KEMAMPUAN BAKTERI KITINOLITIK LOKAL DALAM
MENGHAMBAT PERTUMBUHAN Fusarium sp TANAMAN KOPI
SKRIPSI
Diajukan untuk melengkapi dan memenuhi syarat mencapai gelar Sarjana Sains
SARIANTI SIHOMBING
060805041
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS SUMATERA UTARA
PERSETUJUAN
Judul : KEMAMPUAN BAKTERI KITINOLITIK LOKAL DALAM MENGHAMBAT PERTUMBUHAN Fusarium sp. TANAMAN KOPI
Kategori : SKRIPSI
Nama : SARIANTI SIHOMBING
Nomor Induk Mahasiswa : 060805041
Program Studi : SARJANA (S1) BIOLOGI
Departemen : BIOLOGI
Fakultas : MATEMATIKA DAN ILMU PENGETAHUAN ALAM (FMIPA) UNIVERSITAS SUMATERA UTARA
Diluluskan di
Medan, Desember 2010
Komisi Pembimbing :
Pembimbing 2 Pembimbing 1
Drs. Kiki Nurtjahja, M.Sc Prof. Dr. Dwi Suryanto, M.Sc NIP. 196212111998031001 NIP. 196404091994031003
Diketahui/Disetujui oleh
Departemen Biologi FMIPA USU Ketua,
PERNYATAAN
KEMAMPUAN BAKTERI KITINOLITIK LOKAL DALAM MENGHAMBAT PERTUMBUHAN Fusarium sp. TANAMAN KOPI
SKRIPSI
Saya mengakui bahwa skripsi ini adalah hasil kerja saya sendiri, kecuali beberapa kutipan dan ringkasan yang masing-masing disebutkan sumbernya.
Medan,
PENGHARGAAN
Puji dan syukur penulis panjatkan kepada Allah SWT yang Maha Pengasih Lagi Maha Penyayang atas limpahan rahmat kekuatan dan kemudahan sehingga penulis dapat menyelesaikan skripsi ini dengan judul, “Kemampuan Bakteri Kitinolitik Lokal dalam Menghambat Pertumbuhan Fusarium sp. Tanaman Kopi”.
Skripsi ini saya persembahkan kepada seluruh keluarga yang saya sayangi Ayahanda dan ibunda (Saibun Sihombing dan Nurbaini Ritonga) yang sudah memberikan banyak doa, harapan dan dorongan dan pengorbanan sehingga saya dapat menyelesaikan perkuliahan dan skripsi ini. Kakak dan abangku (Hotlan, lamhot, Arlita, A. Md, Nursaima, Muhammad Syukur, SE, Subuh Panangian dan Nurasimah. penulis ucapkan ribuan terima kasih atas pengorbanan moril maupun material, segala cinta, motivasi, kesabaran serta doa yang tidak akan pernah bisa penulis balas sampai kapanpun.
Ucapan terima kasih penulis sampaikan kepada Prof. Dr. Dwi Suryanto, M.Sc selaku Dosen Pembimbing I dan Drs. Kiki Nurtjahja, M.Sc selaku Dosen Pembimbing II atas bimbingan, arahan dan bantuannya dalam pembuatan skripsi ini. Kepada Dra. Nunuk Priyani, M.Sc dan Dr. Suci Rahayu, M.Si selaku dosen penguji, penulis mengucapkan banyak terima kasih atas arahan dan saran dalam penyempurnaan penulisan skripsi ini.
Ucapan terima kasih juga saya sampaikan kepada Dra. Nunuk Priyani, M.Sc sebagai Pembimbing Akademik dan sekretaris Departemen Biologi FMIPA USU, Prof. Dr. Dwi Suryanto, M.Sc sebagai Ketua Departemen Biologi FMIPA USU, Prof. Erman Munir, M.Sc sebagai Kepala Laboratorium Mikrobiologi, Seluruh Staf Pengajar Departemen Biologi FMIPA USU. Bapak Sukirmanto, Ibu Nurhasni Muluk, Ibu Roslina Ginting dan Bang Erwin selaku staf pegawai Departemen Biologi FMIPA USU.
