USWATUN CHASANAH
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN
Dengan ini saya menyatakan bahwa tesis yang berjudul:
“Pemanfaatan Rizobakteri Pemacu Pertumbuhan Tanaman (RPPT) untuk Menginduksi Resistensi Sistemik Mentimun terhadap Zucchini Yellow Mosaic Potyvirus (ZYMV)
Adalah benar merupakan hasil karya saya sendiri dan belum pernah
dipublikasikan. Semua sumber data dan informasi yang digunakan telah
dinyatakan secara jelas dan dapat diperiksa kebenarannya.
Bogor, Agustus 2007
Uswatun Chasanah
ABSTRAK
USWATUN CHASANAH. Pemanfaatan Rizobakteri Pemacu Pertumbuhan Tanaman (RPPT) untuk Menginduksi Resistensi Sistemik Mentimun terhadap Zucchini Yellow Mosaic Potyvirus (ZYMV). Dibimbing oleh WIDODO dan ENDANG NURHAYATI.
Penelitian bertujuan untuk menyeleksi isolat bakteri yang fluoresen (BF) dan bakteri tahan panas (BTP) yang berpotensi sebagai RPPT, serta menguji kemampuan isolat RPPT untuk menginduksi resistensi pada tanaman mentimun terhadap infeksi ZYMV. Penelitian ini dilaksanakan di laboratorium bakteriologi dan virologi Departemen Proteksi Tanaman IPB, serta instalasi rumah kaca Balai Penelitian Tanah di Sindangbarang. Isolasi bakteri dilakukan dari tiga lokasi pertanaman mentimun yaitu Ciherang, Sindangbarang dan Darmaga. Isolat BF dan BTP yang diperoleh kemudian diuji kemampuannya sebagai RPPT dengan metode penyiraman benih dengan suspensi bakteri. Isolat bakteri yang mampu meningkatkan pertumbuhan perakaran (panjang akar dan jumlah akar lateral) akan digunakan sebagai kandidat RPPT. Lima isolat RPPT terbaik digunakan dalam uji induksi resistensi tanaman mentimun terhadap ZYMV yaitu BTP1P, BTP3G, BTP3M, BTP3O, dan BF2F.
© Hak cipta milik IPB, tahun 2007 Hak cipta dilindungi
Dilarang mengutip dan memperbanyak tanpa izin tertulis dari Institut Pertanian Bogor, sebagian atau seluruhnya dalam bentuk apa pun, baik cetak, foto kopi,
PEMANFAATAN
RIZOBAKTERI PEMACU PERTUMBUHAN TANAMAN (RPPT)
UNTUK MENGINDUKSI RESISTENSI SISTEMIK MENTIMUN
TERHADAP ZUCCHINI YELLOW MOSAIC POTYVIRUS (ZYMV)
USWATUN CHASANAH
Tesis
Sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada
Program Studi Entomologi/Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Pemanfaatan Rizobakteri Pemacu Pertumbuhan (RPPT) untuk Menginduksi Resistensi Sistemik Mentimun terhadap Zucchini Yellow Mosaic Potyvirus (ZYMV)
Nama : Uswatun Chasanah
NRP : P.08500008
Program Studi : Entomologi/Fitopatologi
Disetujui
Komisi Pembimbing
Dr. Ir. Widodo, MS Dr. Ir. Endang Nurhayati, MS Ketua Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana IPB
Dr. Ir. Sri Hendrastuti Hidayat, M.Sc Prof. Dr. Ir. Khairil Anwar Notodipuro, MS
PRAKATA
Alhamdulillah, penulis memanjatkan puji dan syukur kepada Allah SWT
karena rahmat dan hidayah-Nya sehingga tesis dengan judul “Pemanfaatan
Rizobakteri Pemacu Pertumbuhan Tanaman (RPPT) untuk Menginduksi
Resistensi Sistemik Mentimun terhadap Zucchini Yellow
Mosaic Potyvirus (ZYMV)” dapat diselesaikan.
Pada kesempatan ini penulis menyampaikan penghargaan dan terima kasih
kepada :
1. Dr. Ir. Widodo, MS dan Dr. Ir. Endang Nurhayati, MS sebagai ketua dan
anggota komisi Pembimbing atas saran, bimbingan, dan bantuan selama
proses penelitian hingga penulisan tesis ini.
2. Dr. Ir. Budi Tjahjono, MAgr, yang pada awal penelitian turut
membimbing dan mengarahkan penelitian.
3. Dr. Ir. Suryo Wiyono, MSc sebagai penguji luar komisi atas saran dan
arahan untuk perbaikan dalam penulisan tesis ini
4. Ketua Laboratorium Bakteriologi dan Virologi Departemen Proteksi
Tanaman IPB, Ketua Instalasi Rumah Kaca Balai Penelitian Tanah,
beserta segenap teknisi yang telah membantu dan memberi fasilitas
sehingga penulis bisa menyelesaikan penelitian.
5. Segenap Staf Pengajar di lingkungan Sekolah Pascasarjana IPB.
6. Bapak, Ibu, Suami, beserta seluruh keluarga yang telah membantu,
membimbing, dan mendoakan penulis untuk menyelesaikan studi di IPB.
7. Seluruh teman-teman yang senantiasa mengulurkan tali persaudaraan dan
kasih sayang di manapun Anda berada.
Semoga Allah senantiasa memberi rahmat, taufiq, dan hidayah kepada kita semua.
Penulis berharap semoga tesis ini bisa bermanfaat untuk kita semua.
Bogor, Agustus 2007
RIWAYAT HIDUP
Penulis dilahirkan di Rembang pada tanggal 16 Oktober 1976 dari ayah
Asrofi dan ibu Karomah. Penulis merupakan putri pertama dari empat bersaudara.
Penulis menyelesaikan pendidikan SD sampai SMA tahun 1993 di
Rembang, Jawa Tengah. Pada tahun 1998 menyelesaikan pendidikan S1 dari
Jurusan Hama dan Penyakit Tumbuhan Fakultas Pertanian Universitas Jember,
Jawa Timur. Pada tahun 2000 mendapat kesempatan belajar di Program Studi
DAFTAR ISI
Halaman
DAFTAR TABEL ... ix
DAFTAR GAMBAR ... x
PENDAHULUAN Latar Belakang ... 1
Tujuan ... 3
Kegunaan ... 3
TINJAUAN PUSTAKA ... Induksi Resistensi Sistemik melalui RPPT ... 4
Peran RPPT dalam Resistensi Sistemik Terinduksi ... 5
Penyakit Mosaik Kuning Zucchini pada Mentimun ... 8
BAHAN DAN METODE Penyiapan RPPT ... 11
Penyiapan Inokulum ZYMV... 12
Pengujian Induksi Resistensi Mentimun terhadap ZYMV menggunakan RPPT ... 13
HASIL DAN PEMBAHASAN Hasil ... 15
Pembahasan ... 21
KESIMPULAN ... 27
DAFTAR TABEL
Halaman
1. Jumlah bakteri fluoresen (BF) dan tahan panas (BTP) yang
diisolasi dari beberapa lokasi sebagai kandidat RPPT... 15
2. Peningkatan pertumbuhan perakaran mentimun yang diukur dari rata-rata panjang akar dan jumlah akar lateral pada 7
hari setelah tanam (HST) karena perlakuan BF dan BTP ... 16
3. Jumlah tanaman yang bergejala mosaik, kejadian, dan keparahan penyakit dalam uji induksi resistensi mentimun terhadap ZYMV menggunakan RPPT pada pengamatan ke-2
(14 HSI) ... 19
4. Rata-rata panjang batang utama, biomas basah dan kering tanaman mentimun pada uji induksi resisistensi mentimun
terhadap ZYMV menggunakan RPPT ... 20
5. Rata-rata jumlah bunga jantan, betina, dan buah mentimun pada uji induksi resistensi mentimun terhadap ZYMV
DAFTAR GAMBAR
Halaman
1. Gejala penyakit mosaik kuning zucchini pada mentimun
karena infeksi ZYMV ... 17
2. Variasi gejala penyakit mosaic kuning zucchini pada
mentimun karena infeksi ZYMV ... 18
3. Nilai absorban ELISA (NAE) yang terukur dalam uji induksi
resistensi mentimun terhadap ZYMV menggunakan RPPT ... 21
PENDAHULUAN
Latar Belakang
Mentimun merupakan salah satu sayuran buah yang banyak dikonsumsi
segar maupun olahan. Seperti buah Cucurbitaceae lain, kandungan nutrisi
mentimun yang terbesar adalah vitamin dan mineral. Perkiraan kebutuhan buah
Cucurbitaceae secara umum cenderung meningkat dari tahun ke tahun (Zitter et
al. 1998). Di Indonesia, luas pertanaman mentimun mencapai 88.956 ha dengan
rata-rata produksi nasional 3,45 ton/ha. Kemampuan produksi mentimun di
Indonesia masih relatif lebih rendah dibandingkan rata-rata produksi mentimun
negara lain yang mencapai 7 ton/ha (Gunaeni et al. 1999). Salah satu faktor yang
mempengaruhi rendahnya produksi mentimun adalah penyakit tanaman yang
disebabkan oleh virus (Semangun 1993; Gunaeni et al. 1999).
Beberapa virus yang umum menginfeksi pertanaman mentimun adalah
cucumber mosaic virus (CMV), cucumber green mottle virus (CGMV), squash
mosaic virus (SqMV), zucchini yellow mosaic virus (ZYMV), dan watermelon
mosaic virus (WMV). Di antara beberapa virus yang menginfeksi pertanaman
mentimun, ZYMV menyebabkan kerusakan tanaman sejak pertama kali
dilaporkan pada tahun 1981. Hingga saat ini diketahui bahwa penyakit akibat
ZYMV telah tersebar di 22 negara dari lima benua (Zitter et al. 1998). Sutarya
(1991) melaporkan bahwa 83,9% tanaman mentimun dari sentra produksi di Jawa
Barat dan Jawa Tengah terinfeksi oleh ZYMV. Kehilangan hasil akibat infeksi
ZYMV pada beberapa kultivar mentimun yang rentan mencapai 53,4% (Sutarya
& Sumpena 1994).
