EFEKTIVITAS PEMBERIAN ESTRADIOL DAN
PROGESTERON PADA KINERJA REPRODUKSI TIKUS
BUNTING
OLEH:
ARYAN1 SlSMlN SATYANINGTIJAS
PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRAK
ARYAN1
SISMM SATYANINGTIJAS. Efektivitas Pemberian Estradiol dan
Progesteron pada Kineja Reproduksi Tikus Bunting. Di bawah bimbingan TONNY
UNGERER
(Alm)
sebagai
ketua
komisi,
WASMEN
MANALU,
D.T.H.SIHOMBING, S.HAMDAN1 NASUTION dan SRIHADI AGUNGPRIYONO
sebagai anggota.
Dua percobaan pada tikus bunting dilakukan untuk mengetahui apakah pernberian
estradiol dan progesteron dapat meningkatkan penampilan reproduksi.
Pada
percobaan pertama,
378
ekor tikus disuntik secara
subcutan
dengan estradiol
(0,
1,06775, 2,1355
pg/g BB) dan progesteron (0,
4,281, 8,562
qg/g BB) pada hari ke-0,
ABSTRACT
ARYAN1
SISMIN SATYANINGTIJAS. Effects of Estradiol and Progesterone
Administration on Reproductive Performance in Pregnant Rats. Under The Direction
of TONNY UNGERER, WASMEN MANALU, D.T.H.SIHOMBING, S.HAMDANI
NASUTION and SRIHADI AGUNGPRIYONO.
SURAT PERNYATAAN
Dengan
ini
saya menyatakan bahwa disertasi yang berjudul
:E F E K T M T A S PEMBEIUAN ESTRADIOL DAN PROGESTERON PADA
KINERJA REPRODUKSI TIKUS
BUNTING
Adalah benar merupakan hasil karya saya sendiri dan belum pernah dipublikasikan.
Semua sumber data dan informasi yang digunakan telah dinyatakan secara jelas dan
dapat diperiksa kebenarannya.
Bogor, Agustus
2001
EFEKTIVITAS PEMBERIAN ESTRADIOL DAN
PROGESTERON PADA KINERJA REPRODUKSI
TIKUS BUNTING
OLEH:
ARYAN1 SISMIN SATYANINGTIJAS
Disertasi
sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Biologi
PROGRAM PASCA SARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi
:Efektivitas Pemberian Estradiol dan Progesteron
Pada Kinerja Reproduksi Tikus Bunting
Nama
: Ary ani Sismin Saty aningtijasNomor Pokok
:BIO. 95574
Program Studi
:Biologi
Sub Program Studi
:Fisiologi
Meny etujui :
Komisi Pembimbing
Prof. Tonny Ungerer, Ph.D. (Alm)
Ketua
2.
Anggop Komisi
..
Ir. Wasmen Manalu,
Ph.U.Drh. Hamdani Nasution, Ph.D.
Anggota
Anggot a
Prof. D.T.H. Sihombing, P ~ . D .
Drh. Srihadi Appngpriyono, Ph.D.
Anggota
Anggot a
3.
Ketua Program
Dr. Dede Setiadi
Studi Biologi
Br
am Pascasarjana
L.
--
-2RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 14 September 1960 di Bondowoso, Jawa Timur
dari keluarga bpk. Siswojo (alm.) dan ibu Soedarminah (almh.) sebagai anak keempat
dari tujuh bersaudara. Pada tahun 1986 penulis menikah dengan Dadang Sudjana, SE.
dan dikaruniai dua orang anak: Regi Ryanda dan Riza Ryanda.
Pendidikan sekolah dasar di SDK Maria Fatima I, sekolah menengah pertama di
SMPN I dan sekolah menengah atas di SMAN I diselesaikan di Jember, Jawa Timur.
Penulis diterima di EPB pada tahun 1979 melalui Proyek Perintis
I1
dan masuk sebagai
mahasiswa FKH sebagai angkatan 16 (angkatan Gelatin). Penulis menerima gelar
Sarjana Kedokteran Hewan (Dra.Med Vet.) pada tahun 1983 dan menerima gelar
profesi Dokter Hewan (drh.) pada tahun 1984. Pada tahun 1991 penulis mendapatkan
gelar Master of Science (M.Sc.) dari University of Missouri Columbia, USA.
KATA
PENGANTARSyukur Alhamdulillah penulis panjatkan kepada Allah S.W.T. atas rahrnat dan karunia-Nya sehingga penulisan disertasi ini dapat terselesaikan.
Berkat dorongan semangat, bimbingan dan arahan para pembimbing, penulis akhirnya dapat menyelesaikan penelitian dan penulisan disertasi ini, untuk itu penulis mengucapkan banyak terima kasih kepada almarhum Prof. drh. Tonny Ungerer Ph.D. yang telah mendahului kita menghadap Yang Kuasa dan tidak mempunyai kesempatan untuk menguji penulis, Ir. Wasmen ManaIu Ph.D. yang telah banyak memberikan pemikiran dan arahan penelitian, Prof drh. D.T.H. Sihombing Ph.D., drh. S. Hamdani Nasution Ph.D., dan drh. Srihadi Agungpriyono Ph.D. yang membimbing penulis dalam analisis data dan penulisan.
Ucapan terima kasih juga penulis ucapkan kepada Fakultas Kedokteran Hewan khususnya Bagian Fisiologi dan Farmakologi, Program Pascasarjana Institut Pertanian Bogor dan Proyek Tim Manajemen Program Doktor (TMPD) yang telah memberikan kesempatan kepada penulis untuk mendapatkan jenjang pendidikan yang lebih tinggi.
Tanpa dorongan dan semangat dari teman-teman dan sahabat di Bagian Fisiologi dan Farmakologi, FKH-IPS (dik Rini, Hera, Dewi, Koekoeh, Agik, Rita, Isdoni, mbak Ietje, mas Bambang dan semuanya) mungkin disertasi ini tak akan terwujud. Kepada pak Edi, pak Pairin, dan mbak Asmarida terima kasih atas bantuannya. Terima kasih yang tak terhingga penulis ucapkan kepada teman sepejuangan yang teIah banyak membantu terselesainya penelitian dan disertasi ini, Ir. Hernawati M.S dan kepada mahasiswa bimbingan saya, Rohani Samosir. Teman-teman di laboratorium Anatomi dan Histologi, Guris, pak Yoni, mas Adi, dik Tutek, Novie, Dini, Esthi dan pak Maman terima kasih atas segala perhatian dan bantuannya. Semoga segala kebaikannya mendapatkan balasan dari Yang Esa. Tak lupa untuk Cecep, abinya Eldin, dan pak Dede dari HPT-Faperta, terima kasih untuk bantuannya.
Penulis juga tak akan melupakan segala bantuan, perhatian dan dorongan moril maupun materiil yang diberikan oleh sahabat tersayang Sri Utami Handayani ,
Darnayanti Buchori, F. Maulana, dan mbak Ning semoga amal dan kebaikannya akan mendapatkan pahala yang berlebih dari Allah SWT.
Terakhir penulis juga mengucapkan terima kasih kepada bapak dan ibu yang telah pergi menghadap Allah SWT atas doa-doa yang pernah berkumandang untuk penulis tanpa sempat melihat keberhasilan penulis saat ini. Untuk mama-papa 3 a n a Sudjana dan kakak-kakakku (mas Dodi, mas Toni, mas Rudi, Teh Ayu) serta adikku (Ita, Dandi, Edi, Iwan dan Agung) terima kasih atas bantuan dan doanya. Terima kasih yang tak terhingga juga penulis persembahkan untuk suarni tercinta, Dadang Sudjana dan anak-anakku (Regi dan Riza) atas pengertian dan kesabarannya dalam menghadapi sikap penulis selama dalam penyelesaian disertasi ini.
Akhirnya dengan segala kerendahan hati penulis hanya berharap semoga segala jerih payah penulis ini dapat bermanfaat bagi yang memerlukannya.
DAFTAR 1 SI
Halaman
DAFTAR TABEL ... ...
DAFTAR GAMB AR
DAFTAR LAMPIRAN ...
PENDAHULUAN ...
Tujuan ... ...
Manfaat dan Kegunaan Hasil PeneIitian
Hipotesis ... ...
TDJJAUAN PUSTAKA
Biologi Reproduksi Tikus Putih ... ... Hormon Reproduksi ... Estrogen ... Progesteron ...
Kine j a Reproduksi
Asam Nukleat ... ...
