BAB I PENDAHULUAN
Latar Belakang
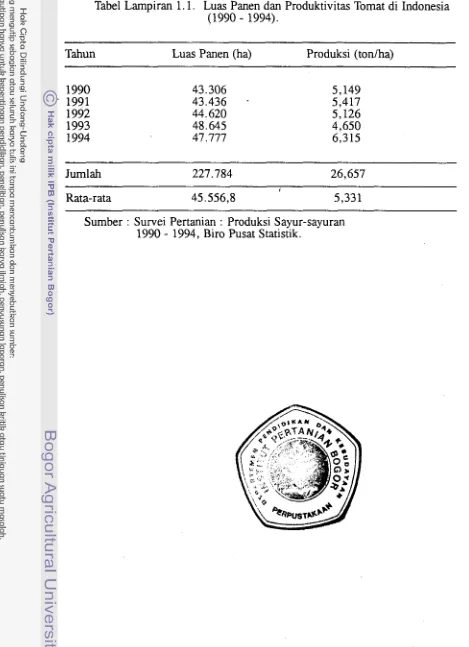
Tomat, Lycopersicon esculentum Mill., merupakan salah satu tanaman sayuran penting di Indonesia. Pada saat ini rataan produksi tomat per hektar
adalah 5,33 tonlha (Tabel Lampiran 1. I), masih jauh lebih rendah dibanding
dengan potensi produksinya yang dapat mencapai 25 tonlha (Sahat 1992).
Rendahnya produksi tersebut antara lain karena serangan hama Helicoverpa armigera (Hiibner) (Lepidoptera : ~octuidbe) (Setiawati et al. 1993).
Helicoverpa spp. (Lepidoptera : Noctuidae) merupakan harna utama tanaman
tomat di berbagai negara (Goncalves et al. 1989; Kennedy et al. 1983; Lange &
Bronson 1981; Martin et al. 1990; Ogunwolu 1989; Walgenbach et al. 1989; Walgenbach et al. 1991 ; Walgenbach & Ester 1992). Larva menimbulkan kerusakan dengan cara menggerek ke dalam buah. Serangan berat dilaporkan
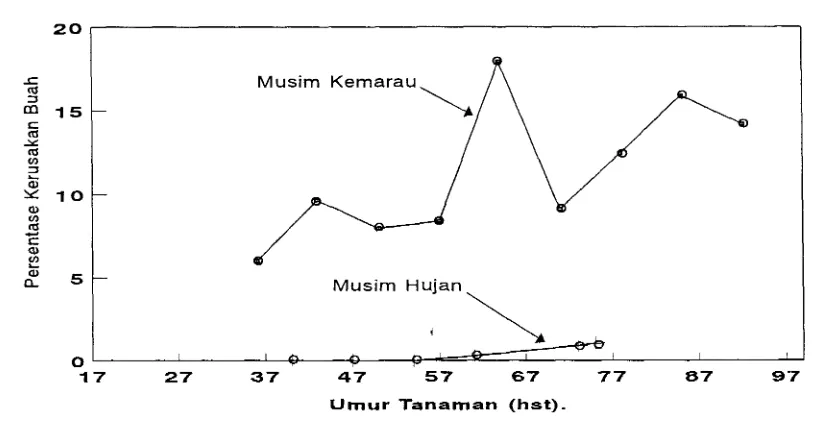
terjadi pada musim kemarau dengan kerusakan buah mencapai 80 % (Uhan &
Suriaatrnadja 1993).
Pada saat ini, pengendalian yang biasa dilakukan petani adalah aplikasi insek-
tisida secara berjadwal, dengan frekuensi dan dosis melebihi yang direkomendasi-
kan (Uhan & Suriaatmadja 1993). Praktek pengendalian seperti itu selain mahal
juga dapat menimbulkan hal-ha1 yang tidak diinginkan seperti resistensi dan
riserjensi hama, pencemaran lingkungan, serta residu insektisida pada buah
tomat.
Ketergantungan petani pada insektisida dalam pengendalian ulat buah tomat
antara lain karena teknik pengendalian alternatif yang tepat dan rasional belum
2 efektif dan efisien tanpa menimbulkan dampak negatif. Teknik pengendalian
tersebut dapat diwujudkan melalui penerapan konsepsi pengendalian hama terpadu
(PHT). Karena PHT pada dasarnya adalah ekologi terapan, maka pengembangan
PHT memerlukan pemahaman ekologi dari hama sasaran. Dengan pemahaman
itu, titik-titik lemah kehidupan hama dan komponen-komponen lingkungan yang
berpengaruh terhadap perkembangan hama dapat diketahui dan dapat dimanipulasi
untuk pengendalian populasinya.
Informasi ekologi H. armigera pada ekosistem tanaman tomat di Indonesia belum banyak diketahui. Kepustakaan yhng berkaitan dengan ulat buah tomat
pada umumnya terbatas pada pengujian aplikasi insektisida (Setiawati et al. 1993; Uhan & Suriaatmadja 1993). Dengan latar belakang hal-ha1 yang diuraikan di
atas maka penelitian ini dilakukan untuk memahami ekologi H. armigera pada pertanaman tomat.
Pemahaman ekologi H. armigera mancakup pengkajian terhadap perkem- bangan populasinya dan faktor-faktor lingkungan yang mempengaruhinya.
Faktor-faktor lingkungan yang dapat mempengaruhi perkembangan populasi hama
meliputi tanaman sebagai sumber daya, faktor abiotik seperti curah hujan, suhu,
cahaya, dan kelembaban, dan faktor biotik yang meliputi predator, parasitoid, dan
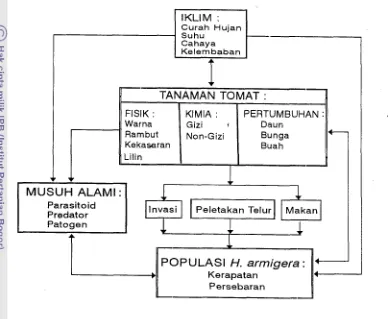
patogen (Clark et al. 1967). Hubungan antara faktor-faktor tersebut dicantum- kan pada Gambar 1.1.
Untuk menerapkan konsepsi PHT secara mantap diperluhan pengetahuan yang
menyeluruh dari semua faktor tersebut, baik masing-masing faktor maupun inter-
aksinya. Untuk mewujudkan ha1 itu perlu dilakukan penelitian secara bertahap
dalarn jangka waktu yang relatif lama. Mengingat keterbatasan waktu yang terse-
3
tor yang memegang peranan kunci dalam mempengaruhi perubahan kelimpahan
populasi.
[image:143.582.110.501.158.477.2]Persebaran
Gambar 1.1. Hubungan Faktor Lingkungan yang Mempengaruhi Perkem- bangan Populasi H. amzigera.
Tanaman tomat merupakan sumber daya makanan dan ruang bergerak dan
berlindung bagi
H.
armigera. Oleh karena itu, kelimpahaq populasi hama tergan- tung dari kesesuaian antara fenologi hama dengan tanaman inang skbagai sumberdaya. Ngengat H. armigera mengin~asi~dan meletakkan telur karena tertarik pada aroma kimia (chemical odour) atau warna dari tanaman tomat. Bunga
diduga merupakan bagian tanaman yang mengeluarkan aroma kimia danlatau
4
Karena kelimpahan bunga selama pertumbuhan tanaman bervariasi, maka ha1 itu
akan berpengaruh pada peletakan telur. Selain itu keberhasilan bertahan hidup
larva yang muncul dari telur tergantung pada ketersediaan buah sebagai makanan-
nya. Informasi tentang hubungan antara peletakan telur dengan pembungaan
tanaman tomat belum tersedia. Di samping itu perkembangan populasi H.
armigera dan kaitannya dengan kerusakan buah masih perlu diteliti.
Pemantauan populasi inerupakan komponen penting dari PHT. Oleh karena
itu hal-ha1 yang berhubungan dengan pemantauan seperti pola persebaran perlu
juga dipahami. Pemahaman pola persebaran diperlukan untuk menentukan
metode penarikan contoh yang tepat, baik untuk pengambilan keputusan pengen-
dalian maupun untuk pendugaan populasi.
Musuh alami merupakan faktor biotik yang sangat besar peranannya dalam
menentukan kelimpahan dan keseimbangan populasi. Pada ekosistem tanaman
tomat di Indonesia, jenis-jenis musuh alami H. armigera belum banyak diketahui. Dengan demikian, penelitian ini juga mencakup pemahaman musuh alami H.
armigera, dengan penekanan pada parasitoid dan predator.
