TOLERAN TERHADAP CEKAMAN KEKERINGAN
DAN RESISTEN TERHADAP PENYAKIT BUSUK
BATANG Sclerotium rolfsii
Oleh :
A. Farid Hemon
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ii
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi yang berjudul : “Efektifitas Seleksi In Vitro Berulang untuk Mendapatkan Plasma Nutfah Kacang Tanah yang Toleran terhadap Cekaman Kekeringan dan Resisten terhadap Penyakit Busuk Batang Sclerotium rolfsii” adalah karya saya sendiri dengan arahan Komisi Pembimbing dan belum pernah diajukan dalam bentuk apapun untuk memperoleh gelar program sejenis di perguruan tinggi lain. Sumber informasi yang berasal atau dikut ip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir disertasi ini.
Bogor, Desember 2006
A. Farid Hemon
iii
ABSTRACT
A. FARID HEMON, 2006. Effectiveness of Repeat Cycling In Vitro Selection to Produce Drought Tolerant and Stem Rot Sclerotium rolfsii Resistant Peanut Germplasm. SUDARSONO, RUSMILAH SUSENO, HAJRIAL ASWIDINNOOR, dan WIDODO.
The main limiting factors of peanut cultivation in dry land are water insufficient and S. rolfsii infection. The effort of peanut cultivation management in upland is use of drought tolerant and disease resistant cultivar. Application of somaclonal variation induction and in vitro selection using selective agent is reliable method to be developed to create new peanut line. For this reason, the objectives of this research were to evaluate effectiveness of 1) repeat cycling in vitro selection using medium containing polyethylene glycol (PEG) and culture filtrate of S. rolfsii to obtain drought tolerant and stem rot disease resistant peanut cultivar, respectively, and 2) double in vitro selection in both medium containing PEG and culture filtrate.
The experiment was inisiated with induction of somaclonal variation in MS medium containing picloram 16 µM (MS-P16). Medium of MS-P16 that added PEG 15% as selective agent for drought stress and MS-P16 medium with culture filtrate 30% as selective agent for disease resistance. Embriogenic calli were placed on PEG medium during three mont hs per cycle. PEG insensitive somatic embryos (SE) were proliferated, and this SE named SE Pi-I. Somatic embryos Pi-I were secondly selected on PEG medium (repeat cycling, two cycles, ES Pi-II). In vitro double selection was done with challenge SE Pi-I against culture filtrate medium, and insensitive SE named SE PFi-I. Repeat cycling and double in vitro selection against culture filtrate medium as same as PEG in vitro selection. In vitro selection on culture filtrate medium was gotten insensitive SE one cycle (Fi-I), SE two cycles (repeat cycling selection, Fi-II). Somatic embryos
Fi-I was selected on PEG mediun (double selection, SE FPi-I).
All insensitive selected SEs from repeat cycling and double in vitro selection were germinated to generate plantlets (R0 generation). These plants were harvested to obtain R0:1 seeds. The seeds were grown (R1 plant) to produce R1:2 seeds. These plants of R2 generation were used to somaclonal plant evaluation against PEG stress, water stress, and pathogen infection.
Result of this research showed 1) after repeat cycling in vitro selection, PEG and culture filtrate insensitive SE and its R0 shoot of cv. Kelinci and Singa were respectively more tole rance against PEG and culture filtrate stress, 2) while one identified from double selection were more tolerance to both PEG and culture filtrate medium, 3) peanut plant of R0 and R1 resulted from repeat cycling and double in vitro selection consisted of qualitative and quantitative variants, 4) plant performance produced from repeat cycling in vitro selection to PEG was lesser leaf necrosed symptom and more survive under PEG stress, 5) somaclonal plants regenerated from repeat cycling in vitro selection against PEG were more tolerance to water defisit, produced higher vegetative growth and dry pod yield, and lower dry pod yield reduction, 6) regenerant peanut plants resulted from selected SE two cycles against culture filtrate were more resistance to S. rolfsii
iv Finally, this research has showed that repeat cycling and double in vitro selection more effective to produce drought tolerance and S. rolfsii resistance peanut germplasm.
v
ABSTRAK
A. FARID HEMON, 2006. Efektifitas Seleksi In Vitro Berulang untuk Mendapatkan Plasma Nutfah Kacang Tanah yang Toleran terhadap Cekaman Kekeringan dan Resisten terhadap Penyakit Busuk Batang Sclerotium rolfsii. SUDARSONO, RUSMILAH SUSENO, HAJRIAL ASWIDINNOOR, dan WIDODO.
Kendala utama penanaman kacang tanah di lahan kering adalah masalah kekurangan air dan serangan cendawan S. rolfsii. Upaya pengelolaan budidaya kacang tanah di lahan kering adalah menggunakan kultivar toleran kekeringan dan resisten infeksi S. rolfsii. Penggunaan teknik induksi variasi somaklonal dan seleksi in vitro dengan menggunakan agens penyeleksi merupakan metode yang layak untuk dikembangkan. Seleksi in vitro yang dilakukan hanya satu siklus (tiga bulan), masih banyak ditemukan tanaman somaklon yang peka pada kekeringan dan rentan terhadap penyakit. Untuk itu, penelitian disertasi ini dilakukan dengan tujuan menguji efektifitas (1) seleksi in vitro berulang dalam media yang mengandung polietilena glikol (PEG) atau filtrat kultur S. rolfsii untuk mendapatkan tanaman kacang tanah toleran kekeringan dan resisten terhadap penyakit busuk batang, secara berturut-turut, dan (2) seleksi ganda dalam media yang mengandung PEG dan filtrat kultur untuk mendapatkan kacang tanah yang toleran cekaman kekeringan dan resisten terhadap penyakit. busuk batang S. rolfsii.
Percobaan diawali dengan menginduksi variasi somaklonal dengan media MS yang mengandung pikloram (MS-P16). Media MS-P16 yang ditambah PEG 15% digunakan sebagai agens penyeleksi untuk cekaman kekeringan dan media MS-P16 dengan penambahan filtrat kultur S. rolfsii 30% sebagai agens penyeleksi resistensi terhadap S. rolfsii. Kalus embriogen ditempatkan dalam media PEG selama tiga bulan per siklus. Embrio somatik (ES) yang insensitif pada PEG diproliferasi, dan ES ini disebut ES Pi-I. Kalus embriogen Pi-I diseleksi kembali pada media PEG (seleksi berulang/dua siklus, ES Pi-II). Seleksi ganda dilakukan dengan menempatkan ES Pi-I pada media filtrat kultur dan disebut ES PFi-I. Seleksi in vitro berulang pada media filtrat kultur sama seperti seleksi pada media PEG. Akhir dari seleksi in vitro pada filtrat kultur diperoleh ES insensitif satu siklus (ES Fi-I) dan ES insensitif dua siklus (seleksi berulang, ES Fi-II). Embrio somatik Fi-I diseleksi pada media PEG dan yang insensitif disebut ES FPi-I.
Semua ES insensitif hasil seleksi in vitro berulang dan seleksi ganda dikecambahkan sampai tumbuh planlet (generasi R0). Tanaman ini dipane n untuk mendapatkan benih R0:1. Benih ini ditanam kembali (generasi R1) untuk mendapatkan benih R1:2. Benih R1:2 (tanaman R2) inilah yang digunakan untuk menguji toleransi terhadap larutan PEG pada media sekam, terhadap cekaman kekeringan, dan resistensi terhadap infeksi cendawan.
vi dihasilkan dari seleksi in vitro berulang pada PEG menunjukkan gejala nekrosis pada daun lebih sedikit dan mampu bertahan hidup lebih lama pada media sekam dengan larutan PEG. Tanaman ini juga memberikan pertumbuhan vegetatif akar dan tajuk yang lebih baik dengan tingkat toleransi yang lebih baik terhadap cekaman PEG, (5) tanaman dari hasil seleksi in vitro berulang pada PEG menghasilkan pertumbuhan vegetatif dan hasil polong yang lebih tinggi dengan persentase penurunan hasil polong lebih kecil. Tanaman ini juga mempunyai tingkat toleransi yang lebih baik terhadap cekaman kekeringan, dan (6) tanaman dari hasil regenerasi seleksi ES selama dua siklus pada media filtrat kultur menghasilkan polong lebih tinggi dengan penurunan hasil polong akibat infeksi cendawan lebih rendah. Tanaman ini lebih tahan terhadap serangan S. rolfsi.
Penelitian ini telah menunjukkan bahwa seleksi in vitro berulang dan seleksi ganda cukup efektif untuk mendapatkan plasma nutfah tanaman kacang tanah yang toleran terhadap cekaman kekeringan dan resisten terhadap penyakit busuk batang S. rolfsii.
vii
EFEKTIFITAS SELEKSI IN VITRO BERULANG UNTUK
MENDAPATKAN PLASMA NUTFAH KACANG TANAH
TOLERAN TERHADAP CEKAMAN KEKERINGAN
DAN RESISTEN TERHADAP PENYAKIT BUSUK
BATANG Sclerotium rolfsii
Oleh :
A. Farid Hemon
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Agronomi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
viii
©
Hak cipta milik Institut Pertanian Bogor, tahun 2006
Hak cipta dilindungi
ix
GLOSARI
ES : embrio somatik, suatu sel tubuh tunggal yang diinduksi untuk ditumbuhkan ke dalam tanaman lengkap.
ES primer : embrio somatik yang pertama terbentuk dari eksplant ES sekunder : embrio somatik yang terbentuk dari embrio somatik
primer setelah disubkultur.
ES Pi-I : embrio somatik insensitif hasil seleksi selama satu siklus (3 bulan) pada media selektif MS-P16 yang mengandung polietilena glikol 15%.
ES Pi-II : embrio somatik insensitif hasil seleksi secara berulang ES Pi-I selama satu siklus lagi pada media selektif MS- P16 yang mengandung polietilena glikol 15% (dua siklus = 6 bulan).
ES PFi-I : embrio somatik insensitif hasil seleksi ES Pi-I diseleksi lagi selama satu siklus pada media selektif MS-P16 yang ditambahkan filtrat kultur 30% S. rolfsii (3 bulan pertama dalam media PEG dan kemudian 3 bulan kedua dalam media filtrat kultur).
ES Fi-I : embrio somatik resisten hasil seleksi selama satu siklus (3 bulan) pada media selektif MS-P16 yang mengandung filtrat kultur 30% Sclerotium rolfsii.