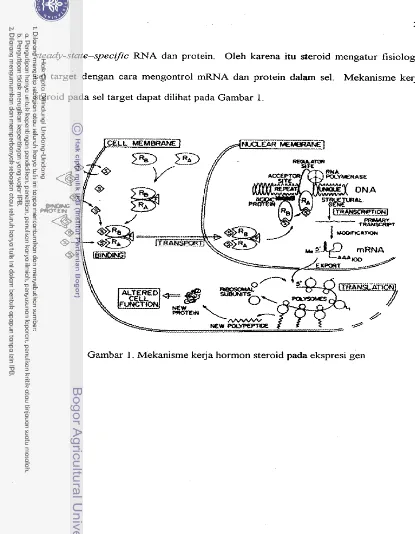
Mekanisme K e j a Hormon Steroid pada Ekspresi Gen
MATElU DAN METODE PENELITIAN ... ...
Rancangan Percobaan
...
Protokol Percobaan
Prosedur Pengambilan Sampel dan Pengukuran Sampel ...
Pembuatan Preparat Histologi dan Pewarnaan ... ...
Analisis Kimia Jaringan Uterus
. .
. .
...
Anal~sls S t a t ~ s t ~ k
HASIL DAN PEMBAHASAN ...
Pengaruh Pemberian Estradiol dan Progesteron pada Tikus pada Hari ke.0. 2. 4. 6. 8. 10. dan 12 Kebuntingan ...
Pengaruh Pemberian Estradiol dan Progesteron pada Tikus Bunting Selarna 5 dan 12 Hari kebuntingan ...
Pengaruh Pemberian Estradiol dan Progesteron Terhadap Pertumbuhan dan Perkembangan Serta Daya Tahan Hidup
...
Anak Sampai Usia Lepas Sapih
...
Pembahasan Umum
DAFTAR TABEL
1. Rataan jumlah korpus luteum, jumlah titik implantasi selisih, jumlah korpus luteum dan titik implantasi tikus selama hari kebuntingan 0 s/d 12 pada berbagai kombinasi
dosis estradiol dan progesteron.. ...
2. Rataan bobot uterus tikus selama hari kebuntingan 0 s/d 12
...
pada berbagai kornbinasi dosis estradiol dan progesteron
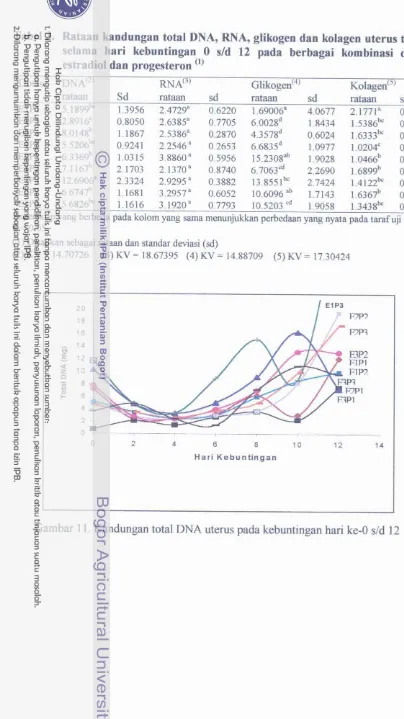
3 . Rataan kandungan total DNA, RNA, gEkogen dan kolagen uterus tikus selama hari kebuntingan 0 s/d 12 pada
berbagai kombinasi dosis estradiol dan progesteron ... 4. Rataan bobot uterus tikus pada penyuntikan sampai masa
...
implantasi (H-5) dan plasentasi (H-12)
5. Rataan kandungan total DNA, RNA, glikogen dan
kolagen uterus tikus bunting pada penyuntikan sampai masa
...
implantasi (H-5) dan plasentasi (H-12)
6 . Rataan jumlah korpus luteum, jumlah titik implantasi, selisih korpus luteum dan titik implantasi, jumlah anak, selisih jurnlah titik implantasi dan anak pada penyuntikan
sampai masa implantasi (H-5) dan plasentasi (H-12) ...
DAFTAR GAMBAR
H a l a m a n
27
3 1
3 3
3 3
Mekanisme kerja hormon steroid pada ekspresi gen ...
Kandang tikus ...
Bagian abdomen tikus ...
Uterus tikus ...
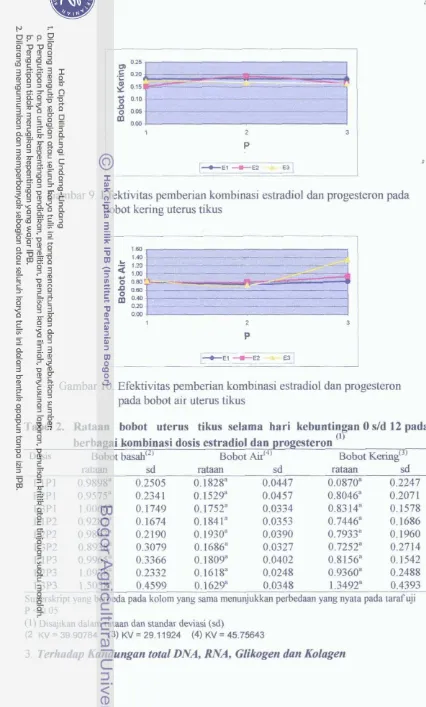
Efektivitas pemberian kombinasi estradiol dan
...
progesteron pada jumlah korpus luteum
Efektivitas pemberian kombinasi estradiol dan
...
progesteron pada jurnlah titik implantasi
Efektivitas pemberian kombinasi estradiol dan progesteron
...
pada selisih jumIah korpus luteum dan titik irnplantasi
Efektivitas pemberian kombinasi estradiol dan
...
progesteron pada bobot basah uterus tikus
Efektivitas pemberian kombinasi estradiol dan
progesteron pada bobot kering uterus tikus ...
Efektivitas pemberian kombinasi estradiol dan
progesteron pada bobot air uterus tikus ...
Kandungan total DNA uterus pada kebuntingan hari 0 d d 12.
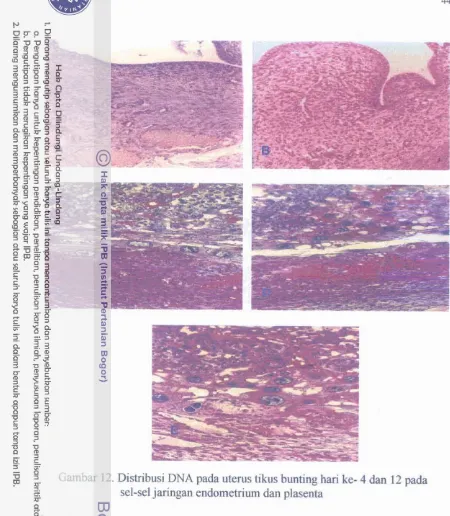
Distribusi DNA pada uterus tikus bunting hari ke 4 dan 12 pada sel-sel jaringan endometrium dan plasenta dengan pewarnaan hemaktoksilin-eosin.. ...
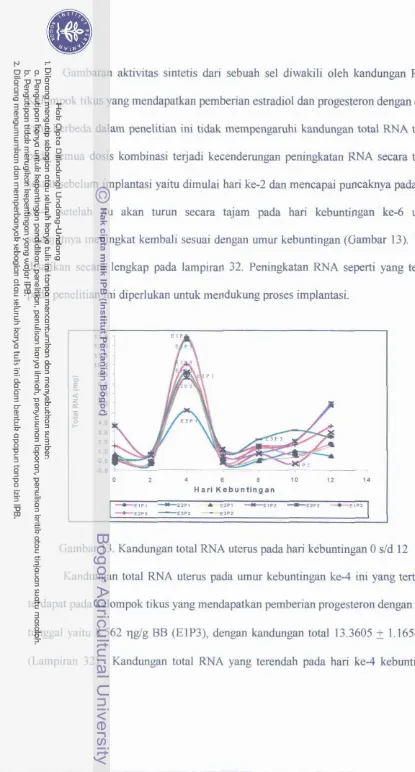
Kandungan total RNA uterus pada hari kebuntingan 0 d d 12 ....
Distribusi RNA pada uterus tikus bunting hari ke-12 pada inti sel-sel jaringan endometrium dan plasenta
' denganpewarnaan hemaktoksilin.. ...
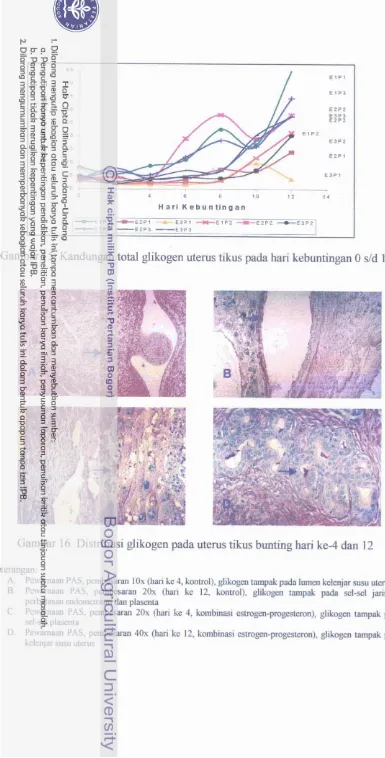
Kandungan total glikogen uterus tikus pada hari
Distribusi glikogen pada uterus tikus bunting hari ke- 4 dan 12 dengan pewarnaan PAS. ...
