistant starch of Ipomoea batatas induced apoptosis via DNA fragmentation. DNA fragmentation was not clear in the pellet of attached cells, but was observed as a separate bands in the pellet of floating cells in the medium. Floating cells in the medium contained dead cells. This imply that SCFA induced apoptosis at very early stage in the attached cells and later stage in the floating cells in the medium which contained of dead cells.
Other researchers had reported that butyrate inhibition towards cancer cells are via apoptosis (Purwani and Suhartono 2010; Ruemmele, et. al 1999; Hinnebusch, et. al 2002). Many studies investigated the mechanism of how SCFA induced apoptosis. Purwani and Suhartono (2010) studies showed that SCFA in bacterial supernatant induced apoptosis and changed the expression of apoptosis related genes such as Bcl-2 and Bax. It also showed that bacterial supernatant consisted of SCFA increased the expression ratio of pro-apoptotic genes.
14
Figure 4 DNA fragmentation of untreated and treated HCT-116 colon cancer cells
Lane Identification
1 : Floating cells in the control medium
2 : Floating cells in the treated medium with SCFA containing 0.625 mM butyrate
3 : Floating cells in the treated medium with SCFA containing 1.00 mM butyrate
4 : Floating cells in the treated medium with SCFA containing 1.25 mM butyrate
5 : Attached cells control
6 : Attached cells treated with SCFA containing 0.625 mM butyrate 7 : Attached cells treated with SCFA
containing 1.00 mM butyrate 8 : Attached cells treated with SCFA
15
Hinnebusch, et. al (2002) treated HT-26 and HCT-116 colon cancer cell with physiologically relevant concentrations of various SCFA, and apoptotic effects were studied by flow cytometry. Butyrate significantly increased apoptosis, whereas the other SCFA studied did not. Hinnebusch, et. al (2002) concluded that SCFA effects on apoptosis, and interestingly, only C4 (butyrate) increased the rate of programmed cell death.
Rummele, et. al (1999) studies showed that butyrate induced apoptosis (maximum 79%) Caco-2 cell line via activation of the caspase-cascade (a key event was the proteolytic activation of caspase-3, triggering degradation of poly-(ADP-ribose) polymerase (PARP)) and the mitochondrial bcl-pathway (butyrate potently up-regulatef the expression of the pro-apoptotic protein bak, without changing Caco-2 cell bcl-2 expression). Bak is a protein that induce pore formation on mitochondrial surface, therefore releasing cytochrome-c out of mitochondrial membrane forming apoptosome that could lead to activation of caspase cascade which could induce apoptosis. The caspase play an important role in apoptosis by activating Dnases, inhibiting DNA repair enzymes and breaking down structural protein in the nucleus (Elmore 2007). In summary, apoptosis is more likely when bax protein levels are high (Wyllie, et. al 1998).
The expected result of this study was clear DNA fragmentation, and there were differences of DNA fragmentation of normal and apoptosis cells. However, DNA fragmentation in this research was not clearly observed. Eventhough, variation of the concentration of agarose gel (0.8%, 1%, and 2%), voltage (75 and 100 Volt), and electrophoresis time (1 and 1.5 hours) was treated, the expected DNA fragmentation was not seen clearly. This might be related with DNA extraction methods that was not optimal, and thus affected the DNA bands in the agarose.
Figure 5. Comparison of two methods for the preparation of apoptotic DNA fragments (Herrmann, et. al 1994) Lane identification:
M : 142 bp ladder
16
17
V.
CONCLUSION AND RECOMMENDATION
A.
CONCLUSION
Based on this research, Short Chain Fatty Acids(SCFA) derived from fermentation of type-3 resistant starch of Ipomoea batatas containing 1,00 and 1, 25 mM butyrate produced DNA bands different from the control when measured with DNA ladder assay. The DNA bands of the treated cells appeared as longer smear. There were separate DNA bands in the floating cells in the treated medium (SCFA containing 0.625, 1.00, and 1.25 mM butyrate) at ±100 bp. These bands were not presence in the control. DNA fragmentation was not clear in the attached cells, but was observed as a separate bands in the floating cells in the medium. This imply that SCFA might induce apoptosis at very early stage in the attached cells and later stage in the floating cells in the medium.
B.
