APLIKASI MARKA ISOENZIM, RAPD, DAN AFLP UNTUK IDENTIFIKASI V ARIABILITAS GENETIK TANAMAN MANGGIS (Garcinia mangostana)
DAN KERABAT DEKATNYA
Soaloon Sinaga1, Sobi.-2, Roedby Poerwanto2, Hajrial Aswidinnoo.-2, Dedy Duryadi2, Resmitasari3, Rudy Lukman4, dan Roswita Amelia5
J Mahasiswa S3 Program Studi Agronomi, Sekolah Pascasarjana IPB; 2Staj Pengajar IPB; 3Sta/ PKT Kebun Raya Bogor; 4StajBISI International dan SEAMEO Biotrop; 5StajSEAMEO Biotrop
ABSTRACT
This research was aimed to study genetic variability among mangosteen accession and other Garcinia sp.
based on isozymes, RAPD, AFLP, and their combinations. Genetic variability and relationships among
several mangosteen and other Garcinia sp were established by using four isozymes, seven RAPD primers,
and three AFLP primer combinations. The level of polymorphism as revealed by isoenzyme, RAPD, AFLP was 88.9 %, 100 %, and 100% respectively. The dendrogram is built based on combined data from
isozymes, RAPD, and AFLP analysis separate clusters mangosteen and other Garcinia sp. The combined
RAPD with AFLP and combined isozymes, RAPD, and AFLP data show that G. mangostana is a close
relative of G. malaccensis, G. porrecta, G. celebica, and G. hombroniana.
Key words: Genetic variabiHty, mangosteen, isozymes, RAPD, AFLP
PENDAHULUAN
Manggis (Garcinia mangostana L.) yang dikenal sebagai Queen of Tropical Fruits merupakan salah satu komoditi ekspor unggulan Indonesia. Peningkatan nilai ekonomis manggis dewasa ini, mendorong perlunya peningkatan produksi. Hal ini dicapai dengan kultur teknis yang lebih maju dan penggunaan bibit unggul dari hasil pemuliaan yang terarah dan strategis.
Program perbaikan genetik tanaman manggis sangat bergantung pada sumber keragaman genetik. Indonesia merupakan salah satu pusat persebaran manggis di Asia Tenggara. Ekplorasi, identifikasi dan karakterisasi manggis dan kerabat dekatnya penting dilakukan untuk memperoleh informasi sumber keragaman genetik bam guna perbaikan genetik dan peningkatan produksi manggis. Studi variasi genetik diarahkan langsung untuk melacak keberadaan variasi aksesi manggis Indonesia (Drew 1997).
Manggis bersifat apomiksis obligat, biji tidak berasal dari fertilisasi dan diduga mempunyai keragaman genetik sempit, sehingga diperkirakan keberadaan manggis di alam satu spesies. Kenyataan di lapang menunjukkan adanya keragaman tanaman manggis, kemungkinan disebabkan faktor lingkungan maupun faktor genetik akibat mutasi alami sejalan dengan sejarah tanaman manggis yang telah berumur ribuan tabun (Ramage et at. 2004). Di Kaligesing, Kabupaten Purworejo, Jawa Tengah diidentifikasi adanya bibit hasil sambungan dan tanaman produktif yang mempunyai trubus dengan warna hijau muda, merah, dan coklat (Supriyanto et at. 1999). Evaluasi keragaman pohon manggis pada sentra produksi manggis di Jawa dan Lombok dengan analisis isoenzim yang dilakukan oleh Supriyanto et al. (1999), menunjukkan minimal ada tiga klon manggis .. Berdasarkan morfologi manggis asal Sumatera Barat dan Sumatera Selatan terdiri atas tujuh klon (Mansyah et al. 1994). Manggis Sumatera Barat dengan analisis isoenzim glucose phosphateisomerase (OPI), 14 sampel yang diujikan menunjukkan pola pita yang sama meskipun secara fenotip bervariasi, dengan kata lain variabilitas genetiknya sempit, tetapi variabilitas fenotipnya luas (Mansyah et at. 1999). Sejumlah analisis DNA dan RNA juga memperlihatkan variasi diantara populasi manggis (Ramage et al. 2004). Kandungan DNA manggis dan kerabat dekatnya (nama lokal Thai, chamuang, mahput, pawa, dan somkhang) menggunakan flow cytometry menunjukkan perbedaan. Selanjutnya, sekuensing DNA genom dengan primer spesifik menunjukkan· manggis lebih dekat kekerabatannya dengan pawa diikuti somkhang dan mahput (Te-Chato dan Lim 2000). Analisis genetik dengan teknik mutakhir terhadap jenis yang lebih luas memungkinkan identifikasi tetua jantan untuk hibridisasi dengan manggis sebagai tetua betina (Osman dan Abdul 2006).
Tanaman manggis mempunyai siklus hidup yang panjang, sehingga studi genetik dengan uji keturunan dan persilangan solit dilakukan, karena itu estimasi variabilitas genotipe menggunakan penanda morfologi dan penanda molekuler (DNA) penting dilakukan. Penggunaan penanda molekuler (RAPD, RAF, dan AFLP) dan isoenzim mempunyai kontribusi penting bagi pemulia tanaman dalam penanganan apomiksis (Ramage et al. 2004). Teknik isoenzim dan RAPD telah dilakukan untuk mengkarakterisasi tanaman kentang (Collares セエ@ al. 2004). Teknik AFLP digunakan untuk pembuatan sidik jari DNA (Vos et al. 1995), untuk menguji hubungan antara polimorfisme molekuler dan penampilan hibrid pada jagung (Ajmone-Marsan et at. 1998). Prosiding Seminar NasionaI Hasil Penelitian yang Dibiayai oleh Hibah Kmnpetitif
Bogor, 1-2 Agustus 2007
Analisis variabilitas tanaman manggis dan genotyping anggota genus Garcinia lainnya penting guna memperoleh beberapa calon tetua dengan melakukan hibridisasi dengan manggis. Penelitian
ini bertujuan mendapatkan 1) infonnasi keragaman genetik dan kekerabatan antara manggis dengan kerabat dekatnya sekaligus mengetahui tetua manggis dan 2) merekomendasikan metode karakterisasi manggis yang paling efektif.
BAHAN DAN METODE Tempat dan Waktu Penelitian
Penelitian dilakukan di Laboratorium Bioteknologi dan Pemuliaan Pohon BIOTROP Bogor, Laboratorium Molekuler Pusat Studi Sumberdaya Hayati dan Bioteknologi IPB, serta di Laboratorium PKBT IPB, Bogor. Penelitian dilakukan dari bulan Juni 2006 sampai dengan Agustus 2007.
Bahan Penelitian
Sample daun tanaman manggis dan kerabat dekatnya dikoleksi dari seluruh Indonesia meliputi 20 provinsi dan Koleksi Kebun Raya Bogor serta taman wisata Mekarsari. Aksesi yang dipakai untuk analisis isoenzim adalah 33, sedangkan untuk analisis RAPD dan AFLP adalah 18 aksesi.