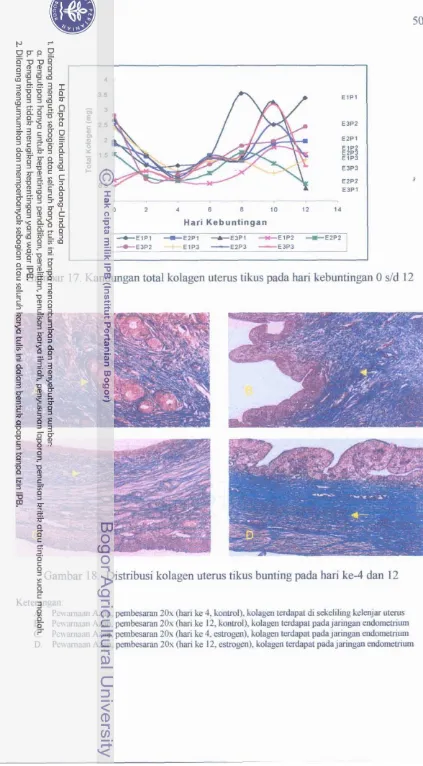
Kandungan total kolagen uterus tikus pada hari
...
kebuntingan 0 s/d 12.
Distribusi kolagen pada uterus tikus bunting
hari ke-4 dan 12 dengan pewarnaan Azan ...
Bobot kering uterus tikus pada penyuntikan sarnpai masa implantasi (H-5) dan plasentasi (H-12) ...
Kandungan total DNA uterus tikus bunting pada penyuntikan
...
sampai masa implantasi (H-5) dan plasentasi (H-12)
Kandungan total RNA uterus tikus bunting pada penyuntikan sarnpai masa implantasi (H-5) dan plasentasi (H-12) ...
Kandungan total glikogen uterus tikus bunting pada penyuntikan sampai masa implantasi (H-5) dan plasentasi (H-12) ...
Kandungan total kolagen uterus tikus bunting pada penyuntikan sampai masa implantasi (H-5) dan plasentasi (H-12) ...
Bobot badan mingguan anak tikus pada penyuntikan
...
sarnpai masa implantasi (H-5)
Bobot badan mingguan anak tikus pada penyuntikan
DAFTAR LAMPIRAN
Daftar sidik ragam total jumlah korpus luteum tikus pada hari kebuntingan 0- 12 pada dosis estradiol dan progesteron yang berbeda ...
Dafiar sidik ragam total jurnlah titik implantasi tikus pada hari kebuntingan 0-1 2 pada dosis estradiol dan progesteron yang berbeda ... .:.. ...
Daftar sidik ragam total selisih jurnlah korpus luteum dan titik implantasi tikus pada hari kebuntingan 0-12 pada dosis
...
estradiol dan progesteron yang berbeda
Daftar sidik ragam bobot basah uterus tikus pada hari kebuntingan 0- 12 pada dosis estradiol dan
progesteron yang berbeda , ...
Daftar sidik ragam bobot kering uterus tikus pada hari kebuntingan 0- 12 pada dosis estradiol dan
progesteron yang berbeda ...
Daftar sidik ragarn bobot air uterus tikus pada hari kebuntingan 0- 12 pada dosis estradiol dan
progesteron yang berbeda ...
Daftar sidik ragam kandungan total DNA uterus tikus pada hari kebuntingan 0-12 pada dosis estradiol dan
progesteron yang berbeda ...
Daftar sidik ragam kandungan total RNA uterus tikus pada hari kebuntingand-12 pada dosis estradiol dan
progesteron yang berbeda ...
Daftar sidik ragam kaildungan total glikogen uterus tikus pada hari kebuntingan 0-12 pada dosis estradiol
dan progesteron yang berbeda ...
Daftar sidik ragam kandungan total kolagen uterus tikus pada hari kebuntingan 0- 12 pada dosis estradiol dan progesteron yang berbeda ...
Halaman
79
79
80
80
8 1
8 1
82
82
83
1 1. Daftar sidik ragam total jumlah korpus luteum uterus tikus pada penyuntikan sampai masa implantasi
(H5) dan plasentasi (H12) ...
12. Dafiar sidik ragam total jumlah titik implantasi uterus tikus pada penyuntikan sampai masa implantasi
(H5) dan plasentasi (HI 2) ... 84
13. Daftar sidik ragam total selisih jumlah korpus luteum dan titik implantasi uterus tikus pada penyuntikan sampai
...
masa implantasi (H5) dan plasentasi (HI 2) 8 5
14. Daftar sidik ragam jumlah anak tikus pada penyuntikan
...
sampai masa implantasi (H5) dan plasentasi ( ~ 1 2 ) 8 5
1 5 . Daftar sidik ragam selisih jurnlah titik implantasi d m anak tikus pada penyuntikan sampai masa implantasi
(H5) dan plasentasi (H12) ... 86 16. Daftar sidik ragam bobot basah uterus tikus pada
penyuntikan sampai masa implantasi (H5) dan plasentasi
(H12) ...
17. Daftar sidik ragam bobot kering uterus tikus pada penyuntikan
...
sarnpai masa implantasi (H5) dan plasentasi (H12) 87
18. Daftar sidik ragam bobot air uterus tikus pada penyuntikan
sampai masa implantasi (H5) dan plasentasi (H12) ... 87
19. Dafiar sidik ragam kandungan total D N A uterus tikus pada penyuntikan sampai masa implantasi (H5) dan
...
plasentasi (H 12) 88
20. Daftar sidik ragam kandungan total RNA uterus tikus pada penyuntikan sampai masa implantasi (H5) dan
...
plasentasi (H12) 8 8
2 1. Daftar sidik ragam kandungan total glikogen uterus tikus pada penyuntikan sampai masa implantasi (H5) dan
...
Daftar sidik ragam kandungan total kolagen uterus tikus pada penyuntikan sampai masa implantasi (H5) dan
...
plasentasi (H 1 2)
Daftar sidik ragam bobot lahir total anak tikus pada penyuntikan sampai masa implantasi (H5) dan
plasentasi (HI 2) ...
Daflar sidik ragam bobot lahir rata-rata anak tikus pada penyuntikan sampai masa implantasi (H5) dan
...
plasentasi (H 12)
Daftar sidik ragam bobot badan anak tikus minggu ke-0
pada penyuntikan sampai masa impIantasi (H5) dan
...
plasentasi (HI 2)
D&ar sidik ragam bobot badan anak tikus minggu ke-1
pada penyuntikan sampai masa implantasi (H5) dan
...
plasentasi (H12)
Daftar sidik ragam bobot badan anak tikus minggu ke-2
pada penyuntikan sampai masa implantasi (H5) dan
plasentasi (HI 2) ...
Daftar sidik ragam bobot badan anak tikus minggu ke-3 pada penyuntikan sampai masa implantasi (H5) dan plasentasi (HI 2) ...
Daftar sidik ragam bobot badan anak tikus minggu ke-4
pada penyuntikan sampai masa impIantasi (H5) dan
plasentasi (HI 2) ...
Daftar sidik ragam pertambahan bobot badan harian an&
tikus pada penyuntikan sampai masa impIantasi (H5)
...
dan plasentasi (HI 2)
Rataan kandungan total DNA uterus tikus pada hari
kebuntingan 0-12 pada dosis estradiol dan
progesteron yang berbeda ...
Rataan kandungan total RNA uterus tikus pada hari
kebuntingan 0- 12 pada dosis estradiol dan
3 3 . Rataan kandungan total glikogen uterus tikus pada hari kebuntingan 0-12 pada dosis estradiol dan
progesteron yang berbeda ...
3 4 . Rataan kandungan total kolagen uterus tikus pada hari kebuntingan 0-12 pada dosis estradiol
dan progesteron yang berbeda ...