Sebagai makhluk ciptaanNya yang tidak lepas dari kekhilafan, kesalahan dan keterbatasan, Penulis sangat menyadari bahwa hasil penelitian ini masih jauh dari kesempurnaan. Oleh karena itu penulis mengharapkan kritik dan saran yang bersifat membangun dalam melengkapi kekurangan serta penyempurnaan skripsi ini. Akhirnya dengan penuh ketulusan dan kerendahan hati, penulis mengharapkan hasil penelitian ini bermanfaat bagi kita semua.
Medan, November 2010
ABSTRAK
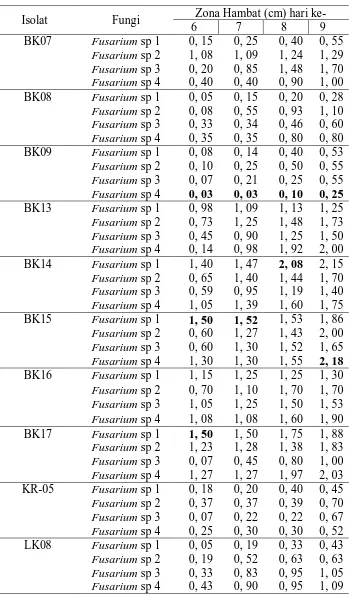
Penelitian tentang kemampuan bakteri kitinolitik dalam menghambat pertumbuhan Fusarium sp. tanaman kopi telah dilakukan dari bulan Juni sampai November. Fungi patogen diisolasi dari daun, buah, ranting yang sakit tanaman kopi di Brastagi. Sedangkan isolasi, identifikasi, dan uji antagonis dilakukan di Laboratorium Mikrobiologi, Departemen Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Sumatera Utara. Penelitian ini bertujuan untuk mengetahui kemampuan bakteri kitinolitik dalam menghambat pertumbuhan Fusarium sp, salah satu penyebab penyakit pada tanaman kopi. Dari hasil isolasi diperoleh 4 jenis Fusarium yaitu Fusarium sp. 1, Fusarium sp. 2, Fusarium sp. 3, dan Fusarium sp. 4. Isolat-isolat bakteri kitinolitik BK07, BK08, BK09, BK13, BK14, BK15, BK16, BK17, KR05, LK08 diperoleh dari Laboratorium Mikrobiologi, Universitas Sumatera Utara. Uji antagonis dilakukan dalam media garam minimum kitin (MGMK) dengan melihat zona hambat terhadap pertumbuhan Fusarium sp. Pada hari kesembilan isolat BK15 memiliki zona hambat bakteri tertinggi dalam menghambat pertumbuhan Fusarium dengan zona hambat sebesar 2,18 cm dan isolat BK09 memiliki zona hambat terendah terhadap Fusarium sp. 4 sebesar 0,25 cm. Hasil pengamatan mikroskopis bakteri kitinolitik mengakibatkan terjadinya hifa abnormal, diantaranya hifa lisis, patah, melilit, menggulung, keriting.
THE ABILITY OF CHITINOLYTIC BACTERIA TO INHIBIT THE GROWTH OF Fusarium sp. IN COFFEE PLANT
ABSTRACT
A study on the ability of chitinolytic bacteria to inhibit the growth of Fusarium sp. isolated from coffee plant, has been conducted from Juni until Desember 2010. The pathogenic fungi were isolated from leaves, fruits, and branches of coffee plant in Brastagi. Whereas fungi isolation, identification and antagonistic examination were conduced in Microbiology Laboratory, Biology Department, Faculty of Mathematic and Natural Sciences, Sumatera Utara University. The purpose of this study was to investigate the chitinolytic bacteria to inhibit the growth of Fusarium sp. pathogenic in the coffee plant. The results showed that four species of Fusarium were obtained from the coffee, which were Fusarium sp. 1, Fusarium sp. 2, Fusarium sp. 3, and Fusarium sp. 4. The chitinolytic bacteria tested were BK07, BK08, BK09, BK13, BK14, BK15, BK16, BK17, KR05, LK08 obtained from Microbiology Laboratory, Department of Biology, Sumatera Utara University. The Antagonistic assay has been conducted using chitin salt minimum media. The antagonistic assay was observed after ninth days of incubation, the results showed that BK15 effective bacteria in inhibiting the growth of the Fusarium with the diameter of clear zone of 2,18 cm, mean while BK09 showed relative low ability to inhibit the growth of the Fusarium sp. 4 with the diameter of clear zone of 0,25 cm. Based on microscopic observation, the chitinolytic bacteria caused abnormal hypae such as lytic, broken, coiled, rolled, and curly hyphae.
