ISOLASI DAN SELEKSI BAKTERI SELULOLITIK
PENGHAMBAT PERTUMBUHAN CENDAWAN PADA
TANAMAN KELAPA SAWIT
DHYAH PURNAMASARI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Isolasi dan Seleksi Bakteri Selulolitik Penghambat Pertumbuhan Cendawan pada Tanaman Kelapa Sawit adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
ABSTRAK
DHYAH PURNAMASARI. Isolasi dan Seleksi Bakteri Selulolitik Penghambat Pertumbuhan Cendawan pada Tanaman Kelapa Sawit. Dibimbing oleh NISA RACHMANIA MUBARIK dan SRI LISTIYOWATI.
Bakteri selulolitik tertentu dapat digunakan sebagai penghambat pertumbuhan cendawan. Penelitian ini bertujuan mengisolasi, menapis, dan menguji potensi bakteri selulolitik yang diisolasi dari tanah perkebunan karet dan kelapa sawit sekitar Taman Nasional Bukit Dua Belas, Jambi. Penapisan dilakukan dengan metode Fokkema, sedangkan pengujian daya hambat terhadap Curvularia sp. dan Colletotrichum sp. dengan metode sumur dan kertas cakram yang berisi kultur atau enzim selulase ekstrak kasar SAHN13.30 dan KAHN15.39. Sebanyak 40 bakteri selulolitik berhasil diisolasi dengan nilai indeks selulolitik 0.03-16.6. Hasil penapisan diperoleh daya hambat bakteri SAHN13.30 dan KAHN15.39 terhadap Curvularia sp. adalah sama (57.5%). Begitu juga daya hambatnya terhadap Colletotrichum sp. (60%). Kultur SAHN13.30 menunjukkan aktivitas penghambatan maksimum (45.95%) terhadap Curvularia sp. dengan metode sumur dibandingkan dengan metode lainnya. Hal yang sama aktivitas maksimum pada kultur KAHN15.39 terhadap Curvularia sp. adalah 46.26%. Sebaliknya, kultur KAHN15.39 memiliki daya hambat maksimum (50%) terhadap Colletotrichum sp. dengan metode kertas cakram. Kultur sel SAHN13.30 dan KAHN15.39 menunjukkan daya hambat terhadap Curvularia sp. maupun Colletotrichum sp. lebih besar dibandingkan dengan enzim selulase ekstrak kasarnya.
Kata kunci: bakteri selulolitik, Colletotrichum sp., Curvularia sp., kelapa sawit
ABSTRACT
DHYAH PURNAMASARI. Isolation and Screening of Cellulolytic Bacteria Inhibit Fungal Growth on Oil Palm Tree. Supervised by NISA RACHMANIA MUBARIK and SRI LISTIYOWATI.
Some cellulolytic bacteria could be used to control growth of fungal pathogens. This study aimed to isolate and to screen the potency of the soil cellulolytic bacteria isolated from rubber and oil palm trees grown surroundings the Taman Nasional Bukit Dua Belas, Jambi. Fourty isolates of cellulolytic bacteria were obtained and they had cellulolytic index activity from 0.03 to 16.6. Two isolates viz SAHN13.30 and KAHN15.39 could inhibit growth of Curvularia sp. and Colletotrichum sp. as much as 57.5% and 60%, respectively by the Fokkema method. Inhibit test to Curvularia sp. and Colletotrichum sp. can do with some method: by using well diffusion method and by using paper disc method of cultures and the crude axract of each cultures of SAHN13.30 and KAHN15.39. Maximum inhibition activity of SAHN13.30 culture on Curvularia sp. was 45.95% by using well diffusion method. Maximum inhibition of KAHN15.39 culture on Colletotrichum sp. was 50% by using paper disc method. On the contrary, maximum inhibition of KAHN15.39 culture on Curvularia sp. was 46.26% by using well diffusion method. The SAHN13.30 and KAHN15.39 cultures had the potency to inhibit fungal growth of Curvularia sp. and Colletotrichum sp. higher than the crude extract of each culture.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biologi
ISOLASI DAN SELEKSI BAKTERI SELULOLITIK
PENGHAMBAT PERTUMBUHAN CENDAWAN PADA
TANAMAN KELAPA SAWIT
DHYAH PURNAMASARI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
Judul Skripsi : Isolasi dan Seleksi Bakteri Selulolitik Penghambat Pertumbuhan Cendawan pada Tanaman Kelapa Sawit
Nama : Dhyah Purnamasari NIM : G34090089
Disetujui oleh
Dr Nisa Rachmania Mubarik, MSi Pembimbing I
Dr Sri Listiyowati, MSi Pembimbing II
Diketahui oleh
Dr Ir Iman Rusmana, MSi Ketua Departemen
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah Subhanahu wa Ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Penelitian yang dilaksanakan sejak bulan Januari 2013 ini ialah biofungisida, dengan judul Isolasi dan Seleksi Bakteri Selulolitik sebagai Penghambat Pertumbuhan Cendawan pada Tanaman Kelapa Sawit. Penelitian ini didanai dari dana start up CRC 990 dan Hibah Desentralisasi Dikti (BOPTN) tahun 2013.