Pengaruh penyakit ini dapat diatasi dengan pengendalian secara terpadu
dan pengelolaan tanaman sehat. Strategi ini bertujuan untuk memperkecil
kehilangan hasil akibat infeksi pathogen dan menjaga kualitas hasil pertanian
sampai pada tingkat yang dapat diterima konsumen (Shetty & Vasanthi 2000).
Berkaitan dengan hal ini, dikembangkan suatu strategi dan metode yang
dapat diaplikasikan secara terpadu sehingga dapat meningkatkan keefektifan
pengendalian dan mengurangi kerusakan ekologi mikrob. Salah satu strategi
pengendalian yang diharapkan dapat memenuhi tujuan ini dan mengarah pada
pengelolaan tanaman sehat adalah tanaman resisten terinduksi. Induksi resistensi
didefinisikan sebagai proteksi sistemik dengan aplikasi suatu agens penginduksi
pada bagian-bagian tanaman tertentu (Liu et al. 1994). Resistensi tanaman
terhadap infeksi patogen bisa diinduksi oleh mikrob patogen maupun non patogen
(Bergstrom et al. 1982; Tuzun & Kloepper 1994), metabolit yang dihasilkan oleh
mikrob, material tanaman, dan bahan kimia organik maupun anorganik (Wheeler
1975; Gottstein & Kuc 1989; Mucharromah 1999).
Mikrob non patogen yang mampu menginduksi resistensi adalah
rizobakteri pemacu pertumbuhan tanaman (RPPT) atau plant growth-promoting
rhizobacteria (PGPR), yaitu beberapa strain rizobakteri yang mampu merangsang
dan meningkatkan pertumbuhan (Leong et al. 1991; Ryder et al. 1994). Beberapa
strain Pseudomonas kelompok fluoresen dan Bacillus merupakan rizobakteri yang
dapat berperan sebagai RPPT dan sekaligus menginduksi resistensi tanaman
terhadap penyakit. Kedua jenis bakteri ini hidup bebas di lingkungan rizosfer dan
memanfaatkan eksudat dan lisat yang disekresikan oleh akar sebagai nutrisi
(Backman et al. 1994; Tuzun & Kloepper 1994; Steiner & Schoenbeck 1995).
Perlakuan dengan RPPT ini dapat mengakibatkan berbagai macam mekanisme
resistensi secara cepat, yang pada tanaman rentan, potensi resistensi ini laten atau
tidak secara cepat diekspresikan untuk melawan patogen (Kuc 1995).
Beberapa penelitian telah membuktikan bahwa beberapa strain RPPT
mampu menginduksi resistensi tanaman dalam berbagai sistem penyakit,
contohnya induksi resistensi tanaman mentimun terhadap Pseudomonas syringae
pv. lachrymans (Liu et al. 1995a), Fusarium oxysporum f.sp. cucumerinum (Liu et
al. 1995b), Colletotrichum orbiculare (Liu et al. 1995c), dan CMV (Raupach et
Tujuan
Tujuan penelitian ini adalah 1) menyeleksi isolat bakteri yang fluoresen
dan bakteri tahan panas yang berpotensi sebagai RPPT, 2) menguji kemampuan
isolat RPPT untuk menginduksi resistensi sistemik pada tanaman mentimun
terhadap infeksi ZYMV.
Kegunaan
Hasil penelitian dapat dimanfaatkan sebagai acuan untuk pengembangan
penelitian serta aplikasi resistensi terinduksi pada tanaman Cucurbitaceae
terhadap infeksi patogen lain dalam spektrum yang lebih luas.
TINJAUAN PUSTAKA
Induksi Resistensi Sistemik melalui RPPT Resistensi Sistemik Terinduksi
Resistensi terinduksi merupakan salah satu tipe resistensi tanaman
terhadap infeksi patogen (Heath 1995). Resistensi terinduksi adalah suatu keadaan
atau kondisi terjadinya peningkatan kemampuan pertahanan yang dikembangkan
oleh tanaman jika terstimulasi secara khusus. Dalam hal ini terjadi pengaktifan
mekanisme resistensi laten, yang diekspresikan jika ada infeksi patogen
selanjutnya (Kuc 1995). Tipe ini merupakan tipe resistensi yang tidak khusus
terhadap patogen tertentu. Satu aplikasi agens penginduksi dapat menyebabkan
tanaman menjadi lebih resisten terhadap berbagai macam patogen. Tipe ini juga
tidak memerlukan gen-gen resistensi khusus untuk patogen tertentu (Heath 1995).
Resistensi terinduksi dapat dipicu oleh bahan-bahan kimia tertentu, mikrob
non patogen, patogen avirulen, ras patogen inkompatibel, atau oleh patogen
virulen yang infeksinya gagal karena kondisi lingkungan yang tidak mendukung.
pertahanan tanaman meningkat di seluruh bagian tanaman. Berdasarkan sifat
tersebut, resistensi terinduksi sering disebut sebagai systemic acquired resistance
(SAR) atau induced systemic resistance (ISR) (Liu et al. 1994; Tuzun & Kloepper
1994; Sticher et al. 1997). Kemampuan tanaman untuk mengekspresikan SAR
dengan spektrum yang lebih luas telah diketahui dengan baik (Ryals et al. 1996,
diacu dalam Pieterse et al. 1999), sedangkan ISR yang diinduksi oleh rizobakteri
non patogenik relatif belum diketahui secara terperinci (van Loon et al. 1998).
Mekanisme Resistensi Sistemik Terinduksi
Secara umum, aplikasi agens penginduksi menyebabkan tanaman menjadi
sensitif sehingga dapat merespon serangan pathogen secara cepat. Perlakuan
dengan agens penginduksi (misalnya RPPT) dapat mengakibatkan berbagai
macam mekanisme resistensi secara cepat, sedangkan pada tanaman rentan,
potensi resistensi ini laten atau tidak secara cepat diekspresikan untuk melawan
pathogen. Penurunan efek patologis dapat dianggap sebagai bukti adanya
resistensi terinduksi. Mekanisme ini meliputi akumulasi fitoaleksin, pembentukan
lapisan pelindung seperti lignin, kalose, dan hydroxyproline-rich glycoproteins
(Hammerschmidt et al. 1984, diacu dalam Tuzun & Kloepper 1994).
Dalam ISR juga terjadi peningkatan aktivitas enzim dalam lintasan
produksi metabolit tertentu dan peningkatan jumlah produksi gen primer seperti
kitinase, β1,3 glukanase, peroksidase, dan/atau protein pathogenesis related (PR)
(Maurhofer et al. 1994; Park & Kloepper 1994). Sintesis protein-protein ini
tampaknya diregulasi pada tingkat mRNA (Park & Kloepper 2000). Kitinase dan
β1,3 glukanase secara biologis aktif terhadap fungi melalui hidrolisis polimer dinding sel, sedangkan peroksidase dan protein PR berasosiasi dengan reaksi
hipersensitif tanaman dan/atau menginduksi resistensi terhadap virus (Wheeler
Peran RPPT dalam Resistensi Sistemik Terinduksi RPPT sebagai Agens Penginduksi Resistensi
Bakteri rizosfer ada dalam jumlah yang besar pada permukaan akar,
memanfaatkan eksudat dan lisat yang disekresikan oleh akar sebagai nutrisi
(Lynch & Whipps 1991). Strain-strain bakteri tersebut ada yang disebut sebagai
Rizobakteri pemacu pertumbuhan tanaman (RPPT) atau plant growth promoting
rhizobacteria (PGPR) karena kemampuannya untuk merangsang dan
meningkatkan pertumbuhan tanaman (Leong et al. 1991; Ryder et al. 1994).
Pseudomonas yang fluoresen dan Bacillus merupakan contoh RPPT yang efektif
untuk menekan penyakit. Bakteri ini mempunyai kebutuhan nutrisi sederhana dan
mampu memetabolisme berbagai macam komponen bahan organik (Leong et al.
1991; Steiner & Schoenbeck 1995).
Pseudomonas fluoresen yang dapat menginduksi resistensi antara lain
adalah P. aeruginosa, P. fluorescens, P. putida, dan P. syringae. Bakteri-bakteri
ini mampu memproduksi dan menyekresikan zat pigmen berwarna hijau
kekuningan, fluoresen, dan larut air. Zat ini mempunyai afinitas terhadap Fe dan
berfungsi sebagai agens transport Fe (siderofor) (Leong et al. 1991).
Pseudomonas juga memproduksi enzim pelisis, metabolit sekunder lain yaitu
antibiotik, fitotoksin, dan hormon pertumbuhan (Leong et al. 1991; Dunne et al.
1996; Bender et al. 1999). Kelompok Bacillus yang dapat menginduksi resistensi
antara lain adalah B. subtilis, B. thuringiensis, dan B. pumilus (Tuzun & Kloepper
1995).
Prosedur umum aplikasi RPPT untuk induksi resistensi adalah dengan cara
penyiraman atau pencampuran tanah dengan suspensi bakteri, pencelupan akar
bibit pada suspensi bakteri pada saat transplantasi, atau perlakuan benih dengan
bakteri sebelum penanaman (Tuzun & Kloepper 1994).
Penelitian-penelitian yang telah dilakukan menunjukkan bahwa induksi
resistensi dapat terjadi pada kultivar-kultivar tanaman rentan pada 25 spesies
tanaman yang meliputi Graminae, Cucurbitaceae, Leguminoceae, Solanaceae,
dikendalikan melalui induksi resistensi oleh RPPT cukup luas, meliputi penyakit
disebabkan oleh bakteri, cendawan, dan virus (Ryder et al. 1994; Hoffland et al.
1996; Raupach et al. 1996, van Loon et al. 1998).