DAFTAR ISI Bab. 2. Bahan dan Metoda 2.1. Waktu dan Tempat 5 Bab 3. Hasil dan Pembahasan 3.1 Isolasi dan Identifikasi Fungi Patogen pada Bab 4. Kesimpulan dan Saran 4.1 Kesimpulan 15
4.2 Saran 15
DAFTAR TABEL
Halaman
Tabel 1. Ciri-ciri Fusarium sp. pada Tanaman Kopi 9 Tabel 2. Pertambahan Zona Hambat (cm) Bakteri Kitinolitik
terhadap 4 jenis Fusarium sp.
DAFTAR GAMBAR
Halaman
Gambar 1. Jenis Fusarium Tanaman Kopi 8
Gambar 2. Mikroskopis Konidia Fusarium 9
Gambar 3. Uji Antagonis Bakteri Kitinolitik terhadap Beberapa Fusarium sp. pada Hari Kesembilan
13
Gambar 4. Perbandingan Hifa Normal dengan Hifa Abnormal Fusarium sp.
DAFTAR LAMPIRAN
Halaman
Lampiran 1. Pembuatan koloidal khitin dengan cara hidrolisis parsial (Rodriquez-Kabana et al., 1983 ).
20
ABSTRAK
Penelitian tentang kemampuan bakteri kitinolitik dalam menghambat pertumbuhan Fusarium sp. tanaman kopi telah dilakukan dari bulan Juni sampai November. Fungi patogen diisolasi dari daun, buah, ranting yang sakit tanaman kopi di Brastagi. Sedangkan isolasi, identifikasi, dan uji antagonis dilakukan di Laboratorium Mikrobiologi, Departemen Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Sumatera Utara. Penelitian ini bertujuan untuk mengetahui kemampuan bakteri kitinolitik dalam menghambat pertumbuhan Fusarium sp, salah satu penyebab penyakit pada tanaman kopi. Dari hasil isolasi diperoleh 4 jenis Fusarium yaitu Fusarium sp. 1, Fusarium sp. 2, Fusarium sp. 3, dan Fusarium sp. 4. Isolat-isolat bakteri kitinolitik BK07, BK08, BK09, BK13, BK14, BK15, BK16, BK17, KR05, LK08 diperoleh dari Laboratorium Mikrobiologi, Universitas Sumatera Utara. Uji antagonis dilakukan dalam media garam minimum kitin (MGMK) dengan melihat zona hambat terhadap pertumbuhan Fusarium sp. Pada hari kesembilan isolat BK15 memiliki zona hambat bakteri tertinggi dalam menghambat pertumbuhan Fusarium dengan zona hambat sebesar 2,18 cm dan isolat BK09 memiliki zona hambat terendah terhadap Fusarium sp. 4 sebesar 0,25 cm. Hasil pengamatan mikroskopis bakteri kitinolitik mengakibatkan terjadinya hifa abnormal, diantaranya hifa lisis, patah, melilit, menggulung, keriting.