Tujuan Penelitian
Penelitian bertujuan untuk memahami ekologi H. amigera, yang meliputi:
(1) hubungan antara peletakan telur dengan pembungaan tanaman tomat; (2) perkembangan populasi telur dan larva H. armigera pada pertanaman tomat; (3) pola persebaran spasial telur dan larva dan pengembangan metode penarikan
Kegunaan Penelitian
Sesuai dengan tujuan penelitian, diharapkan informasi yang diperoleh dalam
penelitian ini dapat dimanfaatkan untuk menentukan saat dan metode penarikan
contoh yang tepat (tujuan 1 dan 3). Hasil penelitian juga bermanfaat dalam
mewaspadai atau memperkirakan kehilangan hasil yang mungkin terjadi (tujuan
2), serta memanfaatkan musuh alami sebagai upaya pengendalian yang rasional
dan efektif dalarn rangka mengurangi ketergantungan terhadap aplikasi insektisida
(tujuan 4 dan 5). I
Daftar Pustaka
Clark, L.R., P. W. Geiler, R.D Hughes, & R.F. Morris. 1967. The Ecology of Insect Population in Theory and Practice. Methuen & Co. Ltd., London. 232 hal.
Goncalves, M., F. Martin, A.M.P. Lavadinho & J.B. Sobreiro. 1989. Infesta- tion levels of Heliothis armigera in tomato in Portugal. Bull. SROP. 13(3)
: 100-103.
Kennedy, G.G., L.R. Romanow, S.F. Jenkins & D.C. Sanders. 1983. Insect and diseases damaging tomato fruits in the Coastal Plain of North Carolina. J.
Econ. Entomol. 76: 168-173.
Lange, W. H. & L. Bronson. 198 1. Insect pests of tomatoes. Annu. Rev. Ento- mol. 26: 345-371.
Martins, F., A.M.P. Goncalves, & J.B. Sobreiro. 1990. Extent of the attack of
Heliothis armigera in tomatoes for industrial processing. Act de Horticul- tura: 154-159.
Sahat, S. 1992. Results of highland vegetable research. Dalam Permadi, A.H. ;
Sahat, S.; Sastrosiswojo, S . ; & Bahar, F.A. (ed). Proceedings of the National Vegetable Workshop : Evaluation and Planning of Vegetable Research and Development in the Indonesian Vegetable Production and Industry. Hal. 43-59.
Setiawati, W., Rustaman, E . , Soeriaatmadja & Duskarno. 1993. Effektivitas feromonoid seks dan ambang kendali terhadap serangan Helicoverpa armigera (Hbn) pada tanaman tomat. Bul. Penel. Hort. 25 (3) : 121-127.
Uhan, T.S. & R. E. Suriaatmadja. 1993. Pengendalian ulat buah tomat (Heliothis armigera Hiibn) dengan insektisida organophosphat dan pirethroid buatan. Bul. Penel. Hort. 25 (4): 29-34.
Walgenbach, J.E. & E.A. Ester. 1992. Economics of insecticide use on staked tomatoes in Western North Carolina, J. Econ. Entomol. 85(3) : 888-894. Walgenbach, J.F., R.B. Leidy & T.J. Sheets. 1991. Persistence of insecticides
on tomato foliage and implications for control of tomato fruitworm (Lepidoptera : Noctuidae). J. Econ. Entomol. 84(3): 978-986.
Tabel Lampiran 1.1. Luas Panen dan Produktivitas Tomat di Indonesia (1 990
-
1994).--
Tahun Luas Panen (ha) Produksi (toniha)
Jumlah 227.784 26,657
[image:147.582.60.529.93.740.2]BAB I1
TINJAUAN PUSTAKA
Genus Heliothis merupakan kompleks serangga kosmopolit yang bersifat polifag (Mitter, Poole & Mathew 1993). Hardwick (1965) mengkaji kompleks
Heliothis di Amerika Utara berdasarkan metode filogenetik. Ia menyimpulkan bahwa di antara kompleks Heliothis itu terdapat kelompok yang memiliki karakter yang unik. Hardwick (1965) mengusulkan genus Helicoverpa untuk kelompok
Heliothis yang disebutkan terakhir ini, sedangkan sisanya tetap digolongkan dalam genus Heliothis, kecuali Heliothis virescens. Analisis filogenetik dan morfologi lebih lanjut membuktikan bahwa genus Helicoverpa terdiri dari beberapa kelom- pok (Mitter et al. 1993) seperti tertera pada Gambar 2.1. Kelompok gelotopoeon
merupakan kelompok primitif, sedangkan kelompok armigera yang meliputi
Helicoverpa armigera dan Helicoverpa helenae adalah kelompok mutakhir (Garnbar 2.1).
Morfologi Larva dan Imago
Hingga kini, larva spesies Helicoverpa belum dapat dibedakan dengan jelas. Meskipun demikian, untuk kepentingan praktis, beberapa penulis mempelajari
morfologi larva untuk membedakan spesies. Misalnya Kirkpatrick (1961)
mencoba menyusun kunci untuk memisahkan empat spesies Helicoverpa yang ada di Australia yaitJ H. armigera, H. punctigera, H. rubrescens dan H. assulta
9
bentuk sklerit pada bagian dorsal dari ruas abdomen I dan berdasarkan warna tungkai.
[AUS TRALO THIS ] HELICOVERPA :
Kelompok Gelotopoeon
4
gelotopoeon titicacaeI bracteae
I
I atacamae
-
Kelompok Puntigera :I
,
7 puntigeraL I Kelompok Hawaiiensis I
I
I - -
C
pallid'!I I ha wallensis I I
4 I Kelompok Zea
minuta assulta - fletcheri
pacifica prepodes tibetensis toddi con fusa
LI
Kelompok Armigera :;-
armigeraL helenae
Persebaran : Indo-Australia
Amerika Selatan Argentina, Paraguay Peru
Chili, Peru
Australia
Hawai Barat Hawai Timur
Hawai Barat
Afrika, Asia, Indo-Australia Afrika
Hawai Timur Australia
Ti bet
Madagascar, Afrika Timur Hawai Timur
Amerika Utara dan Selatan
Eropa, Asia dan Afrika Kepulauan Helena
Garnbar 2.1. Dugaan Hubungan Filogenetik spesies Helicovelpa (Mitter et al. 1993) (-
- - -
- Berarti Hubungan Filogenetik Masih Belum Pasti).Pada H. arrnigera sklerit dorsal ruas I abdomen berbentuk pelana dan tungkainya berwarna gelap, sedangkan pada H. punctigera tidak terdapat pelana pada ruas I
abdomen dan tungkainya berwarna pucat. Daly & Gregg (1985) menyatakan bahwa metode Stanley tersebut belum dapat digunakan secara umum. Hal itu
membedakan spesies
Helicoverpa.
Imago
H. armigera
danH. punctigera
dapat dibedakan dengan jelas berdasar-kan adanya garis melintang pada sayap depan. Pada H.
armigera
garis melintang pada sayap depan sangat jelas, sedangkan padaH. punctigera
tidak begitu jelas.Biologi
Telur
Telur
Helicoverpa
spp. pada umumnya berbentuk bulat dengan diameterI
kurang lebih 0,5 mm. Pada waktu diletakkan telur adalah bening (Setiawati
199 1 ; Zalucki
et al.
1986), kemudian berangsur-angsur berubah menjadi keku- ningan, dan menjelang menetas berbintik hitam. Lama stadia telur tergantungdari suhu. Setiawati (1991) melaporkan bahwa di Lembang pada kondisi labora-
torium dengan suhu 18"
-
28"C, telur H.armigera
menetas dalam waktu 10 sampai 18 hari setelah diletakkan. Pada suhu yang lebih tinggi, stadia telurberlangsung lebih singkat. Elna (1988) melaporkan bahwa di Bogor pada suhu
rata-rata 27"C, telur menetas antara tiga dan empat hari setelah diletakkan.
Perkembangan
Larva
Lama perkembangan larva bervariasi tergantung antara lain dari jenis
tanaman inang. Goyal & Rathore (1988) melaporkan bahwa terdapat variasi lama stadia perkembangan dan indeks pertumbuhan pada berbagai tanaman inang.
Pada tanaman kapas dan tomat perkembangan lebih cepat dan indeks pertumbuhan
lebih tinggi dibanding dengan pada tanarnan lain. Elna (1988) melaporkan bahwa
11
H . armigera yang dipelihara pada kacang buncis segar terdiri dari lima instar.
Singh & Rembold (1988) melaporkan bahwa larva yang dipelihara pada makanan buatan dari bahan kacang buncis dan kedelai terdiri dari lima instar, sedangkan
dari bahan jagung enam instar.