ES Fi-II : embrio somatik resisten hasil seleksi secara berulang ES Fi-I selama satu siklus lagi pada media selektif MS- P16 yang mengandung filtrat kultur 30% Sclerotium rolfsii (dua siklus = 6 bulan).
ES FPi-I : embrio somatik resisten hasil seleksi ES Fi-I diseleksi lagi selama satu siklus pada media selektif MS-P16 yang ditambahkan PEG 15%(3 bulan bulan pertama dalam media filtrat kultur dan kemudian 3 bulan kedua dalam media PEG).
Filtrat kultur : hasil ekstraksi miselia cendawan S. rolfsii
Generasi R0, R1, R2 : tanaman generasi R0 hasil kecambah ES seleksi in vitro : tanaman generasi R1 zuriat dari generasi R0
: tanaman generasi R2 zuriat dari generasi R1
MS : media dasar Murashige dan Skoog untuk kultur jaringan MS-P16 : media MS yang mengandung 16 µM auksin pikloram Planlet atau tanaman
Pi-I
: planlet atau tanaman ya ng berasal dari ES Pi-I
Planlet atau tanaman
Pi-II
: planlet atau tanaman yang berasal dari ES Pi-II
Planlet atau tanaman
PFi-I
: planlet atau tanaman yang berasal dari ES PFi-I
Planlet atau tanaman
Fi-I
: planlet atau tanaman yang berasal dari ES Fi-I
Planlet atau tanaman
Fi-II
x Planlet atau tanaman
FPi-I
: planlet atau tanaman yang berasal dari ES FPi-I
PEG : polietilena glikol dengan berat molekul besar yang dapat menurunkan potensial air
Seleksi berulang : seleksi in vitro yang dilakukan secara berulang atau lebih dari satu siklus (satu siklus = 3 bulan) pada media selektif PEG 15% atau pada filtrat kultur 30%.
Seleksi ganda : seleksi in vitro yang dilakukan sekaligus pada dua media selektif, tiga bulan pertama pada media MS-P16 yang ditambahkan PEG 15%, dan tiga bulan kedua pada media yang ditambahkan filtrat kultur 30%. Atau sebaliknya, tiga bulan pertama pada media yang ditambahkan filtrat kultur 30%, an tiga bulan kedua pada media yang ditambahkan PEG 15%
xi Judul Disertasi : EFEKTIFITAS SELEKSI IN VITRO BERULANG UNTUK MENDAPATKAN PLASMA NUTFAH KACANG TANAH TOLERAN TERHADAP CEKAMAN KEKERINGAN DAN RESISTEN TERHADAP PENYAKIT BUSUK BATANG Sclerotium rolfsii
Nama Mahasiswa : A. Farid Hemon
Nomor Pokok : A361020051
Program Studi : Agronomi
Menyetujui :
1. Komisi Pembimbing
Prof. Dr. Ir. Sudarsono, MSc. Prof. Dr. Ir. Rusmilah Suseno, MSc. Ketua Anggota
Dr. Ir. Hajrial Aswidinnoor, MSc. Dr. Ir. Widodo, MS Anggota Anggota
Mengetahui :
2. Ketua Program Studi Agronomi 3. Dekan Sekolah Pascasarjana
Dr.Ir. Satriyas Ilyas, MS. Prof. Dr. Ir. Khairil Anwar Notodiputro, MS.
xii
RIWAYAT HIDUP
A. FARID HEMON dilahirkan di Kabupaten Dompu Nusa Tenggara Barat pada tanggal 13 April 1963 sebagai anak kedua dari pasangan H. Hemon H. Arsyad dan Hj. St. Hadijah. Pendidikan Sarjana ditempuh di Jurusan Budidaya Pertanian Fakultas Pertanian Universitas Mataram, lulus pada tahun 1987. Pada tahun 1995, dia melanjutkan studi S2 dengan mayor Plant Breeding dan minor
Plant Pathology di Department of Horticulture, College of Agriculture, University of the Philippines Los Banos, dan lulus pada tahun 1997 dengan gelar Master of Science. Pada tahun 2002 dia terdaftar sebagai mahasiswa S3 di Sekolah Pascasarjana Institut Pertanian Bogor pada Program Studi Agronomi dengan menggunakan beasiswa pendidikan pascasarjana (BPPS) dari Departemen Pendidikan Nasional Republik Indonesia
Pada tanggal 29 Mei 1994, dia menikah dengan ULFAH KALSOM, dan dikarunia dua orang anak : Muhammad Dinullah Akbar (lahir 15 Maret 1995) dan Nurul Fidinia Hijah (lahir 20 Maret 1999).
xiii
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Mei 2003 ini ialah seleksi in vitro, dengan judul “Efektifitas Seleksi In Vitro Berulang untuk Mendapatkan Plasma Nutfah Kacang Tanah yang Toleran terhadap Cekaman Kekeringan dan Resisten terhadap Penyakit Busuk Batang Sclerotium rolfsii.
Penulis menyampaikan ucapkan terima kasih yang sebesar-besarnya kepada Bapak Prof. Dr. Sudarsono, Dr. Hajrial Aswidinnoor, Dr. Widodo, dan Ibu Prof. Dr. Rusmilah Suseno, selaku Komisi Pembimbing yang telah banyak meluangkan waktunya untuk membimbing selama studi, pelaksanaan penelitian dan penulisan Disertasi ini. Ucapan yang sama penulis sampaikan kepada bapak dan ibu orang tua, mertua, istri dan anak tercinta, kakak dan adik serta seluruh keluarga, atas do’a, dukungan dan kasih saya ngnya selama studi ini.
Semoga karya ilmiah ini bermanfaat.
Bogor, Desember 2006
xiv
DAFTAR ISI
Halaman
DAFTAR TABEL ... vii DAFTAR GAMBAR... xiii PENDAHULUAN... Latar Belakang... Tujuan Penelitian... Strategi Penelitian... 1 1 3 4 TINJAUAN PUSTAKA... Penyakit Busuk Batang dan Mekasisme Ketahanan Tanaman
terhadap Sclerotium rolfsii... Pengaruh Cekaman Kekeringan dan Mekanisme Toleransi... Mutasi In Vitro... Seleksi In Vitro untuk Resistensi terhadap Penyakit...
Seleksi In Vitro untuk Toleransi terhadap Cekaman Kekeringan 6 6 8 11 13 15
PERCOBAAN BAGIAN I : SELEKSI IN VITRO, DIAWALI
PADA MEDIA POLIETILENA GLIKOL (PEG) 16
SELEKSI BERULANG DAN IDENTIFIKASI EMBRIO SOMATIK KACANG TANAH YANG INSENSITIF POLIETILENA GLIKOL DAN FILTRAT KULTUR Sclerotium rolfsii...
Abstrak... Abstract... Pendahuluan... Bahan dan Metode... Hasil... Pembahasan... Kesimpulan... 18 18 19 20 21 25 33 36
TIPE VARIAN SOMAKLONAL YANG DIAMATI DIANTARA POPULASI TANAMAN KACANG TANAH HASIL SELEKSI IN VITRO BERULANG DAN SELEKSI GANDA...
xv PENAMPILAN TANAMAN KACANG TANAH HASIL SELEKSI
IN VITRO BERULANG DAN SELEKSI GANDA PADA
POLIETILENA GLIKOL DAN FILTRAT KULTUR Sclerotium rolfsii TERHADAP CEKAMAN LARUTAN POLIETILENA GLIKOL...
Abstrak... Abstract... Pendahuluan... Bahan dan Metode... Hasil... Pembahasan... Kesimpulan... 54 54 55 56 57 59 68 70
RESPONS TANAMAN KACANG TANAH SOMAKLON DARI HASIL REGENERASI SELEKSI IN VITRO BERULANG DAN
SELEKSI GANDA TERHADAP KEKERINGAN... Abstrak... Abstract... Pendahuluan... Bahan dan Metode... Hasil ... Pembahasan... Kesimpulan... 71 71 72 73 75 78 94 97
PERCOBAAN BAGIAN II : SELEKSI IN VITRO, DIAWALI
PADA MEDIA FILTRAT KULTUR Sclerotium rolfsii 99
SELEKSI IN VITRO BERULANG DAN SELEKSI GANDA SERTA IDENTIFIKASI PLANLET INSENSITIF CEKAMAN AKIBAT PENAMBAHAN FILTRAT KULTUR Sclerotium rolfsii DAN
POLIETILENA GLIKOL... Abstrak...
Abstract...
Pendahuluan... Bahan dan Metode...
Hasil... Pembahasan... Kesimpulan... 101 101 102 103 104 108 115 118 VARIASI SOMAKLONAL DIANTARA TANAMAN KACANG
TANAH HASIL SELEKSI IN VITRO BERULANG PADA MEDIA FILTRAT KULTUR Sclerotium rolfsii DAN POLIETILENA GLIKOL.
xvi SKRINING TANAMAN KACANG TANAH HASIL REGENERASI
EMBRIO SOMATIK YANG DISELEKSI SECARA BERULANG PADA FILTRAT KULTUR Sclerotium rolfsii DAN POLIETILENA GLIKOL TERHADAP INFEKSI S. rolfsii...
Abstrak... Abstract... Pendahuluan... Bahan dan Metode... Hasil... Pembahasan... Kesimpulan...