THE ABILITY OF CHITINOLYTIC BACTERIA TO INHIBIT THE GROWTH OF Fusarium sp. IN COFFEE PLANT
ABSTRACT
A study on the ability of chitinolytic bacteria to inhibit the growth of Fusarium sp. isolated from coffee plant, has been conducted from Juni until Desember 2010. The pathogenic fungi were isolated from leaves, fruits, and branches of coffee plant in Brastagi. Whereas fungi isolation, identification and antagonistic examination were conduced in Microbiology Laboratory, Biology Department, Faculty of Mathematic and Natural Sciences, Sumatera Utara University. The purpose of this study was to investigate the chitinolytic bacteria to inhibit the growth of Fusarium sp. pathogenic in the coffee plant. The results showed that four species of Fusarium were obtained from the coffee, which were Fusarium sp. 1, Fusarium sp. 2, Fusarium sp. 3, and Fusarium sp. 4. The chitinolytic bacteria tested were BK07, BK08, BK09, BK13, BK14, BK15, BK16, BK17, KR05, LK08 obtained from Microbiology Laboratory, Department of Biology, Sumatera Utara University. The Antagonistic assay has been conducted using chitin salt minimum media. The antagonistic assay was observed after ninth days of incubation, the results showed that BK15 effective bacteria in inhibiting the growth of the Fusarium with the diameter of clear zone of 2,18 cm, mean while BK09 showed relative low ability to inhibit the growth of the Fusarium sp. 4 with the diameter of clear zone of 0,25 cm. Based on microscopic observation, the chitinolytic bacteria caused abnormal hypae such as lytic, broken, coiled, rolled, and curly hyphae.
BAB 1
PENDAHULUAN
1.1Latar Belakang
Kopi adalah spesies tanaman berbentuk pohon dan termasuk dalam famili Rubiaceae. Tanaman ini tumbuh tegak, bercabang dan dapat mencapai tinggi 12 m (Danarti & Najiyati, 1999). Tanaman kopi merupakan komoditas ekspor yang mempunyai nilai ekonomis yang relatif tinggi di pasaran dunia, di samping merupakan salah satu komoditas unggulan yang dikembangkan di Jawa Barat. Sudah hampir tiga abad kopi diusahakan penanamannya di Indonesia untuk memenuhi kebutuhan konsumsi di dalam negeri dan luar negeri (Siswoputranto, 1978). Lebih dari 90% tanaman kopi diusahakan oleh rakyat. Di dunia perdagangan dikenal beberapa golongan kopi, akan tetapi yang paling sering dibudidayakan adalah kopi arabika, robusta, dan liberika (Danarti & Najiyati, 1999).
Penyakit tanaman menyebabkan kerusakan dan kehilangan hasil produksi tanaman akibat adanya patogen (Agrios, 1996). Fungi patogen dapat masuk ke dalam badan tumbuhan melalui luka, lubang alami seperti stomata dan hidatoda, dan patogen dapat masuk langsung menembus permukaan tumbuhan yang utuh. Tanaman tertutup oleh lapisan pelindung seperti tanaman berkayu, sehingga patogen tidak dapat masuk melaui mulut hidatoda, maka patogen ini dapat masuk melalui luka. Luka dapat terjadi secara langsung atau tidak langsung seperti kegiatan manusia, hujan, atau sinar matahari yang terlalu kuat. Lubang alami yang sering dipakai sebagai jalan masuk fungi patogen adalah stomata sehingga patogen dapat menginfeksi tanaman (Semangun, 1996).
tanaman dan mikroorganisme hidup bersama (Rao, 1994). Patogen tular tanah merupakan sekelompok mikroba pengganggu tanaman yang keberadaan dan hidupnya di dalam tanah, yang dapat menimbulkan kerugian hasil panen dalam skala besar, Persaingan sumber nutrisi di lingkungan tanah merupakan hal penting bagi keberadaan beberapa mikroorganisme dalam meningkatkan jumlah dan keaktifannya (Soesanto, 2008).
Di Indonesia tanaman kopi merupakan salah satu tanaman perkebunan yang penting dan termasuk komoditas ekspor. Salah satu penyakit yang terdapat dalam kopi adalah Fusarium. Organisme penyebab penyakit ini biasanya masuk melalui akar muda dan kemudian tumbuh dan berkembang sehingga akan mengkonduksi bagian pembuluh dari akar dan batang. Di bagian pembuluh batang tersumbat dan gagal menyalurkan air ke daun (Miller et al., 1986). Penyakit ini juga menyebabkan kerugian yang sangat besar pada perkebunan kopi di Afrika (Okeniyi, 2007).