Suhu juga mempengaruhi lama perkembangan. Di Australia pada suhu 16"
-
18"C, H. armigera membutuhkan waktu 73 hari untuk berkembang dari telur menjadi dewasa, sedangkan pada suhu 28°C pada musim panas memerlukan
waktu 34 hari (Zalucki et al. 1986). Di Lembang pada suhu 18"-26"C, lama
f
perkembangan H. armigera mulai dari telur sampai ngengat mati adalah 52 - 82 hari (Setiawati 1991).
Pupa
Pupa yang baru terbentuk berwarna kekuningan, kemudian berubah menjadi
kuning-kecoklatan dan menjelang imago keluar berwarna coklat tua. Lama stadia
pupa bervariasi antara 15 dan 21 hari. Bentuk pupa betina dapat dibedakan dari
jantan. Pada ruas terakhir abdomen pupa betina terdapat celah yang berbentuk
segi tiga, sedangkan pada jantan terdapat celah yang bentuknya membulat (Setia-
wati 1991).
Imago
Ngengat biasanya muncul dan aktif pada malam hari. Ngengat betina H. punctigera berkopulasi dalam waktu satu sampai dua hari setelah muncul dari
kepompong, dan dapat berkopulasi hingga enam kali selama hidupnya, serta
menghasilkan telur rata-rata 1395 dan 1437 masing-masing pada suhu 19" dan
4 16 butir (Setiawati 1991).
Berdasarkan hasil pemantauan dengan lampu perangkap, Persson (1976)
melaporkan bahwa ngengat
H.
punctigera lebih banyak tertangkap sebelum tengah malam, sedangkanH.
armigera setelah tengah malam. Persson (1976) menyata- kan bahwa puncak aktivitas betina berkaitan dengan puncak peletakan telur.Aktivitas peletakan telur Helicoverpa spp. umumnya berhubungan dengan pembungaan dan produksi nektar (Adjei-Maafo & Wilson 1983; Wardhaugh,
Room & Greenup 1980; Wilson, Basinski & Thomson 1972; Wilson 1976).
7
Pada kedelai, kapas dan tembakau, ngengat betina mulai meletakkan telur
menjelang pembungaan (Broadley 1978; Wardhaugh et al. 1980; ).
Zalucki et al. (1986) menyatakan bahwa ngengat betina meletakkan telur satu per satu pada berbagai bagian tanaman seperti daun, kuncup tunas, bunga, buah
dan batang. Saour & Causse (1993) mengadakan penelitian tentang perilaku
peletakan telur oleh H. armigera pada tanaman tomat. Kedua peneliti ini mela- porkan bahwa daun paling disukai untuk peletakan telur terutama yang letaknya
dekat tandan bunga dan buah. Sebagian telur diletakkan pada bunga, kuncup
bunga, batang, dan buah.
Helicoverpa spp. sebagai Hama Tanaman
Status serangga fitofag dapat digolongkan ke dalam salah satu dari empat
kelompok hama, tergantung dari posisi keseimbangan umum (PKU) relatif terha-
dap posisi tingkat kerusakan ekonomi (TKE) (Stern et al. 1959). Keempat kelompok tersebut adalah : 1) serangga yang TKE-nya jauh di atas posisi PKU- nya, sehingga serangga tersebut tidak pernah menimbulkan kerugian secara
13
disebut dengan harna kadang-kadang (occasinal pest); 3) serangga yang TKE-nya
sedikit lebih tingg i dar i PKU, sehingga setiap kali populasinya meningkat menca-
pai TKE, dan serangga demikian tergolong dalam hama tahunan (perennial
pest); dan 4) serangga yang TKE-nya di bawah PKU, sehingga merupakan hama sepanjang musim, dan oleh karena itu disebut sebagai hama utarna (severe pest). - Status Helicoverpa spp. sebagai hama tergantung dari jenis tanaman yang
diserangnya. H. zea yang menyerang alfalfa, populasinya tidak pernah mencapai TKE, sedangkan jika menyerang kapas, H. zea termasuk hama tahunan, serta
t
bila menyerang jagung manis H. zea termasuk hama penting (Luckmann &
Metcalf, 1982).
Pertumbuhan Tanaman Tomat dan Peletakan Telur
Pertumbuhan Tanaman Tomat
Sifat pertumbuhan tanaman tomat dapat digolongkan ke dalam determinate,
indeterminate, dan semi determinate. Pada tomat yang determinate, pertumbuhan
vegetatif berhenti bila telah terjadi pertumbuhan generatif atau telah berbunga,
sedangkan pada indeterminate, pertumbuhan vegetatif berlangsung terus meskipun
tanaman telah berbunga.
Varietas Taiwan tergolong tipe indeterminate, dan merupakan varietas impor
yang banyak ditanam oleh petani di dataran tinggi. Saat ini varietas Taiwan (TW)
yang dijumpai di pasar terdiri dari beberapa nomor seri, yaitu TW 303, TW 305,
TW 375. Varietas yang disebut terakhir lkbih disukai oleh petani tomat di Desa Tugu, karena pertumbuhannya lebat, bunga dan buahnya banyak, dan produksi-
nya tinggi.
14
per tandan cukup beragam (Budi, Anggara & Etti 1986). Mereka melaporkan
bahwa tanaman mulai berbunga pada umur antara 45 hingga 60 hari setelah tanam
(hst), dengan dua hingga enam bunga per tandan. Untuk varietas Taiwan yang
pertumbuhannya bersifat
indeterminate
umumnya mulai berbunga pada umur 37- hst (Etti & Ali 1990), dan khusus varietas Taiwan nomor seri 305 (TW 305)
inulai berbunga pada umur 30 hst (Etti
et al.
1990). Pada pengarnatan pendahu-luan oleh penulis (tidak dipublikasikan) pada musim tanam akhir November 1994,
varietas TW 375 mulai berbunga pada umur antara 23 hingga 25 hst, dengan 4 -
i 12 bunga per tandan.
Perilaku Peletakan Telur
Peletakan telur merupakan salah satu tahapan dari proses kolonisasi. Renwick
& Chew (1994) menyatakan bahwa proses peletakan telur menyangkut perilaku pencarian, orientasi, penemuan, turunlhinggap, evaluasi permukaan, dan peneri-
maan inang
.
Perilaku pencarian, orientasi hingga penemuan tanaman inang ,biasanya dituntun oleh adanya senyawa volatil dari tanaman, seperti dilaporkan
untuk
Trichoplusia ni
danPlutella maculipennis
(Khanet
al. 1987; Gupta &Thorsteinson 1960). Liu
et al.
(1 988) melaporkan bahwa 4-hexen- 1-01 acetat,2-2-dimethyl hexanal, dan 2-hexenal merupakan senyawa yang berkaitan dengan
peletakan telur oleh
T.
ni.
llnggap ngengat terhadap senyawa volatil juga dapat berkaitan dengan upaya
mencari sumber nektar. Phenylacetaldehyde, benzaldehyde dan benzyl acetat
pada bunga
Abelia grandiflora
dan bungsmelati(jasmine)
dilaporkan memikatngengat
T.
ni
(Hayneset al.
1991; Heathet al.
1992). Wiesenborn & Baker (1990) menyatakan bahwaPectinophora gossypiella
tanggap terhadap senyawa15
tarsi, antena, probosis, dan ovipositor sangat berperan dalam menentukan kesesu- aian inang (Renwick & Chew 1994). Tekstur permukaan tanaman berkaitan
dengan pemilihan tempat peletakan telur yang sesuai. Permukaan tanarnan yang
berambut dan kasar sesuai untuk peletakan telur, karena ngengat dapat berpijak
dengan baik dan telur tidak mudah terlepas (Renwick & Chew 1994; Callahan. 1957). Hal yang sama dilaporkan pula untuk H. armigera dan H . puntigera
(Cullen 1969; Hassan 1985).
Pola Persebaran
Persebaran hama dalam habitatnya merupakan fenomena ekologi yang khas
untuk setiap spesies (Taylor 1984), yang dapat mengikuti pola acak, mengelom-
pok, atau beraturan. Pola persebaran merupakan cerminan interaksi antara peri-
laku dan keragaman lingkungan, khususnya tanaman inang sebagai sumber daya
makanan dan ruang (Southwood 1978; Iwao 1979).
Persebaran serangga pada umumnya tidak mengikuti pola acak (Taylor 1984).