RECOMMENDATION
DNA FRAGMENTATION ON HCT-116 COLON CANCER CELL
IN THE PRESENCE OF SHORT CHAIN FATTY ACID (SCFA)
EXTRACT
BACHELOR THESIS
NURUL KURNIA SARI
F24080056
FOOD SCIENCE AND TECHNOLOGY
FACULTY OF AGRICULTURAL ENGINEERING
BOGOR AGRICULTURAL UNIVERSITY
DNA FRAGMENTATION ON HCT-116 COLON CANCER CELL
IN THE PRESENCE OF SHORT CHAIN FATTY ACID (SCFA)
EXTRACT
BACHELOR THESIS
NURUL KURNIA SARI
F24080056
FOOD SCIENCE AND TECHNOLOGY
FACULTY OF AGRICULTURAL ENGINEERING
BOGOR AGRICULTURAL UNIVERSITY
DNA FRAGMENTATION ON HCT-116 COLON CANCER CELL IN THE
PRESENCE OF SHORT CHAIN FATTY ACID (SCFA) EXTRACT
Nurul Kurnia Sari and Maggy T Suhartono
Department of Food Science and Technology, Faculty of Agricultural Engineering, Bogor Agricultural University, IPB Darmaga Campus,
PO Box 220, Bogor, West Java, Indonesia
Phone: +62 858 8385 3225, e-mail: [email protected]
ABSTRACT
Type 3 resistant starch (RS3) is retrogaded starch that resists digestion in the small intestine of healthy people and fermented in the large intestine. The main fermentation products are the short chain fatty acids (SCFA) acetate, propionate, and butyrate. Butyrate is the major energy source for the colonocytes and have anti-carcinogenic effects mainly by affecting proliferation, differentiation, and apoptosis of colonocytes. The aim of this study was to analyze apoptotic DNA fragments of HCT-116 human colon cancer cell incubated with SCFA derived from fermentation of RS3 of sweet potato (Ipomoea batatas) by C. Butyricum BCC B2571 using DNA ladder assay. The DNA bands detected from attached cells treated with SCFA containing 1.00 and 1.25 mM butyrate were different from the control, appeared as longer smear. There were separate DNA bands in the floating cells in the treated medium (SCFA containing 0.625, 1.00, and 1.25 mM butyrate) at ±100 bp. These bands were not presence in the control. DNA fragmentation was not clear in the attached cells, but was observed as a separate bands in the floating cells in the medium which contained the dead cells. This research still needs optimazion of DNA extraction to get the best result of DNA fragmentation assay.
Nurul Kurnia Sari. F24080056. DNA Fragmentation on HCT-116 Colon Cancer Cell in the Presence of Short Chain Fatty Acid (SCFA) Extract. Supervised by Maggy Thenawidjaja Suhartono 2012.
SUMMARY
Sweet potato (Ipomoea batatas) is an alternative food rich in starch, made it a potential source of resistant starch. Recent studies showed that resistant starch consumption may reduce colorectal cancer risk. Resistant starch is total amount of starch or the products of starch degradation that resists digestion in the small intestine of healthy people. There are four types of resistant starch.
Type 3 resistant starch (RS 3) is retrogaded starch, may be partially fermented by colonic bacteria. The end-products are H2, CO2, CH4, and short chain fatty acid (SCFA). SCFA consists primarily of butyrate, propionate, and acetate. SCFA, especially butyrate maintenance of colonic health and barrier function, butyrate has drawn most attention as this fatty acid is the major energy source for the colonocytes. Furthermore, butyrate has been shown to have anti-carcinogenic effects mainly by affecting proliferation, differentiation and apoptosis of colonocytes.
The objective of this research was to analyze apoptotic DNA fragments of HCT-116 human colon cancer cell incubated with short chain fatty acids (SCFA) derived from fermentation of type 3 resistant starch (RS3) of sweet potato (Ipomoea batatas) by C. Butyricum BCC B2571 using DNA ladder assay.
DNA FRAGMENTATION ON HCT-116 COLON CANCER CELL IN THE
PRESENCE OF SHORT CHAIN FATTY ACID (SCFA) EXTRACT
BACHELOR THESIS
In the partial fulfillment of the requirement of degree of
SARJANA TEKNOLOGI PERTANIAN
at the Department of Food Science and Technology
Faculty of Agricultural Engineering
Bogor Agricultural University
By :
NURUL KURNIA SARI
F24080056
FACULTY OF AGRICULTURAL ENGINEERING
BOGOR AGRICULTURAL UNIVERSITY
BOGOR
Title
: DNA Fragmentation on HCT-116 Colon Cancer Cell in the
Presence of Short Chain Fatty Acid (SCFA) Extract
Name
: Nurul Kurnia Sari
Student ID : F24080056
Approved by,
Academic Advisor,
(Prof. Dr. Ir. Maggy Thenawidjaja Suhartono)
NIP. 19530507 197701 2 001
Acknowledged by,
Head of Department of Food Science and Technology,
(Dr. Ir. Feri Kusnandar, M. Sc)
NIP 19680526 199303 1 004
STATEMENT LETTER OF THESIS AND SOURCE OF
INFORMATION
Hereby I genuinely stated that the bachelor thesis entitled DNA Fragmentation on HCT-116 Colon Cancer Cell in the Presence of Short Chain Fatty Acid (SCFA) Extract is an authentic work of mine under supervision of academic advisor and never being presented in any forms and universities. All the informations taken and quoted from published or unpublished works of other writers had been mentioned in the texts and attached in the references at the end of the bachelor thesis.