Melihat kemampuan bakteri kitinolitik lokal dalam menghambat pertumbuhan patogen penyebab penyakit pada tanaman harus terus dilakukan. Salah satunya adalah dengan melakukan uji antagonis antara bakteri kitinolitik dengan Fusarium sp. penyebab penyakit tracheomycosis pada tanaman kopi dan melihat pengaruhnya terhadap perkembangan hifa fungi tersebut. Berdasarkan hasil survei lapangan dan pengumpulan informasi dari masyarakat sekitar, petani kopi biasanya menggunakan pestisida untuk membunuh serangan hama dan penyakit, namun adakalanya para petani menebang pohon kopi yang dianggap tidak memberikan hasil lagi, bahkan membiarkan begitu saja dan menganggap penyakit tersebut tidak berbahaya bagi tanaman lainnya. Telah banyak dilakukan penelitian tentang kemampuan bakteri kitinolitik, diantaranya adalah isolat LB01, LK01, KR05, LK03 dan KR03 mampu menghambat pertumbuhan P. citrinum dan isolat DS01, LK01, LB02, KR03 dan LK03 mampu menghambat fungi F. semitectum dan G. boninense (Suryanto & Munir, 2006) dan mampu menghambat fungi F. oxysporum penyebab layu Fusarium pada kecambah cabai merah (Patonah, 2008), sehingga diperlukan pengujian dengan memanfaatkan isolat bakteri kitinolitik yang potensial untuk mengendalikan fungi patogen Fusarium sp. pada tanaman kopi.
1.2Permasalahan
Fusarium sp. merupakan patogen penyebab penyakit tracheomycosis pada tanaman kopi, sehingga perlu dilakukan pengujian dengan menggunakan bakteri kitinolitik lokal yang diduga mampu menghambat pertumbuhan Fusarium sp.
1.3Tujuan Penelitian
1.4Hipotesis
Bakteri kitinolitik memiliki kemampuan untuk menghambat pertumbuhan Fusarium yang diisolasi dari tanaman kopi sehingga dapat digunakan sebagai agen pengendali hayati.
1.5Manfaat
BAB 2
BAHAN DAN METODE
2.1. Waktu dan Tempat
Penelitian ini dilaksanakan dari bulan Juni sampai November tahun 2010 bertempat di Laboratorium Mikrobiologi Departemen Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Sumatera Utara. Sampel tanaman sakit diambil di daerah Brastagi.
2.2 Alat dan Bahan
Alat yang digunakan dalam penelitian ini adalah cawan Petri, tabung reaksi, rak tabung reaksi, cork borer, Erlenmeyer, spatula, jarum ose, bunsen, beaker glass, pro pipet, pipet serologi, gelas ukur, kertas saring, corong, autoclave, stirer, hot plate, inkubator, cutter, label tempel, vortex, jangka sorong, timbangan analitik. Bahan-bahan yang diperlukan adalah bakteri kitinolitik, akuades, media potato dextrose agar (PDA), potato dextrosa broth (PDB), koloidal kitin, larutan Mc Farland, kertas cakram, HCl 10N, larutan garam (K2HPO4, KH2PO4, MgSO4.7H2O, FeSO4.7H2O, ZnSO4, MnCl2), dan agar-agar powder.
2.3 Isolat Bakteri Kitinolitik Lokal
6,8 dan diinkubasi pada suhu 30oC. Pembuatan koloidal kitin dilakukan dengan cara hidrolisis parsial menurut Rodriquez-Kabana et al., (1983). Proses pembuatan dapat dilihat pada lampiran 1 halaman 19.
2.4 Isolasi dan Identifikasi Fungi Patogen pada Tanaman Kopi
Pengambilan sampel tanaman terserang fungi pada tanaman kopi dilakukan di Brastagi, Kabupaten Karo. Sampel tanaman yang terserang (daun, pucuk, ranting, dan buah) dimasukkan ke dalam plastik yang sudah diberi alkohol 70% kemudian dibawa ke Laboratorium Mikrobiologi Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Sumatera Utara. Daun, pucuk, ranting, dan buah dipotong sebesar 3 cm dan dimasukkan ke dalam cawan petri berisi media PDA, ditutup dan diinkubasi selama 2-5 hari. Hasil isolasi fungi yang berupa biakan murni, diidentifikasi berdasarkan morfologi makroskopis, kemudian dilakukan identifikasi di bawah mikroskop dengan menggunakan panduan buku identifikasi (Pitt & Hocking, 1997; Gandjar et al., 1999) dengan metode block square.