35. Rataan pertambahan bobot badan harian anak tikus pada penyuntikan sampai implantasi (H5)
PENDAHULUAN
Dalam upaya pemenuhan kebutuhan pangan asaI protein hewani diperlukan
adanya peningkatan kuantitas (populasi) maupun kualitas hewan ternak yang dimasa
sekarang ini masih merupakan pilihan konsumsi utama masyarakat. Akan tetapi
pengelolaan hewan ternak secara tradisional saja seperti yang masih banyak terdapat
pbJa rakyat pedesaan dewasa ini dirasakan kurang dapat mencapai peningkatan
seperti yang diharapkan Suatu usaha yang dapat meningkatkan kuantitas dan
kualitas hewan ternak dapat dilakukan melalui suatu penelitian dasar di bidang
fisiologi-reproduksi, terutama usaha pengembangan ternak politokus (ternak yang
dapat menghasilkan anak lebih dari satu dalam satu kali kebuntingan) sehingga usaha
peningkatan kebutuhan pangan asal protein hewani dapat cepat tercapai. Dalarn
berbagai penelitian, tikus putih dipakai sebagai hewan model karena tikus
mempunyai sifat yang cepat berkernbangbiak, selang generasinya pendek, siklus
reproduksinya singkat, cara pemeliharaannya mudah dan biayanya relatif murah
(Malole dan Pramono, 1989).
Keberhasilan peningkatan populasi suatu spesies tidak hanya berdasarkan
pada jurnlah pertemuan antara sperma dan sel telur saja, akan tetapi pengaruh hormon
kebuntingan juga sangat penting. Progesteron dan estradiol adaIah hormon-hormon
yang banyak sekali mengaiami perubahan pada saat birahi aan pada saat kebuntingan.
Selama kebuntingan, pertumbuhan dan perkembangan uterus dipengaruhi oleh
Anderson, 1986; Tucker, 1987). Kambing yang termasuk ternak politokus, insiden
kematian prenatalnya cukup tinggi, yang disebabkan
oleh tidak dihasilkannya progesteron pada plasenta atau sekresi progesteron tidak
mencukupi untuk mempertahankan kebuntingan (Nalbandov, 1976).
Aktivitas reproduksi ternak berlangsung setelah dewasa keIamin tercapai,
diawali dengan terjadinya ovulasi yang merupakan proses pelepasan sel telur yang
sudah matang dari ovarium. Apabila terdapat sperma pada saluran reproduksi maka
akan terjadi fertilisasi yaitu peleburan antara sel telur yang diovulasikan dengan
spermatozoa. Selanjutnya dua sel kelamin yang telah bersatu tersebut membutuhkan
tempat untuk berkembang dalam uterus melalui perlekatannya pada dinding uterus,
yang dikenal dengan istilah implantasi. Konsentrasi progesteron dan estradiol yang
cukup diperlukan untuk berlangsungnya proses ini.
Selama kebuntingan, estrogen dan progesteron dihasilkan oleh korpus luteum
dan plasenta. Pada tikus, korpus luteum merupakan penghasil utama progesteron
(Taya dan Greenwald, 1981). Pola sekresi estrogen dan progesteron selama
kebuntingan erat kaitannya dengan kebutuhan akan hormon-hormon tersebut, dan
peningkatan sekresi hormon tersebut erat kaitannya dengan peningkatan ukuran dan
aktivitas kelenjar penghasil hornion yaitu korpus luteum pada awal kebuntingan,
kemudian diikuti oleh plasenta pada pertengahan kebuntingan (Ichikawa et al., 1974;
Taya dan Greenwald, 1981).
Progesteron adalah hormon yang b e h n g s i untuk proliferasi sel-sel uterus
(dalam proses implantasi) dan untuk merangsang kelenjar susu uterus yang akan
Selama kebuntingan progesteron b e f i n g s i menekan produksi prostaglandin (Wilson
and Connell, 199 1) dan mengatur pengambilan relaksin (Downing and Hollingsworth, 1993) sehingga progesteron dapat menghambat kontraksi miometrium
yang menjamin ketenangan pemukiman embrio. Lebih lanjut progesteron berfungsi
menggertak pertumbuhan plasenta untuk keperluan penyaluran rnakanan dari induk
ke fetus dan zat buangan dari fetus ke induk. Progesteron juga bekerja sama dengan
estradiol untuk merangsang pertumbuhan kelenjar ambing pada induk yang meliputi
perbanyakan saluran (percabangan) dan sel-sel alveoli yang mensekresi susu selama
laktasi (Reeves, 1987). Kerja dari progesteron selalu diawali oleh kerja dari estradiol.
Estradiol merangsang perkembangan mukosa uterus di awal kebuntingan dan
perkembangan kelenjar susu selama kebuntingan (Guyton, 1994). Bersama-sama
dengan laktogen plasenta, estradiol berperan sebagai luteotropik yaitu bersifat
mempertahankan keberadaan korpus luteum supaya tetap mensekresikan progesteron
(Gibori et al., 1979).
Pola hormonal pada saat awal kebuntingan akan mempengaruhi pola
perkembangan dan implantasi embrio (Lapolt et al., 1990). Peningkatan estradioI
sebelum owlasi yang diikuti oleh peningkatan progesteron setelah ovu1asi berperan
dalam persiapan lingkungan uterus untuk proses implantasi dan perkembangan
embrio selanjutnya (Manalu dan Sumaryadi, 1995). Pertumbuhan dan perkembangan
embrio selama awal kebuntingan Iebih banyak ditentukan oleh pembentukan
lingkungan mikro uterus dan perkembangan embrio selanjutnya sangat bergantung
Sejauh ini informasi yang ada mengenai fingsi estradiol dan progesteron pada
peranannya dalam mempertahankan kebuntingan dilakukan dengan suatu teknik
superovulasi yang ditujukan untuk memperbanyak jumlah folikel dan jumlah sel telur
yang diovulasikan. Sedangkan penelitian ini dilakukan untuk melihat Iangsung
efektivitas pemberian estradiol dan progesteron selama masa kebuntingan dari
yeriode praimplantasi (hari ke-0, 2 dan 4), implantasi (hari ke-6), praplasentasi (hari
ke-8 dan 10) dan plasentasi (hari ke-12) pada tikus putih (Rattus sp.), serta pengaruh
pemberian estradiol dan progesteron selama 5 hari kebuntingan dan 12 hari
kebuntingan, dengan harapan bahwa pemberian hormon tersebut akan lebih
meningkatkan kondisi lingkungan mikro uterus sehingga jumlah anak yang dilahirkan
lebih banyak dan akan memiliki kondisi yang lebih baik dan hidup sehat sampai usia
lepas sapih.
Implantasi adalah suatu aktivitas fisiologis yang terjadi pada awal
kebuntingan dimana embrio akan melekat pada dinding uterus. Keberhasilan
implantasi merupakan salah satu indikasi tingkat fertilitas ternak. Dalam sektor
peternakan ini dapat diartikan sebagai suatu peningkatan produksi. Peningkatan
keberhasilan implantasi ini dimaksudkan untuk mengurangi kematian embrional
dengan perbaikan pertumbuhan prenatal sehingga induk dapat menghasilkan anak
yang sehat dengan daya tahan hidup yang baik dan sifat reproduksi yang lebih baik.
Ada hasil penelitian yang mengungkapkan bahwa perubahan pada Iingkungan
praimplantasi embrio dapat mempengaruhi perkembangan embrionya. Penambahan
progesteron pada sapi yang sedang bunting (1 sampai 4 hari urnur kebuntingan) akan
mendapatkan penambahan progesteron selama kebuntingannya. Pengaruh perubahan
lingkungan ini dikatakan sangat nyata pada hari-hari pertama dari perkembangan
fetus (awal kebuntingan). Demikian juga dengan estradiol (17-beta dan estrone)
dilaporkan mempunyai pengaruh terhadap implantasi (Garret et al., 1988)
Penelitian ini bertujuan untuk melihat bagaimana pertumbuhan dan
perkembangan sel-sel jaringan uterus tikus bunting yang telah disuntik dengan
estradiol dan progesteron dan apakah penyuntikan estradiol dan progesteron tersebut
dapat mempertahankan implantasi yang ada sehingga akhirnya dapat menghasilkan
anak sesuai dengan jumlah implantasi yang terjadi tanpa adanya peluruhan dalam
perjalanan masa kebuntingannya dan anak tersebut diharapkan akan mempunyai daya
tahan hidup sampai masa lepas sapih.
T u j u a n Penelitian
Melihat efek penyuntikan estradiol dan progesteron pada:
1. Tahapan perkembangan sel-sel jaringan uterus tikus selama masa kebuntingan.
2. Keberhasilan implantasi dengan mempertahankan titik implantasi yang ada.
3. Perkembangan embrio dan fetus.
4. Jumlah anak yang dilahirkan.
5 Pertumbuhan bobot badan anak sampai usia lepas sapih.
M a n f a a t d a n Kegunaan Hasil Penelitian
Hasil penelitian ini diharapkan dapat berguna sebagai landasan pengetahuan
seperti ternak ruminansia kecil kambing dan dornba juga pada babi. Dengan
keberhasilan peningkatan sifat/efisiensi reproduksi ini diharapkan akan dapat
meningkatkan kuantitas ataupun kualitas hewan ternak.