Berbagai penelitian mengenai pengaruh RPPT pada tanaman inang
menunjukkan 1) RPPT mempunyai pengaruh yang menguntungkan pada tanaman
melalui berbagai mekanisme, 2) kemampuan meningkatkan pertumbuhan (growth
promoting) dan pengendalian hayati sering diekspresikan oleh strain RPPT yang
sama, 3) beberapa metabolit bakteri berasosiasi dengan zat–zat metabolit yang
berperan dalam pengendalian hayati. Zat-zat metabolit ini antara lain,
siderophore, hidrogen cyanida (HCN), antibiotik, enzim pelisis, dan faktor-faktor
antifungal. Zat metabolit ini tidak termasuk dalam mekanisme resistensi
terinduksi (Tuzun & Kloepper 1994).
Faktor Determinan Bakteri dalam Resistensi Sistemik Terinduksi
Beberapa strain Pseudomonas memicu SAR melalui produksi asam
salisilat (AS) pada permukaan akar. Strain-strain ini memproduksi AS sebagai
metabolit sekunder. AS ini merupakan siderofor prekursor pyochelin, hormon
tanaman, dan sebagai sinyal transduksi dalam SAR. AS diproduksi maksimal pada
kondisi lingkungan yang kekurangan Fe (Leong et al. 1991; Dunne et al. 1996).
Pada beberapa kasus, RPPT memicu lintasan sinyal lain yang tidak memerlukan
AS (van Loon et al. 1998; Pieterse 1999). Faktor determinan lain yang dapat
mememicu resistensi terinduksi adalah lipopolisakarida dan siderofor (van Loon
et al. 1998).
1. Lipopolisakarida
Lipopolisakarida (LPS) membran luar beberapa strain Pseudomonas
mampu menginduksi resistensi sistemik, dibuktikan pada induksi resistensi
tanaman anyelir terhadap layu fusarium oleh P. fluorescens WCS417. Ekstrak
dinding sel atau LPS murni P. fluorescens WCS374 atau WCS417 atau LPS
murni yang mengandung lipid A-inner core-O-antigenik side chain (OA) mampu
berperan dalam pertumbuhan dan bertahannya (survival) bakteri pada tanaman
dengan cara membantu kolonisasi, pengaturan lingkungan mikro yang sesuai,
sebagai barier terhadap komponen-komponen pertahanan tanaman, dan modulasi
reaksi inang. Inokulasi tanaman tembakau dengan LPS dari sejumlah bakteri dapat
mencegah reaksi hipersensitif terhadap bakteri inkompatibel dan menurunkan
tingkat keparahan gejala pada interaksi kompatibel. Namun, mekanisme
penginduksian resistensi sistemik masih belum diketahui. OA pada LPS
merupakan faktor determinan dalam ISR jika kondisi Fe cukup (van Loon et al.
1998).
2. Siderofor
Pada kondisi rendah Fe, faktor determinan lain yang menginduksi
resistensi sistemik adalah siderofor. Sebagai contoh, P. fluorescens WCS374
memproduksi siderofor berupa pseudobactin-pyoverdin yang dapat menginduksi
resistensi. Beberapa bakteri penginduksi resistensi dapat memproduksi AS sebagai
sebagai suatu siderofor pada kondisi Fe terbatas. Strain WCS374 memproduksi
AS 50 µg/ml dan strain WCS417 memproduksi AS 10µg/ml dalam medium
suksinat standar (SSM) (Leeman et al. 1996). Jika gen yang mengatur
pembentukan AS ditransfer ke strain WCS358 Sid -, mutan ini mampu memproduksi suatu siderofor yang fluoresen biru kehijauan di bawah sinar UV.
Siderofor ini dinamakan fluorebactin (Mercado-Blanco et al. 1997, diacu dalam
van Loon et al. 1998). Sekresi AS yang cukup tinggi dalam SSM oleh strain
WCS374 merupakan suatu artefak, karena ketiadaan substrat yang dibutuhkan
untuk produksi fluorebactin. Fluorebactin ini lebih merupakan faktor penginduksi
resistensi daripada AS (van Loon et al. 1998).
Suatu siderofor lain juga dapat menginduksi ISR, misalnya pseudobactin
358 yang diproduksi oleh strain WCS358 (Leong et al. 1991). Pseudobactin 358
ini dapat berperan sebagai penginduksi ISR pada Arabidopsis. Cara siderofor ini
3. Asam Salisilat (AS)
Eksperimen dengan P. aeruginosa 7NSK2 sebagai penginduksi dan
tobacco mosaic virus (TMV) sebagai challenger pada tanaman tembakau dan
Botrytis cinerea (challenger) pada tanaman buncis menunjukkan bahwa bakteri
memproduksi AS untuk memicu ISR (van Loon et al. 1998). Di bawah kondisi Fe
terbatas, strain 7NSK2 memproduksi tiga siderofor, yaitu pyoverdine, pyochelin,
dan AS. Peran pyochelin dalam ISR tidak dapat diabaikan karena AS sebagai
prekursor pyochelin (De Meyer & Hofte 1997).
Pada lobak, ISR oleh strain WCS374 dan strain WCS417 berasosiasi
dengan kemampuan bakteri untuk memproduksi AS (Leeman et al. 1996), tetapi
keterlibatan AS dalam induksi masih harus dibuktikan (van Loon et al. 1998).
P. fluorescens CHAO juga memproduksi AS untuk induksi resistensi pada
tembakau (Maurhofer et al. 1994).
Hal-hal tersebut menunjukkan bahwa strain-strain rizobakteri menginduksi
resistensi melalui mekanisme yang berbeda. Sebagai contoh, strain-strain
menginduksi ISR melalui mekanisme konstitutif, produksi AS, atau memicu
resistensi lebih lanjut melalui produksi AS pada kondisi Fe terbatas.
Penyakit Mosaik Kuning Zucchini pada Mentimun Gejala
Infeksi virus penyebab penyakit mosaik kuning zucchini pada mentimun
dapat terjadi pada semua fase pertumbuhan tanaman, yaitu fase pembibitan,
pertumbuhan vegetatif, pembungaan, dan pembuahan. Hampir semua bagian
tanaman yaitu daun, batang, dan buah juga dapat terinfeksi oleh virus ini (CABI
1999).
Gejala penyakit mosaik kuning pada tanaman mentimun adalah mosaik,
belang, kerdil, klorosis, dan distorsi pada daun, mosaik dan distorsi pada buah
(Lisa & Lecoq 1984; Purciful et al. 1984; Al-Shahwan et al. 1995). Gejala pada
tanaman mentimun rentan adalah klorosis pada daun, mosaik hijau pucat,
(blister), dan pertumbuhan tanaman terhambat. Pada tanaman muda, gejala yang
tampak adalah tulang daun berkelok-kelok dan tepi daun bergerigi (Sutarya &
Duriat 1995). Gunaeni et al. 1999 juga melaporkan bahwa gejala tampak sangat
nyata pada tanaman mentimun muda, sedangkan pada tanaman yang agak tua,
gejala tampak lebih ringan atau berupa mosaik lemah.
Agens Penyebab Penyakit
Penyakit ini disebabkan oleh zucchini yellow mosaic potyvirus (ZYMV).
Partikel virus berbentuk benang panjang, berukuran 750 – 800 nm, yang
mempunyai untai RNA tunggal posistif. Isolat ZYMV Singapura berukuran 800 x
11 nm. RNA mempunyai berat molekul 2,93 x 106 Da, dan protein selubung berukuran 36 KDa. Tiap protein subunit dalam protein selubung mengandung 279
asam amino (Lisa & Lecoq 1984; CABI 1999). Virus ini masih infekif dalam sap
zucchini setelah pemanasan 10 menit pada temperatur 55 oC tetapi tidak pada 60 o
C, virus ini juga masih infektif setelah pengenceran sampai 10-4 dengan air suling tetapi tidak infektif pada pengenceran 10-5, dan masih infektif setelah 3 hari penyimpanan pada temperatur ruang (Lisa & Lecoq 1984).
Virus ini mempunyai beberapa strain yang dapat menginduksi gejala
berbeda. Ada satu strain yang menginduksi nekrotik dan reaksi layu pada kultivar
melon yang mempunyai gen semi dominan Fn. Selanjutnya ada 22 isolat virus
yang dikelompokkan ke dalam tiga patotipe berdasarkan reaksi tanaman melon
galur PI414723 yang resisten terhadap beberapa isolat virus. Ada juga strain dari
Amerika Serikat (AS) yang menyebabkan gejala yang sama pada tanaman
labu-labuan dengan gejala yang disebabkan oleh watermelon mosaic virus (WMV)
strain 1 pada tanaman labu-labuan. Strain dari Perancis menginduksi gejala yang
sama dengan yang diinduksi oleh WMV strain 2 pada tanaman melon.
Strain-strain ini juga mempunyai perbedaan penularan oleh kutu daun. Beberapa peneliti
juga melaporkan adanya isolat California, Connecticut, Florida, Mesir, pulau
Penularan dan Kisaran Inang Virus
ZYMV ditularkan oleh sejumlah kutu daun secara non persisten, meliputi
Acyrthosiphon pisum, A. kondoi, Aphis craccivora, A. citricola, A. gossypii, A.
middletonii, A. spiraecola, Macrosiphum euphorbiae, Myzus persicae, Toxoptera
aurantii, dan Uroleucon ambrosiae. ZYMV juga dapat ditularkan secara mekanik
dengan mudah serta melalui alat-alat pemotong yang telah terkontaminasi virus
tersebut. ZYMV dapat menginfeksi spesies liar Melothria perdula sebagai inang
alternatif yang dapat menjadi sumber inokulum penting di pertanaman
Cucurbitaceae. Selain menginfeksi tanaman inang Cucurbitaceae, ZYMV juga
dapat menginfeksi tanaman dari famili Aizoaceae, Amaranthaceae, Apiaceae,
Asteraceae, Chenopodiaceae, Fabaeae, Lamiaceae, Ranunculaceae,
Scrophulariaceae, dan Solanaceae (Zitter et al. 1998; CABI 1999)
BAHAN DAN METODE
Penelitian dilaksanakan di laboratorium Bakteriologi Tumbuhan dan
Virologi Tumbuhan Departemen Proteksi Tanaman IPB dan di instalasi rumah
kaca Balai Penelitian Tanah Departemen Pertanian di Sindangbarang.