Terima kasih penulis ucapkan kepada Dr Nisa Rachmania Mubarik, MSi dan Dr Sri Listiyowati, MSi selaku pembimbing. Di samping itu, penghargaan penulis sampaikan kepada Dr Ir Raden Roro Dyah Perwitasari, MSc atas saran dan diskusi yang diberikan, Teman-teman seperjuangan (Agus, Lili, Dini, Routh, Ulfah), Bapak Jaka sebagai laboran Mikrobiologi, Bapak Suharyanto di Balai Bioteknologi Perkebunan Indonesia, Bogor, dan rekan kerja di laboratorium Mikrobiologi yang telah banyak membantu dan memberikan saran dalam proses penelitian. Ungkapan terima kasih juga disampaikan kepada Papa, Mama, Dede, serta seluruh keluarga, dan keluarga Biologi 46 atas segala doa dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL ix
DAFTAR GAMBAR ix
DAFTAR LAMPIRAN ix
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 1
METODE 2
Bahan 2
Isolasi Bakteri Selulolitik 2
Isolasi dan Identifikasi Cendawan yang diduga Patogen asal Kelapa Sawit 3
Identifikasi Bakteri Isolat Terpilih 3
Uji Antagonis 3
Pembuatan Kurva Tumbuh dan Uji Daya Hambat Bakteri terhadap Cendawan 3
HASIL 4
Isolasi Bakteri Selulolitik 4
Isolasi dan Identifikasi Cendawan yang Diduga Patogen asal Kelapa Sawit 4
Identifikasi Bakteri Isolat Terpilih 5
Uji Antagonis 6
Pembuatan Kurva Tumbuh dan Uji Daya Hambat Bakteri terhadap Cendawan 7
PEMBAHASAN 10
SIMPULAN 12
DAFTAR PUSTAKA 12
LAMPIRAN 14
DAFTAR TABEL
1 Indeks selulolitik (IS) dari 40 isolat bakteri asal tanah perkebunan sawit dan karet dari sekitar Hutan Taman Nasional Bukit Dua Belas,
Jambi 4
2 Zona hambat isolat bakteri terhadap cendawan Curvularia sp. dan
Colletotrichum sp. 6
DAFTAR GAMBAR
1 Morfologi koloni cendawan yang diduga patogen Curvularia sp. dan Colletotrichum sp. dari daun kelapa sawit asal Balai Penelitian
Bioteknologi Perkebunan Indonesia, Bogor 5
2 Morfologi spora cendawan yang diduga patogen Curvularia sp. dan Colletotrichum sp. dari daun kelapa sawit asal Balai Penelitian
Bioteknologi Perkebunan Indonesia, Bogor 5
3 Morfologi sel bakteri hasil pewarnaan Gram SAHN13.30 dan
KAHN15.39 6
4 Penghambatan cendawan Colletotrichum sp. oleh isolat bakteri SAHN13.30 dan KAHN15.39 serta penghambatan cendawan Curvularia sp. oleh isolat bakteri SAHN13.30 dan KAHN15. 39 7 5 Kurva pertumbuhan isolat SAHN13.30 dan daya hambat terhadap
cendawan Curvularia sp. dan Colletotrichum sp. selama 60 jam inkubasi dengan metode kertas cakram dan metode sumur 8 6 Kurva pertumbuhan isolat KAHN15.39 dan daya hambat terhadap
cendawan Curvularia sp. dan Colletotrichum sp. selama 60 jam inkubasi dengan metode kertas cakram dan sumur 8 7 Kurva pertumbuhan isolat SAHN13.30 dan daya hambat terhadap
cendawan Curvularia sp. dan Colletotrichum sp. selama 60 jam inkubasi dengan metode kertas cakram dan sumur 9 8 Kurva pertumbuhan isolat KAHN15.39 dan daya hambat terhadap
cendawan Curvularia sp. dan Colletotrichum sp. selama 60 jam inkubasi dengan metode kertas cakram dan sumur 10
DAFTAR LAMPIRAN
1 Sampel tanah asal perkebunan kelapa sawit dan perkebunan karet kawasan Hutan Taman Nasional Bukit Dua Belas, Jambi 14 2 Komposisi media pertumbuhan bakteri dan cendawan 15 3 Kurva standar isolat bakteri SAHN13.30 dan KAHN15.39 16 4 Hasil uji biokimia isolat SAHN13.30 dan KAHN15.39 dengan
PENDAHULUAN
Latar Belakang
Upaya peningkatan produksi kelapa sawit dilakukan secara ekstensifikasi, usaha perbaikan teknik budidaya, penggunaan klon unggul, dan usaha pengelolaan hama dan penyakit dilakukan secara terpadu. Peningkatan terlihat pada tahun 1967 produksi kelapa sawit sebesar 167.000 menjadi 18 juta pada tahun 2009 (Ditjenbun 2009). Penyakit yang menyebabkan kerugian pada perkebunan kelapa sawit di antaranya cendawan patogen, seperti Ganoderma boninense (Widiastuti et al. 2003) penyebab busuk pangkal batang yang menyebabkan kerugian terbesar pada produksi kelapa sawit, Colletotrichum acutatum sebagai penyebab penyakit antraknosa (Utomo 1987), dan Curvularia eragrostidis penyebab penyakit hawar daun (Aji et al. 2013).
Pengendalian penyakit yang diakibatkan cendawan dapat dilakukan dengan menggunakan fungisida, namun pemberian fungisida berlebihan dalam jangka waktu panjang dapat memberikan dampak negatif. Salah satu dinding sel cendawan tersusun atas kompleks selulosa sehingga penanggulangan cendawan penyebab penyakit pada kelapa sawit dapat dilakukan dengan introduksi bakteri penghasil enzim selulase sebagai biokontrol terhadap cendawan patogen. Degradasi sempurna selulase merupakan proses yang melibatkan beberapa aktivitas enzim, yaitu endo-β1.4-glukanase, ekso-β1.4-glukanase, dan β -glukosidase.