135 135 136 137 139 141 156 160 PEMBAHASAN UMUM... 161 SIMPULAN DAN SARAN... 168 DAFTAR PUSTAKA... 171
xvii
DAFTAR TABEL
Halaman
1. Pertumbuhan ES kacang tanah cv. Kelinci atau Singa hasil seleksi satu (Pi-I), dua siklus (Pi-II) dalam media MS-P16 dengan PEG, dan setelah seleksi ganda dalam media dengan PEG dan filtrat kultur S. rolfsii (PFi-I)
26
2. Respons terhadap cekaman PEG dari ES kacang tanah cv. Kelinci dan Singa insensitif PEG yang telah melalui satu (Pi-I), dua siklus (Pi-II) dalam media MS-P16 dengan PEG serta ES insensitif terhadap PEG dan filtrat kultur setelah melalui seleksi ganda dalam media dengan PEG dan filtrat kultur (PFi-I)
28
3. Respons terhadap cekaman filtrat kultur dari ES kacang tanah cv. Kelinci dan Singa insensitif PEG yang telah melalui satu (Pi-I), dua siklus (Pi-II) dalam media MS-P16 dengan PEG serta ES insensitif terhadap PEG dan filtrat kultur setelah melalui seleksi ganda dalam media dengan PEG dan filtrat kultur (PFi-I)
29
4. Perkecambahan dan regenerasi planlet dari ES kacang tanah cv. Kelinci dan Singa insensitif PEG yang telah melalui satu (Pi-I), dua siklus (Pi-II) dalam media MS-P16 dengan PEG atau insensitif terhadap PEG dan filtrat kultur setelah melalui seleksi ganda dalam media dengan PEG dan filtrat kultur (PFi-I)
30
5. Respons terhadap cekaman PEG dari stek pucuk planlet R0 kacang tanah cv. Kelinci dan Singa hasil regenerasi dari ES insensitif PEG satu (Pi-I) atau dua siklus (Pi-II) dalam media MS-P16 dengan PEG serta ES insensitif PEG dan filtrat kultur setelah melalui seleksi ganda dalam media PEG dan filtrat kultur (PFi-I)
31
6. Respons terhadap cekaman filtrat kultur dari setek pucuk planlet R0 kacang tanah cv. Kelinci dan Singa hasil regenerasi dari ES insensitif PEG satu (Pi-I) atau dua siklus (Pi-II) dalam media MS-P16 dengan PEG serta ES insensitif terhadap PEG dan filtrat kultur setelah melalui seleksi ganda dalam media PEG dan filtrat kultur (PFi-I)
32
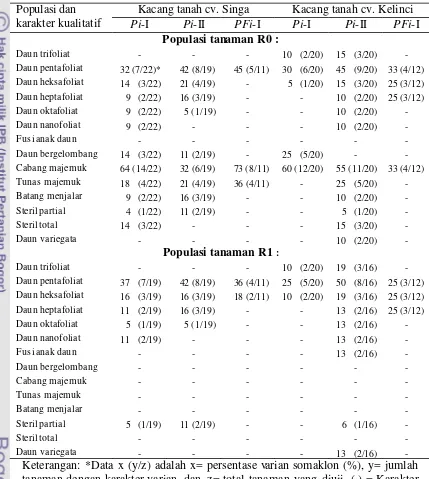
7. Macam dan persentase varian kualitatif yang diamati diantara populasi tanaman R0 yang diregenerasikan dari ES kacang tanah cv. Singa dan Kelinci hasil seleksi satu (Pi-I), dua siklus (Pi-II) dalam media PEG serta hasil seleksi ganda dalam media PEG dan diikuti dalam media filtrat kultur (PFi-I) serta populasi tanaman R1
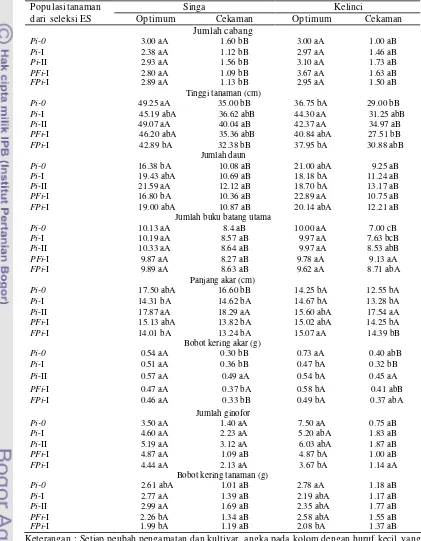
xviii 8. Jumlah cabang (JC), tinggi tanaman (TT), umur berbunga (UB),
jumlah daun (JD), bobot polong kering (BPK) dan jumlah polong kering (JPK) yang diamati pada tanaman cv. Kelinci dan Singa standar (Std), serta tanaman R0 yang diregenerasikan dari ES hasil seleksi in vitro pada media seleksi PEG siklus I (Pi-I), siklus II (Pi -II), dan seleksi ganda pada media PEG dan filtrat kultur (PFi-I)
47
9. Nomer galur tanaman generasi R2 zuriat dari generasi R1 kacang tana h hasil seleksi in vitro berulang dan seleksi ganda yang dievaluasi pada cekaman larutan PEG
57
10. Rata-rata jumlah hari tanaman untuk dapat bertahan hidup dan persentase (%) tanaman yang masih hidup sampai umur 50 hari dalam media yang diberi perlakua n PEG 15% pada populasi tanaman Pi-0 (tanpa seleksi in vitro) dan somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci hasil seleksi in vitro berulang pada media PEG dan seleksi ganda pada PEG dan filtrat kultur S. rolfsii
60
11. Pengaruh cekaman PEG 15% terhadap pertumbuhan tanaman populasi tanaman Pi-0 (tanpa seleksi in vitro) dan populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci hasil seleksi in vitro berulang pada media PEG dan seleksi ganda pada PEG dan filtrat kultur S. rolfsii
62
12. Indeks sensitivitas (S) terhadap cekaman PEG berdasarkan sejumlah karakter pertumbuhan pada populasi Pi-0 (tanpa seleksi in vitro) dan populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci ha sil seleksi in vitro berulang pada media PEG dan seleksi ganda pada PEG dan filtrat kultur S. rolfsii
64
13. Indeks sensitivitas (S) terhadap cekaman PEG berdasarkan sejumlah karakter pertumbuhan pada populasi Pi-0 (tanpa seleksi in vitro) dan galur populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci hasil seleksi in vitro berulang pada media PEG dan seleksi ganda pada PEG dan filtrat kultur S. rolfsii
66
14. Nomer galur tanaman generasi R2 zuriat dari generasi R1 kacang tanah hasil seleksi in vitro berulang dan seleksi ganda yang dievaluasi pada cekaman kekeringan
76
15 Pengaruh cekaman kekeringan terhadap pertumbuhan tanaman pada populasi Pi-0 (tanpa seleksi in vitro) dan populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci hasil seleksi in vitro berulang pada media PEG dan seleksi ganda pada PEG dan filtrat kultur S. rolfsii
xix 16. Persentase penurunannya bobot kering dan jumlah polong bernas
pada populasi Pi-0 (tanpa seleksi in vitro) dan populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci hasil seleksi in vitro berulang pada media PEG dan seleksi ganda pada PEG dan filtrat kultur S. rolfsii terhadap kondisi cekaman kekeringan dan optimum
81
17. Indeks sensitivitas kekeringan (S) berdasarkan sejumlah karakter agronomi pada populasi Pi-0 (tanpa seleksi in vitro) dan populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci hasil seleksi in vitro berulang pada media PEG dan seleksi ganda pada PEG dan filtrat kultur S. rolfsii
88
18. Indeks sensitivitas kekeringan (S) berdasarkan bobot kering dan jumlah polong bernas pada populasi Pi-0 (tanpa seleksi in vitro) dan galur populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci seleksi dua siklus pada PEG (seleksi in vitro berulang)
89
19. Kandungan prolin dan gula total pada populasi Pi-0 (tanpa seleksi in vitro) dan populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci hasil seleksi in vitro berulang pada media PEG dan seleksi ganda pada PEG dan filtrat kultur S. rolfsii
terhadap kondisi cekaman dan optimum
92
20. Pertumbuhan ES kacang tanah cv. Kelinci dan Singa setelah melalui satu (Fi-I), dua siklus (Fi-II) dalam media MS-P16 dengan filtrat kultur atau seleksi ganda dalam media dengan filtrat kultur dan PEG (FPi-I)
108
21. Respons ES kacang tanah hasil seleksi in vitro siklus I (Fi-I), siklus II (Fi-II), dan seleksi ganda (FPi-I) terhadap cekaman yang diinduksi oleh penambahan filtrat kultur S. rolfsii atau PEG dalam media
110
22. Perkecambahan dan regenerasi planlet dari ES kacang tanah cv. Kelinci dan Singa insensitif filtrat kultur yang telah melalui satu (Fi-I) atau dua siklus (Fi-II) dalam media MS-P16 dengan filtrat kultur serta ES insensitif terhadap filtrat kultur dan PEG setelah melalui seleksi ganda dalam media dengan filtrat kultur dan PEG (FPi-I)
111
23. Respons terhadap cekaman PEG dari setek pucuk planlet R0 kacang tanah cv. Kelinci dan Singa hasil regenerasi dari ES insensitif filtrat kultur melalui satu (Fi-I) atau dua siklus (Fi-II) dalam media MS-P16 dengan filtrat kultur serta ES insensitif terhadap filtrat kultur dan PEG setelah melalui seleksi ganda dalam media dengan filtrat kultur dan PEG (FPi-I)
xx 24. Respons terhadap cekaman filtrat kultur dari setek pucuk planlet R0
kacang tanah cv. Kelinci dan Singa hasil regenerasi dari ES insensitif filtrat kultur melalui satu (Fi-I) atau dua siklus (Fi-II) dalam media MS-P16 dengan filtrat kultur serta ES insensitif terhadap filtrat kultur dan PEG setelah melalui seleksi ganda dalam media dengan filtrat kultur dan PEG (FPi-I)
114
25. Macam dan persentase varian karakter kualitatif dibanding tanaman standar yang diamati diantara populasi tanaman generasi R0 dan R1 hasil seleksi ES satu (Fi-I), dua siklus (Fi-II) dalam media filtrat kultur serta hasil seleksi ganda dalam media filtrat kultur dan diikuti dalam media PEG (FPi-I) cv. Kelinci
126
26. Macam dan persentase varian karakter kualitatif diband ing tanaman standar yang diamati diantara populasi tanaman generasi R0 dan R1 hasil seleksi ES satu (Fi-I), dua siklus (Fi-II) dalam media filtrat kultur serta hasil seleksi ganda dalam media filtrat kultur dan diikuti dalam media PEG (FPi-I) cv. Singa
126
27. Jumlah cabang (JC), tinggi tanaman (TT), umur berbunga (UB), jumlah daun (JD), bobot polong kering (BPK) dan jumlah polong kering (JPK) tanaman kacang tanah cv. Kelinci dan Singa standar (Std), serta tanaman R0 hasil seleksi ES satu (Fi-I), dua siklus (Fi -II) pada media filtrat kultur serta hasil seleksi ganda dalam media filtrat kultur dan diikuti dalam media PEG (FPi-I)
127
28. Jumlah cabang (JC), tinggi tanaman (TT), umur berbunga (UB), jumlah daun (JD), bobot polong kering (BPK) dan jumlah polong kering (JPK) tanaman kacang tanah cv. Kelinci dan Singa standar (Std), serta tanaman R1 zuriat dari tanaman R0 hasil seleksi ES satu (Fi-I), dua siklus (Fi-II) pada media filtrat kultur serta seleksi ganda pada media filtrat kultur dan diikuti dalam media PEG (FPi-I)
128
29. Nomer galur tanaman kacang tanah hasil seleksi in vitro berulang dan seleksi ganda ES pada tanaman generasi R2 yang diskrining pada infeksi S. rolfsii
140
30. Persentase (%) tanaman mati dan jumlah hari hidup dari awal inokulasi sampai tanaman mati pada tanaman yang diinokulasi dengan cendawan S. rolfsii pada populasi tanaman Fi-0 (tanpa seleksi in vitro) dan somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci hasil seleksi in vitro berulang pada media dengan penambahan filtrat kultur 30% dan seleksi ganda pada filtrat kultur S. rolfsii dan PEG
xxi 31. Pengaruh inokulasi cendawan S. rolfsii terhadap pertumbuhan
tanaman pada populasi Fi-0 (tanpa seleksi in vitro) dan populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci hasil seleksi in vitro berulang pada media filtrat kultur S. rolfsii dan seleksi ganda pada filtrat kultur S. rolfsii dan PEG
143
32. Persentase penurunannya bobot kering dan jumlah polong bernas pada populasi Pi-0 (tanpa seleksi in vitro) dan populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci hasil seleksi in vitro berulang pada media filtrat kultur S. rolfsii dan seleksi ganda pada filtrat kultur S. rolfsii dan PEG terhadap infeksi cendawan S. rolfsii
144
33. Indeks kerentanan terhadap penyakit S. rolfsii (K) berdasarkan bobot kering dan jumlah polong bernas pada populasi Fi-0 (tanpa seleksi in vitro) dan populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci hasil seleksi in vitro berulang pada media filtrat kultur S. rolfsii dan seleksi ganda pada filtrat kultur S. rolfsii dan PEG
150
34. Indeks kerentanan terhadap penyakit (K) berdasarkan bobot kering polong dan jumlah polong pada populasi Fi-0 (tanpa seleksi in vitro) dan galur populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci dua siklus (seleksi in vitro berulang)
152
35 Aktivitas enzim peroksidase dan enzim kitinase menggunakan substrat dimer (pNP-NAcGlc) akibat infeksi S. rolfsii pada pangkal batang pada populasi Fi-0 (tanpa seleksi in vitro) dan populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci hasil seleksi in vitro berulang pada media filtrat kultur S. rolfsii dan seleksi ganda pada filtrat kultur S. rolfsii dan PEG