Penyiapan RPPT a. Isolasi kandidat RPPT dari rizosfer
Pelaksanaan isolasi kandidat RPPT dilakukan di laboratorium Bakteriologi
Tumbuhan Departemen Proteksi Tanaman IPB. Kandidat RPPT diisolasi dari tiga
lokasi yaitu pertanaman mentimun di Ciherang dan Sindangbarang dan
pertanaman mentimun yang ditanam dalam pot di Darmaga. Bakteri diisolasi dari
rizosfer tanaman mentimun serta gulma disekitarnya. Tanah yang menempel
dipermukaan perakaran dirontokkan, kemudian perakaran dikocok dalam air steril
dengan menggunakan rotary shaker (Chambel et al. 1994). Suspensi yang
Isolasi bakteri fluoresen dilakukan dengan medium King’s B yang
ditambah antibiotik sikloheksimid dengan konsentrasi akhir dalam medium 100
ppm, ampisilin 50 ppm, dan kloramfenikol 12,5 ppm. Suspensi tanah diencerkan
hingga 10-7, agar setelah ditumbuhkan pada medium dapat diperoleh koloni-koloni tunggal. Suspensi pada pengenceran 10-7 – 10-9 diambil 0,1 ml, diteteskan dan ditebarkan secara merata pada medium, kemudian diinkubasikan pada
temperatur kamar selama 48 jam. Koloni yang fluoresen di bawah sinar NUV
selanjutnya diisolasi (Widodo et al. 1993; Chambel et al. 1994). Isolat bakteri
fluoresen yang diperoleh diberi kode BF (bakteri fluoresen) dan akan diuji
kemampuannya sebagai RPPT.
Isolasi bakteri tahan panas dilakukan dengan menggunakan medium
tryptic soy agar (TSA) 10%. Suspensi tanah dipanaskan dalam penangas air pada
temperatur 80 oC selama 30 menit, kemudian ditunggu hingga dingin. Suspensi diambil 0,1 ml, diteteskan dan ditebarkan secara merata pada medium, kemudian
diinkubasikan pada temperatur kamar selama 48 jam. Koloni yang tumbuh
diisolasi dan diberi kode BTP (bakteri tahan panas), dan akan diuji
kemampuannya sebagai RPPT (Kim et al. 1997).
b. Evaluasi Potensi RPPT
Pengujian isolat bakteri yang berpotensi sebagai RPPT dilakukan menurut
metode Chambel et al. (1994) yang dimodifikasi. Isolat bakteri kandidat PGPR
diperbanyak pada medium TSA dalam cawan pada temperatur kamar selama 24
jam, kemudian disuspensikan dalam air steril dengan kerapatan 108-1010 CFU/ml. Benih mentimun disterilisasi dengan larutan natrium hipoklorit 1% selama
1 menit, kemudian dibilas dengan akuades. Benih tersebut kemudian ditanam
pada nampan pembibitan yang bersisi campuran tanah, pasir, dan pupuk kandang
steril dengan perbandingan 2:1:1. Tiap benih ditetesi dengan 1 ml suspensi
bakteri. Perlakuan kontrol ditetesi dengan air steril. Tiap perlakuan isolat bakteri
dan kontrol diulang 12 kali. Pembibitan tersebut dipelihara selama 7 hari pada
temperatur kamar di bawah sinar matahari langsung. Kemudian tanaman dicabut
dengan hati-hati agar tidak merusak perakarannya. Panjang akar dan jumlah akar
jumlah akar lateral mentimun yang diberi perlakuan RPPT dibandingkan
perlakuan kontrol. Lima isolat RPPT terbaik dalam peningkatkan pertumbuhan
perakaran digunakan dalam pengujian induksi resistensi.
Penyiapan Inokulum ZYMV
Penyiapan inokulum dilakukan di laboratorium Virologi Tumbuhan
Departemen Proteksi Tanaman IPB. Isolat ZYMV yang digunakan adalah koleksi
laboratorium Virologi Tumbuhan Departemen Proteksi Tanaman IPB. Isolat
diperbanyak pada tanaman kabocha (Cucurbita moschata) yang diinokulasikan
secara mekanik. Daun tanaman sumber inokulum digerus menggunakan mortar
steril dalam larutan penyangga fosfat 0,01 M pH 7 dengan perbandingan 1g daun
terinfeksi dalam 5 ml larutan penyangga (1:5 b/v). Sap ini segera diinokulasikan
dengan cara dioles menggunakan kapas steril pada kotiledon dan daun primer
(primary leaf ) kabocha umur 5 hari setelah tanam (HST) yang telah ditaburi
serbuk karborundum 600 mesh. Kotiledon dan daun kabocha yang telah
diinokulasi disemprot dengan air secara hati-hati untuk mencuci sisa-sisa sap.
Tanaman dipelihara dan diamati tiap hari untuk mengetahui perkembangan gejala
dan masa inkubasinya. Daun yang bergejala sistemik dipanen pada 14 hari setelah
inokulasi (HSI) dan digunakan sebagai sumber inokulum.
Pengujian Induksi Resistensi Mentimun terhadap ZYMV melalui RPPT Pengujian kemampuan RPPT untuk menginduksi resistensi mentimun
terhadap ZYMV dilakukan di Instalasi Rumah Kaca Balai Penelitian Tanah di
Sindangbarang. Percobaan disusun menurut rancangan acak lengkap (RAL)
dengan perlakuan lima isolat RPPT terbaik dari hasil seleksi RPPT yaitu BTP1P,
BTP3G, BTP3M, BTP3O, BF2F, kontrol sehat (tanpa aplikasi RPPT maupun
inokulasi ZYMV), dan kontrol sakit (hanya diinokulasi ZYMV). Masing-masing
perlakuan diulang 15 kali. Percobaan dilakukan sampai masa berbuah yang
optimal yang mana tanaman mampu berbuah dengan kualitas yang baik (buah
Benih mentimun disterilisasi dengan natrium hipoklorit 1% selama
1 menit, kemudian dibilas dengan air steril. Benih ditanam dalam pot berdiameter
35 cm yang berisi campuran tanah dan pupuk kandang yang tidak distrerilisasi
dengan perbandingan 2:1. Tiap pot ditanami 3 butir benih.
Pada perlakuan isolat RPPT, tiap benih mentimun ditetesi dengan 1 ml
suspensi bakteri dengan kerapatan 108-1010 CFU/ml, kemudian ditutup dengan tanah. Benih untuk perlakuan kontrol sehat dan kontrol sakit ditetesi dengan air
steril. Pada 9 hari setelah tanam (HST), kotiledon tanaman mentimun dan daun
primer (primary leaf) yang telah muncul (untuk perlakuan RPPT dan kontrol
sakit) diinokulasi secara mekanik seperti dijelaskan pada penyiapan inokulum
ZYMV. Tanaman diamati tiap hari untuk diketahui perkembangan gejala dan
masa inkubasinya. Kejadian dan keparahan penyakit diamati dan dicatat pada 7-28
HSI dengan interval 1 minggu (Raupach et al. 1996). Kejadian dan keparahan
penyakit ditentukan dengan rumus (Dolores & Valdez 1988 diacu dalam Gunaeni
et al. 1999) :
Kejadian Penyakit = [ Jumlah tanaman sakit / Jumlah tanaman uji] x 100%
Keparahan Penyakit = [ ∑ ( n x v)] / [ N x V] x 100%
Keterangan :
n : Jumlah tanaman dalam tiap kategori keparahan v : Nilai skala tiap kategori serangan
N: Jumlah tanaman uji
V: Nilai skala keparahan tertinggi
Nilai sklala keparahan :
0 = Tanaman tidak menunjukkan gejala
1 = Tanaman menunjukkan gejala mosaik ringan
2 = Tanaman menunjukkan gejala mosaik sedang tanpa malformasi 3 = Tanaman menunjukkan gejala mosaik disertai malformasi
Jumlah bunga jantan dan bunga betina dicatat selama 2 minggu sejak awal
pembungaan dengan interval 3 hari. Jumlah buah dicatat sejak awal masa berbuah
sampai tanaman tidak bisa menghasilkan buah secara baik (dilihat dari bentuk
buah dan kemampuan berbuah). Pada akhir masa tanam, tanaman dicabut dengan
dikeringkan untuk mengetahui biomas keringnya. Data dari parameter jumlah
bunga, buah, panjang batang, biomas basah dan kering dianalisis dengan One-Way
Analisis of Variance (AOV) dengan program Statistix 8.
Konsentrasi virus dalam tanaman diukur dengan uji indirect ELISA
(enzyme-linked immunosorbent assay) (Al Shahwan et al. 1995; Raupach et al.
1996). Pengambilan sampel dilakukan dua kali, pertama pada waktu tanaman
berumur 20 HST atau 11 HSI, kedua pada waktu tanaman sudah berbunga yaitu
berumur 29 HST atau 20 HSI. Suatu perangkat komersial ELISA (dari Agdia Inc.)
digunakan untuk mendeteksi ZYMV. Pengujian sampel I menggunakan antiserum
potyvirus sedangkan sampel II menggunakan antiserum ZYMV. Prosedur
pelaksanaan ELISA disesuaikan dengan petunjuk dari Agdia Inc. Absorban
tiap-tiap sampel dibaca pada ELISA reader (Biorad model 550) pada panjang
gelombang 405 nm.