Hipotesis
1. Pemberian estradiol dan progesteron secara eksogen selarna kebuntingan akan
memperbaiki pertumbuhan dan perkembangan jaringan dan kelenjar uterus
2. Pemberian estradiol dan progesteron secara eksogen selarna kebuntingan akan
-
-meningkatkan keberhasilan implantasi sehingga jumlah embrio akan lebih banyak
dan dapat tumbuh dan berkembang menjadi fetus dan anak yang mempunyai daya
TINJAUAN PUSTAKA
Peningkatan efisiensi reproduksi dapat ditempuh dengan memperbaiki kondisi
kehidupan anak sejak periode di dalam kandungan (selama kebuntingan) dan setelah
anak tersebut dilahirkan (Dziuk, 1992) Melalui pengamatan yang sudah pernah
dilakukan pada domba ataupun kambing, pertumbuhan dan perkembangan anak selama
dalam kandungan dibagi menjadi dua yaitu fase uterus (fase embrional, sejak blastosis
sampai awal pembentukan fetus) dan fase plasenta (fase fetus, sejak pembentukan
fetus sampai dilahirkan) (Tomaszewska et al., 199 1 ; Manalu dan Sumaryadi, 1996a).
Dalarn perkembangannya selama di dalam uterus, embrio atau fetus mempunyai
beberapa kemungkinan yaitu embrio/fetus dapat melangsungkan hidupnya di dalam
uterus secara normal sampai saatnya dilahirkan, embrio/fetus yang sedang berkembang
tidak dapat melanjutkan hidupnya karena sesuatu hal atau ernbrio/fetus tersebut dapat
hidup sampai dilahirkan dengan pertumbuhannya yang menyimpang
(Hardjopranjoto,l995). Banyak faktor yang memegang peranan penting dalam
mempengaruhi pertumbuhan dan perkembangan embrio/fetus di dalam uterus induk.
Faktor-faktor penyebab kematian embrio dini pada berbagd spesies hewan diantaranya
faktor genetik, yang t e ja d i karena adanya perkawinan inbreeding, dapat menyebabkan
33 % kematian dini (King and Linarcs, 1983); faktor ketidakseimbangan hormonal antara estrogen dan progesteron dapat rnempengaruhi perjalanan embrio dari tuba
falopii ke uterus. Pada induk domba, menurut Davies dan Beck (1992) t e j a d i
kematian embrio yang tinggi selama 3 minggu pertarna kebuntingan, sehingga
makin menurun Hal ini berdasarkan kenyataan bahwa kekurangan korpus luteum
yang terbentuk diakibatkan kekurangan nutrisi dan stress. Berdasarkan laporan
Randell (1986), faktor infeksi menyebabkan hampir 90 % dari induk sapi perah yang barn melahirkan masih memperlihatkan adanya bakteri di dalam uterus 10 hari setelah
melahirkan yang kemudian dapat bersarang dalam alat kelamin. Faktor suhu
lingkungan juga sangat menentukan dalam mempertahankan embrio. Edwards dkk
(1968) melaporkan bahwa embrio babi sangat peka terhadap peningkatan suhu pada
usia kebuntingan dua minggu Embrio domba akan dapat mengalami kematian dini
sebesar 75 % bila berada dalam suhu yang terus menerus meningkat tanpa
mengganggu timbulnya birahi yang berikutnya. Sedangkan pada sapi dilaporkan
terjadi kematian dini setiap ada peningkatan suhu tubuh sebesar 1,5 "C. Faktor pakan
juga dapat menyebabkan kematian dini pada ternak secara langsung, terutama karena
kandungan zat-zat di dalamnya. Faktor kapasitas uterus dapat menyebabkan kematian
dini embrio karena keterbatasan tempat untuk bersarang pada uterus ataupun
persaingan dari embrio (pada hewanlternak multipara/politokus) untuk mendapatkan
nutrisi dari induk melalui sirkulasi di dalarn tubuh induk. Ribeiro et al., (1996)
melaporkan bahwa jumlah anak yang dilahirkan oleh seekor induk sangat tergantung
dari status fisiologis induk tersebut yaitu pada kecepatan ovulasi (ovulation rate),
ketahanan hidup prenatal (prenatal survival), aan kapasitas uterus. Semua ha1
tersebut tidak lepas dari peranan estrogen dan progesteron.
Selama dalam kandungan, zat-zat makanan yang dibutuhkan untuk
embrional sumber zat makanan yang diperlukan untuk pertumbuhan berasal dari
kelenjar uterus, sedangkan pada fase fetus sumber zat makanan berasal dari sistem
sirkulasi induk Oleh karena itu, pertumbuhan dan perkembangan jaringan uterus dan
plasenta sangat penting untuk meningkatkan sekresi zat-zat makanan yang diperIukan
oleh fetus domba ataupun kambing sejak dalam kandungan (Manalu dan Sumaryadi,
1996b). Perkembangan kelenjar uterus dan plasenta yang kurang baik ,kan
menyebabkan pertumbuhan dan perkembangan embrio yang rendah atau terjadi
kematian embrio dan fetus sehingga secara keselumhan akan menyebabkan rendahnya
jumlah anak yang dilahirkan Akibat lain yang bisa terjadi adalah bobot lahir anak
rendah sehingga kemarnpuan bertahan hidupnya sangat kurang dan akan terjadi
kematian pada awal kelahiran
Pertumbuhan dan perkembangan jaringan uterus dan plasenta berada di bawah
pengaruh hormon yang sekresinya bembah drastis seiring dengan umur kebuntingan,
jumlah korpus luteum, dan jumlah anak yang dikandung. Hormon yang dimaksud
adalah estrogen (diproduksi oleh korpus luteum dan plasenta), relaksin (korpus
luteum), progesteron (korpus luteum dan plasenta ) dan laktogen plasenta (pIasenta) (Tomaszewska ei al., 199 1)
Peningkatan sekresi hormon akan meningkatkan pertumbuhan dan
perkembangan lingkungm uterus (kelenjar utems d m plasenta) dombz ztzupun
kambing sehingga akan lebih banyak menghasilkan zat-zat makanan untuk
pertumbuhan dan perkembangan embrio dan fetus, akan memperbaiki perkembangan
penunhan kematian prenatal) dan Iebih pesat pertumbuhannya sehingga bobot lahir
yang optimum tercapai dan daya tahan hidup anak lebih baik (Manalu dan Sumaryadi,
Zat-zat makanan yang diperlukan untuk pertumbuhan dan perkembangan
embrio disediakan oleh kelenjar yang terdapat pada dinding uterus yang dikenal
dengan istilah kelenjar susu uterus (Mc. Donald, 1980). Pertumbuhan kelenjar ini
berada di bawah pengaruh hormon estrogen dan progesteron, namun hormon dan
faktor pertumbuhan lain yang dihasilkan oleh korpus luteum maupun uterus itu sendiri
tetap berperan bersama-sama dengan estrogen dan progesteron tersebut. Estradiol dan
IGF (Insulin-like Growth Factor) dapat mempengaruhi kemampuan sel uterus
memberikan respons terhadap progesteron dengan meningkatkan aktivitas CAMP
seluler (Aronica dan Katzenellenbogen, 1991). Pada babi, IGF-I yang dihasilkan oleh
uterus akan meningkat pada awal kebuntingan dan mencapai puncaknya pada waktu
umur kebuntingan yang ke-12 seiring dengan makin panjangnya blastosis (Letcher et
a1.,1989). IGF-I ini diduga akan merangsang aktivitas enzim aromatase pada
konseptus untuk meningkatkan biosintesis estradiol (Hofig et al., 1991). IGF-I dengan
kadar yang rendah terdapat pada fetus tikus sedangkan IGF-11 pada fetus tikus dan
serum tikus neonatus didapatkan pada kadar yang tinggi. Kadar ini akan menurun
sesudah beberapa saat setelzh kelahir= seiring dengan meningk~tnya IGF-I (Heath
and Smith, 1989). Progesteron dan estrogen mengalami banyak perubahan selama
siklus estrus dan kebuntingan mengiringi perubahan histologis dan kimiawi uterus.