Indonesia merupakan salah satu negara yang memiliki keanekaragaman hayati yang tinggi, sehingga potensi mikrobnya sangat besar untuk terus dieksplorasi dan diteliti. Taman Nasional Bukit Dua Belas (TNBDB), Jambi menjadi salah satu daerah yang mendapati sorotan karena konversi lahannya. Selain itu, publikasi mengenai mikrob selulolitik di daerah tersebut belum ada, sehingga eksplorasi selulolitik di daerah tersebut sangat menarik untuk diteliti.
Tujuan Penelitian
Penelitian ini bertujuan mengisolasi dan menapis bakteri selulolitik dari tanah perkebunan kelapa sawit dan karet disekitar kawasan hutan TNBDB, Jambi serta menyeleksi isolat yang berpotensi menghambat pertumbuhan cendawan yang diduga patogen terhadap tanaman bibit kelapa sawit.
METODE
2
Bahan
Bahan-bahan yang digunakan adalahs ampel tanah yang diambil dari kawasan perkebunan (hutan transformasi) karet dan kelapa sawit di kawasan Hutan TNBDB Jambi (Lampiran 1) serta daun kelapa sawit yang terserang cendawan yang diduga patogen berasal dari Balai Penelitian Bioteknologi Perkebunan Indonesia, Bogor dengan ciri-ciri daun terdapat bercak coklat di sekitar pinggir daun pada bibit berumur 2-7 bulan.
Isolasi Bakteri Selulolitik
Sebanyak 2.5 gram tanah dimasukkan ke dalam 25 mL media nutrient broth (NB) yang mengandung carboxymethylcelullose (CMC) 1% (Lampiran 2) di dalam Erlenmeyer 100 mL. Suspensi lalu digoyang di atas mesin penggoyang dan diinkubasi selama 24 jam. Selanjutnya, suspensi diencerkan secara serial (10-5, 10-6, 10-7, 10-8 ) dan 0.1 ml dari masing-masing pengenceran disebar pada medium nutrient agar (NA) yang mengandung CMC 1% (Lampiran 2) dan diinkubasi selama 24 jam pada suhu ruang. Koloni bakteri yang menghasilkan zona bening dimurnikan dengan metode cawan gores (Hadioetomo 1993). Penegasan zona bening dilakukan dengan menitikkan masing-masing isolat murni pada medium NA dan diinkubasi selama 24 jam. Selanjutnya medium NA tersebut diwarnai dengan pewarna merah kongo 1% selama 15 detik dan dibilas dengan NaCl 0.85% sebanyak tiga kali.Indeks Selulolitik (IS) dihitung dengan menggunakan rumus: IS = Ø zona bening – Ø koloni
Ø koloni
Isolasi dan Identifikasi Cendawan yang diduga Patogen asal Kelapa Sawit
Cendawan patogen diisolasi dari sampel daun kelapa sawit yang diperoleh dari Balai Penelitian Bioteknologi Perkebunan Indonesia, Bogor. Sampel daun dipotong ± 25 mm dan dicuci dengan air mengalir. Selanjutnya sampel daun direndam natrium hipoklorit 1% selama 1 menit, dibilas air steril dan dikeringkan dengan tisu steril, serta ditumbuhkan pada medium potato dextrose agar (PDA) yang mengandung kloramfenikol 0.5% (Lampiran 2), dan diamati hingga terdapat pertumbuhan miselium. Miselium yang tumbuh dipindahkan ke medium PDA
(Lampiran 2) sampai mendapat biakan murni pada medium PDA yang
mengandung kloramfenikol 0.5%. Identifikasi cendawan diawali dengan membuat
preparat dari biakan murni hasil peremajaan. Preparat diamati dengan menggunakan mikroskop pada perbesaran 400x. Identifikasi cendawan berdasarkan Barnett dan Hunter (1987).
Identifikasi Bakteri Isolat Terpilih
200 µL medium kit API 50 CHB. Sedangkan tabung pertama diisi dengan 200 µL medium kit API 50 CHB tanpa bakteri (kontrol). Hasil diinterpretasi dengan menggunakan perangkat lunak API web dengan database untuk diidentifikasi (Lampiran 4).
Uji Antagonis
Uji antagonis dilakukan dengan metode Fokkema (1973) yaitu memasangkan inokulum cendawan (± 20 mm) dengan goresan bakteri pada jarak 3 cm pada medium PDA. Pasangan kultur tersebut diinkubasi selama 6 hari kemudian dihitung zona hambat bakteri terhadap cendawan dibandingkan dengan kontrol. Kontrol menggunakan cendawan dan air destilata tanpa bakteri. Persentase penghambatan antara bakteri terhadap cendawan yang diduga patogen dapat dihitung dengan mengukur jari-jari cendawan ke arah tepi cawan (R1), dan mengukur jari-jari cendawan ke arah bakteri (R2). Persentase Penghambatan dihitung menggunakan rumus = (R1 – R2) x R1-1x 100%.
Pembuatan Kurva Tumbuh dan Uji Daya Hambat Bakteri terhadap Cendawan
4
HASIL
Isolasi Bakteri Selulolitik
Bakteri selulolitik yang berhasil diisolasi dari 10 sampel tanah hutan transformasi sebanyak 20 isolat dari sampel tanah perkebunan kelapa sawit dan 20 isolat dari sampel tanah perkebunan karet dengan nilai indeks selulolitik (IS) terendah 0.03 dan tertinggi 16.6. Sebanyak 12 isolat bakteri memiliki nilai IS > 6 (Tabel 1).