Populasi serangga sebagai mahluk hidup mempunyai saling ketergantungan
antara satu dengan yang lain. Serangga dari golongan Lepidoptera dalam siklus
hidupnya mempunyai lebih dari satu fase perkembangan yang berbeda. Telur
dan larva merupakan fase perkembangan yang relatif kurang aktif, dan perse- barannya sangat ditentukan oleh ngengat betina yang aktif mencari dan mendapat-
kan inang yang sesuai' (Renwick & Chew 1994). Perilaku ngengat meletakkan telur beragam. Ada yang meletakkan telu; satu per satu atau berkelompok, dan
peletakan telur umumnya terbatas pada tanaman tertentu yang sudah berbunga
(Southwood 1978). Price (1986) menjelaskan pengaruh kondisi tanaman inang
16
Pastinaca sativa (parsnip)
yang diserang oleh ulat lamat(webworm).
Padatanaman yang sedikit menghasilkan bunga ditemukan sedikit ulat, sedangkan
pada yang banyak menghasilkan bunga ditemukan banyak ulat. Interaksi perilaku
serangga dengan kondisi tanaman inang seperti yang disebutkan itu menentukan
pola persebaran. Selain sumber daya, kerapatan populasi juga turut menentukan '
persebaran. Misalnya pada kerapatan populasi rendah, persebaran populasi
mungkin mengikuti pola acak (Taylor 1984). Tetapi dengan meningkatnya kera- patan, persebaran berubah menjadi mengelompok. Di samping itu, perilaku
kanibalisme juga dapat mempengaruhi pola persebaran. Kanibalisme biasanya
terjadi jika sumber daya makanan terbatas. Pada serangga yang bersifat kanibal,
persebaran populasinya cenderung mengikuti pola acak atau teratur. .
Musuh Alamai Helicoverpa spp.
Kerapatan populasi hama dapat berubah dari waktu ke waktu dan dari tempat ke tempat. Salah satu faktor yang mempengaruhi perubahan itu adalah musuh
alami, yang meliputi parasitoid, predator, dan patogen.
Parasitoid
Parasitoid merupakan unsur pengendali populasi hama yang umumnya bersifat
spesifik, sehingga dapat menekan populasi inangnya pada tingkat yang lebih
rendah. Sifat itulah yang menyebabkan parasitoid lebih sering digunakan dalam
pengendalian hayati dibanding dengan serangga predator (Price 1975).
Telur dan larva
Helicoverpa
spp. dapat diparasit oleh berbagai jenis parasi-toid. Pada masing-masing tanaman inang , jenis parasitoid serta tingkat parasitisa-
17
mendapatkan serangga inang pada umumnya melalui beberapa tahapan. Menurut
Doutt (1973) tahap-tahap tersebut adalah 1) penemuan habitat serangga inang, 2)
penemuan serangga inang, 3) penerimaan inang dan 4) kesesuaian inang. Dalam
proses tersebut tanaman inang memegang peranan penting sebagai habitat
-
serangga inang. Dout (1973) menjelaskan lebih jauh bahwa pada tanaman yangrnenghas ilkan madu atau nektar, betina parasitoid akan tertarik pada tanaman
tersebut walaupun tidak ada serangga inangnya. Hasil percobaan Sathe (1990)
memperlihatkan bahwa betina parasitoid Eriborus argenteopilosus awalnya terta- rik oleh daun tanaman inang dan kemuditm aktif melakukan pencarian serangga
inang. Faktor-faktor tanaman seperti adanya eksudat yang bersifat lengket pada
permukaan daun tanaman, adanya rambut yang banyak pada permukaan tanaman,
dan mekanisme antibiosis oleh tanaman merupakan ha1 yang dapat menjelaskan
mengapa komunitas parasitoid dan tingkat parasitisasinya dapat berbeda-beda
(Nyambo, 1 990).
Studi parasitoid Helicoverpa spp. telah banyak dilakukan di berbagai negara pada berbagai komoditi. Di Virginia (Arnerika Serikat), H. zea yang menyerang tanaman kedelai diparasit oleh Microplitis croceipes (Cresson) (Hymenoptera :
Braconidae). Parasitoid tersebut merupakan parasitoid larva instar I11 dan IV
dengan tingkat parasitisasi 93 hingga 98 %. Parasitoid larva yang kurang penting
adalah Cotesia marginiventris (Cresson) (Hymenoptera : Braconidae) , Compoletis flavicincta (Ashmead) (Hymenoptera : Ichneumonidae), Architas marmoratus
Tabel 2.1. Parasitoid Larva H. armigera pada Beberapa Tanaman Inang Di Tanzania. (Nyambo 1990).
Jenis parasitoid Tanarnan inang
Hymenop tera
Braconidae
Cardiochiles spp.
Apanteles sp.
Chelonus sp.
Meteorus sp.
Ichneumonidae Charops sp.
Netalia sp.
Pristomerus sp . Emelucha sp. Diptera
Tachinidae
Paradrimo halli (Curr)
Palexorista laxa (Curr)
C1, t, ch, k, s, j. C1, t, k, s, j.
C1, ch, k, s, j. C1, k.
C1, t, ch, k, s, j. CL, t, k, s, j. C1, t, k, s, j. C1.
C1, j, s, k, ch, t. .C1, j, s, k, ch, t.
Keterangan : C1 = Cleome sp., t = tomat, ch = chickpea, k = kapas, s = sorghum, j = jagung.
Di Indonesia larva H. armigera pada tanaman kapas diparasit oleh sejumlah parasitoid yang selengkapnya tercantum pada Tabel 2.2 (Nurindah & Bindra
1988).
Trichogramma spp. merupakan parasitoid telur Helicoverpa. Parasitoid penting dari telur H. zea di Amerika Serikat adalah T. exiguum, T. semifunctum, T. pretiosum dengan tingkat parasitisasi 46 - 100 % (Graham 1970; Hoffmann et al. 1990; Oatman & Platner 1971, 1978). Di Portugal telur H. armigera pada pertanaman tomat diparasit oleh T. rhenana Voegele & Rum dan Telenomus spp.
*
1990), sedangkan di Australia adalah T. australicum Girauet dan
T.
ivelae Pang& Chen (Zalucki et al. 1986). Di Indonesia telur H. armigera pada tanaman
kapas diparasit oleh Trichograrnma australicum, T. chilonis, T. chilotraea, T. japonicum, T. minutum, Trichogrammatoidea armigera, T. guanensis, dan T.
nana (Nurindah & Bindra 1988).
Tabel 2.2. Parasitoid Larva H. armigera pada Tanaman Kapas Di Indonesia (Nurindah & Bindra 1988)
Or do Farnili Spesies
Hymenoptera Braconidae Apanteles sp. Chelonus sp.
Microgaster demolitor Ichneumonidae Diadegma sp.
Enicospilus sp.
Eriborus argenteopilosus Sphecidae Ammophila insolita Vespidae Antepipona sp
.
Diptera ~achinidae* Carcelia kockiana
Carcelia sp.
Compsilura concinnata Exorista sp.
* oleh Nurindah & Bindra (1988) digolongkan dalam famili Sarcophagidae.
Predator
Selain parasitoid, predator mempakan faktor mortalitas yang mempengaruhi
perkembangan populasi hama. Berdasarkan pengamatan Nyambo (1990) di
Tanzania, predator H. armigera meliputi Chrysopa sp., semut, Coccinella sp.
,
larva Syrphidae, dan belalang Mantidae. ' Larva Chrysopa memangsa telur dan larva instar awal, dan semut memangsa semua instar larva (Reed 1965).Tabel 2.3. Predator H. armigera pada Tanaman Kapas di Indonesia . (Nurindah & Bindra 1988).
Inang Jenis Predator
-
Ordo Famili Spesies
Telur Coleoptera Coccinellidae Coccinella repanda
Menochilus sexmaculatus Scyrnnus roepkei
Verania discolor
V lineata
Nitidulidae
,
Cybocephalus sp. Staphylinidae Paederus fasciatus Hemiptera Anthocoridae Orius tantillusLy gaeidae Geocoris sp. Germalus sobrinus
Miridae Compylomma diversicomis
Deraeocoris indianus Neuroptera Chrysopidae Chrysopa sp.
Hemerobiidae Micromussp. Larva Coleoptera Staphylinidae Paederus fasciatus
Dermaptera Carcinophoridae Euborielia annulipes Forficulidae Exypnus pulchripennis Hemiptera Anthocoridae Orius tantillus
Lygaeidae Geocoris sp. Miridae Cryptopeltis sp
.
Deraecoris indianus Pentatomidae Eocanthecona furcellata
E. rufescens Reduviidae Rhinocoris fuscipes
Rhinocoris sp
.