Bogor, July 2011 The undersigned,
© Created right owned by Nurul Kurnia Sari, 2012 All rights reserved
AUTHOR BIOGRAPHY
PREFACE
First and foremost, Praise to Allah for giving me strength to accomplish my research and thesis. All Your blessing motivated my self to achieve my bachelor degree. This bachelor thesis which is entitled “DNA Fragmentation on HCT-116 Colon Cancer Cell in the Presence of Short Chain Fatty Acid (SCFA) Extract”, was based on research conduced from September 2011 to Maret 2012. The author would like to thank:
1. My father Mulyono, my mother Pariyah, and my brother Achmad Kurniawan. Thank you for all your love, support, and pray.
2. Prof. Dr. Ir. Maggy Thenawidjaja Suhartono as my academic supervisor. Thenk you for her enormous help both academically and financially throughout the completion of my thesis. I am eternally grateful both her inspiring advice, warm support and her encouragement. 3. Dr. Endang Yuli Purwani for providing all my research requirements, all her guidances, and
also as my thesis examiner.
4. Dr. Nancy Dewi Yuliana as my thesis examiner, whose critism and suggestion would help me accomplish my thesis.
5. Silmi Mariya, S.Si, Uus Saepuloh, S.Si, M.Biomed, and Ms. Sela for their guidances in the laboratory. I am grateful for their helpful suggestions and advices.
6. Lecturers of Department of Food Science and Technology for the knowledge that enable me to do this research.
7. My lovely nephew: Ayesha Ahmad, Kayysah Ahmad, and Yumna Kinasih.
8. Desy Ayu as my partner in research. Thanks for the times that we have spent together for disscussion, culinary, and etc.
9. My beloved “TACOS” member ITP45.
10. Al-Fattaah Private Tutoring member. Thank you for being my family, supporting me, make me learn more how to manage my self and how to manage people.
11. Member of Freaky: Meutia, Arum, Anita, Nanda, and Icha. Thanks for being my partner in crime, doing crazy and silly things.
12. Staff of UPT Food Science and Technology Department: Bu Novi and Mbak Anie
Bogor, Mei 2012
TABLE OF CONTENT
Page TABLE OF CONTENT ... ix LIST OF FIGURE ... x I. INTRODUCTION ... 1 A. BACKGROUND ... 1 B. OBJECTIVE ... 2 II. LITERATURE REVIEW ... 3 A. RESISTANT STARCH (RS) ... 3 B. SHORT CHAIN FATTY ACID (SCFA) ... 3 C. APOPTOSIS ... 5 D. DNA LADDER ASSAY ... 8 III. MATERIALS AND METHODS ... 10
A. MATERIALS AND EQUIPMENTS ... 10 1. MATERIALS ... 10 2. EQUIPMENTS ... 10 B. METHODS ... 10 1. Cell Culture ... 10 2. Incubation with SCFA ... 11 3. DNA Extraction (Qiagen DNA Extraction Kit) ... 11 4. DNA Ladder Assay ... 11 IV. RESULTS AND DISSCUSSION ... 12 A. INHIBITION TO HCT-116 COLON CANCER CELL ... 12 B. DETECTION OF APOPTOSIS USING DNA LADDER ASSAY ... 13 V. CONCLUSION AND RECOMMENDATION ... 17 A. CONCLUSION ... 17 B. RECOMMENDATION ... 17 REFERENCES ... 18
LIST OF FIGURE
[image:30.595.120.514.65.814.2]Page Figure 1. Illustration of the morphological features of apoptosis ... 6 Figure 2. The two main apoptotic signalling pathways ... 7 Figure 3. Morphological change of untreated or treated HCT-116 with cell free supernatant
contain SCFA ... 12 Figure 4 DNA fragmentation of untreated and treated HCT-116 colon cancer cells 14
1
I.
INTRODUCTION
A.
BACKGROUND
Sweet potato (Ipomoea batatas) is an alternative food rich in starch. In Indonesia, economical value of sweet potato is low, compared with other starch sources. However, the sweet potatoes which are rich in starch made it a potential source of resistant starch. At present, various studies about resistant starch had been conducted. Recent studies showed that resistant starch consumption may reduce colorectal cancer risk.