Metode
Analisis Isoenzim mengikuti prosedur Soltis dan Soltis (1989). Enzim yang dianalisis adalah peroxidase (PER), Asam Fosfatase (ACP), Malat Dehidrogenase (MDH), dan Esterase (EST). Analisis molekuler, ekstraksi DNA mengikuti prosedur CT AB oleh Doyle and Doyle (1987) dengan beberapa modifikasi. Teknik RAPD mengikuti metode William et al. (1990). Tujuh random primer yang dipakai adalah SBH12 (S'-ACGCGCATGT-3'), SBH13 (S'-GACGCCACAC-3'), SBH19 (S'-CTGACCAGCC-3'), OPA14 (S'-TCTGTGCTGG-3'), OPA16 (S'-AGCCAGCGAA-3'), OPA17 (S'-GACC-GCTTGT-3'), dan OPA18 (5'-AGGTGACCGT-3'). Analisis AFLP mengikuti protokol yang dikembangkan oleh Vos et al. (1995) yang dimodiflkasi Schwarz et al. (2000). Sekuen adaptor EcoRI adalah 5' -CTCGTAGACTGCGTACC-3', 3' -CTGACGCATGGTTAA-5', dan sekuen dari adaptor MseI adalah 5' -GACGATGAGTCCTGAG-3', 3'-TACTCAGGACTCAT -S'. Primer untuk ampliflkasi preselektif adalah EcoRI+A dan MseI+C. Selanjutnya, amplifikasi selektif menggunakan primer EcoRl + ANN dan Mse 1 +CNN. Elektroforesis PAGE menggunakan 5 % gel polyacrilamide pada tegangan 2 SOO Volt selama 4 jam. Data awal dikoleksi dengan software ABI PRISM1M v.I. 1 dan dianalisis dengan software GENESCAN 1M v.2.1 (Applied Biosystems).
Pita dari teknik isoenzim, RAPD, dan AFLP diterjemahkan menjadi data biner (diberi nilai 1 bila ada pita dan 0 bila tidak ada pita). Data ini digunakan untuk menyusun matriks kesamaan genetik berdasarkan rumus Nei dan Li (1979) dengan metode UPGMA (Unweighted Pair-Group Method Aritmetic) menggunakan program NTSys (Numerical Taxonomy and Multivariate System) versi 2.02 (Rolf 1998).
BASIL DAN PEMBAHASAN
Analisis Variabilitas dengan Penanda Isoenzim
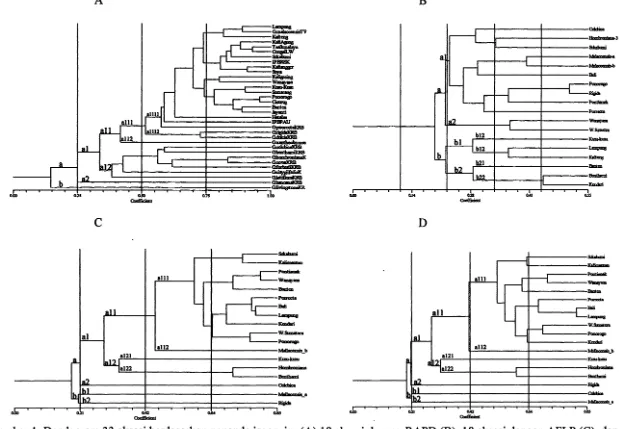
Fragmen 33 aksesi manggis dan kerabat dekatnya dengan empat isoenzim berjumlah 27 pita (Tabel 1), mampu mengungkap keragaman ke-33 aksesi dengan polimomsme tinggi (88.9%). Analisis kluster menghasilkan dendrogram khas yang memisahkan tanaman manggis terhadap kerabat dekatnya pada indeks ketidaksamaan 42.8 %, kecuali G. malaccensis (Gambar lA). Pengelompokan manggis dan kerabat dekatnya terbentuk pada koefisien kemiripan 14-96%. Kelompok manggis mempunyai tingkat kemiripan yang lebih tinggi (57.2-96%) dibanding kerabat dekatnya (14-83 %). Dengankoeflsien kesamaan S7.2 % diperoleh 11 kelompok aksesi. Keragaman ini tennasuk tinggi untuk tanaman opomiksis obligat, dibanding dengan tanaman Taraxacum (19 %) (Ford dan Richards, 1985). Variasi yang muncul pada apomiktik terjadi dengan kecepatan yang lebih besar dari pada mutasi (Hughes dan Richards, 1985). Aksesi
G. celebica dan G. hombroniana yang diduga sebagai tetua manggis justru pada kemiripan genetik yang lebih rendah (33 %). Richards (1990), menyatakan tanaman manggis merupakan allotetraploid (2n=90)
turunan
dari G. malaccensis (2n=42) dan G. hombroniana (2n=48), dengan morfologi intennediet kedua spesies diploid tersebut. Nilai korelasi matriks kesamaan MxComp sebesar r=O.914. Artinya, dendrogram yang dihasilkan dengan goodness of.fit sangat sesuai menggambarkan pengelompokan tersebut di atas (Rolf 1998).f_ No Kar. 2 3 4 5 6 7 8 9 10
Tabel 1. Jumlah Pita dan Tingkat Polimorfisme 5 Isoenzim pada 33 Aksesi Manggis dan Kerabat No 1 4 7 9 Dekatnya Isoenzim Esterase Peroksidase Acid phosphatase Malat dehidrogen JumlahPita 10 7 5 5 Pita Polimorfik 10 (100%)
6 (85.%) 4 (80%) 4 (80%) Pita Monomorfik
o
1 11
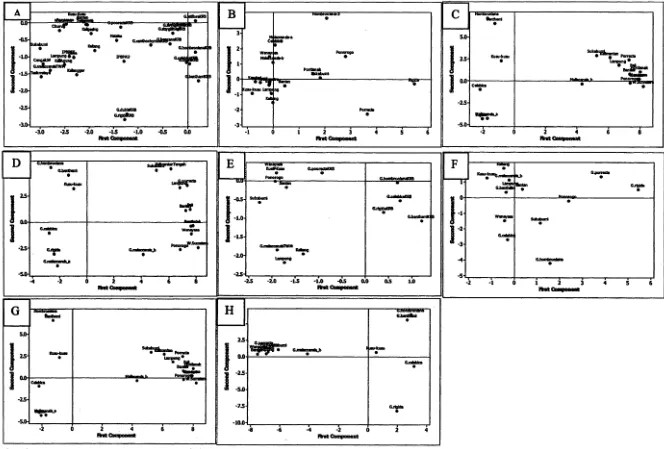
Akumulasi perseritasi tiga komponen utama pertama mewakili 59.94 % keragaman (Tabel 2) dari total 100 % pada 27 karakter. Keragaman 70 % dari 27 karakter bam dapat diperoleh dari 5 komponen utama. Terdapat 22 karakter yang tidak terlalu berpengaruh terhadap
ploting 33 aksesi tersebut.
Tabel 2. Nilai Ciri Komponen Utama Isoenzim, RAPD, AFLP, Kombinasi RAPD dan AFLP, serta Kombinasi Isoenzim dan AFLP
Isoenzim RAPD AFLP· RAPD+AFLP Izoenzim+AFLP
NC (%) K NC (%) K NC (%) K NC (%) K NC (%) K
10.259 37.997 37.997 32.080 18.331 18.331 18.508 22.122 22.122 19.888 18.611 18.611 20.971 22.90 22.90
4.188 15.512 53.510 15.146 8.665 26.986 8.638 10.324 32.446 10.254 9.595 28.206 11.861 12.90 35.80
1.736 6.432 59.942 12.757 7.290 34.276 8.057 9.630 42.077 9.958 9.318 37.525 10.126 11.00 46.80
1.543 5.716 65.659 11.446 6.540 40.817 6.597 7.885 49.962 8.287 7.755 45.280 9.349 10.20 57.00
1.471 5.450 71.109 11.338 6.479 47.297 6.424 7.678 57.461 7.912 7.404 52.684 8.192 8.90 66.00
10.096
5.769 53.066 5.915 7.070 64.711 7.222 6.758 59.442 7.331 8.00 74.00
9.817
5.610 58.676 5.334 6.376 71.087 6.963 6.516 65.958
9.235
5.277 63.953 6.546 6.126 72.084
7.628
4.359 68.313
6.994
3.997 72.309
Ket.: Kar = karakter, NC = Nilai eiri, K = kumulatif
Hasil ekstraksi komponen 1 vs 2 membentuk 3 kelompok utama (Gambar 3A). Analisis ini mendukung pengelompokan pada dendrogram. Aksesi G. hombroniana dan G. celebica yang diduga sebagai tetua manggis cenderung selalu dalam kelompok yang sama, sedangkan G. malaccensis selalu mengelompok dengan manggis.