2.5 Uji Antagonisme Bakteri Kitinolitik terhadap Fungi Patogen Tanaman
Biakan fungi kopi diinokulasikan pada media garam minimum kitin (MGMC). Komposisi dapat dilihat pada lampiran 2 halaman 20, dengan jarak 3,5 cm dari cakram tempat inokulum bakteri pada bagian tengah. Suspensi bakteri kitinolitik yang telah dibuat dengan konsentrasi ≈108 sel/ml (Standar Mc. Farland), kemudian diambil 0,01 ml dan diteteskan di atas kertas cakram dan diinokulasikan ke dalam media di bagian pinggir, setiap jenis bakteri kitinolitik dibuat 2 titik pengulangan dalam satu cawan Petri. Biakan diinkubasi pada suhu 30o C. Zona hambat terhadap miselia jamur kopi diamati mulai dari hari ke-2 sampai ke-9. Zona hambat dihitung dengan mengukur diameter koloni fungi yang tidak terhambat bakteri kitinolitik yang dikurangi dengan diameter koloni fungi patogen yang menjauhi isolat bakteri kitinolitik (terhambat), kemudian dibagi dua (Martorejo et al., 2001). Pengukuran diameter zona hambat dengan menggunakan jangka sorong.
2.6 Pengamatan Struktur Hifa Fungi Setelah Uji Antagonis
BAB 3
HASIL DAN PEMBAHASAN
3.1 Isolasi Fungi dari Tanaman Kopi yang Terserang Penyakit
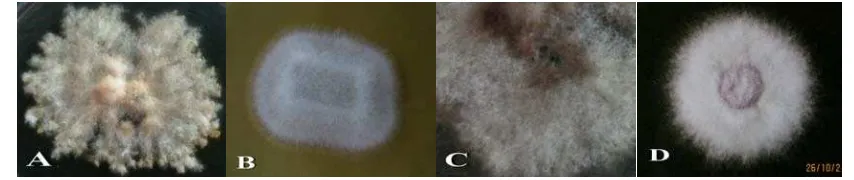
Isolasi fungi dari tanaman kopi diperoleh 4 jenis fungi Fusarium yang diduga patogen tanaman kopi yang terlihat berbeda. Identifikasi dilakukan berdasarkan bentuk koloni dan penegak hifa (Gambar 1). Fungi ini dapat bertahan dalam bibit dan merupakan jamur tular tanah yang mampu bertahan lama di dalam tanah sebagai klamidospora sehingga sulit dikendalikan (Brown & Ogle, 1997). Patogen tular tanah merupakan sekelompok mikroba pengganggu tanaman yang keberadaan dan hidupnya di dalam tanah, yang dapat menimbulkan kerugian hasil panen dalam skala besar (Soesanto, 2008).
Gambar 1. Jenis Fusarium Tanaman Kopi, A. Fusarium sp. 1, B. Fusarium sp. 2, C. Fusarium sp. 3, D. Fusarium sp. 4
cukup besar di perkebunan kopi di Afrika oleh xylarioides (Okeniyi, 2007), oxysporum pada tanaman pisang (Brown & Ogle, 1997) dan cabe (Patonah, 2008).