Tabel 1 Indeks selulolitik (IS) dari 40 isolat bakteri asal tanah perkebunan kelapa sawit dan karet dari sekitar Hutan TNBDB, Jambi
A.Kelapa sawit B. Karet
Isolasi dan Identifikasi Cendawan yang Diduga Patogen asal Kelapa Sawit
Kelapa sawit yang diduga terserang cendawan diambil dari Balai Penelitian Bioteknologi Perkebunan Indonesia, Bogor. Sebanyak dua jenis cendawan patogen berhasil diisolasi dari daun kelapa sawit Balai Penelitian Bioteknologi Perkebunan Indonesia, Bogor yaitu Curvularia sp. (Gambar 1a) dan Colletotrichum sp. (Gambar 1b).Curvularia sp. memiliki karakteristik morfologi meliputi warna koloni awal putih, kemudian menjadi abu-abu, merah muda, dan setelah dewasa berwarna cokelat atau hitam (Gambar 1a), dan konidiofor berwarna cokelat yang sederhana, serta konidium yang membesar pada bagian tengahnya (Gambar 2a) (Barnett dan Hunter 1987; Larone 1995). Colletotrichum sp. memiliki karakteristik morfologi meliputi warna koloni oranye tua, konidiofor
No. Kode Isolat Gram Indeks
sederhana yang memanjang, konidia hialin, 1 sel, berbentuk ovoid (berbentuk telur dengan satu ujungnya menyempit) atau oblong (Barnett dan Hunter 1987).
Gambar 1 Morfologi koloni cendawan yang diduga patogen (a) Curvularia sp. dan (b)
Colletotrichum sp. dari daun kelapa sawit asal Balai Penelitian Bioteknologi Perkebunan Indonesia, Bogor
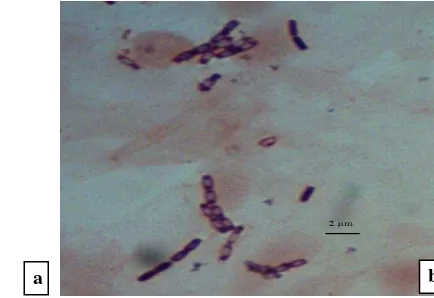
Gambar 2 Morfologi spora cendawan yang diduga patogen (a) Curvularia sp. dan (b)
Colletotrichum sp. dari daun kelapa sawit asal Balai Penelitian Bioteknologi Perkebunan Indonesia, Bogor
Identifikasi Bakteri Isolat Terpilih
Identifikasi bakteri dilakukan berdasarkan uji biokimia (fisiologi) menggunakan kit API 50 CHB untuk isolat bakteri SAHN13.30dan KAHN15.39. Isolat SAHN13.30 merupakan bakteri Bacillus cereus dengan tingkat kemiripan 88% dengan kit API 50 CHB yang menunjukkan hasil yang positif terhadap uji karbohidrat yang ditandai mampu menggunakan D-xilosa, mannosa, amygdalin, salisin, laktosa, trehalosa, dan xylitol (Lampiran 4). Isolat KAHN15.39 merupakan bakteri Bacillus thuringiensis yang menunjukkan hasil yang baik untuk identifikasi menggunakan kit API 50 CHB, positif terhadap uji karbohidrat yang ditandai mampu menggunakan D-xilosa, fruktosa, mannosa, sorbosa, amygdalin, salisin, selobiosa, maltosa, laktosa, inulin, glikogen dan xylitol (Lampiran 4). Hasil pewarnaan Gram merupakan Gram positif baik isolat SAHN13.30 dan KAHN15. (Gambar 3).
a b
6
Gambar 3 Morfologi sel bakteri hasil pewarnaan Gram (a) SAHN13.30 dan (b) KAHN15.39
Uji Antagonis
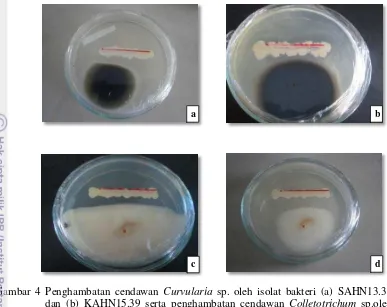
Sebanyak dua isolat dari dua belas isolat terpilih yaitu SAHN13.30 dan KAHN15.39 (Tabel 2) memiliki IS > 6 dan memiliki kemampuan dalam menghambat cendawan Curvularia sp. serta Colletotrichum sp. melalui metode Fokkema. Daya hambat isolat SAHN13.30 dan KAHN15.39 terhadap Curvularia sp. sebesar 57.5%, sedangkan terhadap Colletotrichum sp. sebesar 60% (Gambar 4). Kontrol dengan perlakuan akuades untuk kedua cendawan patogen menunjukkan tidak ada aktivitas penghambatan karena masing-masing cendawan dapat tumbuh maksimum memenuhi cawan petri.
Gambar 4 Penghambatan cendawan Curvularia sp. oleh isolat bakteri (a) SAHN13.30 dan (b) KAHN15.39 serta penghambatan cendawan Colletotrichum sp.oleh isolat bakteri (c) SAHN13.30 dan (d) KAHN15.39
Pembuatan Kurva Tumbuh danUji Daya Hambat Bakteri terhadap Cendawan
Kurva pertumbuhan bakteri digunakan untuk mengetahui waktu optimum dalam kemampuan menghambat pertumbuhan cendawan, dan kemampuan enzim selulase ekstrak kasarnya pada waktu optimum tersebut. Kurva pertumbuhan menunjukkan bahwa fase eksponensial berakhir pada jam ke-12 dengan jumlah bakteri berkisar 3.995 x 10-8 sel/mL, setelah itu diikuti fase stasioner hingga jam ke-60 dengan jumlah bakteri berkisar antara 3.995 x 10-8sel/mL sampai 4.115 x 10-8sel/mL untuk isolat SAHN13.30. Jumlah sel bakteri pada keadaan fase tersebut dengan nilai logaritmiknya 0.6. Sedangkan fase eksponensial KAHN15.39 berakhir pada jam ke-12 dengan jumlah bakteri berkisar 3.937 x 10-8 sel/mL dan memasuki fase stasioner hingga jam ke-60 dengan jumlah bakteri berkisar antara 3.937 x 10-8sel/mL sampai 4.767 x 10-8sel/mL dengan nilai logaritmiknya 0.9.