Resistant starch has been defined as total amount of starch, and the products of starch degradation that resists digestion in the small intestine of healthy people. There are four types of resistant starch. Type 1 resistant starch (RS1) is starch that escapes digestion in the small intestine due to physical protection by the food matrix, such as grains. Type 2 resistant starch (RS2) is raw starch granules (ungelatinized) with compact structure which limits accessibility of digestive enzymes. Type 3 resistant starch (RS3) is retrogaded starch in which parts of the starch chains can crystallize into components that are less digestible. Most often, this occurs by cooking and cooling starch-containing foods. And the last, type 4 resistant starch (RS4) is not found naturally in foods. Starch that has been chemically modified to introduce bonds that are not digestible by human enzymes.
Starches that resist small intestinal breakdown are fermented by the resident bacteria in the
large intestine, producing a variety of end products, the most significant of which is the short chain
fatty acids (SCFA). SCFA, especially butyrate maintenance of colonic health and barrier function, butyrate has drawn most attention as this fatty acid is the major energy source for the colonocytes (Roediger 1990). Furthermore, butyrate has been shown to have anti-carcinogenic effects mainly by affecting proliferation, differentiation and apoptosis of colonocytes (Scheppach, et. al 1995).
There are several bacterial species which produce butyric acid. It is an anaerobic process. The genera Clostridium, Butyribacterium, and Butyrivibrio are the mostly studied microorganism. For commercial purposes, Clostridium species are preferred for butyric acid production. C. butyricum, C. tyobutyricum, C. thermobutyricum, C. beijerinckii, and C. populeti are of interest for the production of butyrate. Purwani, et. al (2012) observed that RS3 derived from sago and rice starch could be well utilized as substrate by Clostridium butyricum, and revealed that good proportion of acetate: propionat: butyrate in that fermentation.
Apoptosis occurs normally during development and aging and as a homeostatic mechanism to maintain cell populations in tissues and the cell is an active participant in its own demise (“cellular suicide”). Apoptosis is a form of programmed cell death characterized by cytoplasmic condensation, plasma membrane blebbing and nuclear pycnosis, leading to nuclear DNA breakdown into multiples of ~200 bp oligonucleosomal size fragments (Chen and Ioannou 1996).
2
in the appearance of “DNA laddering” when the DNA is analyzed by agarose gel electrophoresis (Arends, et. al 1990). Principle of this assay is apoptotic DNA binds quickly to glass fiber fleece in the presence of a chaotropic salt, guanidine hydrochloride (guanidine HCl). After cellular impurities are washed off the fleece, the DNA is released from the fleece with a low salt buffer (Wyllie, et. al 1998).
B.
OBJECTIVE
3
II.
LITERATURE REVIEW
A.
RESISTANT STARCH (RS)
Resistant starch (RS) has been defined as total amount of starch, and the products of starch degradation that resists digestion in the small intestine of healthy people then enter the large bowel. It occurs for various reasons including chemical structure, cooking of food, chemical modification, and food mastication (Topping and Clifton 2001).There are four types of resistant starch. Starch found in plant cell walls that is inaccessible to amylase activity is one type of resistant starch, designated type 1 resistant starch (RS1). Food sources of RS1 include partially milled grains and seeds. Resistant starch also may be formed during food processing. Ungelatinized granules of starch are typically resistant to enzymatic digestion and are designated type 2 resistant starch (RS2). This type of starch can be found in potatoes and unripe (green) bananas. Cooking and cooling starchy foods by moist heat or extrude starchy foods, for example, generates retrogade starch called type 3 resistant starch (RS3). Chemical modifications of starch, such as formation of starch esters, or cross-bonded starches, also result in resistant starch, called type 4 resistant starch (RS4). RS3 and RS4 may be partially fermented by colonic bacteria (Gropper, et. al 2009). Interestingly, if RS2 granules are heated to over 100ºC, the granularity is lost and starch is gelatinized or the granules swell thereby increasing the availability of the starch to amylase. However, when the starch cools, there is some recrystallization of the starch. This is called retrogradation, which is resistant to α–amylase hydrolysis (Medeiros and Wildman 2012).
Starch that is trapped within whole plant cells or within the food matrix, and some starch granules that have not been fully gelatinized, are hydrolysed only very slowly by a-amylase and therefore may escape complete digestion in the small intestine. The energy yield to the body from this source is less than that provided if starch is digested and absorbed in the small intestine (Englyst, et. al 1996). The glycemic effects of starch is therefore lowered, which decreases insulin secretion. In addition to glycemic management resistant starches may play role in weight management as well (Medeiros and Wildman 2012).