ovulasi (fase proliferasi dan sekresi), progesteron meningkat. Akibat pengaruh kedua
hormon tersebut, aktivitas uterus berubah. Pada saat estrogen meningkat akan terjadi
peningkatan aktivitas enzim yang berperan dalarn proses pembelahan sel, sintesis
protein, glikogen dan glikoprotein (Norman and Litwack, 1987). Daiam
rnempersiapkan uterus menjadi lingkungan yang cocok untuk pertumbuhan dan
perkembangan embrio serta feras, estrogen bekerja dengan cara mempengaruhi sintesis
kolagen sehingga mengubah struktur kolagen uterus (Pastore et al., 1992),
meningkatkan kandungan glikogen pada uterus dengan mempengaruhi aktivitas
glikogen sintetase, meningkatkan metabolisme fosfolipid (Gould et aZ., 1978) serta
meningkatkan sintesis DNA dan proliferasi sel-sel uterus (Yamashita et al., 1990)
Secara histologis, pada awal kebuntingan akan tampak terjadinya pembelahan sel-sel
kelenjar dan stroma endometrium, kelenjar menjadi memanjang membentuk lekukan,
kemudian arteri pada endometrium tumbuh membentuk spiral @rickson, 1987; Berne
and Levy, 1988; Keys and King, 1995) Proses selanjutnya yang tejadi pada saat
progesteron meningkat adalah kelenjar uterus terus tumbuh membentuk lekukan yang
semakin banyak dan mendalam, sel-sel kelenjarnya mulai mensintesis giikogen dan
terbungkus pada vakuola besar di bagian dasar sel kelenjar. Pada saat yang sama
pembuluh darah uterus juga tumbuh pesat membentuk lilitan. Dengan semakin
meningkatzya sekresi progesteroc, sintesis glikcgen (Erickson 1987; Berne and Levy,
1988) dan senyawa glikoprotein dan protein (Norman and Litwack, 1987; Wheeler et
al., 1987) juga semakin pesat dan isinya dibebaskan ke d J a m lumen uterus. Secara
unsur-unsur jaringan epitel dan darah yang keluar dari nodus limfa yang diperlukan
untuk diferensiasi dan pertumbuhan embrio. Sehingga dapat dikatakan bahwa
perkembangan blastula menjadi embrio sampai terbentuk fetus sangat dipengaruhi oleh
perkembangan kelenjar susu uterus (Miller and Zhang, 1984).
Pertumbuhan dan perkembangan kelenjar susu uterus dimuiai dari masa
sebelum owlasi sampai periode implantasi sehiiigga pada periode ini diperlukan
ketersediaan progesteron dan estrogen yang cukup. Pada periode ini estrogen dan
progesteron dihasilkan oleh sel-sel teka folikel. dan sel-sel granulosa korpus luteum
(Stubbing et al., 1986; Southee e f al., 1988; Schiewe et al., 1990; Schiewe et al.,
1991). Estrogen akan meningkat sebelum owlasi kemudian turun selama fase luteal
dan akan meningkat secara perlahan-lahan selama periode kebuntingan berikutnya
sampai periode plasentasi dan akan meningkat drastis pada sisa kebuntingan
selanjutnya (Umo et aE., 1976; Trounson et al., 1977; M c Donald, 1980; Refsal et al.,
1991; Manalu e2 a!. , 1995a;b; Sumaryadi dan Manalu 1995 a,b;c; Tuju dan Manalu, 1995). Progesteron akan mulai meningkat setelah sel-sel granulosa korpus luteum
berkembang dan aktif mensintesis hormon tersebut (Umo et al., 1976; Trounson et
aL 1977; Boulfekhar and Brudieux, 1980; M c Donald, 1980; Refsal et a!., 1991;
Manalu et al., 1995a;b; Sumaryadi dan Manalu 1 995a;b;c; Tuju dan Manalu, 1995).
Penelitian tentang penyuntikan estrogen secarz 1.M (intrc muscular) dengan
dosis 5 ml pada awal kebuntingan (umur 11-15 hari kebuntingan) meningkatkan
perkembangan sistem pembuluh darah subepitel uterus babi (Keys and King, 1995).
dilaporkan menyebabkan peningkatan kematian ernbrional pada tikus (Halling el
aZ., 1993), dan penambahan progesteron telah ditunjukan rneningkatkan perturnbuhan
uterus (Krauss and Katzenellenbogen, 1993). Pada domba juga telah dilaporkan
bahwa penambahan progesteron pada awal kebuntingan rneningkatkan pertumbuhan
fetus (Kleernann el al., 1994). Pada babi dilaporkan bahwa penambahan progesteron
dapat meningkatkan pertumbuhan anak babi (Hard and Anderson, 1979; Ashworth,
1991). Penarnbahan progesteron dari luar tubuh pada tikus dan babi yang bunting
ternyata dapat rneningkatkan jumlah anak yang dilahirkan dan bobot lahir (Kendall and
Hays, 1960; Hard and Anderson, 1979; Ashworth, 1991). Pada mencit, estrogen yang
dihasilkan sesaat sebelum ovulasi akan merangsang proliferasi sel-sel epitel uterus pada
umur kebuntingan hari pertama dan kedua, sedangkan progesteron dari korpus luteum
yang barn terbentuk akan meningkatkan proliferasi sel-sel stroma yang potensial
dengan cara meningkatkan estrogen selama masa prairnplantasi (Parandoosh ei al.,
1995)
Ketersediaan zat-zat rnakanan di plasenta sangat erat kaitannya dengan
mobilisasi zat-zat rnakanan dalam darah induk (Egan, 1984), juga selanjutnya sangat
dipengaruhi oleh status hormonal induk terutama insulin, glukagon, kortisol,
somatotropin, tiroksin, prolaktin dan laktogen plasenta (Fain, 1979; Baurnan e t al.,
1982; Lewis e t aZ., 1988). .Uzn tetapi stztus fisiologis kebctingan iru sendiri terk&t
dengan kebutuhan zat-zat makanan bagi embrio atau fetus dan peningkatan estradiol
hormon-hormon metabolis seperti tiroksin dan kortisol pada kambing (Manalu et a].,
1995a; Manalu el al., 1997b) dan domba (Manalu dan Surnaryadi, 1996b).
Pertumbuhan anak setelah lahir sangat dipengaruhi oleh berat lahir anak dan
produksi susu induk. Berat sapih anak sangat dipengaruhi oleh produksi susu induk
dan berat lahir anak (Sumaryadi dan Manalu, 1995b) yang merupakan akumuIasi
pertumbuhan embrio sa;-.-ipai fetus Berat lahir juga sangat menentukan daya tahan
hidup anak selama periode prasapih (Bell, 1984; Tiesnamurti, 1992) serta menentukan
berat sapih.
Secara ringkas dapat disimpulkan bahwa pertumbuhan dan perkembangan
uterus, embrio dan fetus dipengaruhi terutama oleh konsentrasi horrnon reproduksi,
khususnya estrogen dan progesteron dalam darah induk selama kebuntingan.
Biologi Reproduksi Tikus Putih
Tikus merupakan hewan yang bersifat politokus dengan jumlah anak berkisar
antara 6 sampai 12 ekor setiap kali melahirkan (Harkness and Wagner, 1989). Tikus
Iaboratorium bisa hidup 2 hingga 3 tahun, mencapai usia dewasa antara 40 sampai 60
hari dan biasanya akan melakukan perkawinan pertarna pada saat mencapai usia 10
rninggu yang akan dilakukan saat estrus (Smith dan Mangkoewidjojo, 1987). Masa
pubertas (dewasa kelamin) dicapai pada umur 50 sampai 6 0 hari. Tikus siap
dikawinkan pada saat umur 65 sampai 110 hari dimana tikus betina dan jantan masing-
masing sudah mencapai bobot badan sekitar 250 gram dan 300 gram. Lama siklus
kelompok yaitu 1) proestrus (sekitar I2jam) , 2) estrus (sekitar 12jarn) , 3) metestrus I (15 jam); metestrus I1 (6 jam) dan 4) diestrus (57jam) (Baker, 1979). Masa
kebuntingan meliputi masa dari sejak terjadinya kopulasi, fertilisasi, implantasi sampai
saat anak dilahirkan berkisar antara 22 sampai 23 hari. Setelah terjadi fertilisasi pada
bagian ampula dari tuba falopii, sel telur yang telah dibuahi akan ditransportasikan ke
dalam uterus. Pada mamalia perjalanan ini memerlukan waktu 2 sampai 4 hari. Pada
tikus putih, ~embelahan menjadi 2 sel terjadi pada hari pertama dan kedua, pembelahan
menjadi 4 sel pada hari ke-2 dan 3 dan pembelahan menjadi 16 sel terjadi pada hari ke
4 kebuntingan. Pada mencit dan tikus, implantasi akan terjadi sehari setelah embrio
memasuki uterus. Pada tikus, waktu implantasi ini terjadi pada hari ke-5 kebuntingan
yaitu pada saat tahap biastosis sudah dicapai. Pada ternak, jarak antara waktu
fertilisasi dan proses implantasi bervariasi antara spesies, pada kambing dari hari ke-16
dan pada kuda dari hari ke-36 sampai dengan hari k e 4 0 kebuntingan (Hodgen dan
Itskowitz, 1988). Perubahan yang terjadi pada area implantasi yaitu adanya
peningkatan permeabilitas dari pembuluh darah yang diikuti adanya oedema atau
penebalan stroma yang mengelilingi blastosit. Aksi dari otot-otot uterus sangat
penting untuk penyebaran embrio sepanjang uterus bagi hewan atau ternak politokus.