Patogen
Sejumlah patogen yang ditemukan menginfeksi larva Helicoverpa spp. antara
lain NPV dan bakteri (Nyambo 1990). Zalucki et al. (1986) menyatakan bahwa cendawan (Beauveria bassiana dan Nomu+aea rileyi), protozoa (Noserna heliothi- dis) , dan nematoda {Heterorhabditis bacteriophora) merupakan patogen yang
dapat menyebabkan kematian pada larva Helicoverpa. Di Indonesia .larva H. annigera diserang oleh NPV dan jarnur Metarrhizium (Kalshoven 1981; Nurindah
[image:160.580.89.532.86.721.2]Pemanfaatan Parasitoid Telur
Pengendalian hayati merupakan inti dari pengendalian hama terpadu, dan
umumnya dilakukan dengan menggunakan parasitoid. Berdasarkan kepustakaan
.. yang ada, parasitoid telur telah banyak diteliti untuk pengendalian hama (Smith
1996). Pengendalian pada fase telur lebih baik karena telur belum menyebabkan
kerusakan.
Keberhasilan pengendalian hayati dengan parasitoid telur tergantung dari
beberapa faktor, di antaranya adalah spesjles yang digunakan, kualitas dan cara
pembiakan massal (Smith 1996). Pemilihan spesies yang baik untuk pelepasan
merupakan faktor yang paling penting karena spesies tertentu hanya dapat bera-
daptasi dengan baik pada kondisi iklim, habitat dan inang tertentu. Hassan (1994)
menyatakan bahwa spesies lokal lebih baik digunakan karena spesies tersebut
sudah beradaptasi dengan kondisi lingkungan setempat. Oleh karena itu, sebelum
pelepasan perlu dilakukan inventarisasi spesies parasitoid (Wajnberg 1994).
Kualitas parasitoid perlu mendapat perhatian berkaitan dengan keberhasilan
program pelepasan. Smith (1996) menyatakan bahwa parasitoid yang berkualitas
baik adalah yang mempunyai keperidian tinggi, lama hidup imago panjang,
persentase keturunan betina lebih banyak, preferensi yang tingg i, kemampuan
mencari inang yang baik, dan toleransi yang tinggi terhadap cuaca setempat.
Salah satu indikator yang biasa digunakan untuk mencirikan kualitas adalah
ukuran tubuh, yang ditunjukkan oleh panjang tibia tungkai belakang dan panjang
sayap. Kedua parameter tersebut dilaporkan berkorelasi positif dengan
keperidian, lama hidup, kemampuan mencari inang dan daya pencar (Bai et al.
22
Dalam pembiakan massal parasitoid, perhatian umumnya lebih banyak dituju-
kan pada mudahnya pelaksanaan pembiakan agar diperoleh parasitoid dalam
jumlah besar, dan kurang memperhatikan kualitas parasitoid yang dihasilkan.
Oleh karena itu, tidak jarang hasil pelepasan parasitoid tidak memuaskan (Smith
1996).
Upaya perbanyakan massal parasitoid telur umumnya menggunakan telur
Sitotroga cerealella, Ephestia kuehniella dan Corcyra cephalonica (Flanders
1930). Namun hasil penelitian belakangan ini menunjukkan bahwa dengan
menggunakan telur inang yang berukuran lebih besar, seperti telur ulat sutera
(silkworm) dan Ostrinia nubilalis, dapat dihasilkan parasitoid yang lebih besar
dan persentase betina y.ang lebih banyak (Burbutis & Koepke 198 1; Li 1982).
Di Amerika Serikat, parasitoid telur Tnchogramma telah dimanfaatkan dalam
program PHT
H.
zea (Oatman & Platner 1971, 1978; Oatman et al. 1983). Di Indonesia, pelepasan Trichogramma sebagai pengendali hayati H. armigera pernah dilakukan pada pertanaman kapas (Nurindah 1989). Dari pengalaman diluar negeri, hasil pelepasan Tnchogramrna dalarn menekan hama sangat beragam.
Oatman & Platner (1 97 1) melaporkan bahwa pelepasan 32.800 T. pretiosumlminggu/0,4 ha, menghasilkan rataan parasitisasi 55,7 % , sedangkan
dengan pelepasan 40.200 parasitoid/minggu/0,4 ha parasitisasi meningkat menjadi
8 1,2 %
.
Penelitian lain menunjukkan dengan pelepasan 200.000 parasitoid/0,4ha diperoleh tingkat parasitisasi 53,l %, sedangkan dengan pelepasan 3 18.000
parasitoid/0,4 ha selama pertumbuhan tanahan tomat diperoleh tingkat parasitisa-
si 85,4 %. Oatman & Platner (1971) menyimpulkan bahwa keberhasilan pelepasan parasitoid Trichogramrna ditentukan oleh banyaknya parasitoid yang
Daftar Pustaka
Adjei-Maafo, I.K. & L.T. Wilson. 1983. Association of cotton nectar production with Heliothis punctigera (Lepidoptera : Noctuidae) oviposition. Environ. Entomol. 12 : 1166-1170.
Bai, S., S. Cobanoglu & S. M. Smith. 1995. Assessment of Trichogramma spe- cies for biological control of forest lepidopteran defoliators. Entomol. Exp. Appl. 75 : 135
-
143.Bigler, F. 1994. Quality control in Trichogramma production. Dalam Wajnberg, E. & S. A. Hassan (ed). Biological control with egg parasitoids. Oxon, UK. CAB International. Hal. 93
-
1 12.Broadley, R. H. 1978. Distribution of Hekothis spp. (Lepidoptera : Noctuidae) eggs and 1st instar larvae on pre-flowering flue-cured tobacco plants in north Queensland. Queensl. J. Agric. Anim. Sci. 35 : 73-76.
Brown, E.H. & B.K. Cantrell. 1978. Key to the larva of the common species of army worm, cutworm and budworms found in Queensland. Queensl. J. Agric. Anim. Sci. 35 : 91-94.
Budi, J., Anggara H. P. & Etti P. 1986. Introduksi dan evaluasi tomat (Lycoper- sicon esculentum) asal AVRDC. Bul. Pen. Hort. 16 (2) : 1-7.
Burbutis, P.P. & C.H. Koepke. 1981. European corn borer control in peppers by
Trichogramma nubilale. J . Econ. Entomol. 74 : 246 - 247.
Callahan, P.S. 1957. Oviposition response of the corn eanvorm to differences in surface texture. J. Kans. Entomol. Soc. 30 : 59
-
63.Common, I. F.B. 1953. The Australian species of Heliothis (Lepidoptera : Noc- tuidae) and their pest status. Aust. J. Zool. 1 : 319-344.
Cullen, J.M. 1969. The reproduction and survival of Heliothis punctigera
Wallengren in South Australia, Disertasi, University of Adelaide.
Daly,J. & P. Gregg. 1985. Genetic variation in Heliothis in Australia : Species identification and gene flow in the two pest species H. armigera (Hubner) and H. punctigera (Wallengren) (Lepidoptera : Noctuidae). Bull. Ento- mol. Res. 75 : 169-184.
Doutt, R.L. 1973. Biological characteristics of entomophagous adult. Dalam
DeBach, P. (Ed). Biological Control of Insect Pests and Weeds; Chap- man and Hall Ltd., London. Hal. 145-167.
Etti, P. & Ali, A. 1990. Seleksi varietas tomat untuk perbaikan kualitas. Bul. Pen. Hort. 20 (1): 98-105.
.,
Hanudin, A. Ali, & G. Setiani. 1990. Seleksi progeni tomat tahan terhadap bakteri layu dan berkualitas baik di dataran rendah. Bul. Pen. Hort. V18 (2): 89-97.Flanders, S.E. 1930. Mass production of egg parasites of the genus Trichogram- ma. Hilgardia. 4 : 465 - 501.
Goyal, S. P. & V. S. Rathore. 1988. Pattern of insect plant relationship deter- mining susceptibility of different hosts to Heliothis armigera Hiibner. Indian J. Entomol. 50(2) : 193-201.
Graham, H.M. 1970. Parasitism of eggs of bollworms, tobacco budworms and loopers by Trichogramma semifunctum (Hymenoptera : Trichogrammati- dae) in the lower Rio Grande Valley, Texas. J. Econ. Entomol. 63 : 686- 688.
Gupta, P.D., & A.J. Thorsteinson. 1960. Food plant relationships of the dia- mondback moth (Plutella maculipennis (Curt.)) 11. Sensory regulation of oviposition the adult female. Entomol. Exp. Appl. 3 : 305
-
314.Hassan, S
.
T.