Analisis Variabilitas dengan Penanda RAPD
Semua primer acak mampu mengamplifikasi semua DNA aksesi (Tabel 3) dengan total 69 pita DNA. Jumlah pita tiap primer bervariasi antara 2-13 pita dengan rata-rata 10 pita DNAlsampel. Primer OPA 14 menghasilkan pita paling sedikit (6 pita), sedangkan pita terbanyak dihasilkan oleh primer SBH 19 (13 pita). Ukuran pita yang· diamplifikaSi berkisar antara 100-1600 pb. Jumlah pita yang dihasilkan oleh tiap primer tergantung pada sebaran situs yang homolog dengan sekuen primer pada genom. Semria primer menghasilkan tingkat polimorfis tinggi yaitu 69 pita (100 %). Perbedaan jumlah dan ukuran pita menentukan tingkat keragaman genetik aksesi manggis dan kerabat dekatnya.
Tabe13. Jumlah pita hasil amplifikasi tujuh primer dengan teknik RAPD
Primer. Ukuran pita (Pb). Jumlah pita monomorfik Jumlah pita polimorfik
SBH 12 250 - 1600 0 9
SBH 13 200 -1600 0 12
SBH 19 150 -1400 0 13
OPA 14 200 -1100 0 6
OPA 16 . 100 - 1500 0 10
OPA 17 100-1800 0 10
OPA 18 100 -1500 0 9
Total 0 69.(100%)
Jumlah pita 9 12 13 6 10 10 9
69
Dendrogram 18 akSesi manggis dan kerabat dekatnya dengap. teknik RAPD mempunyai tingkat kemiripan 19.6-50.8 % (Gambar IB). Teknik ini tidak mengelompokkan aksesi manggis dalam satu kelompok dan kerabat dekatnya tersebar diantara aksesi manggis. Analisis RAPD ini tidak kongruen dengan penanda isoenzim dan. AFLP yang mampu memisahkan grup manggis dengan kerabat dekatnya. Diduga pita-pita yang teramplifikasi bu1cim merupakan penciri yang mampu menunjukkan tingkat kekerabatan antar 18 aksesi yang diuji. Penggunaan primer yang lebih banyak dengan tingkat polimorfisme tinggi diharapkan dapat memberikan pengelompokan yang lebih komprehensif. Chen et al. (2000), menyatakan data molekuler sangat tergantung pada pemilihan primer yang digunakan. Berbeda dengan penanda lainnya dendrogram yang dihasilkan
Prosiding Seminar Nasional Hasil Penelitian yang Dibiayai oleh Hibah Kompetitif Bogor, 1-2 Agustus 2007
teknik. RAPD mampu mengelompokkan G. hombroniana dan G. celebica dengan tingkat kemiripan 30.8 %. Selanjutnya, G. malaccensis_a dan G. malaccensis_b berada pada kelompok yang sama dengan tingkat kemiripan 39.5 % padabaJ pada dua penanda lainnya kedua aksesi pada kelompok berbeda. Dendrogram penanda RAPD mempunyai nilai korelasi matriks r=0.587 dengan goodness of fit sangat tidak sesuai.
Tiga komponen utama pertama penanda RAPD mempunyai persentase akumulasi 34.27 % keragaman (Tabel 2). Keragaman 70 % dari 175 karakter baru dapat diperoleh dari 10 komponen utama. Terdapat 165 karakter yang tidak terlalu berpengaruh terhadap ploting 18 aksesi yang dianalisis. HasH ekstraksi komponen 1 vs 2 (Gambar 3B) membentuk 4 kelompok utama. Analisis komponen utama memberikan gambaran yang berbeda pengelompokan pada dendrogram (Gambar IB).
Analisis Variabilitas dengan Penanda AFLP
Analisis AFLP 18 aksesi manggis dan kerabat dekatnya dengan tiga kombinasi primer: ACC_CAG, ACT_CAA, dan ACT_CAC menghasilkan 220 pita yang seIuruhnya (100 %) polimorfik. Jwnlah pita yang dihasilkan dari tiap kombinasi primer bervariasi antara 19-94 pita. Rata-rata 73.3 pita tiap sampel/primer. Primer ACT_CAA menghasilkan pita polimorfik terbanyak (94 pita) diikuti primer ACT_CAA 70 pita dan primer ACC_CAG 56 pita. Ukuran fragmen yang diamplifikasi berkisar antara 50-500 pasang basa (Pb).
Tingkat kemiripan 18 aksesi manggis dan kerabat dekatnya dengan penanda AFLP berkisar antara 18.48-80 % (Gambar lC). Penanda AFLP memisahkan aksesi manggis pada indeks kesamaan 45.3 %, kecuali kusu-kusu. Kelompok manggis mempunyai tingkat kemiripan yang Iebih tinggi (58-81 %) dibanding kerabat dekatnya (18-71 %). HasH analisis kluster membagi 18 aksesi menjadi 7 kelompok pada kemiripan 58 %. Aksesi G. malaccensis _ b, dan
G. hombroniana yang diduga sebagai tetua manggis berada pada kemiripan genetik 45.3 % dan 29%. Berdasarkan dendrogram AFLP hipotesis tersebut dapat diterima. Nilai r=O.978, artinya dendrogram yang dihasilkan dengan goodness of fit sangat sesuai untuk menggambarkan pengelompokan tersebut.
Tiga komponen utama pertama mempunyai akuinulasi 42.07 % keragaman (Tabel 2). Keragaman 70 % dari 607 karakter diperoleh dari 7 komponen utama. Terdapat 600 karakter yang tidak terlalu berpengaruh terhadap ploting 18 aksesi. Hasil ekstraksi komponen 1 vs 2 membentuk 6 kelompok utania (Gmbar 2C). Analisis komponen utama mendukung pengelompokan pada dendrogram (Gambar 1 C).
Analisis Variabilitas Menggunakan Kombinasi RAPD dan AFLP , , Tingkat kemiripan 18 aksesi manggis dan kerabat dekatnya berkisar antara 19.32-80.8 % (Gambar ID). Kombinasi kedua penanda memisahkan aksesi manggis pada indeks kesamaan 42 %. Kelompok manggis mempunyai'tingkat kemiripan yang lebih tinggi (53-81 %) dibanding kerabat dekatnya (19-60 %). Analisis kluster membagi 18 aksesi menjadi 7 kelompok (53 %). G. malaccensis, dan G. hombroniana yang diduga sebagai tetua manggis berada pada kemiripan genetik 42 % dan 28 %. Berdasarkan dendrogram kombinasi penanda AFLP dan RAPD hipotesis tersebut dapat diterima. Analisis MxComp menghasilkan nilai r=0.972 dengan goodness of fit
sangat sesuai untuk menggambarkan pengelompokan.
Tiga komponen utama pertama mempunyai nilai akumulasi 37.5 % keragaman. Keragaman 70 % dari 777 karakter baru dapat diperoleh dari 8 komponen utama. Terdapat 769 karakter yang tidak terlalu berpengaruh terhadap ploting 18 aksesi tersebut. Ekstraksi komponen 1 vs 2 membentuk 4 kelompok Utama (Gambar 3D). Analisis komponen Utama mendukung pengelompokan pada dendrogram (Gambar ID).