The material included in the definition of RS reaches the human large intestine and thus becomes a substrate for microbial fermentation. The end-products are H2and CO2, CH4 in about half the population, and short-chain fatty acids. SCFA consist primarily of butyrate, propionate and acetate (Englyst, et. al 1996). SCFA contribute to normal large bowel function and prevent pathology through their actions in the lumen and on the colonic musculature and vasculature, through their metabolism by colonocytes (Topping and Clifton 2001).
B.
SHORT CHAIN FATTY ACID (SCFA)
4
varying amounts depending on the diet and the composition of the intestinal microbiota (Marleen,
et. al 2003). SCFA constitute approximately two-thirds of the colonic anion concentration (70-130 mmol/l), mainly as acetate, propionate, and butyrate (Mortensen and Clausen 1996). Various population data show that SCFA production is in order of acetate > propionate > butyrate in a molar ratio of approximately 60:20:20 or 3:1:1, respectively in the proximal and distal colon (Cummings, et. al 1987; Topping and Clifton 2001).
The major source of this in the human colon is thought to be plant cellwall polysaccharides such as cellulose, pectins, and hemicelluloses, currently referred to in human nutrition as dietary fibre. Starch would also be a suitable substrate if it were to reach the colon in significant quantities. Plant cell-wall polysaccharides are composed of hexose (glucose and galactose), pentose (xylose and arabinose), and uronic acid monomers which are fermented by gut micro-organisms along a variety of pathways. The important feature to remember of this metabolism is that it is anaerobic. Many lines of evidence therefore support the idea that fermentation occurs in the colon but, because of the difficulties in obtaining access to the human large bowel during normal digestion, most of the evidence for fermentation has, of necessity, been indirect (Cummings, et. al 1987).
There are several bacterial species which produce butyric acid. Most butyric acid-producing bacteria form acetic acid in addition to butyric acid as their major fermentation products.
Clostridium butyricum, C. tyobutyricum, C. thermobutyricum, C. beijerinckii, and C. populeti are of interest for the production of butyrate. There are some benefits using Clostridium butyricum.
These bacteria needs simple growth medium, produces high metabolite, and easy to isolate. Therefore, Clostrium butyricum also has butyrogenic activity, produce more butyric acid than acetic acid and propionic acid. C. butyricum had the highest specific growth rates compared with other genera. On the other hand, Clostridium spp. are generally less dominant in the human colon and hence the role of Clostridium in the production of butyrate in humans is unclear (Wang 1999).
SCFA will be absorbed and contribute about 3 kilocalories per gram (Medeiros and Wildman 2012). SCFA are rapidly absorbed and have shown to have distinct bioactivity depending on their chain length. With regard to maintenance of colonic health and barrier function, butyrate has drawn most attention as this fatty acid is the major energy source for the colonocytes (Roedriger 1990). Furthermore, butyrate has been shown to have anti-carcinogenic effects mainly by affecting proliferation, differentiation, and gene expression of colonocytes (Scheppach, et. al 1995). Butyrate concentration didn‟t show a significant correlation with pH or
propionic concentration (McIntyre, et. al 1993). The production of propionate is by two main, but circuitous, routes, firstly, involving fixation of CO2, to form succinate which is subsecquently decarboxylated (the 'decarboxylic acid pathway') or secondlly, from lactate and acrylate (the 'acrylate pathway'). Acetate is usually formed by the oxidative decarboxylation of pyruvate and butyrate by reluction of acetoacetate formed from acetate (Cummings, et. al 1981).
5
1. Cells of the ceco-colonic epithelium that use butyrate as a major substrate for the maintenance of energy producing path-ways;
2. Liver cells that metabolize residual butyrate with propionate used for gluconeogenesis; 50% to 70% of acetate is also taken up by the liver;
3. Muscle cells that generate energy from the oxidation of residual acetate.
In ruminants and other herbivores, SCFA are absorbed and transported via the portal vein to the liver, and the fraction not absorbed is distributed to the other body organs and tissues for metabolism. In herbivores, peripheral venous SCFA concentrations are high due to comparatively low visceral extraction and high rates of absorption into the circulation. However, human peripheral venous blood concentrations are normally low, and only acetate is present in measurable amounts. This profile reflects the lower SCFA production rates and greater visceral extraction in omnivores, meaning that human peripheral venous SCFA are not representative of those in the portal circulation. Human experimentation has been confined largely to fecal measurements, which are also limited as 95% of SCFA are produced and absorbed within the colon (Topping and Clifton 2001).
Purwani, et. al (2012) study revealed that type 3 resistant starch (RS3) derived from sago and rice starch that treated with pullulanase could be well utilized as substrate by Clostridium butyricum and showed good proportion of acetate: propionate: butyrate in the bacterial culture filtrate. The fermentation of resistant starch type 3 (RS3) derived from sago starch treated with pullulanase (RSSP) by C. butyricum produced 17.65 mM of butyrate, and fermentation of RS3 derived from rice starch treated with pullulanase (RSRP) by C. butyricum produced 21.76 mM, higher concentration than fermentation of RSSP. This study implied that difference of starch source resulted difference of SCFA concentration.