S . 1985. Distribution of Heliothis armigera (Hiibnner) and Heliothis punctigera (Wallengren) (Lepidoptera : Noctuidae) eggs and larvae, and insecticide spray droplets on cotton plants. Disertasi, University of Queensland.Hassan, S.A. 1994. Strategies to select Trichogramma spesies for use in biologi- cal control. Dalam Wajnberg, E. & S. A. Hassan (ed). Biological control with egg parasitoids. Oxon UK. CAB International. Hal. 55
-
73.Haynes, K.F., J.Z. Zhao, & A. Latif. 1991. Identification of floral compounds from Abelia grandij7ora that stimulate upwind fligth in cabbage looper moths. J. Chem. Ecol. 17 : 637
-
646.Heath, R.R, P.J. Landolt, B. Dueben, & B. Lenczewski. 1992. Identification of floral compounds of nigth-blooming Jessamine attractive tocabbage looper moths. Environ. Entomol. 21 : 854 - 859.
Hoffmann, M.P., L.T. Wilson, F.G. Zalon & R. J. Hilton. 1990. Parasitism of Heliothis zea (Lepiodoptera : Noctuidae) eggs : Effect on pest manage- ment decision rules for processing'tomatoes in the Sacramento Valley of California. Environ. Entomol. 19(3) : 753-763.
Kakar, K.L., J.P. Sharma, & G.S. Dogra. 1990. Feasibility of using
Tricho-
gramma
spp. againstHeliothis armigera
Hilbner on tomato. Indian J. Plant Protec. 18(2): 237-239.Khan, Z.R., A. Ciepiela, & D.M. Norris. 1987. Behavioral and physiological responses of cabbage looper,
Trichoplusia ni
(Hbn), to steam distillates from resistant versus susceptible soybean plants. J. Chem. Ecol. 13 : 1903-
1915.Kirkpatrick, T. H
.
196 1. Comparative morphological studies ofHeliothis
species (Lepidoptera : Noctuidae) in Queensland. Queensl. J. Agric. Sci. 18 :179-194.
Li, L.Y. 1982. Integrated rice pest control in the Guangdong Province of China. Entomophaga. 27 : 81
-
88.t
Liu, S. H., D.M. Norris, & E. Marti. 1988. Behavioral responses of female adult
Trichoplusia ni
to volatiles from soybeans versus a preferred host, lima bean. Entomol Exp. Appl. 49 : 99-
109.Luckmann, W.H. & R.L. Metcalf. 1982. The pest management concept.
Dalam
Metcalf & Luckmann (Ed), Introduction To Insect Pest Management. John Wiley & Sons, Inc. New York, London, Sydney, Toronto. Hal. 1-29.Meierrose, C., I. Silva, & J. Araujo. 1991. Egg parasitoids from
Heliothis
armigera
on tomato in Southern Portugal. Colleques-de-1 IRNA. No. 56 :22 1-226.
Meierrose, C. & J. Araujo. 1986. Natural egg parasitism of
Helicoverpa (Helio-
this) armigera
Hbn. (Lepidoptera : Noctuidae) in South Portugal. Z.Angew. Entomol. 101: 11-18.
Mitter, C., R.W. Poole & M. Mathew. 1993. Biosystematics of the Heliothinae (Lepidoptera : Noctuidae). Annu. Rev. Entomol. 38 : 207
-
225.Nurindah. 1989. Studies on
Trichogramma
spp.
(Hymenoptera : Trichogram- matidae) in the control ofHeliothis armigera
(Hiibner) (Lepidoptera :Noctuidae). Biotrop Spec. Publ. 36 : 165 - 172.
.
& O.S. Bindra. 1988. Studies on biological control of cotton pests. Indust. Crops Res. J. l(1) : 59-
83.Oatrnan, E.R., J.A. Wyman, R.A. van Steenwyk & M.W. Johnson. 1983. Inte- grated control of the tomato fruitworm (Lepidoptera : Noctuidae) and other lepidopterous pests on fresh-market tomatoes in Southern California. J. Econ. Entomol. 76 : 1363-1369.
Oatman, E.R. & G.R. Platner. 1971. Biological control of tomato fruitworm, cabbage looper and hornworms on processing tomatoes in Southern Cali- fornia, using mass releases of Trichogramma pretiosum. J. Econ. Ento- mol. 64 : 501-506.
.
1978. Effect of mass releases of Tn'cho- gramma pretiosum against lepidopterous pests on processing tomatoes in Southern California, with notes on host egg population trends. J. Econ. Entomol. 71 : 896-900Persson, B. 1976. Influence of weather an'd nocturnal illumination on the activity and abundance of populations of noctuids (Lepidoptera) in south costal Queensland. Bull. Entomol. Res. 66 : 33-63.
Price, P. W. 1975. Insect Ecology. John Wiley & Son, Inc. New York, London, Sydney, Toronto, 5 14 hal.
Price, P.W. 1986. Ecological aspects of host plant resistance and biological control : Interactions among three trophic level. Dalam Boethel, D.J. & R.D. Eikenbary (ed). Interactions of plant resistance and parasitoids and predators of insects. Hal. 1
-
30.Reed, W. 1965. Heliothis armigera (Hb) (Noctuidae) in Western Tanganyika 11. Ecology and natural and chemical control. Bull. Entomol. Res. 56: 127- 140.
Renwick, J. A. A. & F. A. Chew. 1994. Oviposition behaviour in Lepidoptera. Annu. Rev. Entomol. 39 : 377 - 400.
Saour, G. & R. Causse. 1993. Oviposition behaviour of Heliothis armigera
' Hiibner (Lepidoptera : ~octucdae) on tomato. J. Appl. Entomol. 115(2):
203-209.
Sathe, T.V. 1990. Impact of host food plant on parasitization behaviour in a larval parasitoid of Heliothis armigera (Hubner). Anim. Sci. 99(3) : 233- 242.
Setiawati, W. 1991. Daur hidup ulat buah tomat, Heliothis armigera Hbn (Lepi- doptera : Noctuidae). Bul. Penel. Hort. 20 (4): 1 12-1 17.
Singh, A.K. & H. Rembold. 1988. Developmental value of chickpea, Cicer arie- tinum, soybean, Glycine max and maize,zea mays flour for Heliothis armigera (Lepidoptera : Noctuidae) larvae. J. Appl. Entomol. 106 (3) :
Smith, S. M. 1996. Biological control with Trichograrnma : Advances, successes, and potential of their use. Annu. Rev. Entomol. 41 : 375
-
406.Southwood, T. R. E. 1978. Ecological Methods with Particular Reference to The Study of Insect Populations. London. Chapman & Hall. 524 hal.
Stanley, S. M. 1978. Comparative interactions between the larvae of Heliothis armigera (Hiibner) and Heliothis punctigera (Wallengren) (Lepidoptera :
Noctuidae). Ph.D. Thesis. Australian National University, Carnberra.
Stern, V.M., R.F. Smith, R. van den Bosch & K.S. Hagen. 1959. The Integrated control concept. Hilgardia 29 (2): 81
-
101.Taylor, L. R. 1984. Assessing and interpreting the spatial distributions of insect populations. Annu. Rev. Entomol. 29 : 321 - 357.
I
Wajnberg, E. 1994. Intra-population genetic variation in Trichogramrna. Dalam
Wajnberg, E. & S. A. Hassan (ed). Biological control with egg parasi- toids. Oxon, UK. CAB International. Hal. 245
-
273.Wardhaugh, K. G., P. N. Room, & L. R. Greenup. 1980. The incidence of Helio- this armigera and H. punctigera (Lepidoptera : Noctuidae) on cotton and other host plants in the Namoi Valley of New South Wales, Australia. Bull. Entomol. Res. 70: 113-132.
Wiesenborn, W. D. & T. C. Baker. 1990. Upwind fligth to cotton flowers by
~ e c t i n o ~ h o r a gossypiella ( ~ e ~ i d o p t e r a : ~elechGdae). Environ. ~ntomoi. 19 : 490 - 493.
Wilcox, J., A.F. Howland, & R.E. Campbell. 1956. Investigations of the tomato fruitworm : Its seasonal history and methods of control. USDA. Tech. Bull. No. 1147. 47 hal.
Wilson, A.G.L. 1976. Varietal responses of grain sorghum to infestation by
Heliothis armigera. Exp. Agric. 12: 257- 265.
Wilson, A.G.L., J.J. Basinski & N. J. Thomson. 1972. Pests, crop damage and control practices with irrigated cotton in a tropical environment. Cotton Grow. Rev. 49 : 308
-
340.Zalucki, M.P., G. Daglish, S. Firempong & P. Twine. 1986. The biology and ecology of Heliothis armigera (Hubner) and H . punctigera Wallengren (Lepidoptera : Noctuidae) in Auaralia : What do we know?. Aust. J. 2001. 34 : 779 - 814.