Analisis Variabilitas Menggunakan Kombinasi Isoenzim dan AFLP
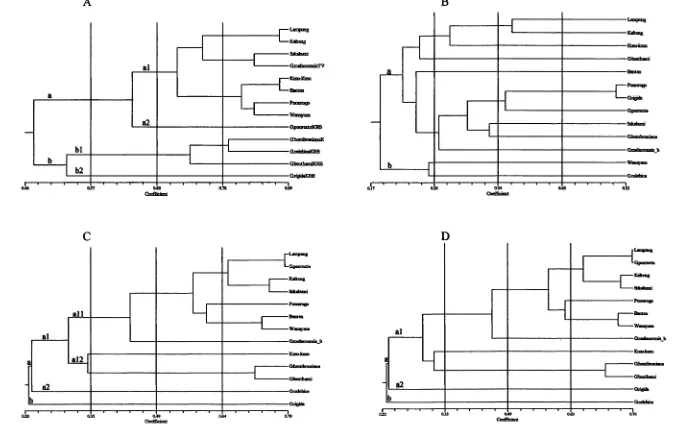
Kombinasi penanda diharapkan mengungkap kekerabatan antat aksesi dengan lebih baik karena karaktemya Iebih banyak dan dari region genom yang berbeda. Jarak genetik 13 aksesi manggis dan kerabat dekatnya dengan kombinasi isoenzim dan AFLP berkisar 23-76.1 % (Gambar 2D). Kombinasi kedua penanda memisahkan aksesi manggis dengan indeks kemiripan 30.8 %. Kelompok manggis mempunyai tingkat kemiripan yang Iebih tinggi (30.8-76 %) dibanding kerabat dekatnya (23-70.3 %). Analisis kluster membagi 13 akSesi menjadi 6 kelompok (58.3 %). Dilihat dari kekerabatannya penanda isoenzim Iebih sesuai dalam pengelompokan. Kerabat dekat sudah'memisah pada 64 % (G. porrecta) kecuali G. Malaccensis,
G. Hombroniana, dan G. celebica dalam kelompok yang, sama (79 %), G. rigida merupakan
kerabat terjauh (47.5 %) dibanding manggis. Sebaliknya pada dendrogram AFLP dan kombinasi kedua penanda G. porrecta masuk kelompok manggis (71.6 % dan 75.5 %), kusu-kusu masuk kerabat dekat (30 % dan 30.8 %). G. hombroniana dan G. benthami dalam kelompok yang sama (71 %), sedangkan kerabat terjauh G. rigida (20.7 %) pada penanda AFLP dan G. celebica
(22 %) pada kombinasi kedua penanda. G. Malaccensis dan G. homhroniana yang diduga sebagai tetua manggis berada pada kemiripan genetik 45.9 % dan 30.8 %. Hipotesis tersebut berdasarkan dendrogram kedua penanda dapat diterima. Nilai r=0.975, dendrogram mempunyai
goodness of fit sangat sesuai. .
Tiga komponen utama pertama mempunyai nilai akumulasi yang mewakili 46.8 % keragaman (Tabel 2). Keragaman 70 % dari 634 karakter diperoleh dari 6 komponen utama. Terdapat 628 karakter yang tidak terlalu berpengaruh terhadap ploting 13 aksesi yang dianalisis. Ekstraksi komponen 1 vs 2 membentuk 6 kelompok utama (Gambar 3H). Analisis komponen utama mendukung pengelompokan pada dendrogram (Gambar 2D). Pengelompokan 13 aksesi pada dendrogram masing-masing penanda dengan tingkat kemiripan 55 % berbeda dengan pengelompokan hasil komponen utama, kecuali oleh AFLP. Secara umum dari 13 aksesi semua penanda dan kombinasinya memisahkan manggis dengan kerabat dekatnya, kecuali RAPD.
AFLP berkontribusi besar terhadap kluster kombinasi penanda. Secara konsisten nilai r tetap tinggi (> 90%) dan mampu memisahkan aksesi manggis dengan kerabat dekatnya. Hipotesis tetua manggis G. malaccensis dan G. homhroniana dapat diterima dengan pembuktian penanda AFLP dan kombinasinya dengan isoenzim. Persentase polimorfik yang tinggi bukan hal yang umum bagi tanaman manggis sebagai tanaman apomiksis obligato Kemungkinan hal ini
disebabkan oleh G. mangostana dihasilkan tidak dari persilangan tunggal diantara tetuanya. Hibridisasi berulang diantara tetua manggis memungkinkan munculnya variasi genetik yang lebih luas diantara tetuanya (ipbfruit@indo.net.id). Variasi yang tinggi diantara genotipe manggis merupakan potensi genetik untuk menghasilkan tipe unggul. Hal ini dapat dilakukan dengan metode seleksi Massa pada beberapa tanaman untuk menghasilkan varietas baru.
KESIMPULAN
Variabilitas genetik dan kekerabatan diantara tanaman manggis dan kerabat dekatnya dapat diungkapkan dengan empat isoenzim, tujuh primer RAPD, dan tiga kombinasi primer AFLP dengan tingkat polimorfisme masing-masing 88.9 %, 100 %, dan 100 %. G. mangostana
sebagai tanaman apomiksis obligat justru mempunyai variasi genetik yang luas. Tanaman manggis relatif dekat dengan G.malaccensis, G. porrecta, G. celebica, and G. hombroniana.
Hipotesis yang menyatakan G. mangostana merupakan hibrid dari G. malaccensis dengan G. homhroniana berdasarkan penanda AFLP dan kombinasinya dengan isoenzim dapat diterima.
Dendrogram berdasarkan penanda RAPD sangat berbeda pengelompokannya dibanding penanda isoenzim dan AFLP. Tingkat kesesuaian isoenzim lebih kongruen dengan AFLP. Penanda isoenzim dan AFLP mempunyai tingkat kesesuaian yang sangat baik dalam menggambarkan keragaman antar aksesi serta mampu mengelompokkan aksesi manggis secara terpisah dari kerabat dekatnya.
UCAPAN TERIMA KASIH
Kepada Kantor Kementerian Riset dan Teknologi melalui program RUSNAS Pengembangan Buah-buahan Unggulan Indonesia di Pusat Kajian Buah-Buahan Tropika (PKBT), LPpM-IPB periode 2000-2015, kepada Kementrian Departemen Pertanian melalui program KKP3t periode 2007, dan kepada PSRPT melalui program DIPA.
DAFfAR PUSTAKA
Ajmone-Marsan, P., P. Castiglioni, F. Fusari, M. Kuiper, and M. Motto. 1998. Genetic diversity and its relationship to hybrid performance in maize as revealed by RFLP and AFLP markers. Theor. Apll. Genet. 98:219-227.
Chen, Z., W. Song, and A. Warren. 2000. Studies on Six Euplotes spp. (Ciliophora: Hypotrichida) Using RAPD Fingerprinting, Including a Comparison with Morphometric Analyses. Acta. Protozool. 39: 209 - 216.
Collares, E. A. S., E. Choer, and A. S. Pereira. 2004. Characterization of potato genotypes using moleculer markers. Pesq. Agropec. Bras., Brasilia 39(9): 871-878.
Prosiding Seminar Nasional Hasil Penelitian yang Dibiayai oleh Hibah Kompetiti! Bogor, 1-2 Agustus 2007
Doyle, J. J. and J. L. Doyle. 1987. A rapid dna isolation procedure for small quentities of fress leaf tissue. Phytochemical Bulletin 19 (1): 11-15.
Drew, R. A. 1997. The Application of Biotechnology to the Conservation and Improvement of Tropical and Subtropical Fruit Species. Seed and Plant Genetic Resources Service. Food and Agriculture Organization of the United Nations. Rome. 92p
Ford, H., and A. J. Richards. 1985. Isozyme variation within and between
Taraxacum
agamospesies in a single locality. Heredity 55: 289-291.