xxii
DAFTAR GAMBAR
Halaman
1. Diagram alir skenario penelitian dan keterkaitan antar percobaan 5
2. Respons perkembangan ES yang ditanam dalam media MS-P16 dan media selektif. PEG
27
3. Regenerasi ES insensitif PEG dan filtrat kultur untuk membentuk planlet.
30
4. Contoh fenotipe varian yang diamati diantara populasi tanaman generasi R0 dan R1 yang diregenerasikan dari ES hasil seleksi in vitro berulang dalam media selektif PEG 15% atau hasil seleksi dalam media dengan PEG, diikuti dengan media filtrat kultur (seleksi ganda)
44
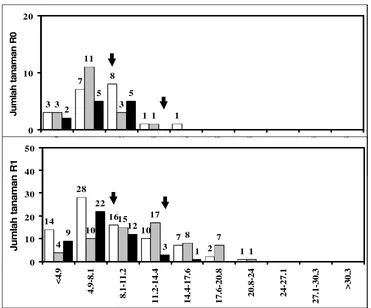
5. Jumlah tanaman R0 yang diregenerasikan dari ES cv. Kelinci hasil seleksi in vitro selama satu atau dua siklus dalam media PEG serta satu siklus dalam media PEG, diikuti media filtrat kultur (seleksi ganda) dan R1 zuriat dari tanaman R0 yang menghasilkan kisaran bobot polong kering tertentu
48
6. Jumlah tanaman R0 yang diregenerasikan dari ES cv. Singa hasil seleksi in vitro selama satu atau dua siklus dalam media PEG serta satu siklus dalam media PEG, diikuti dengan media filtrat kultur (seleksi ganda) dan R1 zuriat dari tanaman R0 yang menghasilkan kisaran bobot polong kering tertentu
49
7. Representasi respons tanaman kacang tanah terhadap cekaman PEG 59
8. Pengaruh cekaman kekeringan terhadap perumbuhan tanaman, akar dan hasil polong
82
9 Jumlah tanaman standar dan generasi R2 yang diregenerasikan dari ES kacang tanah cv. Singa hasil seleksi in vitro dan menghasilkan bobot polong kering (g) setelah mengalami cekaman kekeringan dan dibandingkan denga n tanaman dalam konsisi optimum
83
10. Jumlah tanaman standar dan generasi R2 yang diregenerasikan dari ES kacang tanah cv. Singa hasil seleksi in vitro dan menghasilkan jumlah polong kering setelah mengalami cekaman kekeringan dan dibandingkan dengan tanaman dalam konsisi optimum
84
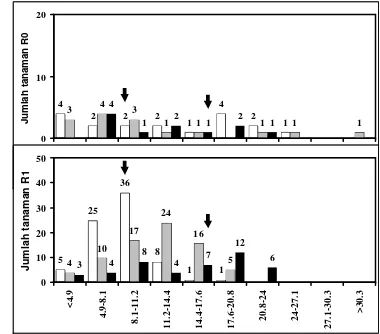
11. Jumlah tanaman standar dan generasi R2 yang diregenerasikan dari ES kacang tanah cv. Kelinci hasil seleksi in vitro dan menghasilkan bobot polong kering (g) setelah mengalami cekaman kekeringan dan dibandingkan denga n tanaman dalam konsisi optimum
xxiii 12. Jumlah tanaman standar dan generasi R2 yang diregenerasikan dari
ES kacang tanah cv. Kelinci hasil seleksi in vitro dan menghasilkan jumlah polong kering setelah mengalami cekaman kekeringan dan dibandingkan dengan tanaman dalam konsisi optimum
86
13 Kadar prolin pada populasi somaklon generasi R2 yang diregenerasikan dari ES cv. Singa dan Kelinci hasil seleksi in vitro satu siklus (Pi-I), dua siklus II (Pi-II) pada media PEG dan seleksi ganda pada PEG dan filtrat kultur S. rolfsii(PFi-I atau FPi-I)
93
14 Respons perkembangan ES yang ditanam dalam media selektif filtrat kultur S. rolfsii 30% dan PEG 15 %
109
15 Regenerasi ES insensitif filtrat kultur dan PEG untuk membentuk planlet
111
16 Varian kualitatif yang teramati diantara populasi tanaman generasi R0 dan R1 yang regenerasi ES dari hasil seleksi in vitro berulang dalam media selektif dengan penambahan filtrat kultur dan seleksi ganda dalam media filtrat kultur dan PEG
125
17. Berat polong kering per tanaman (g) dari nomer- nomer galur somaklon generasi R1 cv. Kelinci yang dihasilkan dari seleksi ES satu siklus (Fi-I), dua siklus (Fi-II), dan seleksi ganda (FPi-I) dibandingkan lebih ringan atau lebih berat dari populasi tanaman standar
130
18. Berat polong kering per tanaman (g) dari nomer- nomer galur somaklon generasi R1 cv. Singa yang dihasilkan dari seleksi ES satu siklus (Fi-I), dua siklus (Fi-II), dan seleksi ganda (FPi-I) dibandingkan lebih ringan atau lebih berat dari populasi tanaman standar
131
19. Jumlah tanaman standar dan generasi R2 yang diregenerasikan dari ES kacang tanah cv. Singa hasil seleksi in vitro dan menghasilkan bobot polong kering (g) setelah diinokulasi dengan S. rolfsii dan dibandingkan dengan tanaman sehat
146
20 Jumlah tanaman standar dan generasi R2 yang diregenerasikan dari ES kacang tanah cv. Singa hasil seleksi in vitro dan menghasilkan jumlah polong kering setelah diinokulasi dengan S. rolfsii dan dibandingkan dengan tanaman sehat
147
21 Jumlah tanaman standar dan generasi R2 yang diregenerasikan dari ES kacang tanah cv. Kelinci hasil seleksi in vitro dan menghasilkan bobot polong kering (g) setelah diinokulasi dengan S. rolfsii dan dibandingkan dengan tanaman sehat
xxiv 22. Jumlah tanaman standar dan generasi R2 yang diregenerasikan dari
ES kacang tanah cv. Kelinci hasil seleksi in vitro dan menghasilkan jumlah polong kering (g) setelah diinokulasi dengan S. rolfsii dan dibandingkan dengan tanaman sehat
149
Latar Belakang
Kacang tanah (Arachis hypogaea L.) merupakan salah satu sumber protein nabati yang cukup penting dalam pola menu makanan setelah tanaman kedelai. Kebutuhan kacang tanah dalam negeri menunjukkan kenaikan cukup besar yaitu 4,4% per tahun, sedangkan produksi kacang tanah hanya mengalami kenaikan 2,5%. Akibatnya untuk memenuhi kekurangan produksi tersebut harus diimpor (Adisarwanto 2003).
Luas pertanaman kacang tanah menempati urutan keempat setelah padi, jagung, dan kedelai. Di kawasan Asia, Indonesia menempati urutan ketiga dengan luas penanaman mencapai 650.000 Ha dan merupakan negara ketujuh terbesar penghasil kacang tanah di dunia. Namun, produksi kacang tanah Indonesia masih rendah yaitu sekitar 1 ton/Ha polong kering, sedangkan negara lain melebihi 2 ton/Ha (Adisarwanto 2003; Biro Pusat Statistik 1999). Penanaman kacang tanah sebagian besar (70%) ditanam pada lahan kering dan sisanya ditanam pada lahan basah (Saleh & Adisarwanto 1996).