C.
APOPTOSIS
6
Figure 1 Illustration of the morphological features of apoptosis (Wyllie, et. al 1998)
Apoptosis is characterised by an initial shrinkage of the cell, which in a tissue means breaking cell-to-cell contacts with neighbours and rounding up. The consequence of this stage tends to be that the volume of the cell becomes smaller, and the cytoplasmic internal membranes, ribosomes, mitochondria, and other cytoplasmic organelles are more concentrated in the cytoplasm, which then consequently looks darker. The organelles remain intact and healthy looking very late into the process of death, suggesting that the cell continues metabolic activity for some considerable time. This indeed can be demonstrated in many cell systems by using various inhibitors of metabolic processes (RNA and protein synthesis inhibitors), which delay the progression of a cell through the apoptotic sequence. Apoptosis characteristically involves single isolated cells and not clusters. The DNA or chromatin material in the nucleus condenses very extreme (Potten and Wilson 2004). Pyknosis is the result of chromatin condensation and this is the most characteristic feature of apoptosis. On histologic examination with hematoxylin and eosin stain, apoptosis involves single cells or small clusters of cells. The apoptotic cell appears as a round or oval mass with dark eosinophilic cytoplasm and dense purple nuclear chromatin fragments. Electron microscopy can better define the subcellular changes. Early during the chromatin condensation phase, the electron-dense nuclear material characteristically aggregates peripherally under the nuclear membrane although there can also be uniformly dense nuclei.
7
do not release their cellular constituents into the surrounding interstitial tissue; (2) they are quickly phagocytosed by surrounding cells thus likely preventing secondary necrosis; and (3) the engulfing cells do not produce anti-inflammatory cytokines (Savill and Fadok 2000; Kurosaka, et. al 2003).
The biochemical hallmark of apoptosis is the fragmentation of the genomic DNA, an irreversible event that commits the cell to die and occurs before changes in plasma membrane permeability (prelytic DNA fragmentation). The fragmentation of chromosomal DNA is a hallmark of apoptosis and may facilitate apoptosis by terminating DNA replication and gene transcription. In many systems, this DNA fragmentation has been shown to result from activation of an endogenous Ca2+ and Mg2+ dependent nuclear endonuclease. This enzyme selectively cleaves DNA at sites located between nucleosomal units (linker DNA) generating mono- and oligonucleosomal DNA fragments (Arends et. al, 1990; Wyllie, et. al 1998).
[image:37.595.101.508.49.842.2]In priciple, there are two main apoptotic pathways: the extrinsic or death receptor pathway and the intrinsic or mitochondrial pathway. In both pathways, cysteine aspartyl-specific proteases (caspases) are activated that cleave cellular substrates, and leads to the biochemical and morphological changes that are characteristic of apoptosis. However, there is now evidence that the two pathways are linked and that molecules in one pathway can influence the other (Igney and Krammer 2002). Figure 2 shows the two main apoptotic signalling pathways.
8
Apoptotic cells exhibit several biochemical modifications such as protein cleavage, protein cross-linking, DNA breakdown, and phagocytic recognition that together result in the distinctive structural pathology described previously (Hengartner 2000). Caspases are widely expressed in an inactive proenzyme form in most cells and once activated can often activate other procaspases, allowing initiation of a protease cascade. Some procaspases can also aggregate and autoactivate. This proteolytic cascade, in which one caspase can activate other caspases, amplifies the apoptotic signaling pathway and thus leads to rapid cell death (Elmore 2007).
Although DNA fragmentation into oligonucleosomal ladders is characteristic of apoptosis, recent evidence indicates that not all cells undergo such extensive DNA fragmentation (Cohen, et. al1. 1992 in Chen and Ioannou 1996). In fact, fragmentation of DNA into kilobase-size fragments appears to be an early event in apoptosis, preceding the complete digestion of DNA into multiples of nucleosomal size fragments (Cohen and Sun 1994 in Chen and Ioannou 1996).
D.
DNA LADDER ASSAY
One of the most easily measured features of apoptotic cells is the break-up of the genomic DNA by cellular nucleases. These DNA fragments can be extracted from apoptotic cells and result in the appearance of “DNA laddering” when the DNA is analyzed by agarose gel electrophoresis (Arends, Morris, and Wyllie 1990). Principle of this assay is apoptotic DNA binds quickly to glass fiber fleece in the presence of a chaotropic salt, guanidine hydrochloride (guanidine HCl). After cellular impurities are washed off the fleece, the DNA is released from the fleece with a low salt buffer (Wyllie, et. al 1998).