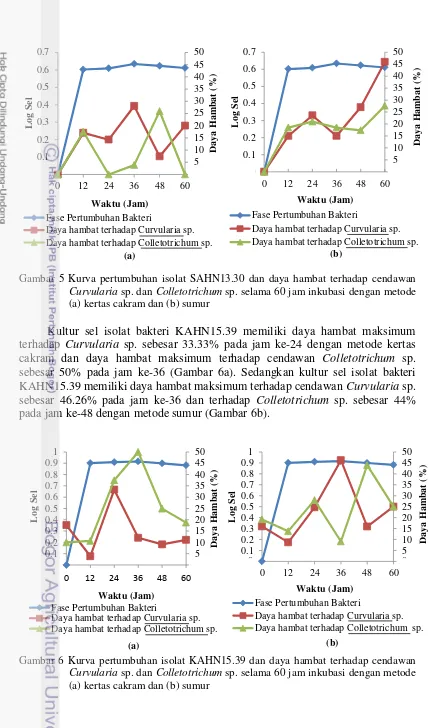
Kultur sel isolat bakteri SAHN13.30 memiliki daya hambat maksimum terhadap cendawan Curvularia sp. sebesar 28.06% pada jam ke-36 dan terhadap Colletotrichum sp. sebesar 25.92% pada jam ke-48 dengan metode kertas cakram (Gambar 5a). Pada metode sumur kultur sel isolat bakteri SAHN13.30 memiliki daya hambat maksimum pada jam ke-60, baik terhadap Curvularia sp. maupun Colletotrichum sp. yaitu masing-masing sebesar 45.95% dan 27.58% dengan metode sumur (Gambar 5b).
a
c d
8
Gambar 5 Kurva pertumbuhan isolat SAHN13.30 dan daya hambat terhadap cendawan
Curvularia sp. dan Colletotrichum sp. selama 60 jam inkubasi dengan metode (a) kertas cakram dan (b) sumur
Kultur sel isolat bakteri KAHN15.39 memiliki daya hambat maksimum terhadap Curvularia sp. sebesar 33.33% pada jam ke-24 dengan metode kertas cakram dan daya hambat maksimum terhadap cendawan Colletotrichum sp. sebesar 50% pada jam ke-36 (Gambar 6a). Sedangkan kultur sel isolat bakteri KAHN15.39 memiliki daya hambat maksimum terhadap cendawan Curvularia sp. sebesar 46.26% pada jam ke-36 dan terhadap Colletotrichum sp. sebesar 44% pada jam ke-48 dengan metode sumur (Gambar 6b).
Gambar 6 Kurva pertumbuhan isolat KAHN15.39 dan daya hambat terhadap cendawan
Curvularia sp. dan Colletotrichum sp. selama 60 jam inkubasi dengan metode (a) kertas cakram dan (b) sumur
(b) Daya hambat terhadap Colletotrichum sp.
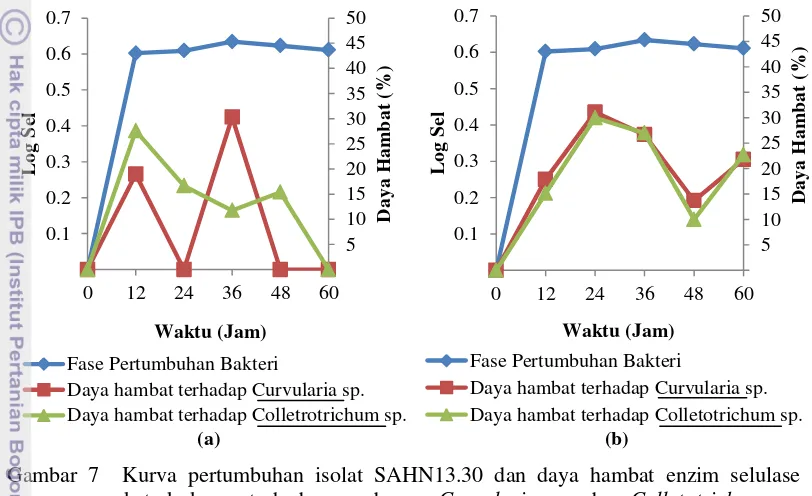
Enzim selulase ekstrak kasar isolat bakteri SAHN13.30 memiliki daya hambat maksimum terhadap cendawan Curvularia sp. sebesar 30.3% pada jam ke-36 dan terhadap Colletotrichum sp. sebesar 27.58% pada jam ke-24 dengan metode kertas cakram (Gambar 7a). Enzim selulase ekstrak kasar isolat bakteri SAHN13.30 memiliki daya hambat maksimum dengan nilai 31.08% dan 30% terhadap cendawan Curvularia sp. dan Colletotrichum sp. pada jam ke-24 dengan metode sumur (Gambar 7b).