•
Hughes, J. and A. J. Richards. 1985. Isozyme in heritance in diploid
Taraxacum
hybrid. Heredity 54: 245-249.Mansyah, E., Edison Hs., dan M. Winarno. 1994. Eksplorasi dan Studi Keragaman Manggis
(Garcinia Mangostana
L.) di Sumatera Selatan. Laporan Hasil Penelitian Balai PenelitianHortikultura Solok.
Mansyah, E., M. Jawal, A. Lukitariati, dan A. Susiloadi. 1999. Variabilitas genetik tanaman manggis melalui analisis isoenzim dan kaitannya dengan variabilitas fenotipik. Zuriat 10(1):1-9.
Nei, M. and W. H. Li. 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc Natl acad Sci USA 76(10): 5269-5273
Osman, M. and R. M. Abdul. 2006. Mangosteen
Garcinia mangostan
L. Southampton Center for Underutilised Crops, University of Southampton, UK. I69p.Ramage, C. M., L. Sando, C. P. Peace, B. J. Carroll, and R. A. Dew. 2004. Genetic diversity revealed in the apomictic fruit species Garcinia mangostana L. (mangosteen). Euphytica 136(1):1-10.
Richards, A. J. 1990. Studies in Garcinia, dioecious tropical forest trees: the origin of the mangosteen (G
mangostana).
Botanical Journal of The Linn. Soc. 103:103-308.Rolf, F. J. 1998. NTSys-pc. Numerical Taxonomy and Multivariate Analysis Syatem. Version 2.02. Exerter Software. New York.
Schwarz, G., M. Herz, X. Q. Huang, W. Michalek, A. Jahoor, G. Wenzel, and V. Mohler. 2000. Application of fluorescence-based semi-automated AFLP analysis in barley and wheat. Theor. Appl. Genet. 100: 545-551.
Soltis, D. E., and P. S. Soltis. 1989. Isoenzimes in plant biology. Discorides Press. Portland, Oregon. 268p.
Supriyanto, A., A. Muharam, dan B. Hariyanto. 1999. Evaluasi Keragaman Pohon Manggis pada Sentra Produksi di Jawa dan Lombok dengan Analisis Isoenzim. Bul. Plasma Nutfah 5(1):6-10.
Te-Chato, S. and M. Lim. 2000. Improvement of mangosteen micropropogation through meristematic nodular callus formation from in vitro-derived leaf explants. Scientia Horticulturae 86(4): 291-298.
Vos, P., R. Hogers, M. Bleekers, M. Reijans, T. Van de Lee, M. Homes, A. Frijters, J. Pot, J. Peleman, E. Jacobsen, J. Helder, and J. Bakker. 1995. AFLP: a new technique for DNA fingerprinting. Nucl. Acids Res. 23:4407-4414.
William, J. G. K., A. R. Kubelik, K. J. Livak, J. A. Ravalski, and S. V. Tingey. 1990. DNA polimorphisms amplified by arbitrary primer are useful as genetic markers. Nucleic Acid Research 18(22):6531-6535.
"
i'.'
ャセ@
A
セ@
S:
1-1 S·
,(" OQ h!'$!!!..;av
セイ@
セ@
E -.
セセ@
lij
セ@
nセ@ セ@
ァセ@ セGX@
"0'
==
[
セ@セ@
=
セ@
セ@:::: セ@
セ@
=-セ@
セ@
:::: セ@
So
qセ@
=
セ@1
セ@
セ@
セ@
S2
セ@
E
セ@ OJ:lO G.2S Clx:fIicicD1セ@
=セ@
セ@
!:!. Q
セ@
C
セ@
セ@
-{
...
iセ]@
セ@ 1 _
-
..
-ol
I
8112I
c:=::
_ _ bal21 Km>Ioal
-1122 _ _
0IkI0ic0
MIII8ceosis_B
セ@
. ftAc J;;
0.21 セ@ Q64
oJ. B a D all al21 8122 bl2 bl2 b21 b22
...
0ld60i0at alII a112 0A2 0>0ffidaII GA. OM Cddioa_3
--セ@ BIIi-...
RipIo--
- .
セ@ w._ K>II>lam I.ompIIs KoIIq ----fill-
----
セ@-
- ....
BIIi セ@ W.Sualaa-...
--..b
Km>bo>----
RipIo"""'""
MollooomIs_8;J.,
セ@
Gambar 1. Dendogram 33 aksesi berdasarkan penanda isoenzim (A) 18 aksesi dengan RAPD (B), 18 aksesi dengan AFLP (C), dan 18 aksesi berdasarkan gabungan RAPD dan AFLP 18 aksesi (D)
[image:7.903.128.747.53.482.2]. ) I' A a bl b b2
....
OS!C _all al
セ@
a2 hIUD
....
al a2
....
0Jd6cicaI 0A9 OIefIlcientY
0.'78I
l
o.iS4 I I l.qlmg KaItcog-...
GIIIIIIacocmisTY JCuiD.KmI---
w_
GJICI<lftl<IlIKR <lbambmIaaaK GcddioaKRB Gbc:oIhImiKRB Grl!JjdIKRB 0.89 ' -a_a KaItcog-
---
w_ ...
..
GmaIocccosB_b
-
セ@ a...,; a _ Q,;pIa 0.78 -0: B I-1
I
r--L-I
- I
-
L-.jb
Nセ@ 034
....
0>0IIici00t D
Y
al セ@ a2 b - - - .-022 03$
....
ClodI'am
I
I
... l セ@ ICoItq-
a _-
..
--
GJVoh a ...-
セ@ セ⦅「@w_
a _ 0 OS! r l 1.ompaaa Gpomcta Weos-
---
セ@ GmoIooxmsis_b-
ClhombtooUoaoa_
Qriaidt
a_
[image:8.899.120.794.48.482.2]0.76
Gambar 2. Dendogram 13 aksesi dengan penanda isoenzim (A), RAPD (B), AFLP (C), dan kombinasi isoenzim dan AFLP (0)
,.
セ@
:§
セ@
txI"tI
セ@ d
Lセ@
S:
I-ol - .
セセ@
セイ@
1
ij'
Xセ@
" c). §.[
セ@
f
セ@ セ@j
51. C)""
;:to;;:
セ@
{
セ@
Sl
JI i' II
A
BJ
Cセ@
_ '""'iii&:...
"'P-:- - ; - セ@
セ@
;:;...
• セ@"---3
セ@•
•
-
」セイZM s.o-05
セ@
-
•
G , セイ@ .. " ' • .cAIiGlセ@
:1"''# ·
セ@セ@
1 :
::Iセ@
-...
1
25Ku""" ...
..
セBZG@...
-... u
•
•
•
•
• NLNNセ@-1.5-セ@
-r
11-:--10
• .-* ....
J
0...
セ@..
J
u-_.
...=-=-)
MRNセ@
NNNN[Zセ@ Iae..
•
c_
•
•
.
...
-1
.
...
セ@ -25
-25 ..- -2
-セ@ • セ@
I
-30ft. -3 -5.0
-3.0 -25 -2.0 -LS
-LO
-os u -1 0 1 2 3 4 5 6 -2 0 2 4 6 8iィエセ@ Mセ@ iィエセ@
D セNNNM セM
•
•
E :;:. セ@ FJ
セ@• •
セセ@ lou
0-, . - - .
•
-...-h
•--
セ@ ."":'" -........
""-• "-:- • •
)25
I:
...
-
. .-1-:
• • •
"'tt セ@
セiッ@ w_
...
-• •
)
...-•
-
•
•
) -LS ) -2 セ@NNBNNBNBNセ@ _ M -3
-25
T セ@
•
•-•
Mセ@ •'""":"' -4 セ@
..-..