DNA ladder is asolution of DNA molecules of different lengths used in agarose gel electrophoresis. it is applied to an agarose gel as a reference to estimate the size of unknown DNA molecules. In addition it can be used to approximate the mass off a band by comparison to a special mass ladder. Different DNA ladders are commercially available depending on expected DNA length. The 1 kb ladder with fragment ranging from about 0.5 kbp to 10 or 12 kbp and the 100 bp ladder with fragment ranging from 100 bp to just above 1000 bp are the most frequent. DNA ladders are often produced by a suitable restriction digest of plasmid.
DNA electrophoresis is an analytical technique used to separate DNA fragments by size. An electric field forces the fragments to migrate through a gel. DNA molecules normally migrate from negative to positive potential due to the net negative potential due to the net negative charge of the phosphate backbone of the DNA chain. At the scale of the length of DNA molecules, the gel looks much like a random, intricate network. Longer molecules migrate more slowly because they are more easily „trapped‟ in the network. After the separation is completed, the fractions of DNA fragments of different length are often visualized using a fluorescent dye spesific for DNA, such as ethidium bromide. The gel shows bands corresponding to different DNA molecules populations with different molecular weight. Fragment size is usually reported in “nucleotides”, “base pairs” or
9
separated. Fragment size determination is typically done by comparison to commercially available DNA ladders containing linear DNA fragments of known length.
The types of gel most commonly used for DNA electrophoresis are agarose (for relatively long DNA molecules) and polyacrylamide (for high resolution of short DNA molecules, for example in DNA sequencing). Electrophoresis techniques used in the assessment of DNA damage include alkaline gel electrophoresis and pulsed field gel elctrophoresis. The measurement and analysis are mostly done with a specialized gel analysis software.
10
III.
MATERIALS AND METHODS
A.
MATERIALS AND EQUIPMENTS
1. MATERIALS
The sample used for this research was the fermentation product of type III resistant starch (RS3) derived from Ipomoea batatas var. Jago provided by Indonesian Center of Agricultural Post Harvest Research and Development, Bogor in the form of cell free supernatant consisted of 52.95 mM acetic acid, 66.93 mM propionic acid, and 92.41 mM butyric acid. HCT-116 colon cancer cells was provided by Primate Research Center, Bogor Agricultural University. Materials used in this research were Dulbecco‟s Modified Eagle‟s Medium (DMEM) Gipco USA, Fetal Bovine Serum (FBS) Gipco USA, antibiotics (penicillin and streptomycin) Invitrogen USA, Phosphate Buffered Saline (PBS) Gipco USA, Trypsin 0.25% Difco USA, Qiagen DNA extraction kit (lysis buffer, Aw 1, Aw 2, and AE buffer), agarose powder Promega, ethidium bromide Gipco USA, gel loading buffer, double-distilled water, and TAE buffer 50x Gipco USA (0.484 gram Tris-Base, 0.1142 mL acetic acid, and 0.2 mL 0.5M EDTA pH 8.0).
2. EQUIPMENT
The equipments used in this research were biosafety cabinet, dissposable gloves, dissposable pipettes (1 mL, 2 mL, 5 mL, 10 mL, 25 mL, and 50 mL), micropipette, T25 and T75 tissue culture flask, incubator (37oC, 5% CO2), refrigerator, water-bath, inverted microscope, centrifuge, centrifuge tube, glass bottle, gel doc, cuvette, and DNA electrophoresis.
B.
METHODS
1. Cell Culture
HCT-116 colon cancer cells were warmed up from cryogenic state using waterbath with 37 ºC. Then, the cells were pelleted using centrifuge 750 g and media was discarded. HCT-116 colon cancer cells have been cultured in Dulbecco‟s Modified Eagle‟s Medium (DMEM) supplemented with 10% Fetal Bovine Serum (FBS) and 1% antibiotics (penicillin 100 unit/mL; streptomycin 100µg/mL). Cells (1 x 105) were seeded into T25 tissue culture flask. The cells were incubated at 37oC in 5% CO2 incubator.
11
hemocytometer to seed the cell into flasks in an acurate amount. 1 × 105 cells were seeded into T25 flask every subculture for maintaining.