HASIL DAN PEMBAHASAN
Hasil Isolasi Kandidat RPPT dari Rizosfer
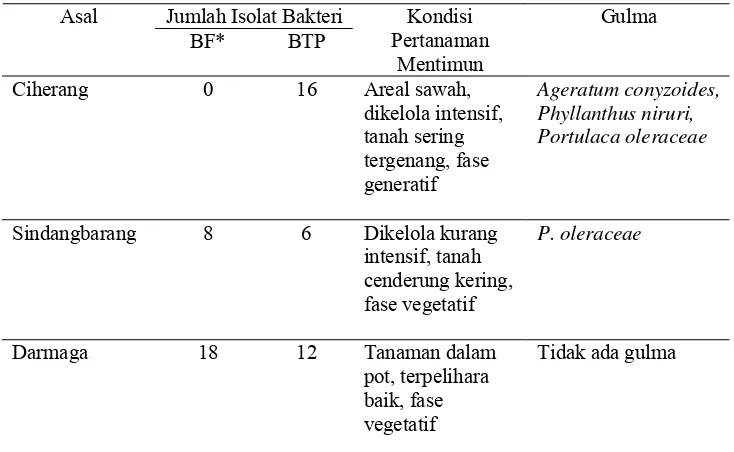
Kandidat RPPT diisolasi dari tiga lokasi yaitu Ciherang, Sindangbarang,
dan Darmaga. Tabel 1 menampilkan asal, jumlah isolat BF dan BTP, kondisi
pertanaman mentimun, serta gulma yang menyertai di pertanaman mentimun.
Hasil isolasi kandidat RPPT dari beberapa lokasi menunjukkan perbedaan jumlah
dan jenis isolat bakteri. Isolat BF hanya diperoleh dari Sindangbarang dan
Darmaga, sedangkan BTP dapat diperoleh Ciherang, Sindangbarang, dan
Tabel 1. Jumlah bakteri fluoresen (BF) dan tahan panas (BTP) yang diisolasi dari beberapa lokasi sebagai kandidat RPPT
Jumlah Isolat Bakteri
Sindangbarang 8 6 Dikelola kurang
intensif, tanah
Ket : * : isolasi BF dilakukan dari suspensi tanah pada pengenceran 10-7 - 10-9
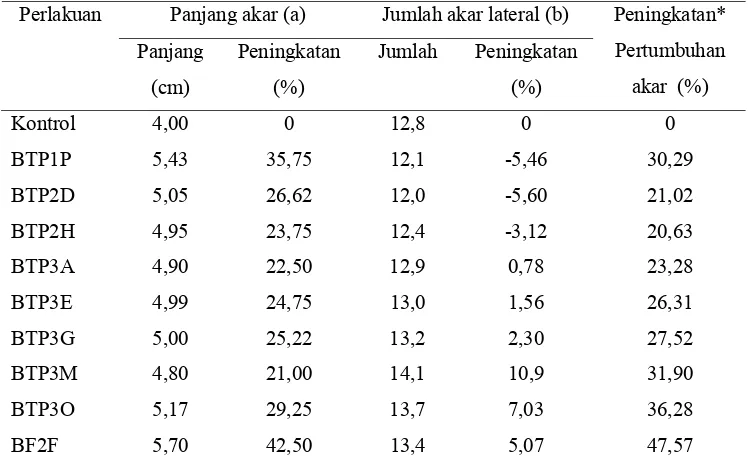
Seleksi RPPT didasarkan pada kemampuan bakteri tersebut untuk
meningkatkan pertumbuhan tanaman yang diukur pada awal pertumbuhan dengan
parameter panjang akar dan jumlah akar lateral (Chambel et al. 1994). Secara
keseluruhan diperoleh 60 isolat BTP dan BF yang diisolasi dari Ciherang,
Sindangbarang dan Darmaga, yang kemudian diuji kemampuannya sebagai RPPT
(Tabel 1). Tabel 2 menampilkan isolat BTP dan BF yang mampu meningkatkan
Tabel 2. Peningkatan pertumbuhan perakaran tanaman mentimun yang diukur dari rata-rata panjang akar dan jumlah akar lateral 7 hari setelah tanam (HST) karena perlakuan isolat BF dan BTP
Panjang akar (a) Jumlah akar lateral (b) Perlakuan
Ket.: * : Peningkatan pertumbuhan akar dihitung dari jumlah peningkatan panjang akar dan jumlah akar lateral dibandingkan kontrol
Selanjutnya isolat yang digunakan dalam pengujian induksi resistensi
adalah lima isolat terbaik dalam peningkatan pertumbuhan perakaran, yaitu
BTP1P, BTP3G, BTP3M, BTP3O, dan BF2F.
Induksi Resistensi Mentimun terhadap ZYMV menggunakan RPPT 1. Gejala
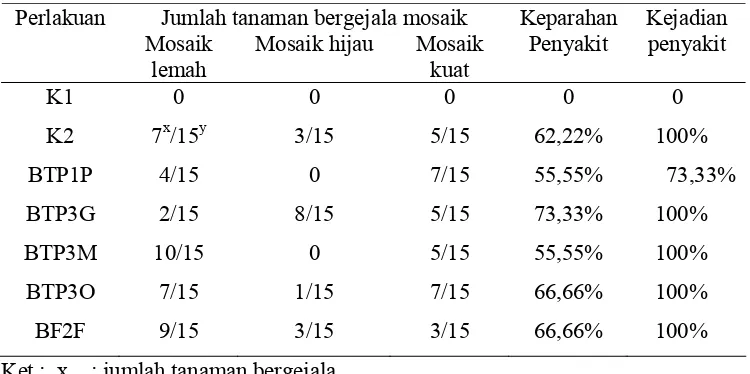
Hasil pengujian menunjukkan bahwa hampir semua tanaman mentimun
yang terinfeksi ZYMV dan diberi RPPT masih menunjukkan gejala mosaik yang
bervariasi (Tabel 3). Kejadian penyakit perlakuan kontrol sakit maupun perlakuan
isolat RPPT mencapai 100%, kecuali pada perlakuan BTP1P yaitu 73,33%.
Keparahan penyakit karena infeksi ZYMV pada semua perlakuan kontrol dan
RPPT mencapai 55,55% - 73,33%.
Gejala muncul antara 6-7 hari setelah inokulasi (HSI) pada semua
tulang daun pada primary leaf yang diinokulasi menjadi pucat, kekuningan yang
terlihat kontras dengan lamina daun. Pada 9-10 HSI, gejala mulai muncul pada
daun pertama, selanjutnya diikuti dengan daun yang tumbuh berikutnya. Pada
10-11 HSI gejala terlihat bervariasi di antara tanaman. Variasi gejala ini tidak
tergantung pada perlakuan RPPT.
A B
C D
Gambar 1. Gejala penyakit mosaik kuning zucchini pada mentimun karena infeksi ZYMV; A. daun sehat, B. klorosis tulang daun pada bergejala, C. klorosis tulang daun pada daun yang lebih tua, D. klorosis tulang daun berkembang ke lamina daun nampak mosaik hijau kuning
Berdasarkan variasi gejala yang ada, gejala penyakit akibat infeksi ZYMV
pada mentimun bisa dikelompokkan sebagai mosaik lemah, mosaik hijau dan
mosaik kuat (Gambar 2).
1) Mosaik lemah dicirikan dengan adanya klorosis tulang daun, berwarna
2) Mosaik sedang dicirikan dengan adanya klorosis tulang daun, berwarna
kuning atau hijau pucat, mulai menyebar di lamina daun, tampak mosaik
hijau-kuning, atau hijau-pucat disertai penebalan daun (daun terasa lebih
kaku), tanpa disertai malformasi;
3) Mosaik kuat dicirikan dengan klorosis tulang daun, adanya mosaik
hijau-kuning, disertai penebalan daun, melepuh, dan malformasi daun.
A B
C D
Tabel 3. Jumlah tanaman yang bergejala mosaik dan kejadian penyakit dalam uji induksi resistensi mentimun terhadap ZYMV menggunakan RPPT pada pengamatan ke-2 (14 HSI.)
Gejala penyakit juga muncul pada bunga tetapi hanya pada bunga betina
berupa mosaik hijau pada mahkota bunga serta malformasi, tetapi tidak berlanjut
sampai ke buah.
2. Perkembangan Penyakit dan Karakteristik Pertumbuhan Tanaman Sejak gejala muncul pada 6 – 7 HSI, gejala terus berkembang pada
daun-daun yang muncul selanjutnya. Mulai 10 HSI, tanaman yang menunjukkan gejala
mosaik kuat dan disertai malformasi daun tidak dapat melakukan pemulihan
(recovery), sehingga pertumbuhan tanaman terganggu dan menjadi kerdil.
Tanaman yang menunjukkan gejala mosaik lemah dan mosaik hijau dapat
melakukan pemulihan sehingga tanaman tetap tumbuh dan berkembang. Pada
semua perlakuan RPPT maupun kontrol sakit, pemulihan dapat dilakukan oleh
tanaman yang bergejala mosaik lemah dan mosaik hijau. Ada perbedaan respon
tanaman sakit terhadap infeksi virus karena perlakuan RPPT. Hal ini terlihat pada
parameter pertumbuhan tanaman seperti biomas tanaman kering dan basah, serta
Tabel 4. Rata-rata panjang batang utama, biomas basah dan kering tanaman mentimun pada uji induksi resistensi mentimun terhadap ZYMV menggunakan RPPT
Ket: *: Perbedaan huruf pada kolom yang sama menunjukkan beda nyata berdasarkan uji Tukey pada taraf nyata 5%
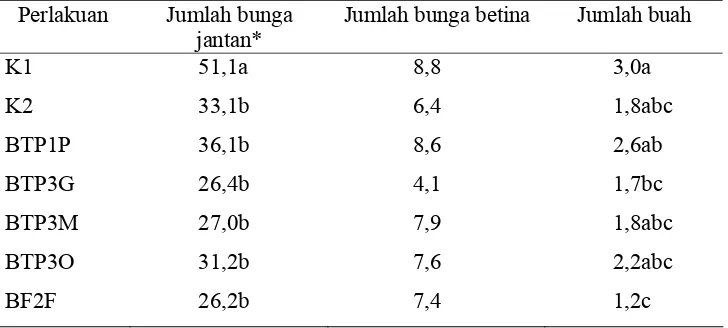
Tidak ada perbedaan rata-rata panjang batang maupun jumlah bunga
karena perlakuan RPPT. Tanaman yang diberi perlakuan RPPT dan perlakuan
kontrol dapat tumbuh dan berkembang sama, kecuali pada parameter jumlah
bunga jantan yang mana jumlah bunga pada K1 berbeda dengan perlakuan yang
lain. (Tabel 5).