Hal ini juga untuk mencegah tejadinya pengumpulan embrio pada satu daerah saja
yang biasanya dapat menyebabkan ksmatian embrio pada akhir kebuntingan (hlc.
Laren, 1982). Pembesaran abdomen biasanya terjadi pada hari ke-13 kebuntingan
(Baker, 1979). Tikus dapat menjalani perkawinan lagi (remaling) 24 jam setelah
16
Telah diketahui bahwa jumlah fetus yang dikandung erat kaitannya dengan
jumlah folikel yang berovulasi dan plasenta (Bradford et a/., 1986; Piper and Bindon,
1984). Dengan demikian sekresi progesteron dan estradiol sangat tergantung pada
jumlah korpus luteum dan massa plasenta Selama siklus estrus, uterus akan
mengalami vaskularisasi sarnpai 10 kali lipat sehingga dapat menyebabkan peningkatan
aliran darah dan menyebabkan perubahan keseimbangan antara estrogen dan -
progesteron pada saluran reproduksi (Schramm et al., 1984). Pada tikus (Taya and
Greenwald, 1981), mencit (Pointis et nL, 1981) dan hamster (Edwards et al., 1994)
korpus luteum merupakan sumber utama progesteron selama kebuntingan.
Peningkatan ukuran korpus luteum pada tikus selama pertengahan kebuntingan
ternyata mempengaruhi peningkatan sekresi progesteron (Ichikawa ef al., 1974).
Berbagai galur tikus yang dipakai dalam penelitian antara lain adalah Sprague-
Dawley, Wistar dan Long-Evans. Sprague-Dawley merupakan galur tikus albino
dengan kepala ramping dan ekor lebih panjang dari badannya, sedangkan Wistar
mempunyai kepala yang lebih lebar dan ekor yang lebih pendek. Galur Long-Evans
mempunyai bulu yang lebih gelap pada bagian atas kepala dan anterior tubuhnya
(Smith dan Mangkoewidjojo, 1987)
Berdasarkan pertimbangan bahwa siklus reproduksi tikus cukup pendek dan
mudah peme!iharaannya maka tikus sangat iepat untuk digunakan sebagai hewan
Horrnon Reproduksi
Estrogen dan progesteron adalah 2 hormon steroid yang paling banyak
peranannya dalam mengendalikan siklus dan proses yang terjadi dalam reproduksi
dengan perubahan yang terjadi pada konsentrasinya. Pada umumnya steroid
mempunyai struktur dasar yang sama yaitu eyelopenfano-perhydro-phenanhee.
Perubahan daya kerja steroid tergantung dari jumlah atom karbon yang terdapat dalam
struktur dan letak grup fungsionalnya, grup hngsional yang dimaksud adalah aldehide,
hidroksil, keton, klor, hidrokarbon yang jenuh, asam karbon dan metil
(Partodihardjo;l992) Semua hormon steroid berasal dari kolesterol yang kemudian
akan diubah menjadi pregnenolon. Oleh enzim yang spesifik kemudian pregnenolon
ini akan diubah langsung menjadi progesteron, dan melalui serangkaian proses/reaksi
yang panjang pregnenolon juga akan menghasilkan estrogen
Estrogen.
Estrogen adalah steroid yang secara kirnia maupun potensinya terdapat dalam
berbagai bentuk yaitu estrone yang mempunyai potensi yang rendah, estriol yang
berasal dari plasenta dan juga mempunyai potensi yang rendah, estradiol yang berasal
dari ovarium yang mempunyai potensi yang paling kuat. Estrogen mempunyai 18
atom karbon Pada bangsa mamalia betina indeks pertumbuhan dan perkembangan
dari folikel dapat diketahui dari produksi serum atau plasma estrogen, bila estrogen
meningkat berarti ada pertumbuhan dan perkembangan folikel, dasar fisiologis dari
hubungan ini adalah induksi dari aktivitas aromatase oleh FSH (Lasley et aL, 1988).
pertumbuhan tulang, pada pelunakan kulit, pada pertambahan berat jaringan uterus
karena hipertrofi dan hiperplasia endometrium dan miometrium, pada aktivitas
pergerakan fimbrie, pada kornifikasi vagina, pada pertumbuhan saluran-saluran dalam
kelenjar susu. Pada hewan betina, estrogen disintesis dan dibebaskan dalam sirkulasi
darah oleh ovarium baik oleh sel teka maupun oleh sel granulosa, plasenta dan adrenal
kortek. Konsentrasi estradiol dalam cairan antrum a r i folikel Graafian yang sedang
berkembang dilaporkan meningkat lebih dari 100 kali sebelum tejadinya ovulasi. Pada
percobaan yang dilakukan secara in vivo dan in vifro peningkatan ini merupakan
indikasi efek pengaturan kemampuan steroidogenesis sel-sel granulosa yang sedang
berkembang (Veldhuis et a ] . , 1986). Pada umumnya sintesis estrogen tidak ditimbun
dalam kelenjar penghasil tetapi tetap disintesis secara kontiniu dengan kadar yang
sangat rendah sekali dalam darah dan efek biologis yang ditimbulkannya sangat pendek
(Partodihardjo, 1992; Johnson dan Everitt, 1988). Banyak efek yang ditimbulkan oleh
estrogen pada proses metabolisme yang t e j a d i pada uterus termasuk meningkatkan
kandungan air uterus, merangsang pembentukan asam lemak dari asetat dan
meningkatkan metabolisme fosfolipid serta meningkatkan konsentrasi dan jumlah total
protein bukan kolagen. Estrogen merangsang metabolisme glikogen, baik melalui
perangsangan glikogenesis maupun melalui pengharnbatan terhadap glikogenolisis
tetapi perangsangan glzkogenesis Iebih ctamz. Pengzcn~h estrogen terhadap
metabolisme glikogen ini melibatkan enzim fosfirilase dan UDPG-glikogen sintetase.
UDPG-glikogen sinfefase dilaporkan banyak terdapat pada lapisan luar otot
19
Estrogen akan mengawali sejumlah perubahan kimiawi uterus yang berakibat adanya
sintesis protein, karena adanya peningkatan kecepatan enzim (Williams dan Provine,
1966). ~ a d a hewan rnultiparafpolitokus seperti tikus dan rnencit, estrogen akan
menyebabkan perubahan vaskularisasi pembuluh darah dan pertumbuhan endometrium
yang melibatkan adanya reaksi desidual di tempat terjadinya implantasi. Estrogen
dapat menyebabkan peningkatan aliran darch secara tidak langsung yaitu melalui
terjadinya peningkatan prostaglandin yang dapat menyebabkan vasodzlatasi pembuluh
darah pada rniometrium maupun pada endometrium (Schramm et a l , 1984). Estrogen
akan menyebabkan hiperemia uterus yang pada umurnnya berhubungan dengan
peningkatan sekresi cairan lurninal sehingga t e j a d i distensi lumen uterus. Estrogen
akan mengawali terjadinya implantasi pada bangsa rodensia melalui peningkatan
pertumbuhan sel uterus dan reseptor progesteron (mekanisme up regulation) serta
aksinya terhadap faktor-faktor pertumbuhan seperti EGF (epidermal growth factor)
atau T G F - a (transforming growth factor a), dimana estrogen akan meningkatkan
EGF dan TGF-a (Johnson dan Chatterjee, 1993). Reseptor dari EGF terdapat pada
semua tipe sel-sel uterus, dan dilaporkan bahwa reseptor ini akan meningkat setelah
adanya treatment estrogen pada tikus (Hanada et al., 1998). Estrogen secara spesifik
meningkatkan IGF-I dan EGF pada bangsa rodensia (Ohtani et aL, 1996). Pada sapi
juga dilaporkan adanya perubahan distribusi dari IGF-I dan EGF endometrium se!arna
siklus estrus (Ohtani et al., 1996). Dalam peranannya sebagai luteotropik, estrogen
berfUngsi untuk memelihara korpus luteum agar tetap ada dan mensekresikan
biosintesis kolesterol, mengatur aktifitas asil Co-A: kolesterol asillransferase ( A C A T )
agar tetap tersedia kolesterol untuk pembentukan progesteron (Azhar ei al., 1989).