2. Incubation with SCFA
Analysis was done using eight T25 flasks which contained 1 x 105 cells as the starting cell number. The flasks was incubated for 2 x 24 hr or to reach 50% confluency. Next, incubation was continued for another 4 x 24 hr in the absence (control) or presence of various concentration of cell free supernatant contained SCFA (0.625 mM, 1.00 mM, and 1.25 mM butyrate in the cell free supernatant). For the treatment of the experimental cultures, the cell free supernatant contained SCFA was diluted with DMEM. At the end of 4 x 24 hr incubation period, the cells were observed using inverted microscope with magnification (4x10). Then, the medium contained of dead cells was collected into centrifuge tube and the attached cells were washed with phosphate buffered saline (PBS). The medium contained of dead cells that floating in the medium and made the medium became turbid. After that, the attached cells were trypsinized with trypsin 0.25% to free the cells from adhesion to the flask followed by incubation in 37 ºC for 5-10 minutes and pipetted into centrifuge tube. Next, the attached cells in the trypsin and the medium were pelleted with centrifugation 750 g for 5 minutes. After that, the supernatant was discarded. The pellet were resuspended using 10 mL of PBS steril and centrifuged as before. Then, the supernatant was discarded. The pellet were resuspended using 1 mL PBS steril, then pipetted into eppendorf. Next, attached and dead cells were centrifuged 1000 g for 5 menit. The supernatant was discarded. And the last, the cuvettes were stored at -20oC.
3. DNA Extraction (Qiagen DNA Extraction Kit)
The cells pellet were treated for 10 minutes with 200 μl lysis buffer. Then, the sample was mixed with ethanol 95% and pipetted into filter tube (silica gel). The filter tube was centrifuged 8,000 g for 1 minute and the flow through was discarded. After that, the treated and untreated cells were washed with wash buffer (Aw 1) and centrifuged as before. Next, the treated and untreated cells were washed with wash buffer (Aw 2) and centrifuged 8,000 rpm for 3 minutes. Then, the sample was mixed with 100 μL elution buffer (AE buffer) and centrifuged 8,000 rpm for 1 minute.
4. DNA Ladder Assay
12
IV.
RESULTS AND DISCUSSION
A.
INHIBITION TO HCT-116 COLON CANCER CELL
After 4 x 24 hr incubation in the presence and absence of the cell free supernatant contained SCFA, observation was conducted using inverted microscope with magnification (4x10). Cellular debris in the media could be observed on HCT-116 cells that were treated with SCFA containing 0.625, 1.00, and 1.25 mM butyrate (figure 3). This research showed that the attached cells of control became multi layer. The treated cells with 1.00 mM butyrate showed higher confluency than the treated cells with 1.25 mM butyrate, but lower than the treated cells with 0.625 mM butyrate. This results implied that higher concentration of butyrate showed higher inhibition to HCT-116 colon cancer cell. The inhibition effect was implied at butyrate concentration as low as 0.625 mM and was lower than those reported by Purwani, et. al (2010) that the inhibition effect was observed at butyrate concentration as low as 2.6 mM. SCFA derived from fermentation of RS3 of sago decreased the viability, the total cell number, and increased the inhibition growth of HCT-116 colon cancer cell in a dose dependent manner (p>0.05).
Other research concluded that butyrate inhibited Caco-2 cell proliferation at concentration as low as 0.1 mM (Ruemmele, et. al 1999). This research implied inhibition at higher concentration of butyrate. This difference might caused by cancer cell difference that was used in each research, therefore difference in effect may occur.
(1)
(2)
(3)
(4)
13
B.
DETECTION OF APOPTOSIS USING DNA LADDER ASSAY
DNA fragmentation of HCT-116 colon cancer cell was observed with DNA ladder assay using electrophoresis gel. Observartion data showed different DNA bands between control and cells treated with SCFA (figure 4). DNA bands from attached cells treated with SCFA containing 0.625 mM butyrate was not significantly different from the control. However, the attached cells treated with SCFA containing 1.00 and 1.25 mM butyrate had DNA bands that were significantly different from the control: appeared as longer smear.
This research also observed DNA fragmentation of the floating cells in the medium. There were different result of DNA bands of the floating cells in the control medium and in the treated medium. There were thin DNA bands in the floating cells in the treated medium (SCFA containing 0.625, 1.00, and 1.25 mM butyrate) at ±100 bp (compared with commercial ladder), lower density than normal DNA bands. These bands were not presence in the control. This result showed that DNA of floating cells in the treated medium was fragmented and had lower density than control, the bands observed at 180-200 bp.
The appearance of the nucleosomal DNA ladder in agarose gels has become the hallmark of programmed cell death via apoptosis. This research implied that SCFA derived from fermentation of type-3 resistant starch of Ipomoea batatas induced apoptosis via DNA fragmentation. DNA fragmentation was not clear in the pellet of attached cells, but was observed as a separate bands in the pellet of floating cells in the medium. Floating cells in the medium contained dead cells. This imply that SCFA induced apoptosis at very early stage in the attached cells and later stage in the floating cells in the medium