BAB I11
HUBUNGAN ANTARA PELETAKAN TELUR
Helicoverpa armigera
(HUBNER) (LEPIDOPTERA : NOCTUIDAE) DENGAN PEMBUNGAAN TANAMAN TOMAT
ABSTRAK
-
Penelitian bertujuan untuk mengetahui hubungan antara peletakan telur Helicoverpa annigera
(Hiibner)(Lepidoptera : Noctuidae) dengan pembungaan tanaman tomat. Penelitian dilakukan di
Desa Tugu Selatan, Kecamatan Cisarua, Kabupaten Bogor pada musim kemarau 1995. Penelitian
dilakukan pada lahan seluas f 600 m2 yang ditanami tomat varietas TW 375. Populasi H. armigera berasal dari infestasi secara alami. Pengamatan telur dilakukanpada 20 satuan contoh, sedangkan pengamatan jumlah bunga dilakukan patla 10 satuan contoh, dimulai sejak tanaman
berumur 15 hst hingga f 100 hst. Pengamatan telur dibedakan berdasarkan letalcnya pada daun,
bunga, batang, dan buah. Hasil penelitian menunjukkan bahwa terdapat korelasi yang kuat antara
peletakan telur dengan jumlah bunga mekar. H. armigera mulai meletakkan telur pada perta- naman tomat pada saat kuncup bunga mulai mekar. Perkembangan populasi telur mengikuti
perkembangan ketersediaan bunga mekar. Puncak populasi telur terjadi hampir bersamaan dengan
puncak ketersediaan bunga mekar. Telur pada umumnya diletakkan pada permukaan atas daun
serta tangkai bunga, rambut tangkai bunga, dan kelopak bunga. Sebagian kecil dari telur diletak-
kan pada kelopak buah dan kadang-kadang pada permukaan buah serta pada batang. Telur lebih
banyak diletakkan pada setengah bagian atas tajuk, tempat terdapatnya bunga mekar. Dengan
demikian, upaya pengamatan telur selayaknya dimulai pada saat kuncup bunga mulai mekar dan
pemeriksaan telur itu diarahkan pada tajuk bagian atas.
Pendahuluan
Ulat
H. armigera
adalah hama utama pada pertanaman tomat. Hama inimenimbulkan kerugian karena larvanya menggerek ke dalarn buah. Pada keadaan
serangan berat, kerusakan buah tomat dapat mencapai 80 % (Uhan & Suriaat- madja 1993). Usaha untuk mendapatkan cara pengendalian yang efektif memer-
lukan pemahaman yang menyeluruh mengenai kehidupan hama, termasuk di
2 9
pemahaman ini, waktu pemantauan dan bagian tanaman yang diamati dapat
ditentukan dengan tepat. Hubungan peneluran Helicoverpa spp dengan pembun- gaan tanaman inang seperti kedelai, kapas, tembakau, sorghum telah banyak
dilaporkan (Broadly 1978; Wardhaugh et al. 1980; Adjei-Maafo & Wilson 1983; Wilson, Bansiski & Thomson 1972; Wilson 1976). Pada tanaman tomat, infor-
masi peneluran Helicoverpa spp. masih terbatas pada preferensi peletakan telur pada bagian tanaman (Rodriguez, Leigh, & Lange 1982; Snodderly & Lambdin
1982; Saour & Cause 1993). Informasi tentang hubungan peneluran H. I
armigera dengan pembungaan tanaman tomat belum banyak diketahui. Penelitian ini bertujuan untuk mengkaji hubungan antara peneluran H. armigera dengan pembungaan tanaman tomat.
Bahan dan Metode
Penelitian dilaksanakan di lahan milik petani di Desa Tugu Selatan, Keca-
matan Cisarua, Kabupaten Bogor mulai Juli hingga Oktober 1995. Lokasi peneli-
tian berjarak f 2 krn dari Perkebunan Teh Gunung Mas. Sepanjang tahun, lahan
di sekitar lokasi penelitian ditanami sayur-sayuran terutama tomat, kubis, buncis,
petsai, bawang daun, wortel, dan jagung. Pada umumnya petani setempat
menanarn tomat secara tumpang gilir dengan kubis atau petsai. Kubis atau
kadang-kadang petsai ditanam lebih awal, dan setelah tanaman berumur satu
bulan atau lebih di antara tanaman kubis atau petsai ditanami tomat. Kadang-
kadang setelah tomat menghasilkan buahl9di antara tanaman tomat ditanami lagi
dengan buncis.
Lahan seluas f 600 m2 ditanami tomat varietas Taiwan 375, salah satu
30
digunakan adalah 50 cm
x
60 cm, sehingga seluruhnya terdapat f 2.000 tanaman. Budidaya tomat yang dilakukan pada petak percobaan mengikuti cara petanisetempat. Pertanaman tomat dipupuk dua kali dengan NPK (15- 15- 15) dengan dosis 650 kglha. Pemupukan pertama (setengah dari dosis) dilakukan pada saat
tanaman berumur 14 hst, dan pemupukan kedua pada umur 35 hst. Selain itu
dilakukan juga pemangkasan, pembuatan guludan, penyiangan, penyiraman,
pengajiran, dan pengendalian penyakit dengan penyemprotan fungisida. Pemang-
kasan tunas atau cabang non-produktif dilakukan sebanyak tiga kali, yaitu pada
I
saat tanaman berumur 28, 42, dan 56 hst. Pembuatan guludan dan penyiangan
gulma dilakukan setelah pemupukan pertama, dan pengajiran dilakukan pada saat
tanaman berumur 30 hst. Tiap empat tanaman yang berdekatan, ujung atas ajir-
nya diikat menjadi satG agar tanaman tidak mudah rebah.
Pengamatan dilakukan terhadap banyaknya telur, saat berbunga, dan
banyaknya bunga mekar. Telur diamati pada 20 satuan contoh tetap selama
pertumbuhan tanaman. Tiap satuan contoh terdiri dari empat tanaman yang ada
dalarn satu ikatan ajir. Satuan contoh ditentukan secara sistematik, dan diupaya-
kan menyebar di seluruh petak pertanaman. Telur yang ditemukan dibedakan
berdasarkan letaknya pada daun, bunga, buah dan batang.
Pengamatan bunga dilakukan
pads'
10 satuan contoh dari 20 satuan contohyang digunakan untuk pengamatan telur. Penentuan 10 dari 20 satuan contoh
tersebut dilakukan dengan mengambil secara acak dua satuan dari empat satuan
contoh yang ada pada tiap baris tanaman* contoh. Pengamatan dilakukan setiap
Hasil
Hubungan Penelwan dengan Pembungaan
Peletakan telur mempunyai kaitan yang erat dengan pembungaan. Telur
pertama kali ditemukan pada saat tanaman mulai menghasilkan bunga mekar,
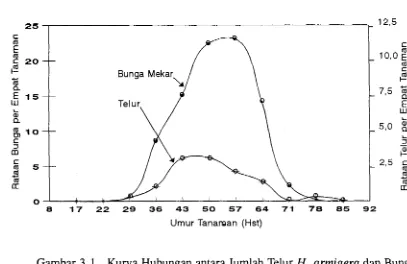
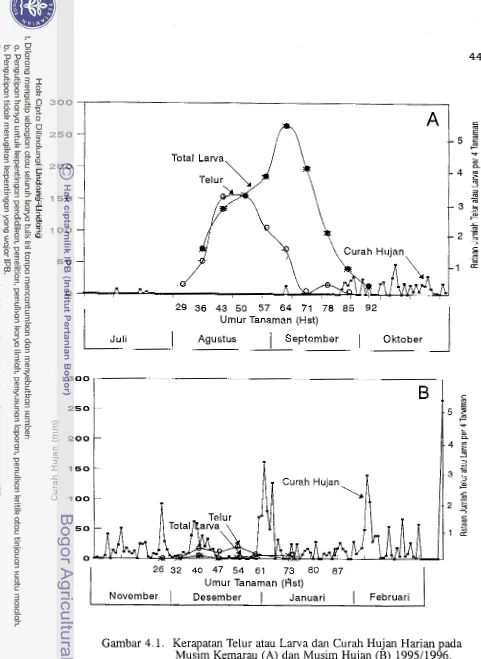
yaitu pada umur 29 hst (Gambar 3.1). Populasi telur meningkat seiring dengan
bertambahnya jumlah bunga mekar. Puncak peletakan telur (3,l butir per empat
tanaman) terjadi pada umur 50 hst, atau seminggu sebelum puncak pem- bungaan (23,l bunga mekar per empat tanaman). Setelah itu, populasi telur
menurun hingga pada umur 85 hst, dan pada umur 92 hst telur tidak ditemukan lagi (Gambar 3.1). Pada umur itu tanaman tidak menghasilkan bunga lagi se- hingga tidak menarik bagi ngengat H.
amzigera.