Gambar 7 Kurva pertumbuhan isolat SAHN13.30 dan daya hambat enzim selulase ekstrak kasar terhadap cendawan Curvularia sp. dan Colletotrichum sp. selama 60 jam inkubasi dengan metode (a) kertas cakram dan (b) sumur
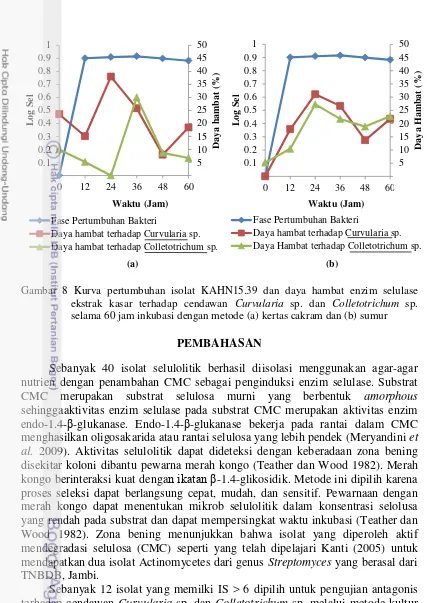
Enzim selulase ekstrak kasar isolat KAHN15.39 memiliki daya hambat maksimum terhadap cendawan Curvularia sp. sebesar 38% pada jam ke-24 dan terhadap Colletotrichum sp. sebesar 30% pada jam ke-36 dengan metode kertas cakram(Gambar 8a). Enzim selulase KAHN15.39 memiliki daya hambat terhadap cendawan Curvularia sp. sebesar 31.08% dan terhadap Colletotrichum sp. sebesar 27.27% pada jam ke-24 dengan metode sumur (Gambar 8b).
10
Gambar 8 Kurva pertumbuhan isolat KAHN15.39 dan daya hambat enzim selulase ekstrak kasar terhadap cendawan Curvularia sp. dan Colletotrichum sp. selama 60 jam inkubasi dengan metode (a) kertas cakram dan (b) sumur
PEMBAHASAN
Sebanyak 40 isolat selulolitik berhasil diisolasi menggunakan agar-agar nutrien dengan penambahan CMC sebagai penginduksi enzim selulase. Substrat CMC merupakan substrat selulosa murni yang berbentuk amorphous sehinggaaktivitas enzim selulase pada substrat CMC merupakan aktivitas enzim endo-1.4-β-glukanase. Endo-1.4-β-glukanase bekerja pada rantai dalam CMC menghasilkan oligosakarida atau rantai selulosa yang lebih pendek (Meryandini et al. 2009). Aktivitas selulolitik dapat dideteksi dengan keberadaan zona bening disekitar koloni dibantu pewarna merah kongo (Teather dan Wood 1982). Merah kongo berinteraksi kuat dengan ikatan β-1.4-glikosidik. Metode ini dipilih karena proses seleksi dapat berlangsung cepat, mudah, dan sensitif. Pewarnaan dengan merah kongo dapat menentukan mikrob selulolitik dalam konsentrasi selolusa yang rendah pada substrat dan dapat mempersingkat waktu inkubasi (Teather dan Wood 1982). Zona bening menunjukkan bahwa isolat yang diperoleh aktif mendegradasi selulosa (CMC) seperti yang telah dipelajari Kanti (2005) untuk mendapatkan dua isolat Actinomycetes dari genus Streptomyces yang berasal dari TNBDB, Jambi.
Sebanyak 12 isolat yang memilki IS > 6 dipilih untuk pengujian antagonis terhadap cendawan Curvularia sp. dan Colletotrichum sp. melalui metode kultur berpasangan (Fokkema 1973). Pengujian antagonis isolat bakteri terhadap kedua cendawan merupakan pengujian awal untuk mendapatkan isolat yang memiliki kemampuan dalam menghambat pertumbuhan cendawan yang diduga patogen. Hasil uji menunjukkan hanya 2 isolat (SAHN13.30 dan KAHN15.39) yang menunjukkan aktivitas penghambatan terhadap kedua cendawan yang diduga patogen. Kedua isolat masing-masing memiliki kemampuan menghambat
cendawan Curvularia sp. sebesar 57.5% dan terhadap Colletotrichum sp. sebesar 60%.
Pada penelitian ini fase adaptasi bakteri tidak tampak pada kurva petumbuhan karena interval waktu pengamatan terlalu besar yaitu pada setiap 12 jam. Tampak fase eksponensial berakhir pada jam ke-12, setelah itu memasuki fase stasioner tampak hingga jam ke-60. Pada fase stasioner, sel yang membelah sama jumlahnya dengan sel yang mati. Pada fase ini keadaan sel menjadi tahan terhadap kondisi ekstrim.
Uji antagonis kultur sel bakteri dan enzim selulase ekstrak kasar terhadap kedua cendawan uji yang dilakukan bersama dengan kurva pertumbuhan membuktikan, bahwa baik kultur sel bakteri maupun enzim selulase ekstrak kasar berperan dalam mekanisme penghambatan Curvularia sp. dan Colletotrichum sp. Berdasarkan kurva pertumbuhan diketahui bahwa SAHN13.30 memiliki kemampuan dalam menghambat cendawan yang diduga patogen yaitu Curvularia sp. pada fase stasioner dengan metode menggunakan kultur sel bakteri, sedangkan KAHN15.39 memiliki kemampuan menghambat cendawan yang diduga patogen yaitu Colletotrichum sp. pada fase stasioner dengan enzim selulase ekstrak kasar. Berbagai jenis selulase diperlukan dalam mendegradasi sempurna struktur selulosa yang kompleks (Yang et al. 2006). Aktivitas enzim selulase akan mengalami penurunan yang dapat disebabkan diantaranya oleh jumlah substrat yang semakin berkurang, feedback inhibition, dan keberadaan aktivitas proteolisis (White 1995).