•
•.eft. -25
-5
-4 -2 0 2 4 6 8 -25 -2.0 -1.5 -1.0 -o.s 0.0 o.s 1.0 -2 -1 0 1 2 3 4 5 6
iィエセ@
Mセ@ Mセ@
G
1.-.,-
H.:;:-• •
I
-s.o
25
1
25-
セMI
IW· ...
セ@ セ@• • • 0.0
BBGZBBGセ@ ..-•
-:..
)
-
セ@I:
セ@ • • • -25-75
""-...
•セB@
..
-So -10.0
-2 0 2 4 6 8 -8 -6 -4 -2 0 2 4
Iht a.o.i>a-t
Mセ@
- -
[image:9.902.127.791.40.489.2]ISBN : 978-979-15549-2-2
PROSIDING SEMINAR NASIONAL HASIL PENELITIAN YANG DIBIAYAI
OLEH HIBAH KOMPETITIF
m m u r a z m
PENINGKATAN PEROLt-HAN HKX DARX HASIL
PENELITIAN YANG DIBIAYAI OLEH
HIBAH KOMPETITIF
BOGOR, 1-2 AGUSTUS 2007
Dalam rangka
Purnabakti Prof. Jajah Koswara
KEPJASAMA
FAKULTAS PERTANIAN IPB
DITJEN PENDIDIKAN TINGGI DEPDIKNAS
PUSAT PERLINDUNGAN VARIETAS TANAMAN DEPTAN
DEPARTEMEN AGRONOMI DAN HORTIKULTURA FAKULTAS PERTANIAN
Seminar ini diselenggarakan oleh Fakultas Pertanian lPB bekerja sama dengan
Direktorat Jenderal Pendidikan Tinggi Depdiknas
dan
Pusat Perlindungan Varietas
Tanaman (PPVT) Deptan dalam rangka Pumabakti Prof., Dr. Jajah Koswara.
Copyright
©
2007 Departemen Agronomi dan Hortikultura Faperta lPB
JI. Merantl Kampus lPB Darmaga Bogor 16680
Telp.lFaks. (0251) 659353 e-mail: agronipb@indo.net.id
lsi dikutip dengan menyebutkan sumberoya
Departemen Agronomi
dan
Hortikultura Fakultas Pertanian Institut Pertanian Bogor.
2007. Peningkatan Perolehan HKI dari Hasil Penelitian yang Dibiayai oleh Hibah
Kompetitif. Prosiding Seminar Nasional Hasil Penelitian yang Dibiayai oleh Hibah
Kompetitif. Bogor, 1-2 Agustus 2007.
xxxv +458
."
セャN@
t; ェNセNセ@ セ@
セエNLN@
)I,
KATAPENGANTAR
Puji syukur kepada Allah SWT atas terselenggaranya Seminar Nasional Hasil Penelitian yang Dibiayai oleh Hibah Kompetitif dalam rangka Purnabakti Prof Dr. Jajah Joswara pada tanggal 1-2 Agustus 2007, hingga diterbitkannya prosiding seminar tersebut. Seminar ini bertema "Peningkatan Perolehan HKI dari Basil Penelitian yang Dihiayai oleh Bibah Kopetitif' .
Seminar diselenggarakan atas ketjasama Fakultas Pertanian IPB, Direktorat Jenderal Pendidikan Tinggi Depdiknas serta Pusat Perlindungan Varietas Tanaman (PPVT) Deptan, dan sebagai panitia pelaksana adalah Departemen Agronomi dan Hortikultura Fakultas Pertanian IPB. Seminar dihadiri 160 peserta yang terdiri atas dosen, mahasiswa dan peneliti. Mengawali presentasi makalah, dilaksanakan sidang pleno dengan topik Sosialisasi HKI-PVT oleh pejabat terkait dari IPB dan Deptan serta praktisi dari kalangan swasta. Terkait dengan Purnabakti Prof. Dr. Jajah Koswara, sebuah buku semi autobiografi karya Prof. Jajah betjudul "Pelajaran hidup yang Tak Pernah Usai : Jalan Masih Panjanft' telah diterbitkan secara terpisah.
Dalam seminar dipresentasikan hasil penelitian yang baru dilaksanakan maupun review hasil-hasil penelitian multi tahun dari sumber dana tunggal maupun beberapa sumber yang berbeda. Review tersebut sangat baik menggambarkan kemanfaatan hibah kompetitif multi tahun yang dirintis oleh Prof. Dr. Jajah Koswara, serta menggambarkan kemajuan pelaksanaan penelitian bersangkutan. Dengan demikian dapat dideteksi potensi HKI-PVT dari hasil-hasil penelitian tersebut.
Makalah presentasi dalam pro siding ini betjumlah 64 terbagi ke dalam 40 makalah presentasi oral dan 24 makalah presentasi poster. Bidang bahasan difokuskan pada tanaman mencakup aspek Agronomi, Pemuliaan Tanaman, Benih, dan Bioteknologi, serta penunjang budidayanya, termasuk penggunaan mikroba. Beberapa makalah yang dipresentasikan dalam seminar tidak diterbitkan dalam pro siding ini atas pertimbangan penulisnya.
Terimakasih disampaikan kepada semua fihak yang telah berpartisipasi mensukseskan Seminar Nasional Hasil Penelitian yang Dibiayai oleh Hibah Kompetitif ini yang terangkai dalam kegiatan Purnabakti Prof. Dr. Jajah Koswara. Disadari masih terdapat kekurangan dalam penyusunan prosiding ini. Meskipun demikian semoga prosiding ini bermanfaat bagi semua pihak yang memerlukannya.
Bogor, Desember 2007 Ketua Departemen Agronomi dan Hortikultura Fakultas Pertanian IPB
Prof. Dr. Ir. Bambang S. Purwoko, MSc.
Prosiding Seminar Nasional Hasil Penelitian yang Dibiayai oleh Hibah KompetitiJ Bogor, 1-2 Agustus 2007
,
DAFTARISI
Halaman KATAPENGANTAR ... 1 DAFTARISI ... ..
SAMBUTAN DEKAN FAKULTAS PERTANIAN IPB ... ..
MAKALAH SIDANG PLENO Perlindungan Varietas Kelapa Sawit
11
VII .
Dwi Asmono ... xi Menjadi Market Trendsetter atau Follower (pengalaman dalam
Perbenihan Sayuran)
Abdul HaIIlid ... XXIX
MAKALAH ORAL
Peran Bahan Organik dalam Meningkatkan Produksi Pertanian M. H. Bintoro, Douglas Manurung, Ishak TanH. Djawahir, dan Wahju
Sujatrniko ... 1 Penambahan COl Internal Tanaman Kapas dengan Pemberian Metanol Guna
Meningkatkan Produksi Melalui Deteksi 14C
Badron Zakaria, Dannawan, dan Nurlina Kasim ... . Mekanisme Fisiologi Tanaman Kedelai pada Kondisi Jenuh Air dan Kering serta Kaitannya dengan Biosintesis Etilen
Munif GhulaIIlahdi ... .. Evaluasi Kualitas Buah Pisang Ambon pada Tingkat Kematangan yang
Berbeda Selama Penyimpanan
10
19
SlaIIlet Susanto, Dina Sabrina, Deliana, Dewi SuIana, dan Sutrisno .... ... 28 Kajian Pertumbuhan, Ekspresi Seks Tanaman, dan Kualitas Buah Pepaya
Genotipe IPB 1 dan IPB 2 dengan Pupuk Organik
Ketty Suketi, Sriani Sujiprihati, Mellyawati, dan Devis Suni ... 36 Pengaruh Ukuran Kawat dan Ukuran Cabang untuk Strangulasi terhadap
Pembungaan Jeruk Besar (Citrus grandis (L.) Osbeck)
Arifah Rahayu, Setyono, dan SlaIIlet Susanto... ... ... ... ... 44 Pengaruh Pemberian Nitrogen terhadap Pertumbuhan dan Produksi Rebung
Bambu Betung (Dendrocalamus asper (Schults F.) Backer Ex Heyne)
Sandra Arifin Aziz ... 51 Adaptasi Pertumbuhan dan Kandungan Flavonoid Daun Dewa (Gynura
pseudochina (L.) Dc) Asal Kultur In Vitro pada Intensitas Cahaya Rendah
Nirwan, Munif GhulaIIlahdi, dan Sandra A. Aziz ... ... ... 60 Struktur Populasi Eriborus argenteopilosus Cameron (Hymenoptera :
Ichneumonidae) pada Beberapa Tipe Lansekap di Sumatera Barat
Novri Nelly dan Y aherwandi ... 69 Sebaran Populasi Nematoda Entomopatogen Steinernema spp. pada Beberapa
Kawasan Pertanian Lahan Gambut di Kalimantan Selatan
Anang Kadarsah dan Jumar ... .. 76 Studi Patogen Penyebab Antraknosa pada Pepaya
Siti Hafsoh ... . 83
ii Daftar lsi
.iI.... セN@
J
f
-"
.!!I---.. ,
/
.