Estrogen menyebabkan peningkatan berat dan fingsi korpus luteum selam pertengahan
kebuntingan (Nakamura dan Ichikawa, 1978 ; Rodway dan Rothchild, 1980).
Konsentrasi estradiol pada tikus yang sedang bunting tidak berubah mulai dari hari ke-
2 ad-mpai hari ke-12 kebuntingan, setelah itu meningkat berangsur-angsur sampai akhir
kebuntingan yaitu hari ke-22 (Taya dan Greenwald, 1981), sementara itu Tuju (1996)
melaporkan bahwa konsentrasi estradiol tidak berubah dari hari ke-4 sampai dengan
.. . hari ke-12 kebuntingan dan selanjutnya akan berangsur meningkat sampai akhir
kebuntingan
Progesteron.
Progesteron adalah steroid yang terdiri dari 21 atom karbon. Progesteron
merupakan substansi intermedia dari sintesa androgen, estrogen atau kortisol, oleh
sebab itu pada suatu keadaan keseimbangan yang terganggu pada organ tubuh yang
menghasilkan steroid, seperti ovarium, testis, korteks adrenal dan plasenta maka akan
dihasilkan progesteron (Turner dan Sagnara, 1988). Fungsi fisiologis progesteron
yaitu mempunyai efek anti radang seperti kortisol, pada miometrium akan menghambat
kontraksi dan menekan respons uterus terhadap pengaruh estrogen dan oksitosin, pada
endometrium akan merangsang tumbuhnya kelenjar-kelenjar susu uterus, pada spesies
tertentu akan merangsang tumbuhnya sel-sel pada permukaan endometrium
(deciduoma) untuk implantasi, progesteron juga dapat menyebabkan te jadinya
kelenjar ambing pada spesies tertentu seperti kucing, tikus dan babi. Pada hewan yang
bunting progesteron meniadakan kemungkinan terjadinya ovulasi (Partodihardjo,
1992; Nalbandov, 1976). Progesteron merangsang differensiasi sel-sel endometrium
dan menyiapkan implantasi, bersamaan dengan ini terjadi penurunan proliferasi dan
penurunan reseptor estrogen (down regulation) (Okulicz dan Balsamo, 1993).
Konsentrasi progesteron dalam serum saat siklus birahi pada domba lokal peranakan
ekor kurus berkorelasi secara positif dengan jumlah korpus luteum artinya semakin
banyak korpus Luteum yang terbentuk maka makin banyak kadar progesteron dalam
serum (Satyaningtijas dan Isdoni, 1997). Demikian juga pada hewan yang bunting
dilaporkan bahwa serum progesteron berkorelasi positif dengan jumlah anak yang
dikandung (Manalu dan Sumaryadi, 1996a; Manalu et al., 1995b). Konsentrasi
progesteron meningkat setelah terjadinya ovulasi dan akan semakin meningkat
terutama setelah periode plasentasi (Manalu dan Sumaryadi, 1995; Sumaryadi dan
Manalu, 1 9 9 5 ~ ) Tuju (1996) melaporkan bahwa konsentrasi progesteron pada tikus
bunting tidak mengalami perubahan dari umur kebuntingan ke-4 sampai dengan umur
kebuntingan ke-12 Sedangkan Rodway dan Rothchild (1980) menyatakan bahwa
dalam serum tikus bunting konsentrasi progesteron mulai meningkat mulai umur
kebuntingan ke- 12 sarnpai ke- 18. Peningkatan konsentrasi progesteron dalam serum
induk diikuti oleh peningkatan konsentrasi reseptor hormon tersebut (Vu E a i ef al.,
K i n e r j a Reproduksi
Reproduksi adalah suatu proses yang ditujukan untuk menghasilkan keturunan,
dimulai sejak bersatunya sel telur dan sel manilsperma sehingga menjadi mahluk hidup
baru. Reproduksi merupakan serangkaian proses majemuk yang disebabkan oleh
banyak faktor baik itu faktor dari dalam tubuh ataupun faktor dari luar tubuh. Tidak
munculnya salah satu faktor tersebut dapat menyebabkan terhambatnya reproduksi
sehingga dapat terjadi gangguan reproduksi. Proses reproduksi yang normal akan
diikuti produktifitas yang tinggi, dan sebaliknya bila daya reproduksi rendah akan
menghasilkan produktifitas temak yang rendah pula. Salah satu faktor penyebab
adanya gangguan reproduksi itu adalah ketidak-seimbangan hormonal. Faktor
hormonal ini erat kaitannya dengan kinerja reproduksi karena akan dapat
mempengaruhi ovulasi, fertilisasi, implantasi, hilangnya ovum serta mortalitas
embrional (Nalbandov, 1976) atau akan mempengaruhi pertumbuhan perkembangan
anak setelah dilahirkan sampai anak tersebut mencapai usia lepas sapih.
Fungsi fisiologis dari sistem reproduksi betina sangat kuat dipengaruhi oleh
hormon steroid yang dihasilkan oleh ovarium (Pineda, 1989 dan Murray, 1992) dan
hormon tersebut adalah estrogen dan progesteron yang menjalankan aksinya rnelalui
reseptor intraselulamya pada sel target atau jaringan. Reseptor bagi kedua macam
hormon ir.i sangat bervaiasi tergantung dslri s i k l ~ s repr~duksi dan dsii konseiltrasi
hormon tersebut yang berada dalam sirkulasi (Re et al., 1995).
Produk sekretoris dari uterus yang dihasilkan akibat adanya steroid sangat
plasentasi (Murray, 1992). Produk sekretoris tersebut termasuk protein dari susu yang
dihasilkan oleh sel epitel dari kelenjar uterus. Murray (1992) melaporkan bahwa efek
pemberian estrogen dan progesteron terhadap perubahan struktur pada sel epitelium
kelenjar uterus domba yang sudah diovariektomi temyata menunjukkan adanya
perubahan morfologi. Penambahan
17-P
estradiol menyebabkan hipertrofi dariepitelium uterus dan organel yang memproduksi protein. Seperti dikatakan di atas
bahwa ada faktor lain yang dapat mempengamhi implantasi yaitu faktor pertumbuhan
seperti EGF dan TGF-a. Estrogen dan progesteron sangat dibutuhkan untuk
pengaturan reseptor dari keduanya (Hanada et al., 1998)
Tingkat sekresi hormon gonadotropik juga menentukan banyaknya folikel yang
masak dan jumlah ovum yang diovulasikan. Secara genetik setiap jenis ternak
mempunyai kemampuan menghasilkan anak per kelahiran yang berbeda-beda. Jumlah
anak per kelahiran dipengaruhi oleh progesteron yang bekerja sama secara sinergis
dengan estrogen untuk menstimulasi ovulasi dengan menggertak pelepasan LH (Cole
dan Cupps, 1969). Kadar estrogen yang rendah dan progesteron yang tinggi dalam
proporsi yang tepat akan menyebabkan perpindahan sel telur yang telah dibuahi dari
ampula menuju ke uterus. Kematian embrional sebagian besar t e j a d i pada umur
kebuntingan dini dan daya hidup (survival) embrio selama periode tersebut tergantung
pada kondisi lingkungan uterus yang ada di bawah pengaturan progesteron
(Nalbandov, 1976 dan Betteridge, 1986). Manalu dan Sumaryadi (1995) melaporkan
bahwa peningkatan sekresi progesteron dan estradiol selama kebuntingan dan akhir
fetus. Peningkatan sekresi ini erat kaitannya dengan fungsi kedua hormon tersebut
pada endometrium sebagai wadah dan sumber nutrisi bagi fetus. Efek estrogen pada
endometrium adalah meningkatkan pertumbuhan sel atau fase proliferasi dan
mengawali kerja progesteron dengan meningkatkan reseptor bagi progesteron
sedangkan progesteron menghambat proliferasi sel dengan cara menurunkan reseptor
untuk estrogen dan menyebabkan diferensiasi berbagai macam tipe sel endometrium
untuk persiapan implantasi dan sekresi susu uterus (Okulicz dan Balsamo, 1993).
Tanda-tanda awaI dari adanya implantasi adalah adanya peningkatan
permeabilitas uterus yang terjadi secara lokal yang akan menyebabkan terjadinya
oedema. Pembengkakan ini diduga merupakan suatu rangsangan untuk mitosis dari
sel-sel stroma sekitarnya dan pertumbuhan uterus. Peningkatan kecepatan proliferasi
sel endotel endometrium sangat jelas dari hari kebuntingan tikus yang ke 3 sampai
dengan hari ke-5 yang menyebabkan pertumbuhan pembuluh darah dan secara
bersama-sama juga pertumbuhan jaringan uterus lainnya