Perbedaaan persentase penghambatan dengan menggunakan sel bakteri secara langsung maupun melalui metode kertas cakram dan metode sumur disebabkan antara lain oleh jenis dan jumlah senyawa antimikrob, konsentrasi dan kualitas dari senyawa antimikrob yang dihasilkan (Hwang et al. 2001), dan mekanisme penghambatan yang berbeda terhadap cendawan yang diduga patogen. Keberadaan dua puncak aktivitas enzim pada kedua isolat diduga terdapat jenis enzim yang berbeda. Menurut Ahlgren et al. (1967), dua puncak aktivitas enzim dapat menunjukkan keberadaan dua jenis enzim yang berbeda. Variasi perbedaan penghambatan cendawan dengan bakteri selulolitik dapat disebabkan oleh kespesifikan spesies, perbedaaan aktivitas selulase bakteri, komposisi selulosa dari dinding sel cendawan, dan keberadaan metabolit anti cendawan (Novitasari 2013). Komponen utama penyusun dinding sel cendawan uji pada penelitian ini adalah kitin. Dinding sel cendawan tidak hanya tersusun atas kitin, tetapi juga terdiri atas selulosa, dan kitinase kemungkinan merupakan enzim yang bertanggung jawab dalam degradasi dinding sel cendawan (Gohel et al. 2006; Anand dan Reddy 2009).
12
Bakteri selulolitik sebagai biokontrol alami diharapkan mampu dalam menghambat pertumbuhan cendawan yang diduga patogen. Colletotrichum sp. penyebab penyakit antraknosa yang menyerang kelapa sawit pada stadium bibit 2-4 bulan (Utomo 1987) dan dilaporkan penyebab penyakit antraknosa pada tanaman cabai (Wilia 2010). Stadium bibit umur tiga bulan merupakan umur bibit yang paling berpotensi terhadap bercak Curvularia sp., sedangkan stadium bibit empat bulan diduga sudah lebih tahan terhadap penularan bercak Curvularia sp. (Solehudin et al. 2012; Aji et al. 2013).
SIMPULAN
Sebanyak 40 isolat bakteri selulolitik berhasil diisolasi dari sampel tanah perkebunan kelapa sawit dan karet di sekitar kawasan Hutan Taman Nasional Bukit Dua Belas Jambi. Isolat bakteri yang memiliki indeks selulolitik (IS) tertinggi sebanyak 12, dengan nilai IS > 6. Isolat SAHN13.30 dan KAHN15.39 menunjukkan kemampuan menghambat cendawan Curvularia sp. dan Colletotrichum sp. Identifikasi SAHN13.30 dan KAHN15.39 dengan kit API 50 CHB menunjukkan bahwa isolat tersebut masing-masing termasuk Bacillus cereus dan Bacillus thuringiensis. Enzim selulase ekstrak kasar dan kultur sel SAHN13.30 dan KAHN15.39 terbukti berpotensi menghambat pertumbuhan cendawan Curvularia sp. dan Colletotrichum sp.
DAFTAR PUSTAKA
[Ditjenbun] Direktorat Jenderal Perkebunan. 2009. Statistik Perkebunan Indonesia. Jakarta (ID): Ditjenbun.
Ahlgren E. Eriksson K, Vesterberg O. 1967. Characterization of cellulases and related enzymes by isoelectric focusing, gel filtration and zone electrophoresis. Acta Chem Scand. 21(4):937-944.
Aji AF, Munajat Q, Pratama AP, Kalamullah H, Aprinaldi, Setiyawan J, Arymurthy AM. 2013. Detection of palm oil leaf disease with image processing and neural network classification on mobile device. Int J Comp Theory Eng. 5(3):1-5.
Anand S, Reddy J. 2009. Biocontrol potential of Trichoderma sp. against plant pathogens. Int J Agric Sci. 1(2):30-39.
Barnett HL, Hunter BB. 1987. Illustratred Genera of Imperfecti Fungi, Fourth Edition. New York (US): Macmilan Publishing Company.
Fokkema NJ. 1973. The role of saprophytic fungi in antagonism against Drechslera sorokiniana (Helminthosporium sativum) on agar plates and on rye leaves with pollen. Phys Plants Pathol. 3(1):195-205.
Gohel V, Singh A, Vimal M, Ashwini P, Chhatpar HS. 2006. Bioprospectiong and antifungal potential of chitinolytic microorganisms [ulas balik]. Afr J Biotechnol. 5(2):54-72.
Hadioetomo RS. 1993. Mikrobiologi Dasar Dalam Praktek. Jakarta (ID): PT Gramedia Pustaka Umum.
Hwang BK, Lee JY, Kim BS, Lim SW, Moon SS. 2001. Isolation and in vivo and in vitro antifungal activity of phenylacetic acid and sodium phenylacetate from Streptomyces humidus. J Appl Environ Microbiol. 67(1):3739-3745. Kanti A. 2005. Actinomycetes selulolitik dari tanah Hutan Taman Nasional Bukit
Dua Belas, Jambi. J Biodiversitas. 2(6):85-89.
Kucuk C, Kivanc M. 2004. In vitro antifungal activity of strain of Trichoderma harzianum. J Biol. 25(1):111-115.
Kloepper JW, Ryu CM, Zhang S. 2004. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathol. 94(1):1259-1266. Kondoh M, Hirai M, Shoda M. 2001. Integrated biological and chemical control
of damping-off caused by Rhizoctonia solani using Bacillus subtilis RB14-C and flutolanil. J Biosci Bioeng. 91(2):173-177.
Larone DH. 1995. Medically Important Fungi-A Guide to Identification. Ed ke-3. Washinton DC (US): ASM Pr.
Meryandini A, Widosari W, Maranatha B, Sumarti TC, Rachmania N, Satria H. 2009. Isolasi bakteri selulolitik dan karakterisasi enzimnya. J Makara Sains. 1(13):33-38.
Novitasari P. 2013. Isolasi dan identifikasi bakteri kitinolitik penghambat pertumbuhan cendawan patogen asal kokon Cricula trifenestrata [skripsi]. Bogor (ID): IPB Pr.