-
..
Perkembangan Penelitian Teknologi Benih Aren (Arenga pinnata (Wurmb.) Merr.) di Universitas Tadulako
Muhammad Salim Saleh, Enny Adelina, Maemunah, Nuraeni, Idham, Sakka
Samudin, dan Nur Alam ... 91 Wani Bali (Mangifera caesia Jack.) Tanpa Biji, Prospek Pengembangan dan
Kendala Pembibitannya
I. N. Rai, O. Wijana, dan C. O. A. Semarajaya ... .. 97 Sistem Pembibitan Manggis untuk Distribusi
M.Rahmad Suhartanto, A. Qadir Dan Muzayyinatin ... ... 105 Pertumbuhan dan Hasil Tanaman Kacang Bogor (Vigna subterranea (L.)
Verdcourt) Galur Gresik dan Bogor pada Berbagai Wama Biji
Endah Sri Redjeki ... 114 Perbanyakan Klonal Phalaenopsis sp. In Vitro dari Eksplan Daun dan
Eksplan Tangkai Bunga
Yusnita, Candra Kesuma, Devina Andiviaty, Sri Ramadiana dan Dwi Hapsoro ... 119 Respon Tanaman Anggrek Bulan terhadap Jenis Media Tanam dan
Letak Tanaman Pada Sistem Pertanian Organik secara Vertikultur
Yati Suryati ... 125 Analisis Daya Gabung dan Aksi Gen Ketahanan Cabai (Capsicum annuum L.)
terhadap Antraknosa yang Disebabkan oleh Colletotrichum acutatum
Muhamad Syukur, Sriani Sujiprihati, Jajah Koswara dan Widodo ... 131 Interaksi Genotipe X Musim pada Karakter Hasil dan Komponen Hasil Ubi
27 Genotipe Bengkuang (Pachyrhizus erosus L. Urban) pada Lingkungan Pemangkasan Reproduktif Di Jatinangor
Agung Karuniawan ... . Galur Kacang Tanah Berdaun Hijau Tua : Keunggulan dan
Pengendalian Genetiknya
Yudiwanti ... . Prospek Senyawa Anti Giberelin dalam Memacu Peningkatan Vigoritas Planlet
137
143 Suseno Amien ... 147 Analisis Daya Gabung dan Heterosis Hasil Galur Jagung Dr Unpad melalui
Analisis Dialel
D. Ruswandi, M. Saraswati, T. Herawati, A. Wahyudin, dan N. Istifadah ... 153 Keragaman Fenotipik dan Genetik Mahoni (Swietenia macrophylla) di Jawa
Tengah dan Jawa Timur
Ulfah J. Siregar, Iskandar Z. Siregar, dan Insan Novita ... 160 Pengujian Cabai Hibrida IPB di Dua Lokasi
Muhamad Syukur, Sriani Sujiprihati, dan Rahmi Yunianti ... ... 165 Pendugaan Daya Gabung dan Heterosis Ketahanan terhadap Phytophthora
capsici Leonian pada Persilangan Dialel Penuh Enam Genotipe Cabai (Capsicum annuum L.)
Rahmi Yunianti, Sarsidi Sastrosumarjo, Sriani Sujiprihati,
Memen Surahman, dan Sri Hendrastuti Hidayat ... ... ... ... 172 Tinjauan Ulang Pengembangan Teknologi Haploid Cabai dan Prospeknya
untuk Percepatan Penelitian Genetika dan Pemuliaan Tanaman
Ence Darmo Jaya Supena ... . Uji Daya Adaptasi dan Interaksi Genotipe X Lingkungan Galur Potensial
Keturunan Persilangan Mentik Wangi dengan Poso untuk Perakitan Padi Gogo Aromatik
Totok Agung D.H. Dan Suwarto ... ..
Prosiding Seminar Nasional Hasil Penelitian yang Dibiayai oleh Hibah KompetitiJ Bogor, 1-2 Agustus 2007
179
187
Pemuliaan Padi Gogo Tenggang Aluminium dan Tahan Bias ' melalui Kultur Antera
Bakhtiar, Bamhang S. Purwoko, Trikoesoemaningtyas, M.A. cィッコセ@ Iswari S.
Dewi, dan Mukelar Am.ir ... 197 Seleksi Nenas Hasil Persilangan Cayenne dengan Queen di Jatinangor
Neni Rostini, Citra Bakti, dan Syaiful Muharok ... . Pendugaan Parameter Genetik dan Hubungan antar Hasil dengan Beberapa Karakter Kuantitatif dari Plasma Nutfah Nenas (Ananas comosus L. Merr.) Koleksi PKBT IPB
Muhammad ArifNasution, Roedhy Poerwanto, Sohir, Memen Surahman, dan Trikoesoemaningtyas ... ; ... . Perakitan Padi Gogo Toleran Tanah Masam Dan Berdaya Hasil Tinggi :
Seleksi Dengan Metode Bulk
Surjono H. Sutjahjo, Trikoesoemaningtyas, Desta Wirnas, Rustikawati,
Rosy I. Saputra ... . Uji Daya Hasil Lanjutan Galur Harapan Padi Sawah Tipe Baru
di Tiga Lokasi
Hajrial Aswidinnoor, Willy Bayuardi Suwarno, Intan Gilang Cempaka, Ratna
205
211
218
Indriani, dan Wulandari Siti Nurhidayah ... ... ... .... 222 Perbaikan Sifat Agronomi dan Kualitas Sorgum Sebagai Sumber Pangan,
Pakan Ternak, dan Bahan Industri melalui Pemuliaan Tanaman dengan Teknik Mutasi
Soeranto Human ... ... .... ... .... ... ... ... .... ... ... ... ... 226 Konstruksi Mutan Pseudomonas sp. Crb17 untuk Meningkatkan Produksi
Indole Acetic Acid Melalui Mutagenesis dengan Transposon
Mutiha Panjaitan, Aris Tri Wahyudi, dan Nisa Rachmania ... . Variabilitas Genetik Mutan-Mutan Manggis In Vitro berdasarkan
Marka RAPD
Warid Ali Qosim, R. Poerwanto, G. A. Wattimena, Witjaksono, Sohir, dan
N. Carsono ... . Aplikasi Marka Isoenzim, RAPD, dan AFLP untuk Identif1kasi Variabilitas Genetik Tanaman Manggis (Garcinia mangostana) dan Kerabat Dekatnya
Soaloon Sinaga, Sohir, Roedhy Poerwanto, Hajrial Aswidinnoor, Dedy Duryadi,
234
240
Resmitasari, Rudy Lukman, dan Roswita Amelia. ... ... ... ... ... ... 247 Amplif1kasi CDNA Kedelai dettgan Beberapa Primer SpesnIk Gen Cao
(Chlorophyll A Oxygenase)
Nurul Khumaida, Kisman, dan Didy Sopandie ... 256 Analisis Sekuen Lengkap Gen yang Terkait Adaptasi Kedelai terhadap
Intensitas Cahaya Rendah
Kisman, Nurul Khumaida, dan Sohir ... 261 Seleksi In Vitro K1on-Klon Kentang Hasil Persilangan cv. Atlantik dan
Granola untuk Mendapatkan Calon Kultivar Kentang Unggul
Awang Maharijaya, Muhammad Mahmud, dan Agus Purwito ... 268 Karakterisasi Abnormalitas Embrio Somatik Kelapa Sawit (Elaeis guineensis
Jacq) Berdasarkan Morfologi, RAPD dan Metilasi RP-HPLC
Nesti F. Sianipar, Gustav A.Wattimena, Maggy Thenawidjaya S., Hajrial
Aswidinnoor, dan Nurita Toruan-Mathius ... .. MAKALAH POSTER
Pengaruh Pendinginan Larutan Hara terhadap Pertumbuhan dan Hasil Tanaman Bawang Merah pada Sistem Hidroponik dengan Empat Macam Media Tanam
Agus Margiwiyatno ... ..