Tabel 5. Rata-rata jumlah bunga jantan, bunga betina dan buah mentimun pada uji induksi resistensi mentimun terhadap ZYMV menggunakan RPPT Perlakuan Jumlah bunga
jantan*
Jumlah bunga betina Jumlah buah
K1 51,1a 8,8 3,0a
Berkaitan dengan perkembangan penyakit dan kemampuan tanaman sakit
melakukan pemulihan, konsentrasi virus yang terukur melalui uji ELISA (Gambar
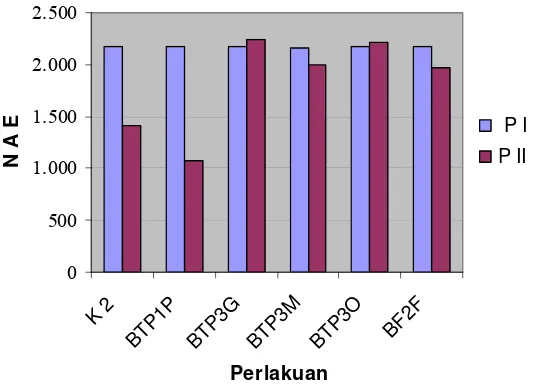
3) dapat digunakan sebagai pembanding. Secara umum terjadi penurunan
konsentrasi virus pada pengukuran ke-2, kecuali pada perlakuan BTP3G dan
BTP3O.
Gambar 3. Nilai absorban ELISA (NAE) yang terukur dalam uji induksi resistensi mentimun terhadap ZYMV menggunakan RPPT. P I: pengukuran ke-1 pada 11 HSI, P II: pengukuran ke-2 pada 20 HSI.
Pembahasan
BF dan BTP dapat diisolasi dari pertanaman mentimun di beberapa lokasi
dengan jumlah dan jenis bakteri yang berbeda. Parke (1991) dan Legocki (1996)
menyatakan bahwa kondisi tanah dan status nutrisi tanaman dapat mempengaruhi
keberadaan bakteri rizosfer. Jika pertumbuhan tanaman baik maka tanaman dapat
mensekresikan eksudat dan lisat yang menjadi sumber nutrisi bagi bakteri rizosfer
(Lynch & Whipp 1991; Nelson 2004).
Kelompok BF tidak diperoleh dari lingkungan pertanaman mentimun di
Ciherang yang kondisi tanahnya cenderung basah dan tergenang. Pada kondisi ini
suspensi perakaran 10-7 – 10-9, BF belum dapat diisolasi. Dalam kondisi sering tergenang, BTP mampu bertahan dalam bentuk endospora. Perlakuan pemanasan
pada saat isolasi dapat menyeleksi endospora BTP (Kim et al. 1997).
Kemungkinan lain yang mempengaruhi rendahnya populasi BF adalah fase
pertumbuhan tanaman mentimun. Pada saat isolasi, tanaman sedang dalam fase
generatif (dalam keadaan berbunga dan berbuah). Seperti yang dilaporkan oleh
Mew et al. (1994), bahwa kepadatan populasi kelompok Pseudomonas lebih
rendah dibandingkan kelompok Bacillus di fase pertumbuhan generatif pada
tanaman padi.
Seleksi BF dan BTP sebagai RPPT berdasarkan pada kemampuan bakteri
tersebut untuk meningkatkan pertumbuhan tanaman yang diukur pada awal
pertumbuhan dengan parameter panjang akar dan jumlah akar lateral (Chambel et
al. 1994). Mekanisme peningkatan pertumbuhan ini dapat dapat melalui beberapa
cara. Germida dan Freitas (1994) menyatakan bahwa mekanisme utama
peningkatan pertumbuhan adalah mekanisme penyerapan nutrisi oleh tanaman.
Keberadaan RPPT menyebabkan tanaman mampu menyerap nutrisi makro
maupun mikro lebih baik. Dalam penelitian lain disebutkan bahwa mekanisme
peningkatan pertumbuhan berkaitan dengan kemampuan RPPT dalam
memproduksi hormon pertumbuhan, siderophore yang dapat mengikat ion Fe dan
membuatnya tersedia bagi perakaran tanaman, serta kemampuan melarutkan
mineral P (Cattelan et al. 1999; Nelson 2004). Produksi hormon auksin dan
1-aminocyclopropane-1-carboxylate (ACC) deaminase (penghambat
pembentukan etilen) oleh RPPT dapat merangsang pertumbuhan sistem perakaran
(Loveless 1991; Srinivasan et al. 1997). Suatu siderofor misalnya pseudobactin
358 (PSB358) yang di produksi oleh P. putida WCS 358 dapat mengikat ion Fe
menjadi Ferric Pseudobactin (FePSB358) yang berguna untuk stimulasi sintesis
klorofil tanaman (Duiff et al. 1994). Kemampuan melarutkan fosfat juga menjadi
keunggulan beberapa isolat RPPT. Senyawa fosfat yang dibutuhkan tanaman tidak
selalu dalam bentuk tersedia dalam lingkungan pertumbuhan tanaman. Isolat
RPPT yang mampu melarutkan fosfat dapat membantu penyediaan fosfat bagi
meningkatkan pertumbuhan tanaman dan akhirnya dapat memacu pelepasan
eksudat tanaman untuk memenuhi kebutuhan RPPT (Kluepfel 1993).
Keberhasilan induksi resistensi oleh RPPT dapat ditentukan dari masa
inkubasi, gejala, kejadian dan keparahan penyakit (Raupach et al. 1996).
Pengujian isolat RPPT untuk menginduksi nresistensi mentimun terhadap ZYMV
menunjukkan bahwa tidak ada perbedaan gejala infeksi ZYMV pada tanaman uji
karena perlakuan beberapa isolat RPPT. Gejala penyakit juga muncul dalam
waktu yang relatif sama (6-7 HSI). Perbedaan gejala berupa mosaik lemah,
sedang, dan kuat beserta variasinya terdapat di semua perlakuan yang diinfeksi
ZYMV. Variasi gejala ini umum terjadi pada tanaman mentimun maupun
Cucurbitaceae lain yang terinfeksi ZYMV (Zitter 1998; CABI 1999).
Berdasarkan nilai kejadian dan keparahan penyakit pada semua perlakuan
isolat RPPT, tanaman mentimun uji belum bisa dikategorikan terinduksi
resistensinya. Untuk semua perlakuan RPPT, kejadian penyakit mencapai 73,33%
- 100%, sedangkan keparahan penyakit mencapai 55,55% - 73,33%. Menurut
Dolores & Valdez (1988 diacu dalam Gunaeni et al. 1999), jika keparahan
penyakit lebih dari 50%, tanaman dikategorikan sangat rentan terhadap infeksi
virus.
Secara umum bisa dikatakan bahwa isolat-isolat RPPT yang diuji belum
menginduksi resistensi tanaman mentimun terhadap infeksi ZYMV, tetapi ada
satu isolat yaitu BTP1P yang mampu berfungsi sebagai pemacu pertumbuhan
(growth-promoting) dan menyebabkan tanaman mentimun mampu berbuah lebih
banyak dengan rata-rata 2,6 buah tiap tanaman dibandingkan kontrol sakit yang
hanya berbuah 1,8 tiap tanaman.
Belum adanya induksi resistensi pada tanaman mentimun oleh isolat RPPT
yang diuji diduga karena teknik aplikasi RPPT dengan cara penyiraman suspensi
bakteri pada benih belum tepat. Dengan cara ini, bakteri yang diaplikasikan
mungkin tidak semuanya mampu mengkolonisasi benih mentimun karena dapat
terbawa air pada saat penyiraman. Teknik aplikasi ini dapat menjadi faktor
penentu keberhasilan induksi resistensi (Cadena-Cepeda et al. 2006). Hal ini
mengkolonisasi perakaran tanaman. Isolat RPPT harus mampu mengkolonisasi
perakaran dan memproduksi suatu pemicu atau sinyal untuk menginduksi
resistensi dan kemudian ditranslokasikan secara sistemik sebelum ada infeksi
patogen (challenge) (Kloepper et al. 1992).
Kemungkinan lain penyebab belum adanya induksi resistensi pada
tanaman mentimun adalah karena isolat RPPT yang diuji bukan merupakan agens
penginduksi. van Loon et al. (1997) menyatakan bahwa kemampuan induksi
resistensi oleh suatu strain RPPT ditentukan oleh suatu faktor determinan berupa
asam salisilat, siderofor, atau lipopolisakarida. Sebagai contoh, suatu
lipopolisakarida menjadi determinan P. fluorescens WCS417 untuk menginduksi
resistensi pada tanaman anyelir, lobak, dan Arabidopsis. Jika isolat-isolat RPPT
yang diuji tidak mempunyai faktor determinan untuk memicu induksi resistensi
maka isolat tersebut tidak dapat menginduksi resistensi pada tanaman mentimun.