Usaha tani pada lahan kering sulit berkembang karena ketersediaan air tidak selalu terpenuhi sepanjang musim tanam. Hal ini menyebabkan produksi kacang tanah menjadi rendah dan bahkan menyebabkan kegagalan panen. Cekaman kekeringan menyebabkan gangguan pada semua fase pertumbuhan tanaman dan menimbulkan berbagai efek seperti pada perluasan daun dan pemanjangan batang, gangguan aktivitas fotosintesis (Riciardi et al. 2001), menurunkan kemampuan untuk melakukan pembuahan (self-fertility) dan menurunkan ukuran biji (Stoddar 1986). Cekaman air menyebabkan pengurangan biomasa daun dan polong kering kacang tanah (Collino et al. 2000) dan penurunan bobot kering polong diduga disebabkan oleh terhambatnya inisiasi dan pemanjangan ginofor (Chapman et al. 1993).
produksi. Kondisi ini sangat bertentangan dengan penerapan pemuliaan moderen yang selalu mensyaratkan menggunakan input teknologi secara intensif (Cercarelli 1996). Salah satu pendekatan yang rasional untuk mengatasi masalah budidaya pada lahan kering adalah dengan mengembangkan kultivar kacang tanah yang berdaya hasil tinggi pada lingkungan tersebut.
Selain masalah cekaman kekeringan, masalah penyakit busuk batang akibat infeksi S. rolfsii juga merupakan faktor pembatas produksi usaha tani kacang tanah di lahan kering. Infeksi patogen dapat menurunkan kuantitas dan kualitas hasil kacang tanah. Menurut Backman dan Brenneman (1997) penurunan hasil akibat serangan patogen ini dapat mencapai 25-80 %. Sclerotium rolfsii
merupakan cendawan tertular tanah (soil borne) dan membentuk sklerotia yang menyebabkan cendawan ini mampu bertahan hidup cukup lama di dalam tanah. Kondisi lahan kering yang sulit diterapkan sistem pengairan, menyebabkan inokulum cendawan sulit dihilangkan pada usaha tani lahan kering (Benhamou & Chert 1996).
Penanaman kultivar resisten yang berdaya hasil tinggi menawarkan harapan dapat mengembangkan budidaya kacang tanah di lahan kering. Pengendalian secara kimiawi dirasakan tidak ekonomis terutama untuk petani lahan kering, tidak efektif karena cendawan soil-born, dan dapat mencemarkan lingkungan. Penggunaan kultivar tahan diharapkan dapat menghambat atau mengurangi perkembangan patogen.
(PEG) yang dapat menstimulasi kondisi cekaman kekeringan untuk mengidentifikasi sel atau jaringan tanaman kacang tanah yang tidak mati oleh cekaman PEG (Dami & Hughes 1997; Widoretno et al. 2003a; Rahayu et al. 2005).
Efektifitas seleksi in vitro sangat tergantung dari keberhasilan untuk menghambat perkembangan sel/jaringan normal yang tidak diinginkan dan memproliferasikan sel/jaringan mutan yang diinginkan dengan menggunakan agens penyeleksi. Penggunaan seleksi in vitro berulang diharapkan dapat meningkatkan akumulasi mutagenesis sel-sel atau jaringan tanaman yang hanya toleran terhadap PEG atau resisten terhadap filtrat kultur atau kedua sifat tersebut sekaligus dapat terakumulasi pada satu genotip. Berdasarkan uraian tersebut, maka penelitian ini sangat diperlukan untuk menguji efektifitas seleksi in vitro berulang untuk mendapatkan kultivar kacang tanah unggul baru yang toleran terhadap cekaman kekeringan dan resisten terhadap penyakit busuk batang S. rolfsii.
Tujuan Penelitian
Secara umum penelitian ini bertujuan untuk menguji efektifitas seleksi in vitro berulang untuk mendapatkan plasma nutfah tanaman kacang tanah yang toleran terhadap cekaman kekeringan dan resisten terhadap penyakit busuk batang
S. rolfsii.
Secara spesifik penelitian ini bertujuan :
1. Menguji efektifitas seleksi in vitro berulang dalam media yang mengandung PEG untuk me ndapatkan tanaman kacang tanah yang toleran kekeringan. 2. Menguji efektifitas seleksi in vitro berulang dalam media yang mengandung
filtrat kultur S. rolfsii untuk mendapatkan tanaman kacang tanah yang resisten terhadap penyakit busuk batang S. rolfsii.
Strategi Penelitian
Untuk me ncapai tujuan penelitian tersebut, strategi penelitian yang ditempuh meliputi beberapa topik penelitian sebagai berikut :
1. Efektifitas seleksi in vitro berulang dalam media selektif yang mengandung PEG untuk mendapatkan varian embrio somatik yang insensitif terhadap PEG.
2. Efektifitas seleksi in vitro berulang dalam media selektif yang mengandung filtrat kultur S. rolfsii untuk mendapatkan varian embrio somatik yang insensitif terhadap filtrat kultur.
3. Efektifitas seleksi in vitro ganda dalam media yang mengandung PEG dan filtrat kultur untuk mendapatkan varian embrio somatik yang insensitif PEG dan filtrat kultur.
4. Evaluasi keragaman karakter kualitatif (morfologis) dan kuantitatif (agronomis) pada populasi kacang tanah hasil seleksi in vitro berulang dan seleksi ganda (pada media yang mengandung PEG dan filtrat kultur S. rolfsii).
1. Embrio somatik insensitif PEG Embrio somatik insensitif PEG dan filtrat kultur
2. Embrio somatik insensitif filtrat kultur
Populasi tanaman varian somaklonal hasil seleksi in vitro
Varian kualitatif dan kuantitatif a. Varian toleran kekeringan b. Varian resisten S. rolfsii
c. Varian karakter ganda
Galur Kacang Tanah Unggul Baru (Toleran kekeringan dan resisten S. rolfsii)
Gambar 1. Diagram alir skenario penelitian dan keterkaitan antar percobaan
Plasma nutfah kacang tanah (cv. Singa dan Kelinci)
Seleksi in vitro berulang dengan 1) PEG dan 2) Filtrat kultur
Seleksi ganda (PEG dan filtrat kultur)
Regenerasi varian somaklonal
Evaluasi karakter kualitatif dan kuantitatif
Identifikasi varian somaklonal a. Toleran kekeringan
Penyakit Busuk Batang dan Mekasisme Ketahanan Tanaman terhadap
Sclerotium rolfsii
Sclerotium rolfsii Sacc. menyebabkan penyakit busuk batang pada kacang tanah dan berbagai spesies tanaman penting lain. Cendawan ini tertular lewat tanah sehingga pengendalian secara kultur teknik menjadi sangat sulit (Backman & Brenneman 1997). Kacang tanah dapat diserang pada setiap stadium pertumbuhan, mulai dari benih, kecambah sampai tanaman dewasa. Serangan pada fase kecambah menyebabkan tanaman rebah kecambah karena pada pangkal batang terjadi pembusukan. Serangan pada tanaman dewasa menyebabkan lesio berwarna coklat pada batang, daun mulai gugur, terjadi pembusukan pada pangkal batang, satu dua cabang menjadi layu, dan akhirnya tanaman mati. Infeksi juga terjadi pada ginofor dan pembusukan pada polong (Agrios 1988; Melouk & Backman 1995; Benhamou & Chert 1996)
Selama kondisi tidak menguntungkan seperti kekurangan nutrisi, kondisi lingkungan panas atau dingin, adanya mikroba yang antagonis, atau tidak ada inang, menyebabkan cendawan akan membentuk sklerotia. Sklerotia membentuk struktur yang kompak, berukuran seragam, berbentuk agak bulat atau tak beraturan, berwarna putih ketika baru terbentuk dan akan menjadi berwarna coklat tua sampai kehitaman ketika telah masak (Hadar & Gorodecki 1991). Sklerotia tersebut mampu bertahan di dalam tanah selama bertahun-tahun, mampu berkolonisasi, dan mampu hidup secara efektif pada berbagai macam bahan organik dan dapat meningkatkan populasi inokulum (Punja 1985).
Efektifitas patogenisitas cendawan S. rolfsii membutuhkan sekresi toksin asam oksalat yang bersifat racun bagi inang. Asam oksalat bersifat toksin bagi tanaman sehingga dapat mematikan jaringan tanaman inang (Backman & Brenneman 1997; Cessna et al. 2000). Asam oksalat yang dikeluarkan dapat meningkatkan virulensi cendawan S. rolfsii karena enzim seperti poligalakturonase akan aktif ketika adanya asam oksalat. Kerjasama asam oksalat dan enzim ini mengakibatkan kerusakan yang lebih parah pada jaringan tanaman. Senyawa asam oksalat dapat menyebabkan klorosis dan nekrosis daun selama stadia pertama perkembangan tanaman. Oksalat dapat menghambat aktivitas
oxidative burst yang merupakan mekanisme fisiologis tanaman untuk melawan patogen.
Pengaruh Cekaman Kekeringan dan Mekanisme Toleransi
Ketersediaan air merupakan pembatas utama produksi tanaman. Tanaman akan terganggu pertumbuhannya ketika ketersediaan air dipermukaan akar berkurang. Konsep cekaman kekeringa n pada tanaman adalah berkurangnya suplai air di daerah perakaran dan permintaan air yang berlebihan oleh daun, yang mana laju transpirasi melebihi laju absorpsi air oleh akar (Kaufmann 1981). Perilaku akar selama cekaman kekeringan dan hubungannya dengan daun (pucuk) tanaman belum dimengerti secara jelas. Pada keadaan tanah kering, perubahan signal metabolisme biokimia akar pada daun adalah dengan menekan pertumbuhan, aktivitas stomata, fotosintesis dan potensial osmotik (Kramer & Boyer 1995). Untuk mempertahankan turgor tanaman dapat dijaga keseimbangan antara laju transpirasi dan penyerapan air oleh akar.
Dua komponen utama resistensi tanaman terhadap kekeringan (Franca et al. 2000) yaitu terhindar kekeringan (drought avoidance) dan toleransi terhadap kekeringan (drought tolerance). Pada kacang-kacangan, mekanisme terhindar kekeringan yang utama adalah sistem perakaran yang baik dan efisiensi pembukaan stomata (Barradas et al. 1994). Mekanisme toleransi kekeringan pada status air tanaman rendah adalah proses yang berfungsi pada tingkat sel, dan yang paling penting adalah pengaturan osmotik (osmotic adjustment) dan proteksi sistem membran (Mullet & Whitsitt 1996).
et al. 1993). Pada tanaman kedelai, defisit air menurunkan luas area dan kandungan klorofil daun (Shimada et al. 1992), menurunkan ukuran polong, biji, dan bobot kering polong (Pookpadi et al. 1990), dan menurunkan kualitas biji (Franca-Neto et al. 1993). Cekaman kekeringan mempengaruhi pula sis tem reproduksi tanaman. Herrero dan Johnson (1981) mempelajari pengaruh cekaman kekeringan pada sistem reproduksi tanaman jagung. Hasil penelitian menunjukkan bahwa pemanjangan rambut jagung (silk) terhenti pertumbuhan pada potensial air daun kira-kira -9 bar. Gejala morfologi yang biasa nampak pada tanaman kekurangan air adalah tanaman layu, daun menggulung, dan kerdil.