Solehudin D, Suswanti I, Supriyanto. 2012. Status penyakit bercak coklat pada pembibitan kelapa sawit di kabupaten Sanggau. J Perkeb Lahan Trop. 2(1):1-6.
Teather RM, Wood PJ. 1982. Use of congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl Environ Microbiol. 43(4):777-780.
Utomo C. 1987. Leaf diseases of oil palm nurseries in north Sumatra. J Bul Perkeb. 18(2):83-88.
White D. 1995.The Physiology and Biochemistry of Prokaryotes. New York(US): Oxford University Press.
Widiastuti H, Guhardja E, Sukarno N, Darusman LK, Goenadi DH, Smith S. 2003. Arsitektur akar kelapa sawit yang diinkulasi beberapa cendawan mikoriza arbuskula. Menara Perkeb 71:28-43.
Wilia W. 2010. Potensi cendawan endofit dan khamir untuk mengendalikan penyakit antraknosa (Colletotrichum acutatum L.) pada tanaman cabai [tesis]. Bogor (ID): IPB Pr.
14
Lampiran 1 Sampel tanah asal perkebunan kelapa sawit dan perkebunan karet kawasan Hutan Taman Nasional Bukit Dua belas, Jambi
Plot CRC Plot No. Sub plot Kode Tanaman
1 B03 A1 - 1259 S
B03 A2 FG01 1249 S
B03 A3 HI01 1250 S
B03 N1 DE45 1261 S
B03 N2 HI23 1253 S
B03 N3 AA01 1246 S
B03 N4 HI23 1307 S
2 B03 A4 JJ45 - S
B03 A5 JJ67 - S
B02 A6 B02 1310 S
B03 N5 JJ89 1311 S
BO2 N6 JJ67 1306 S
B02 N7 FG07 1305 S 3 BR2 A7 BC67 2558 K
BR2 A8 DE67 2576 K
BR2 A9 BR26 2618 K
BR2 A10 FG45 2608 K
BR2 A11 FG67 2610 K
BR2 N8 AA45 2536 K
BR2 N9 BC45 2551 K
BR2 N10 DE45 2578 K
BR2 N11 HI23 2643 K
BR2 N12 HI01 2645 K
4 BR4 A12 BC23 637 K
BR4 A13 BO43 660 K
BR4 N13 BC67 650 K
BR4 N14 FG23 636 K
5 BR4 A14 DEB9 886 K
BR4 A15 BC89 83 K
BR4 A16 FG10 903 K
BR4 N15 BR48 843 K
BR4 N16 FG45 840 K
Lampiran 2 Komposisi media pertumbuhan bakteri dan cendawan Media potato dextrose agar (PDA)
Komposisi Jumlah
Potato dextrose agar 39 g
Akuades 1000 mL
Media nutrient broth (NB) + CMC 1%
Komposisi Jumlah
Nutrient broth 13 g
Carboxymethil cellulose (CMC) 10 g
Akuades 1000 mL
Media nutrient agar (NA) + CMC
Komposisi Jumlah
Nutrient agar 23 g
Carboxymethil cellulose (CMC) 10 g
Akuades 1000 mL
Media potato dextrose agar (PDA) + kloramfenikol 0,5%
Komposisi Jumlah
Potato dextrose agar 39 g
Kloramfenikol 500mg
16
Lampiran 3 Kurva standar isolat bakteri SAHN13.30 dan KAHN15.39 Kurva standar isolat bakteri SAHN13.30
Kurva standar isolat bakteri KAHN15.39
Pengenceran OD Jumlah Sel Pengenceran OD Jumlah Sel (108)
Lampiran 4 Hasil uji biokimia isolat SAHN13.30 dan KAHN15.39 dengan menggunakan kit API 50 CHB
18
RIWAYAT HIDUP
Penulis dilahirkan di Kuningan pada tanggal 20 Februari 1991 sebagai anak pertama dari dua bersaudara dari ayah Arifin dan ibu Nanan Herawati. Penulis menyelesaikan pendidikan di SMA PGRI 109 Kota Tangerang pada tahun 2009 dan pada tahun yang sama diterima di Institut Pertanian Bogor melalui jalur Undangan Seleksi Masuk IPB (USMI). Penulis memilih mayor Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam.
Selama mengikuti perkuliahan, penulis menjadi asisten mata kuliah Biologi Dasar dan Mikrobiologi Dasar pada tahun ajaran 2012/2013. Selama kuliah S1, penulis mendapatkan beasiswa pendidikan yaitu beasiswa BBM tahun 2010-2013. Selain itu, penulis juga aktif di lembaga kemahasiswaan Cybertron Asrama TPB-IPB periode 2009/2010 sebagai pengurus, Himpunan Mahasiswa Biologi (Himabio) sebagai sekretaris II Badan Pengurus Harian (BPH) periode 2010/2011, serta aktif di berbagai kegiatan kepanitiaan BIONIC, Masa Perkenalan Departemen, SPIRIT, dan lain-lain; serta anggota Ikatan Himpunan Mahasiswa Biologi Indonesia Wilayah Jawa 1 (Ikahimbi) periode 2010/2011. Penulis melaksanakan studi lapang pada tahun 2011 dengan judul “Bakteri Filosfer pada Beberapa Tanaman asal Gunung Walat yang Berpotensi Penghasil Senyawa Bioaktif di Hutan Pendidikan Gunung Walat” dan praktik lapangan pada tahun
2012 dengan judul “Pengujian Bakteri Organisme Pengganggu Tumbuhan
Karantina pada Padi asal China di Laboratorium Bakteriologi dan Virologi Balai Karantina Tumbuhan Soekarno Hatta”.