Selain faktor teknik aplikasi dan karakter penginduksi yang harus dimiliki
oleh rizobakteri, temperatur udara di ruang pemeliharaan tanaman juga menjadi
penyebab gagalnya proses induksi resistensi. Pada saat pengujian induksi
resistensi, temperatur udara di ruang pemeliharaan tanaman tidak stabil karena
ruang tersebut tidak dilengkapi dengan alat pengendali temperatur. Selama masa
percobaan, rata-rata temperatur udara pada siang hari adalah 33 oC – 35 oC, bahkan pernah mencapai 40 oC. Gagalnya sistem pertahanan karena pengaruh temperatur ini pernah dilaporkan oleh Weststeijn (1981 diacu dalam van Loon
2000). Ketika tembakau tahan (mengandung gen N ) diinfeksi Tobaccco Mosaic
Virus (TMV) dan temperatur mencapai 30 oC, virus tidak dapat dihalangi penyebarannya sehingga gejala mosaik sistemik mulai berkembang. Naiknya
temperatur pada suatu waktu setelah inokulasi virus menyebabkan rusaknya cincin
penghalang di sekitar lesio, yang mana cincin penghalang ini berfungsi untuk
menghambat pergerakan virus.
Berkaitan dengan perkembangan penyakit, konsentrasi virus yang terukur
melalui uji ELISA (Gambar 3) dapat digunakan sebagai pembanding. Hasil
pengukuran konsentrasi virus ini memperjelas bahwa perlakuan isolat RPPT
pengukuran ke-1 (11 HSI), semua perlakuan yang terinfeksi virus (kontrol sakit
maupun isolat RPPT) mempunyai nilai absorban yang hampir sama antara 2,160 –
2,175. Ini berarti bahwa tanaman tidak mampu menghambat replikasi virus
sehingga konsentrasi virus pada tanaman yang diberi perlakuan RPPT sama
tingginya dengan konsentrasi virus pada tanaman kontrol sakit. Pada pengukuran
ke-2 (20 HSI), hanya perlakuan BTP1P yang menyebabkan konsentrasi virus
dalam tanaman mentimun lebih rendah dibandingkan kontrol sakit.
Penurunan konsentrasi virus pada pengukuran NAE ke-2 dibandingkan
kontrol mengindikasikan adanya penghambatan multiplikasi virus pada mentimun
yang diberi perlakuan BTP1P. Dengan konsentrasi virus yang lebih rendah,
tanaman mampu melakukan pemulihan sehingga dapat tumbuh dan berbuah lebih
baik dibandingkan kontrol sakit. Kemampuan mentimun untuk tetap berbuah
dengan rata-rata 2,6 buah tiap tanaman dibandingkan kontrol sakit yang hanya
berbuah 1,8 tiap tanaman ini dapat dikatakan sebagai tanaman toleran. Fraser et
al. (1986 diacu dalam Fraser 2000) melaporkan bahwa pada tanaman tomat
heterozigot (Tm-1/+) yang diinfeksi oleh Tomato Mosaic Virus (ToMV),
konsentrasi virus hanya 30% dibandingkan pada tanaman tomat rentan yang
mencapai konsentrasi virus maksimal (100%). Pada tingkat konsentrasi virus ini
(30%), produksi gejalanya lebih ringan dibandingkan tanaman rentan dan
dinyatakan sebagai tanaman toleran. Dalam kasus ini, gen TM-1 pada tomat
mungkin mempunyai pengaruh langsung terhadap ekspresi gejala seperti
pengaruh penghambatan terhadap multiplikasi virus.
Toleransi terhadap infeksi ZYMV ini diduga karena 1) karakter-karakter
pemacu pertumbuhan yang dimiliki oleh BTP1P seperti produksi fitohormon,
produksi siderofor atau pelarutan mineral pertumbuhan seperti yang telah
dinyatakan oleh Nelson (2004); 2) ada penghambatan multiplikasi dan penyebaran
virus dalam tanaman. Adanya kalose atau fargmen dinding sel lain yang dapat
menghalangi plasmodesmata sehingga menghambat pergerakan virus misalnya
pergerakan Tobaccco Mosaic Virus (TMV) dari sel ke sel di daun tembakau
(Shimomura 1979 diacu dalam van Loon 2000). Kalose atau fragmen dinding sel
akibat infeksi patogen (van Loon et al. 1998; Tuzun & Bent 1999). Sebagai
contoh, penebalan dinding sel karena adanya kalose dan material fenolik dapat
diinduksi oleh B. pumilus SE34 pada tanaman kacang polong dan P. fluorescens
WCS417 pada tanaman tomat.
Konsentrasi virus pada tanaman yang diberi isolat BTP3M dan BF2F pada
pengukuran ke-2 turun seperti pada kontrol sakit. Hal ini umum terjadi pada pola
perkembangan virus, yang mana perkembangan gejala dan konsentrasi virus
cenderung menurun sesuai dengan pertambahan umur tanaman (Sutarya & Duriat
1995). Produksi buah pada tanaman yang diberi isolat BTP3M sama dengan
jumlah buah tanaman kontrol sakit. Ada dugaan bahwa isolat BTP3M belum
memberikan pengaruh terhadap proses induksi resistensi. Pada perlakuan BTP3G
dan BF2F, produksi buah mentimun per tanaman justru lebih rendah dibandingkan
kontrol sakit. BTP3G dan BTP3M menyebabkan penurunan biomas kering
tanaman mentimun dibandingkan kontrol sakit. Dugaan mengenai penyebab
turunnya potensi pertumbuhan dan produksi adalah stres hormon asam absisi dan
pelepasan hasil metabolisme sebagai respon terhadap infeksi patogen (van Loon
2000).
KESIMPULAN
Berdasarkan hasil penelitian ini dapat disimpulkan bahwa : BTP dan BF
yang dapat diisolasi dari berbagai lokasi berbeda jumlah dan jenisnya. Di antara
BTP dan BF yang diisolasi terdapat beberapa isolat yang berpotensi sebagai
RPPT. Terdapat satu isolat BTP yaitu BTP1P yang berperan agens pemacu
pertumbuhan pada tanaman mentimun dan menyebabkan tanaman lebih toleran
dengan infeksi ZYMV, tetapi belum dapat menginduksi resistensi sistemik
DAFTAR PUSTAKA
Al-Shahwan IM, Abdalla OA, Al-Saleh MA. 1995. Response of greenhouse-grown cucumber cultivars to an isolate of zucchini yellow mosaic virus (ZYMV). Plant Dis 79:898-901.
Backman PA, Brannen PM, Mahaffe WF. 1994. Plant response and disease control following seed inoculation with Bacillus subtilis. Di dalam: Ryder MH, Stephens, PM, Bowen GD, editor. Improving Plant Productivity with Rhizosphere Bacteria. Proceedings of the Third International Workshop on Plant Growth Promoting Rhizobacteria; Adelaide, South Australia, March 7-11 1994. Adelaide: CSIRO. hlm 3-8.
Bender CL, Rangaswamy V, Loper V. 1999. Polyketide production by plant associated Pseudomonads. Annu Rev Phytopathol 37: 175-196.
Bergstrom GC, Johnson MC, Kuc J. 1982. Effect of local infection of cucumber by Colletotrichum lagenarium, Pseudomonas lachrymans, or tobacco necrosis virus on systemic resistance to cucumber mosaic virus. Phytopathology 72: 922-926.
[CABI]. 1999. Crop Protection Compendium. London: CABI.
Cadena-Cepeda, Burelle NK, Runion GB, Kloepper JW. 2006. Assessing soil microbial populations and enzyme activity following the use of microbial inoculants. International Plant Growth Promoting Rhizobacteria Workshop (Abstract).
http://66.218.69.11/search/cache?ei=UTF-8&p=Rhizobacteria...(16 Juni 2007)
Cattelan AJ, Hartel PG, Furhmann. 1999. Screening for plant growth promoting rhizobacteria to promote early soybean growth. Soil Sci Soc of Am J 63: 1670(Abstrak).
http://soil.scijournals.org/cgi/content/abstract/63/6/1670. (16 Juni 2007)
De Meyer G, Hofte M. 1997. Salicylic acid produced by the rhizobacterium Pseudomonas aeruginosa 7NSK2 induces resistance to leaf infection by Botrytis cinerea on bean. Phytopathology 87: 588-593.
Dunne C, Delanay T, Fenton A, Lohrke S, Moenne-Loccoz Y, O’gara F. 1999. The biotechnology and aplication of Pseudomonas inoculants for the biocontrol of phytopathogens. Di dalam: Stacey G, Mullin B, Gresshoff PM, editor. Biology of Plant-Microbe Interactions. Minnesota: APS Press. hlm 325-331.
Duiff BJ, de Kogel WJ, PAHM Bakker, Schippers B. 1994. Significance of pseudobactin for the iron nutrition of plants. Di dalam: Ryder MH, Stephens, PM, Bowen GD, editor. Improving Plant Productivity with Rhizosphere Bacteria. Proceedings of the Third International Workshop on Plant Growth Promoting Rhizobacteria; Adelaide, South Australia, March 7-11 1994. Adelaide: CSIRO. hlm 142-144.
Fraser RSS. 2000. Special aspects of resistance to viruses. Di dalam Slusarenko AJ, Fraser RSS, van Loon LC, editor. Mechanisms of Resistance to Plant Diseases. Dordrecht: Kluwer Academic Publisher. hlm 479-520.
Germida JJ, de Freitas JR. 1994. Growth promotion of Cabbage, lettuce and onion by fluorescent pseudomonads under growth chamber conditions. Di dalam: Ryder MH, Stephens, PM, Bowen GD, editor. Improving Plant Productivity with Rhizosphere Bacteria. Proceedings of the Third International Workshop on Plant Growth Promoting Rhizobacteria; Adelaide, South Australia, March 7-11 1994. Adelaide: CSIRO. hlm 37-39.
Gottstein HD, Kuc JA. 1989. Induction of systemic resistance to antrachnose in cucumber by phosphates. Phytopathology 79: 176-179.
Gunaeni N, Sumpena U, Kusandriani Y. 1999. Evaluasi plasma nutfah
mentimun terhadap penyakit zucchini yellow mosaic virus (ZYMV). Di dalam: Pros Kongr Nas XV dan Seminar Ilmiah PFI; Purwokerto, 16-18 September 1999. Purwokerto: PFI. hlm 346-354.