Pengaruh cekaman kekeringan pada sel dapat dilihat pada kemampuan melakukan pembelahan dan pemanjangan sel. Pengurangan pemberian air pada tanaman menurunkan jumlah dan ekspansi sel (Granier & Tardieu 1999). Pada penelitian stres osmotik menghasilkan penurunan kadar klorofil dan menurunkan aktivitas mitokondria (Gibon et al. 2000).
Pengaruh lanjut dari osmotic adjusment adalah memelihara pertumbuhan akar dan tunas pada kondisi cekaman, melalui pengaturan kontrol turgor sel (Creelman et al. 1990; Mullet & Whitsitt 1996). Integritas struktur membran sel adalah penting untuk dapat bertahan hidup pada periode cekaman kekeringan. Ketahanan pada fase kecambah dan vegetatif adalah penting agar mendapatkan hasil yang baik.
Cekaman kekeringan mempengaruhi proses fisiologi berupa penurunan transpirasi dan potensial air rendah, dan fotosintesis (Pookpadi et al. 1990; Vieira
et al. 1992). Tanaman yang terkena stres kekringan, potensi air daun menurun, pembentukan klorofil daun terganggu (Alberte et al. 1977) dan struktur kloroplas mengalami disintegrasi (Van Doren & Reicosky 1987). Gangguan fotosintesis terutama berhubungan dengan penutupan stomata, sehingga gas CO2 tidak dapat
Cekaman kekeringan menyebabkan respons biokimia atau metabolik pada tanaman. Karakter metabolik berupa akumulasi prolin pada jaringan tanaman merupakan karakter untuk toleransi tanaman terhadap cekaman kekeringan. Tanaman yang toleran terhadap stres osmotik mengakumulasikan prolin jauh lebih tinggi dari tanaman peka. Asam amino ini dipercaya berfungsi sebagai protektor atau menstabilisasi enzim atau struktur membran yang sensitif pada kekeringan atau kerusakan yang diinduksi secara ionik (Madan et al. 1995; Girousse et al. 1996; Iyer & Caplan 1998). Kishor et al. (1995) mengamati terjadinya peningkatan reistensi pada defisit air dan cekaman garam pada tanaman transgenik tembakau dengan over ekspresi ?1-pyroline-5-carboxylate synthetase
(P5CS). Penulis lain menjelaskan bahwa prolin dapat berpartisipasi sebagai penyimpan nitrogen, yang sewaktu-waktu dapat digunakan setelah periode cekaman. Tambahan pula bahwa prolin berperanan dalam meregulasi pH sitoplasma (Pesci & Beffagna 1984). Prolin jelas merupakan senyawa osmotikum dan compatible solut yang dapat terakumulasi pada konsentrasi tinggi pada sel sitoplasma tanpa merusak struktur sel. Akumulasi prolin berfungsi sebagai detoksifikasi radikal dan meregulasi status redoks sel.
Sumber metabolik dari akumulasi prolin pada potensial air rendah belum jelas teramati. Salah satu mekanisme yang dapat menjelaskan adalah meningkatkan sintesis prolin dengan menggunakan glutamin dan ornitin sebagai prekursor. Transkripsi mRNA mengkode ?1-pyroline-5-carboxylate reductase
(P5CR) dan P5CS dan selanjutnya mengkatalisis sintesis prolin melalui glutamin (Madan et al. 1995; Yoshiba et al. 1995).
Ketika konsentrasi ion anorganik tinggi pada vakuola, senya wa gula dapat melindungi enzim sitosolik (Rodrigues et al. 1999).
Ketika cekaman kekeringan, tanaman dapat mensintesis hormon tumbuh berupa abscisic acid (ABA). Pembentukan senyawa ini berkaitan dengan respons terhadap kelayuan (Pruvot et al. 1996; Nambara et al. 1998) yaitu untuk meningkatkan laju penyerapan air oleh akar untuk mengimbangi laju transpirasi (Quintero et al. 1998; Roberts 1998). Penyerapan air di akar ditentukan oleh besarnya gradien hidrostatik atau gradien osmotik. Dalam hal ini ABA berperan sebagai pengatur signal transport air secara radial (Quintero et al. 1998).
Mutasi In Vitro
Munculnya variasi somaklonal selama kultur in vitro berhubungan dengan mutasi titik, chromosomal rearrangement dan rekombinan, transposable element, jumlah kopi sekuens DNA, dan DNA amplifikasi. Sistem kultur jaringan sendiri ikut mengambil bagian dalam sistem mutagenesis karena sel mengalami traumatik setelah diisolasi dari tanaman dan beberapa prosedur atau restruktur isasi dalam proses regenerasi yang berbeda dengan di alam menyebabkan pula munculnya variasi epigenetik pada tanaman ya ng baru diregenerasi (Jain 2000; Larkin 2004). Pengaruh tipe genotipe, tipe eksplan, media kultur, umur tanaman donor, dan jumlah sub kultur berpengaruh pula pada munculnya variasi somaklonal (Veilleux & Johnson 1998; Jain 2001).
Variasi kromosom telah diamati pada beberapa spesies tanaman yang diregenerasikan dari kultur jaringan dan tanaman turunannya (Duncan 1997; Gupta 1998). Eksplan dengan ploidi dan jumlah kromosom tinggi menunjukkan variabilitas lebih tinggi dibanding ploidi dan jumlah kromosom rendah. Pengendalian siklus sel normal dengan mencegah pembelahan sel sebelum mengakhiri replikasi DNA yang terjadi selama kultur in vitro, menyebabkan patahnya kromosom (chromosomal breakage) (Philip et al. 1994). Chromosomal breakage dan konsekuensinya (delesi, duplikasi, inversi, dan translokasi) menyebabkan aberasi kromosom (Duncan 1997). Patahnya kromosom menyebabkan mutasi secara langsung melalui “position effect” atau mengubah ekspresi gen dari chromosomal rearrangement dan menempatkan pada daerah khusus heterokromatin. Metilasi DNA juga menyebabkan patahnya kromosom.
Variasi somaklonal menimbulkan perubahan genetik pada tanaman dan karakter tanaman dapat berubah dari tanaman awal seperti tinggi tanaman, hasil, jumlah bunga per tanaman, kualitas polong, toleran terhadap kekeringan dan resisten terhadap penyakit. Valkonen et al. (1999) mendapatkan tanaman kerdil (pertumbuhannya lambat), ruas-ruas batang pendek, dan daun hijau tua pada tanaman kentang yang diregenerasikan dari kultur antera. Stephens et al. (1991) tidak menemukan perbedaan yang luas antara tanaman turunan homosigot yang diregenerasikan melalui organogenesis pada tanaman kedelai. Ketika dibandingkan dengan tanaman tetua, ternyata kematangan, tinggi tanaman, protein dan minyak biji dan hasil biji berbeda. Variasi somaklonal teramati pada tanaman kedelai yang diregenerasikan melalui proses embriogenesis dan organogenesis. Karakter varian lain yang muncul akibat variasi somaklonal pada tanaman kedelai adalah sterilitas jantan (male sterility), perubahan morfologi daun (Amberger et al. 1992a), dan adanya alel baru dari isozim (Amberger et al. 1992b).
Identifikasi varian somaklonal yang diinginkan biasanya dilakukan dengan menggunakan teknik seleksi in vitro. Kondisi selektif tertentu dapat digabungkan dalam media kultur dan dipakai untuk menumbuhkan varian-varian somaklonal yang telah diperoleh. Tanaman hasil regenerasi dari jaringan yang dapat mengatasi kondisi selektif, besar kemungkinan juga akan menghasilkan fenotipe toleran terhadap kondisi selektif. Hal ini sangat menguntungkan karena proses seleksi in vitro membutuhkan tempat yang relatif sedikit, kondisi selektif dapat dibuat homogen, dan efektifitas seleksi sangat tinggi.
Seleksi In Vitro untuk Resistensi terhadap Penyakit
2003). Pada penelitian Yusnita et al. (2005) juga telah dikembangkan protokol untuk induksi ES, perkecambahan dan regenerasi planlet.
Penggunaan seleksi in vitro dengan menggunakan toksin yang diproduksi oleh patogen telah banyak dilakukan oleh beberapa peneliti. Seleksi in vitro untuk ketahanan terhadap penyakit telah dilakukan seperti Fusarium oxysporum pada tanaman tomat dengan menggunakan agens penyeleksi asam fusarik,
Phytophthora infestans pada kentang dengan menggunakan filtrat kultur,
Xanthomonas oryzae pada padi dengan filtrat kultur (Jayasangkar 2005), dan penyakit busuk batang S. rolfsii pada kacang tanah dengan menggunakan filtrat kultur (Yusnita et al. 2005). Tanaman yang dihasilkan setelah lima atau lebih siklus berulang menghasilkan tanaman resisten terhadap toksin dan patogen. Pada kedelai, tanaman yang diregenerasikan dari kalus terseleksi pada filtrat kultur patotoksik Septoria glycines menyebabkan tertundanya gejala becak coklat dibanding tanaman tanpa seleksi in vitro (Song et al. 1994).
Satu persyaratan untuk keberhasilan sistem seleksi in vitro adalah tersedianya metode regenerasi tanaman. Pada kacang tanah, metode seleksi in vitro telah tersedia (Yusnita et al. 2005). Seleksi in vitro dapat digunakan untuk menapis sel/jaringan varian yang diinginkan. Media selektif akan mematikan dan menghambat petumbuhan sel/jaringan normal yang peka terhadap filtrat kultur dan menumbuhkan sel/jaringan varian yang resisten sehingga dihasilkan tanaman somaklon yang resisten terhadap filtrat kultur cendawan S. rolfsii.
