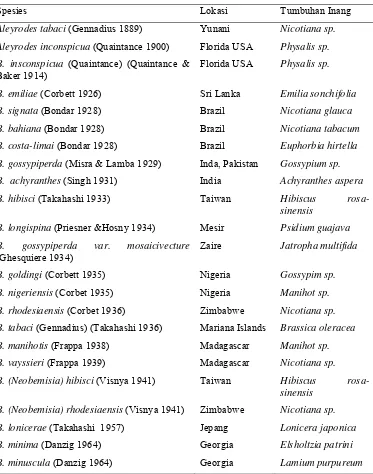
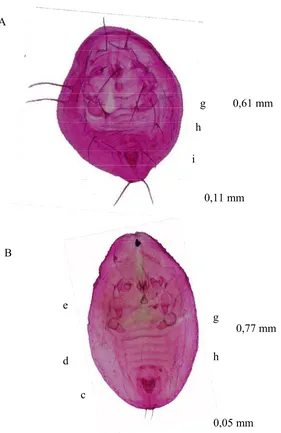
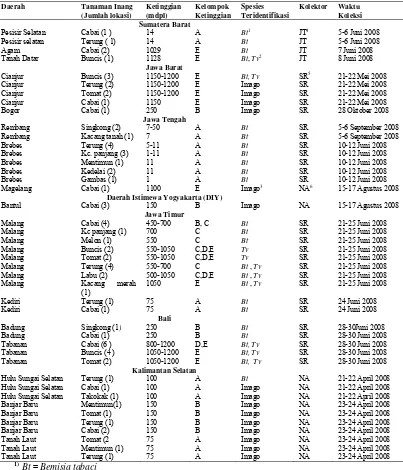
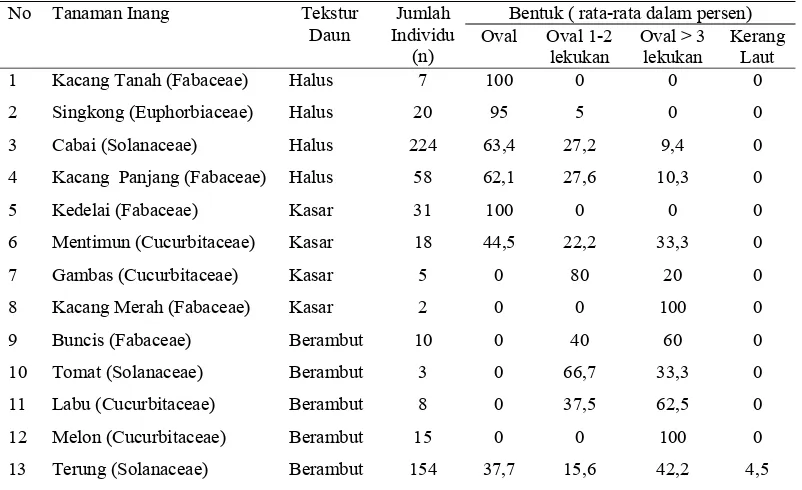
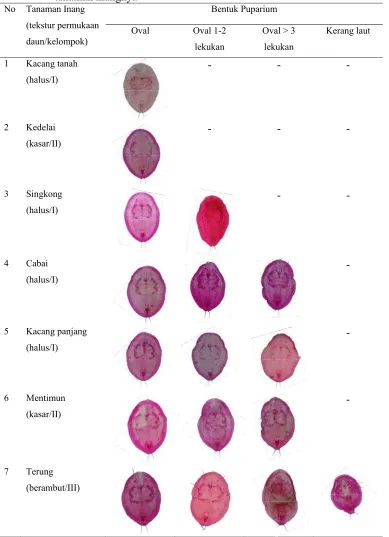
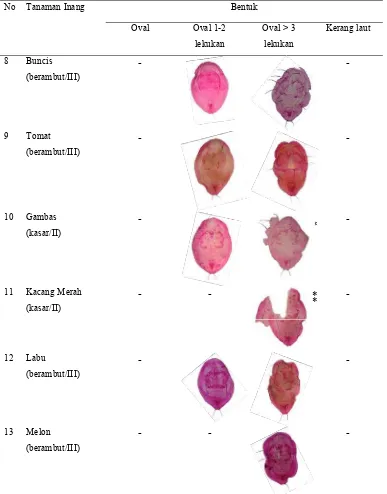
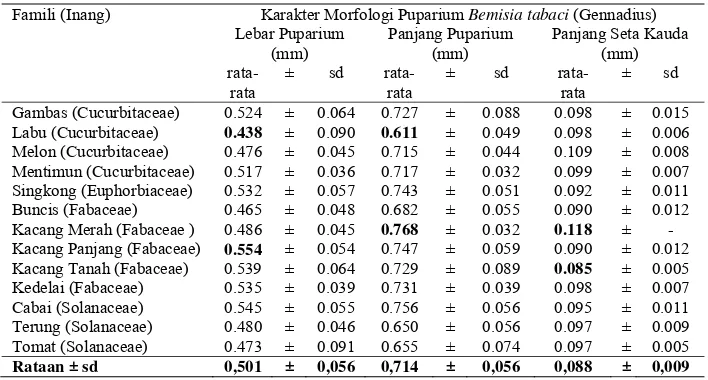
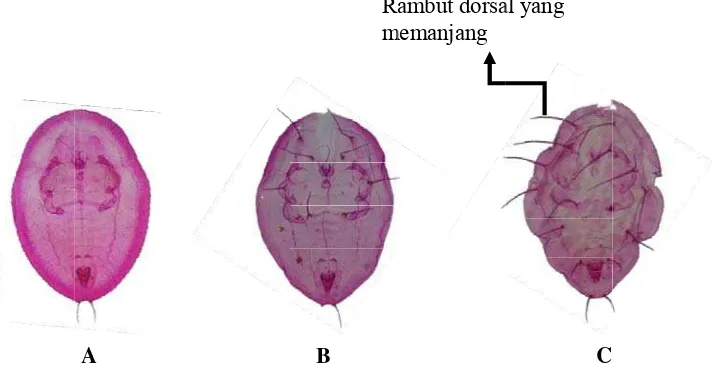
Variasi Morfologi Puparium dan DNA Penyandi Gen Mitokondria Sitokrom Oksidase I Bemisia tabaci (Gennadius) (Hemiptera Aleyrodidae)
Bebas
109
0
0
Teks penuh
Gambar


+7
Dokumen terkait