Hoffland E, Hakulinen J, Van Pelt JA. 1996. Comparison of systemic resistance induced by avirulent and non-pathogenic Pseudomonas species.
Phytopathology 86: 757-762.
Kim DS, Cook RJ, Weller DM. 1997. Bacillus sp. L324-92 for biological control of three root diseases of wheat grown with reduced tillage. Phytopathology 87: 551-558.
Kloepper JW, Wei G, Tuzun S. 1992. Rhizosphere population dynamics and intenal colonization of cucumber by plant growth-promoting rhizobacteria which induce resistance to Colletotrichum orbiculare. Di dalam: Tjamos ES, editor. Biological control of plant diseases. New Yoek: Plenum Press. Hlm 185-191.
Kluepfel DA. 1993. The behaviour and tracking of bacteria in the rhizosphere. Annu Rev Phytopathol 31: 441-472.
Kuc J. 1995. Induced systemic resistance-an overview. Di dalam:
Hammerschmidt R, Kuc J, editor. Induced Resistance to Disease in Plants. Dordrecht: Kluwer. hlm 169-175.
Leeman M, Den Ouden FM, Van Pelt JA, Dirk FPM, Steijl H. 1996. Iron availablity affects inductions of systemic resistance to Fusarium wilt of radish by Pseudomonas fluorescens. Phytopathology 86: 149-155.
Legocki AB. 1996. Emerging areas and future prospects in the field of plant-microbe interactions. Proc of the 8th International Symposium on Molecular Plant-Microbe Interactions, Tennesse 14-19 July 1996. Minnesota: Int Soc for Mol Plant-Microbe Interactions
Leong J, Bitter W, Koster M, Marugg JD, Weisbeek PJ. 1991. Genetics of iron transport in plant growth promoting Pseudomonas putida WCS358. Di dalam: Kleister DL, Cergan PB, editor. The Rhizosphere and Plant Growth.. Dordrech: Kluwer. hlm 271-278.
Lisa V, Lecoq H. 1984. Zucchini yellow mosaic virus. Description of Plant Viruses. England: CMI/AAB. No 282.
Rhizosphere Bacteria. Proceedings of the Third International Workshop on Plant Growth Promoting Rhizobacteria; Adelaide, South Australia, March 7-11 1994. Adelaide: CSIRO. hlm 44-46.
____________________________. 1995a. Induction of systemic resistance in cucumber against bacterial angular spot by plant-growth promoting rhizobacteria. Phytopathology 85: 843-847.
_____________________________. 1995b. Induction of systemic resistance in cucumber against fusarium wilt by plant-growth promoting rhizobacteria. Phytopathology 85: 695-698.
______________________________. 1995c. Induction of systemic resistance in cucumber by plant-growth promoting rhizobacteria: duration of protection and effect of host resistance on protection and root colonization. Phytopathology 85: 1064-1068.
Loveless AR. 1983. Principles of plant biology for the tropics. Alih bahasa. Kartawinata K, Danimiharja S, Soetisna U. 1991. Prinsip-prinsip biologi tumbuhan untuk daerah tropic. Jakarta: Gramedia Pustaka Utama.
Lynch JM, Whipps JM. 1991. Substrate flow in the rhizosphere. Di dalam: Kleister DL, Cergan PB, editor. The Rhizosphere and Plant Growth.. Dordrech: Kluwer. hlm 15-24.
____________, Hase C, Mewly P, Metraux JP, Defago G. 1994. Induction of systemic resistance of tobacco necrosis virus by root colonizing
Pseudomonas fluorescens strain CHAO: influence of the gac A gene and the pyoverdine production. Phytopathology 84: 139-146.
Mew TW, Rosales AM, Maningas GV. 1994. Biological control of rhizoctonia sheath bliaght and blast of rice. Di dalam: Ryder MH, Stephens, PM, Bowen GD, editor. Improving Plant Productivity with Rhizosphere Bacteria. Proceedings of the Third International Workshop on Plant Growth Promoting Rhizobacteria; Adelaide, South Australia, March 7-11 1994. Adelaide: CSIRO. hlm 9-13.
Mucharromah. 1999. Induksi resistensi sistemik tanaman kedele terhadap penyakit antraknosa: jenis dan konsentrasi agen IRS. Di dalam: Pros Kongr Nas XV dan Seminar Ilmiah PFI; Purwokerto, 16-18 September 1999. Purwokerto: PFI. hlm 241-247.
10-1094/cm-2004-0301-05-RV.
http://www.plantmanagementnetwork.org/pub/cm/review/2004/rhizobacter ia [16 Juni 2007)
Park KS, Kloepper JW. 2000. Activation of PR-1a promoter by rhizobacteria that induce systemic resistance in tobacco against Pseudomonas syringae pv. tabaci. Biol Control 18 : 2 – 9.
Parke JL. 1991. Root colonization by indigenous and introduced microorganisms. Di dalam: Keister DL, Cregan PB, editor. The Rhizosphere and plant growth. Dordrecht: lower. Hlm 33-42.
Pieterse CMJ, Van Wees SCM, Ton J, Leon-Kloosterziel KM, Van Pelt JA, Keurentjes JJB, Knoester M, Van Loon LC. 1999. Rhizobacteria-mediated induced systemic resistance (ISR) in Arabidopsis: involvement of
jasmonate and ethylen. Di dalam: De Wit PJGM, Misseling T, Stiekema WJ, editor. Biology of Plant Microbe Vol. 2. Dordrecht: Kluwer. hlm 291-296.
Purciful DE, Adlerz WC, Simone GW, Hiebert E. Christie SR. 1984. Serological relationships and partial characterization of zucchini yellow mosaic virus isolated from squash in Florida. Plant Dis 68: 230-233.
Raupach GS, Murphy JF, Kloepper JW. 1995. Biological control of cucumber mosaic cucumovirus in Cucumis sativus by PGPR-mediated induced systemic resistance. Phytopathology 85: 1167. (Abstrak).
____________, Liu L, Murphy JF, Tuzun S, Kloepper JW. 1996. Induced systemic resistance in cucumber and tomato against cucumber mosaic cucumovirus using plant-growth promoting rhizobacteria (PGPR). Plant Dis 80: 891-894.
Ryder MH, Stephen PM, Bowen GD. 1994. Improving Plant Productivity with Rhizosfer Bacteria. Adelaide: CSIRO.
Semangun H. 1993. Penyakit-Penyakit Tanaman Hortikultura di Indonesia. Yogyakarta: Gadjah Mada University Press.
Srinivasan M, Petersen DJ, Holl FB. 1997. Altered root hair morphogenesis in Phaseolus vulgaris in respons to bacterial coinoculation and presence of aminoethoexy vinyl glycine (AVG). Microbiol Res 152: 151-156.
Steiner U, Schonbeck F. 1995. Induced resistance in monocots. Di dalam:
Hammerschmidt R, Kuc J, editor. Induced Resistance to Disease in Plants. Dordrecht: Kluwer. hlm 86-110.
Sticher L, Mauch-Mani B, Metraux JP. 1997. Systemic acquired resistance. Annu. Rev. Phytopathol.35: 235-270.
Sutariati GAK. 2006. Perlakuan benih dengan agens biokontrol untuk
pengendalian penyakit antraknosa, peningkatan hasil, dan mutu benih cabai. [Disertasi]. Bogor: Institut Pertanian Bogor, Sekolah Pasca Sarjana.
Sutarya R. 1991. Deteksi penyakit yang disebabkan oleh virus pada tanaman
mentimun. Laporan intern ATA-395.
__________, Sumpena U. 1994. Studi kehilangan hasil yang disebabkan oleh ZYMV (zucchini yellow mosaic virus) pada tiga kultivar mentimun. Bul Penel Hort 27: 170-175.
___________, Duriat AS. 1995. Pengaruh waktu inokulasi ZYMV (Zucchini Yellow Mosaic Virus) terhadap pertumbuhan dan hasil pada tanaman mentimun. Kongr Nas XIII dan Seminar Ilmiah PFI, Mataram 27 – 29 September 1995. Mataram: PFI. Hlm 449-452.
Tuzun S, Kloepper JW. 1994. Induced systemic resistance by plant-growth promoting rhizobacteria. Di dalam: Ryder MH, Stephens, PM, Bowen GD, editor. Improving Plant Productivity with Rhizosphere Bacteria. Proceedings of the Third International Workshop on Plant Growth Promoting Rhizobacteria; Adelaide, South Australia, March 7-11 1994. Adelaide: CSIRO. hlm 104-113.
________________. 1995. Practical aplication and implementation of induced resistance. Di dalam: Hammerschmidt R, Kuc J, editor. Induced Resistance to Disease in Plants. Dordrecht: Kluwer. hlm 152-168.
Biochemistry, Ecology, and Agriculture. Minnesota: APS Press. hlm 95-115.
van Loon LC, Bakker PAHM, Pieterse CMJ. 1997. Mechanisms of PGPR-induced resistance against pathogens. Di dalam: Ogoshi A, Kobayashi K, Homma Y, Kodama F, Kondo N, Akino S, editor. Plant Growth-Promoting Rhizobacteria, Present status and future prospects. Sapporo: hlm 50-57.
_____________________________________. 1998. Systemic resistance induced by rhizosphere bacteria. Annu Rev Phytopathol 36: 453-486.
_____________. 2000. Systemic induced resistance. Di dalam Slusarenko AJ, Fraser RSS, van Loon LC, editor. Mechanisms of Resistance to Plant Diseases. Dordrecht: Kluwer Academic Publisher. hlm 521-574.
Wahyuni WS. 1999. Bagaimana respon tanaman tahan terhadap infeksi virus?. Di dalam: Pros Kongr Nas XV dan Seminar Ilmiah PFI; Purwokerto, 16-18 September 1999. Purwokerto: PFI. hlm 717-720.
Wheeler H. 1975. Plant Pathogenesis. Berlin:Springer-Verlag.