Cendawan S. rolfsii membutuhkan sekresi toksin asam oksalat untuk dapat mematikan jaringan tanaman inang (Backman & Brenneman 1997; Cessna
Seleksi In Vitro untuk Toleransi terhadap Cekaman Kekeringan
Potensi osmotik medium adalah penting untuk digunakan dalam seleksi in vitro embrio somatik. Cekaman osmotik yang diterapkan pada kultur in vitro mengurangi kemampuan regenerasi. Namun, setelah diskrining di lapangan, ternyata tanaman regeneran yang dihasilkan dari cekaman osmotik menghasilkan tana man toleran dan hasilnya lebih tinggi dari tetua pada kondisi kekeringan (Mohamed et al. 2000). Beberapa tanaman yang telah diseleksi pada cekaman osmotikum seperti Chrysanthenum morifolium (Shibli et al. 1992), tanaman sorgum (Duncan et al. 1995), padi (Adkins et al. 1995) dan kedelai (Widoretno
et al. 2003a).
PERCOBAAN BAGIAN I :
SELEKSI IN VITRO,
DIAWALI PADA MEDIA
POLIETILENA GLIKOL
DIAGRAM ALIR PERCOBAAN BAGIAN I :
(Seleksi in vitro, diawali pada media selektif polietilena glikol = PEG)
Seleksi I dlm PEG
Proliferasi ES pd media MS -P16
Seleksi I filtrat kultur Regenerasi planlet Seleksi II dlm PEG
Regenerasi planlet Regenerasi tanaman Regenerasi planlet
Regenerasi tanaman Regenerasi tanaman
Keterangan :
= ES insensitif Pi-I, Pi-II, dan PFi-I diuji responsnya pada media PEG 15% dan filtrat
kultur 30%
= Planlet yang berasal dr ES Pi-I, Pi-II, dan PFi-I diuji responsnya pada media PEG
15% dan filtrat kultur 30%
= Tanaman akhir hasil seleksi ES Pi-I, Pi-II, dan PFi-I diamati tipe & macam variasi
somaklonal, diuji responsnya pada cekaman larutan PEG 15% dan cekaman kekeringan
ES INSENSITIF PEG Siklus I (Pi-I)
ES INSENSITIF PEG Siklus I (Pi-I) 50 clump ES INSENSITIF PEG
Siklus I (Pi-I) ES INSENSITIF PEG
Siklus I (Pi-I) 50 clump
ES INSENSITIF PEG & Filtrat kultur, seleksi
ganda (PFi-I)
Planlet dari ES PFi-I
ES INSENSITIF PEG Siklus II (Pi-II)
Planlet dari ES Pi-II Planlet dr ES Pi-I
Kalus ES (50 clump)
Tanaman dari ES PFi-I Generasi R0, R1, & R2
Tanaman dari ES Pi-I, Generasi R0, R1, & R2
POLIETILENA GLIKOL DAN FILTRAT
KULTUR Sclerotium rolfsii
Abstrak
Percobaan dilakukan untuk mengevaluasi efektivitas seleksi berulang dalam media polieilena glikol (PEG) untuk meningkatkan frekuensi didapatkannya embrio somatik (ES) kacang tanah yang insensitif PEG serta seleksi ganda dalam media PEG dan filtrat kultur S. rolfsii untuk meningkatkan frekuensi didapatkannya ES yang insensitif sekaligus pada media PEG dan filtrat kultur. Regenerasi planlet R0 dan evaluasi respons setek pucuk planlet R0 terhadap PEG atau filtrat kultur juga dilakukan untuk menentukan keberhasilan mendapatkan tanaman varian. Embrio somatik diseleksi selama satu, dua siklus dalam PEG, dan satu siklus dalam PEG dan satu siklus dalam filtrat kultur (3 bulan per siklus seleksi). Selanjutnya, ES yang insensitif terhadap kondisi cekaman diproliferasi dan dikecambahkan menjadi tanaman R0. Hasil percobaan menunjukkan bahwa ES dan tunas R0 cv. Singa dan Kelinci yang diseleksi dua siklus dalam media PEG lebih insensitif terhadap PEG. Sebaliknya, ES dan tunas R0 cv. Singa dan Kelinci hasil seleksi ganda lebih insensitif terhadap filtrat kultur.
REPEAT CYCLING SELECTION AND IDENTIFICATION OF
POLYETHYLENE GLYCOL (PEG) AND Sclerotium rolfsii
CULTURE FILTRATE INSENSITIVE SOMATIC
EMBRYOS OF PEANUT
Abstrak
The objectives of this experiment were to evaluate effectiveness of repeat cycling and double in vitro selection to increase frequency of obtaining PEG and
S. rolfsii culture filtrate insensitive somatic embryos (SE) of peanut. Regeneration of the R0 plants and their evaluation against PEG and culture filtrate were conducted to determine the success of identifying tolerance variants. Peanut SE was subjected to one, two cycles of PEG selection, and one cycle of PEG and culture filtrate selection (3 months per cycle). The identified insensitive SE was proliferated and regenerated into R0 shoots and plantlets. Results of the experiments showed after two cycles of in vitro selection, PEG insensitive SE and its R0 shoots of peanut cv. Kelinci and Singa were more tolerance against stress due to PEG. While ones identified from double selection were more tolerance to culture filtrate.
Pendahuluan
Peningkatan keragaman genetik yang merupakan komponen penting dalam program pemuliaan tanaman dapat dilakukan dengan berbagai pendekatan, yaitu hibridisasi, introduksi plasma nutfah, mutagenesis, dan induksi variasi somaklona l dalam kultur in vitro (Neal et al. 1993). Induksi variasi somaklonal yang diikuti dengan seleksi in vitro dilaporkan efektif untuk mengidentifikasi varian tanaman dengan sifat unggul, seperti toleran cekaman kekeringan pada kacang tanah dan kedelai (Rahayu et al. 2005; Widoretno et al. 2003a), resisten terhadap S. rolfsii pada kacang tanah (Yusnita et al. 2005) dan resisten terhadap
Septoria glycines pada kedelai (Song et al. 1994).
Efektivitas seleksi in vitro ditentukan dengan keberhasilan menghambat pertumbuhan sel/jaringan normal yang tidak diinginkan dan memproliferasikan sel/jaringan varian yang diinginkan menggunakan agens penyeleksi (selective agents) tertentu. Seleksi in vitro dengan menggunakan media selektif polietilena glikol (PEG) telah dilakukan untuk mengembangkan galur yang toleran cekaman kekeringan (Rahayu et al. 2005; Widoretno et al. 2003a) dan dengan filtrat kultur
S. rolfsii untuk mendapatkan kacang tanah resisten terhadap infeksi S. rolfsii
(Yusnita et al. 2005). Dalam penelitian tersebut, seleksi in vitro hanya dilakukan terhadap embrio somatik (ES) selama satu siklus seleksi (tiga bulan per siklus seleksi). Sebagian besar tanaman kedelai hasil seleksi in vitro dengan PEG atau kacang tanah dengan filtrat kultur S. rolfsii masih peka terhadap cekaman kekeringan atau rentan terhadap infeksi S. rolfsii (Widoretno et al. 2003b; Yusnita
et al. 2005). Diduga periode seleksi atau kondisi selektif yang digunakan belum cukup efektif untuk menghambat sel/jaringan normal dan memproliferasi sel/jaringan varian.
toleran cekaman PEG dan filtrat kultur. Dalam seleksi ganda, sel/jaringan pertama-tama diseleksi dalam media dengan penambahan PEG. Kalus embriogen dan ES yang insensitif PEG selanjutnya diseleksi dalam media dengan penambahan filtrat kultur untuk mendapatkan varian sel/jaringan yang insensitif cekaman PEG dan filtrat kultur. Sel/jaringan varian hasil seleksi ganda tersebut diharapkan mempunyai sifat toleran terhadap cekaman kekeringan dan resisten terhadap infeksi S. rolsii.
Efektivitas seleksi berulang dan seleksi ganda untuk mendapatkan ES varian tersebut perlu dievaluasi. Menentukan pengaruh seleksi berulang dan seleksi ganda terhadap keberhasilan mengisolasi ES yang insensitif PEG atau filtrat kultur merupakan langkah pertama yang harus dilakukan. Percobaan dilakukan untuk mengevaluasi efektivitas seleksi berulang dalam media dengan PEG untuk meningkatkan frekuensi didapatkannya ES kacang tanah yang insensitif cekaman PEG serta seleksi ganda dalam media denga n PEG dan dengan filtrat kultur untuk meningkatkan frekuensi didapatkannya ES kacang tanah yang insensitif sekaligus pada cekaman PEG dan filtrat kultur. Regenerasi tanaman kacang tanah R0 dari hasil seleksi dan evaluasi respons tunas R0 terhadap cekaman PEG atau filtrat kultur S. rolfsii juga dilakukan untuk menentukan tingkat keberhasilan mendapatkan tanaman varian dari ES kacang tanah hasil seleksi berulang dan seleksi ganda.
Bahan dan Metode
Induksi ES Kacang Tana h
(Murashige & Skoog 1962) dengan penambahan pikloram 16 µM (media MS-P16), campuran vitamin dan asam amino (glisin, tiamin, piridoksin, dan niasin) 0.1 mg/l, sukrosa 2%, dan agar 8 g/l (Sulichantini 1998; Edy 1998). Kalus embriogen dan ES yang didapat diproliferasikan dalam media MS-P16 secara terus- menerus dan selama periode proliferasi diinkubasikan dalam kondisi gelap. Setiap bulan sekali eksplan yang ditanam dipindahkan ke media regenerasi yang masih segar. Sub kultur eksplan dilakukan terus menerus sampai terbentuknya ES primer. Untuk menginduksi pembentukan ES sekunder, eksplan ES primer disubkultur lebih lanjut dan terus menerus dalam media MS-P16. Kultur kalus embriogen dan ES sekunder yang telah berumur enam bulan digunakan dalam percobaan seleksi in vitro berulang dan seleksi ganda.
Seleksi Berulang dalam Media Selektif dengan Penambahan PEG
Ke dalam botol kultur dengan volume 150 ml dituangkan media MS-P16 cair dengan penambahan PEG 15% (media selektif) sebanyak 25 ml. Di atas permukaan media cair diambangkan busa sintetik dan satu lembar kertas saring untuk mencegah agar eksplan kalus embriogen dan ES yang ditanam tidak tenggelam. Konsentrasi PEG 15% yang ditambahkan merupakan konsentrasi sub-letal (Rahayu et al. 2005) yang menghambat proliferasi ES kacang tanah = 95% dibandingkan proliferasi dalam media tanpa PEG (PEG 0%). Media selektif disterilkan dengan suhu hingga 121oC pada tekanan 17.5 psi selama 20 menit.
percobaan berikutnya dan untuk percobaan seleksi ganda dalam media yang mengandung filtrat kultur. Sebagian ES Pi-I yang ada dikecambahkan untuk membentuk planlet Pi-I.