iv
276
285
Daftar lsi
r>\{J_
セ@
\!
r;
-
.
i
,
I
J'
,.
"
Aセ@
Analisis Keanekaragaman Genetik 27 Genotipe Cabai (Capsicum spp.) KoleksiIPB
Ahmad Meka Rosyadi, Sriani SUjiprihati, dan Rahmi Yunianti ... 291
Uji Ketahanan Terhadap Bias Daun Galur-Galur F4:6 Padi Gogo Hasil Seleksi Tanah Masam
Desta Wimas, Trikoesoemaningtyas, Surjono H. Sutjahjo, Khoirul Hidayah, dan Lestari Atmojo ... .
Perlakuan Ec dan Ph Larutan Media Hidroponik pada Bawang Merah Varietas Sumenep, Philipin dan Tiron
Eni Sumarni dan Noor Farid ... .
Akumulasi dan Sekresi Asam Organik pada Padi Gogo Toleran dan Peka Aluminium serta Perannya dalam Mobilisasi P
Etti Swasti dan Nalwida Rozen ... .
Pendugaan Nilai Heritabilitas dan Korelasi Genetik Beberapa Karakter Agronomi Tanaman Semangka (Citrullus lanatus (Thunb.) Matsum & Nakai)
299
305
312
Memen Surahman, Muhamad Syukur, dan Anita Amalia Rahmawati ... 320
Evaluasi Ketahanan Beberapa Persilangan Semangka (Citrullus lanatus (Thunb.) Matsum & Nakai) terhadap Layu Fusarium (Fusarium oxysporum
f. sp. Niveum)
Muhamad Syukur, Efi Toding Tondok, dan Swisci Margaret ... 326
Pengembangan Budidaya Jenuh Air Tanaman Kedelai dengan Sistem Tumpangsari Padi Kedelai di Lahan Sawah
Munif Ghulamahdi, Sandra Arifin Aziz, Maya Melati, Nurwita Dewi,
dan Sri Astuti Rais ... 331
Ketahanan 23 Genotipe Cabai (Capsicum sp.) terhadap Penyakit Antraknosa (Colletotrichum sp.)
Sriani Sujiprihati, Muhamad Syukur, Widodo, Efi Toding Tondok,
Rahmi Yunianti dan Neni Hariati ... .
Tanggap Morfologi dan Fisiologi Padi Gogo Fase Semai pada Kekeringan untuk Memudahkan Seleksi
Noor Farid dan Datjanto ... .
Aplikasi Filter Cahaya dan Teknik Cutting dalam Perbanyakan Vegetatif Tanaman Sansevieria trijasciata 'Laurentii'
Peni Lestari, Nurul Khumaida, dan Ani Kurniawati ... .
Perbanyakan Bambu Betung (Dendrocalamus asper (Schults F.) Backer Ex Heyne) pada Kultur In Vitro
Sandra Arifin Aziz, Fred Rumawas, Livy W. Gunawan, Bambang S. Purwoko,
Hajrial Aswidinnoor, Achmad Surkati Abidin, dan Maggy T. Suhartono ... .
Pengaruh Pepton terhadap Pengecambahan Biji Anggrek Phalaenopsis Amabilis dan Dendrobium Hybrids In Vitro
Sri Ramadiana, Rizka Dwi Hidayati, Dwi Hapsoro dan Yusnita ... .
Determinasi Tipe Seks Pepaya (Carica papaya L.)
Kartika Trias Maknani, Muhamad Syukur, dan Sriani Sujiprihati ... , ... .
Studi Kromosom Anyelir (Dianthus caryophyllus Linn.) Mutan Akibat lradiasi Sinar Gamma
Tia Atisa, Syarifah lis Aisyah, dan M. Syukur ... .
Induksi dan Proliferasi Kalus Embrionik pada Beberapa Genotip Kedelai Peka dan Toleran Naungan
Tri Handayani dan Nurul Khumaida ... .
Prosiding Seminar Nasional Hasil Penelitian yang Dibiayai oleh Hibah Kompetitij Bogar, 1-2 Agustus 2007
337
342
348
357
366
373
379
387
L/'
Keragaman Kandungan Trypsin Inhibitor pada Beberapa Provenan Sengon (Paraserianthes falcataria) di Jawa Sebagai Mekanisme Alami Ketahanan terhadap Hama
Ulfah J. Siregar ... . Hubungan Kekerabatan antar Genotipe dalam Tiga Grup Kultivar Melon
Willy Bayuardi Suwamo dan Sobir ... .. Interaksi Genotipe-Lingkungan untuk Ketahanan terhadap Penyakit
Bercak Daun pada Galur-Galur Kacang Tanah
Chaireni Martasari, S. Sastrosumarjo, A.A. Mattjik, dan Yudiwanti ... .. Pemanfaatan Parasitoid Tetrastichus schoenobii Ferr. (Eulopidae,
Hymenoptera) dalam Pengendalian Penggerek Batang pada Tanaman Padi Arifm Kartohardjono ... . Komparasi Respon Fisiologis Tanaman Kedelai yang Mendapat Cekaman
Kekeringan dan Perlakuan Herbisida Paraquat
Violita, Hamim, Miftahudin, Triadiati dan Soekisman Tjitrosemito ... . Peroksidasi Lipid pada Akar Padi (Oryza sativa L.) sebagai Respon Fisiologis terbadap Cekaman Aluminium
Sri Aninda Wulansarl, Utut Widyastuti Suharsono, Hamim, dan Miftahudin ... Keragaman Aktivitas Nitrat Reduktase (Anr) dan Kandungan KIorof"JI
Beberapa Aksesi Pisang (Musa spp.) di Wilayab Banyumas
Dyah Susanti, B. Prakoso, S. Nurchasanah, dan L.S. Abidin ... .. Pengarub Kualitas Cabaya dan Fotoperiode terbadap Pertumbuban
dan Perkembangan Kastuba In Vitro
Muhannnad Ibrahim Faruq dan Dewi SuIana ... ..
SUSUNAN P ANITIA ... .. SUSUNAN ACARA ... . DAFT AR PESERTA SEMINAR ... . INDEKS PEMAKALAH ... . INDEKS KOMODITAS ... .
vi
397
402
409
413
419
426
432
437
441
443
453
456
458
Daftar lsi