[image:171.586.119.526.381.645.2]Umur Tanaman (Hst)
Gambar 3.1. Kurva Hubungan antara Jumlah Telur H.
armigera
dan Bunga Mekar pada Tanaman Tomat.32
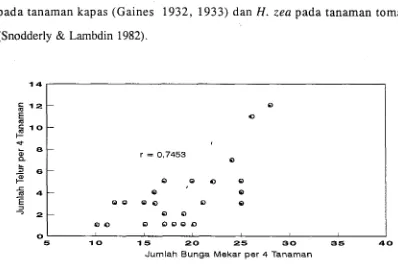
rung berkorelasi positif dengan banyaknya bunga mekar. Adanya keterkaitan
antara peletakan telur dengan pembungaan juga dilaporkan untuk H. obsoleta
pada tanaman kapas (Gaines 1932, 1933) dan H. zea pada tanaman tomat (Snodderly & Lambdin 1982).
Jumlah Bunga Mekar per 4 Tanaman
Garnbar 3.2. Diagram Pencar Hubungk antara Jumlah Telur
H.
armigeradengan Jumlah Bunga Mekar.
Persebaran telur pada tanaman. Telur umumnya ditemukan pada daun dan bunga, tetapi kadang-kadang juga pada batang dan buah. Pada bunga, telur
ditemukan pada rambut tangkai bunga, kelopak bunga, dan kadang-kadang pada
tangkai dan mahkota bunga. Pada daun, telur ditemukan pada permukaan atas
daun, sedangkan pada buah ditemukan pada kelopak buah dan permukaan buah.
Banyaknya telur yang ditemukan pada berbagai bagian tanaman ditunjukkan pada
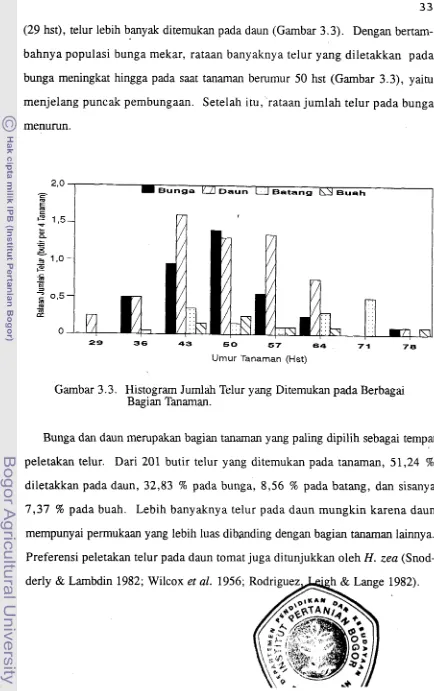
Gambar 3.3. Analisis statistik menunjukkan bahwa rataan banyaknya telur pada
[image:172.582.128.526.140.407.2]3 3
(29 hst), telur lebih banyak ditemukan pada daun (Gambar 3.3). Dengan bertam-
bahnya populasi bunga mekar, rataan banyaknya telur yang diletakkan pada
bunga meningkat hingga pada saat tanaman berumur 50 hst (Gambar 3.3), yaitu
menjelang puncak pembungaan. Setelah itu, rataan jumlah telur pada bunga
menurun.
2,o
m
Bunga k d Daun Batang h Y BuahEII
m
5
c
5
1,s- I29 3 6 43 5 0 57 64 71 78
[image:173.584.101.535.68.759.2]Umur Tanaman (Hst)
Gambar 3.3. Histogram Jumlah Telur yang Ditemukan pada Berbagai Bagian Tanaman.
Bunga dan daun merupakan bagian tanarnan yang paling dipilih sebagai tempat
peletakan telur. Dari 201 butir telur yang ditemukan pada tanaman, 51,24 %
diletakkan pada daun, 32,83 % pada bunga, 8,56 % pada batang, dan sisanya 7,37 % pada buah. Lebih banyaknya telur pada daun mungkin karena daun mempunyai pennukaan yang lebih luas dibanding dengan bagian tanman laimya.
34
Pembahasan
Peletakan telur merupakan tahapan terakhir dari proses penemuan inang.
Proses penemuan inang itu sendiri diawali dengan invasi yaitu penemuan habitat
inang yang diikuti dengan penemuan inang. Dalam tahapan penemuan inang, -.
ngengat tanggap terhadap aroma kimia (chemical odour) atau warna dari tanaman.
Untuk ngengat H. armigera faktor kimia mungkin lebih berperan daripada warna, karena warna tidak penting bagi ngengat yang aktif dan meletakkan telur
pada malam hari (Renwick & Chew 1994). Keterkaitan antara peletakan telur
dengan adanya bunga diduga karena bunia menghasilkan senyawa volatil yang
menarik ngengat untuk datang dan hinggap pada tanaman. Senyawa volatil
tersebut mungkin berhubungan dengan kesesuaian inang.
Ngengat H. armigera tertarik oleh aroma senyawa kimia dari bunga tanaman tomat. Oleh karena itu ngengat H. arnzigera menginvasi dan meletakkan telur pada waktu tanaman mulai menghasilkan bunga mekar. Ngengat H.
armigera berasal dari generasi y ang berkembang pada tanaman inang (tomat,
jagung, dan buncis) di sekitar tempat penelitian. Tanaman inang tersebut ditanam
sepanjang tahun dan pada waktu yang sama terdapat berbagai tingkatan umur
mulai tanaman muda sarnpai tanarnan tua. Sistem pertanaman seperti itu meru-
pakan kondisi yang baik untuk perkembangan
H.
armigera. Dalam satu tahun,H.
armigera menyelesaikan generasinya secara bergantian pada berbagai tanamaninang
.
Setelah menemukan tanaman inang, ngengat melakukan pencarian tempat
peletakan telur yang sesuai. Renwick & Chew (1994) menyatakan bahwa setelah hinggap pada t&xunan, ngengat melakukan evaluasi permukaan tanaman. Dalam
35
peranannya. Ngengat mempunyai indera mekanik dan kimia yang terdapat pada
tarsi, antena, probosis, dan ovipositor untuk mendeteksi keadaan permukaan bagian tanaman yang sesuai untuk peletakan telur (Renwick & Chew 1994).
Tampaknya untuk H. armigera faktor fisik, yaitu tekstur permukaan tanaman berperan dalam menentukan kesesuaian tempat peletakan telur, Ngengat mele-
takkan telur pada bagian tanarnan yang berambut dan kasar seperti tangkai bunga,
kelopak bunga dan buah, batang, dan permukaan atas daun. Perilaku yang sama
ditunjukkan oleh H. punctigera dan H . armigera pada tanarnan kapas (Cullen 1969; Hassan 1985). Renwick & Chew ('1994) menyatakan bahwa pada bagian
tanarnan yang berambut dan kasar, ngengat dapat berpijak dengan baik selarna
peletakan telur. Selain itu telur dapat melekat dengan baik d m tidak mudah terle-
pas.
Daun, batang dan buah yang diletaki telur adalah yang berdekatan dengan
tandan bunga. Karena tandan bunga terletak pada setengah bagian atas tanaman,
maka populasi telur umumnya terdapat pada bagian tersebut. Hal ini mengisya- ratkan bahwa upaya pengamatan telur sebaiknya diarahkan pada setengah bagian
atas tanarnan.
Kesimpulan
Ngengat H. armigera menginvasi pertanaman tomat dan meletakkan telur sejak tanaman menghasilkan bunga mekar. Pola perkembangan populasi telur H. armigera berkaitan erat dengan kelimpahan ketersediaan bunga mekar. Telur ditemukan pada daun, batang dan buah yang berdekatan dengan tandan bunga
yang sedang mekar, yaitu pada setengah bagian atas tanaman. Dengan demiki-
an, upaya pengamatan telur hendaknya dilakukan sejak kuncup bunga mulai
Daftar Pustaka
Adjei-Maafo, I.K. & L.T. Wilson. 1983. Association of cotton nectar production with Heliothis punctigera (Lepidoptera : Noctuidae) oviposition. Environ. Entomol. 12 : 1166-1170.
Broadley, R. H. 1978. Distribution of Heliothis