Sebagian ES Pi-I yang tersisa digunakan sebagai eksplan dalam siklus II seleksi berulang selama tiga bulan, menggunakan media selektif MS-P16 dengan penambahan PEG 15%. Kalus embriogen dan ES yang mampu bertahan hidup dari siklus II seleksi berulang diproliferasi dalam media MS-P16 tanpa PEG untuk meningkatkan biomasanya dan selanjutnya disebut sebagai ES yang PEG insensitif hasil seleksi siklus II (Pi-II). Setelah proliferasi, ES Pi-II yang didapat sebagian digunakan untuk evaluasi respons ES Pi-II terhadap cekaman PEG dan filtrat kultur dalam percobaan berikut dan sebagian yang lain dikecambahkan untuk mendapatkan planlet Pi-II.
Seleksi Ganda dalam Media dengan PEG dan Filtrat Kultur S. rolfsii
Penyiapan filtrat kultur S. rolfsii dilakukan sebagaimana yang telah dilaporkan sebelumnya (Yusnita et al. 2005). Isolat S. rolfsii ditumbuhkan dalam media potato dextose agar (PDA) padat dan kultur cendawan umur 7 hari dipotong-potong dengan ukuran 0.5 x 0.5 cm. Potongan agar dengan hifa cendawan S. rolfsii ditanam dalam media MS padat dengan penambahan campuran (glisin, tiamin, piridoksin, dan niasin) 0.1 mg/l, sukrosa 2%, dan agar 8 g/l. Setelah membentuk sklerotia (± 14 hari), media bersama cendawannya disterilkan dengan autoklaf pada suhu 1210, tekanan 17,5 psi selama 20 menit. Filtrat kultur S. rolfsii yang didapat disaring dan digunakan sebagai agens penyeleksi. Media selektif disiapkan dengan menambahkan 30% filtrat kultur S. rolfsii ke dalam media MS-P16 dan disterilisasi dengan menggunakan autoklaf. Berdasarkan penelitian sebelumnya, 30% filtrat kultur S. rolfsii merupakan konsentrasi sub- letal yang dapat menekan perkembangan ES kacang tanah = 95 % (Yusnita et al. 2005).
(siklus I dengan agens penyeleksi PEG dan siklus II dengan filtrat kultur, masing-masing selama 3 bulan) diproliferasi dalam media MS-P16 untuk meningkatkan biomasa dan selanjutnya disebut sebagai ES yang insensitif terhadap PEG dan filtrat kultur (PFi-I). Setelah proliferasi, ES PFi-I yang didapat sebagian digunakan untuk evaluasi respons ES PFi-I terhadap cekaman PEG dan filtrat kultur dalam percobaan berikut dan sebagian yang lain dikecambahkan untuk mendapatkan planlet PFi-I.
Respons ES Hasil Seleksi terhadap Cekaman PEG dan Filtrat Kultur S. rolfsii
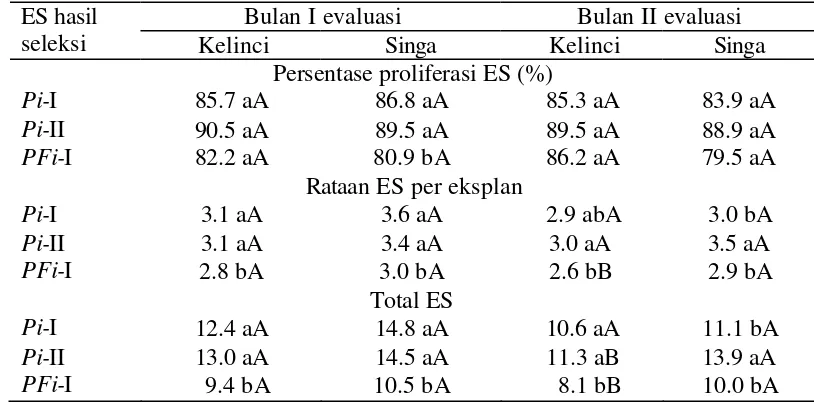
Percobaan dilaksanakan dengan menggunakan Rancangan Acak Lengkap dengan dua faktor perlakuan (populasi ES Pi-I, Pi-II, PFi-I, ES tanpa diseleksi dalam media selektif, dan kultivar kacang tanah). Efektivitas seleksi berulang selama satu atau dua siklus seleksi dalam media dengan PEG 15% untuk mengisolasi ES yang insensitif terhadap PEG ditentukan dengan mengamati respons ES Pi-I, Pi-II, dan PFi-I hasil seleksi terhadap cekaman PEG. Eksplan kalus embriogen yang diproliferasikan dari ES Pi-I, Pi-II, dan PFi-I (masing-masing 50 eksplan atau 5 eksplan per botol) ditanam dalam media selektif dengan PEG 15% dan pertumbuhan kalus embriogen yang dikulturkan diamati setelah dua bulan. Persentase keberhasilan ES hasil seleksi untuk bertahan hidup dalam media selektif, rataan jumlah ES yang terbentuk per eksplan, dan total ES yang didapat diamati selama periode pengamatan.
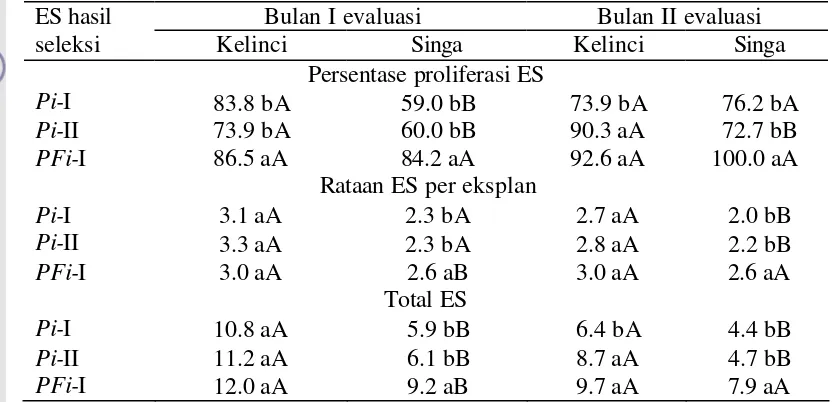
Efektivitas seleksi ganda (siklus I dalam media PEG 15% dan siklus II dalam media filtrat kultur S. rolfsii 30%) untuk mengisolasi ES yang insensitif terhadap PEG dan filtrat kultur S. rolfsii ditentukan dengan mengamati respons ES
Respons Planlet terhadap Cekaman PEG dan Filtrat Kultur S. rolfsii
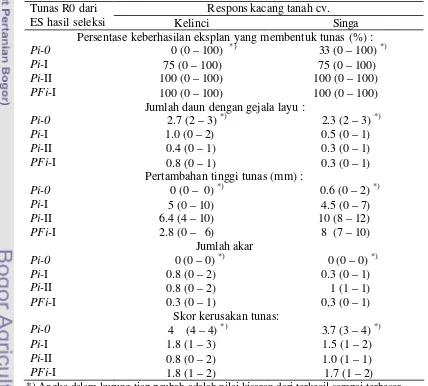
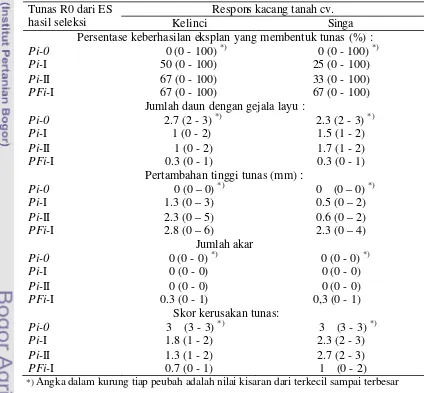
Embrio somatik yang insensitif dari seleksi berulang (ES Pi-I dan Pi-II) dan seleksi ganda (ES PFi-I) dikecambahkan dan diregenerasikan menjadi planlet
Pi-I, Pi-II, dan PFi-I dengan metode yang telah dilaporkan sebelumnya (Yusnita
et al. 2005). Tunas planlet yang didapat dari perkecambahan ES insensitif selanjutnya diperbanyak secara vegetatif. Sebagian tunas digunakan untuk menguji responsnya terhadap cekaman PEG atau cekaman filtrat kultur cendawan. Sebagian yang lain diakarkan dan diaklimatisasi untuk ditanam di rumah kaca.
Setek pucuk (dua buku, dengan 3-4 daun) dari planlet Pi-I, Pi-II, dan PFi-I yang dihasilkan dari percobaan sebelumnya dipisahkan dan ditanam dalam media selektif yang mengandung PEG 15% atau filtrat kultur S. rolfsii 30%. Pengamatan perkembangan setek pucuk yang meliputi jumlah daun layu, pertambahan tinggi tunas, jumlah akar, dan tingkat kerusakan tunas, dilakukan 4 minggu setelah penanaman dalam media selektif. Setek pucuk dari kecambah kacang tanah yang tidak melewati tahapan seleksi in vitro digunakan sebagai pembanding (Pi-0).
Skor kerusakan tunas yang ditanam dalam media dengan PEG ditentukan menggunakan kriteria: skor 0 = eksplan mengalami kerusakan < 5%, skor 1 = eksplan mengalami kerusakan antara 5% – 25%, skor 2 = kerusakan antara > 25% – 50% , skor 3 = kerusakan antara > 50% – 75%, dan skor 4 = kerusakan >75%. Untuk skor kerusakan tunas yang ditanaman dalam media dengan filtrat kultur S. rolfsii ditentukan menggunakan kriteria : skor 0 – jika pucuk tunas sehat dan setek berakar; skor 1 – pucuk sehat tetapi tidak berakar; skor 2 – pucuk sehat dengan 1 atau 2 daun menguning (mengering); dan skor 3 – pucuk mengering, daun nekrosis atau mengering dan tunas mati.
Hasil
Embrio Somatik Hasil Seleksi In Vitro Berulang pada Cekaman PEG
menunjukkan bahwa siklus seleksi dan cv. kacang tanah berpengaruh nyata terhadap persentase proliferasi ES, rataan ES per eksplan, dan total ES (Tabel 1).
Kultivar Singa dan Kelinci yang diseleksi dalam PEG selama dua siklus cend