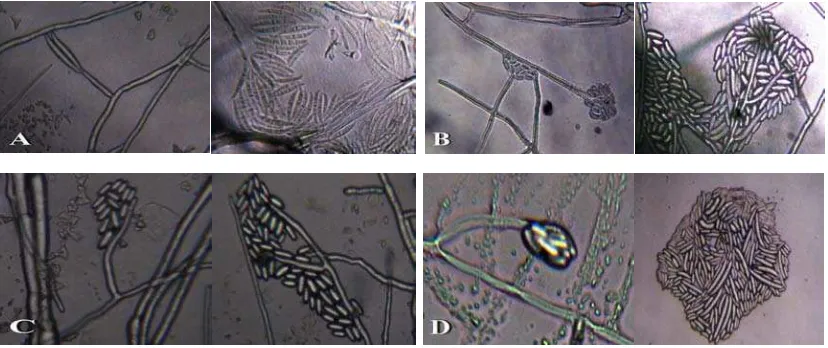
Pada tanaman yang masih sangat muda penyakit dapat menyebabkan tanaman mati secara mendadak, karena pada pangkal batang terjadi kerusakan atau kanker menggelang, sedangkan tanaman dewasa yang terinfeksi sering dapat bertahan terus dan membentuk buah tetapi hasilnya sangat sedikit dan kecil-kecil. Sejumlah cara pengendaliannya telah diteliti, namun belum memberikan hasil yang memuaskan (Semangun, 1996). Berdasarkan hasil pengamatan mikroskopis, dapat dilihat bentuk konidia Fusarium sp. (Gambar 2) tanaman kopi serta ciri-cirinya dapat dilihat pada Tabel 1.
Gambar 2. Mikroskopis Konidia Fusarium, A. Konidia Fusarium sp. 1, B. Konidia Fusarium sp. 2, C. Konidia Fusarium sp. 3, D. Konidia Fusarium sp. 4. (Perbesaran 10 X 40).
Tabel 1. Ciri-ciri Fusarium sp. pada tanaman kopi
Isolat Ciri-ciri
Fusarium sp. 1
Koloni berwarna oranye muda, hifa bersekat, makrokonidia memiliki sekat 3-8, dasar koloni orange muda, permukaannya kasar dan bergerigi. Fusarium
sp. 2
Koloni berwarna putih seperti kapas, hifa bersekat, makrokonidia memiliki sekat 2-4, dasar koloni halus dan putih, permukaannya kasar dan putih
Fusarium sp. 3
Koloni berwarna putih dan bagian tengah berwarna coklat muda, makrokonidia memiliki sekat 2-4, dasar koloni halus dan putih, permukaanya putih dan kasar.
Fusarium sp. 4
3.2 Uji Antagonisme Bakteri Kitinolitik terhadap Fungi Patogen Tanaman Kopi
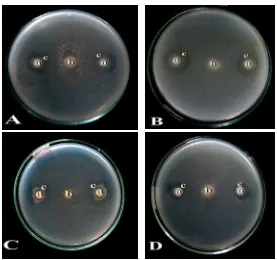
Dari hasil uji antagonis antara bakteri kitinolitik dengan Fusarium sp. menunjukkan kemampuan bakteri dalam menghambat pertumbuhan miselium fungi berbeda, ditandai dengan terbentuknya zona bening yang dihasilkan oleh bakteri kitinolitik (Tabel 2). Zona bening terbentuk di sekitar koloni pertumbuhan bakteri kitinolitik karena adanya aktivitas bakteri yang diduga mensekresikan enzim dalam medium pertumbuhan dan memecah kitin. Faktor yang diduga menjadi penyebab perbedaan kemampuan isolat bakteri kitinolitik dalam uji antagonis terhadap fungi patogen adalah komposisi dan keberadaan kitin dalam miselium fungi patogen, laju pertumbuhan isolat dan miselium fungi dan adanya perbedaan spesies isolat bakteri kitinolitik. Dinding sel fungi kelas Ascomycetes, Basidiomycetes, dan Deuteromycetes terdiri atas kitin sebagai komponen utama penyusunnya (Yurnaliza, 2001). Kitin pada fungi berbentuk mikrofibril dengan panjang yang berbeda tergantung pada spesies dan lokasi selnya (Rajarathnam et al., 1998).
Hasil uji antagonisme antara bakteri kitinolitik terhadap 4 isolat fungi tersebut berbeda. Isolat BK15 memiliki kemampuan tertinggi menghambat miselium Fusarium sp. 4 dengan zona hambat pada hari kesembilan inkubasi dengan diameter sebesar 2,18 cm sedangkan isolat BK09 memiliki zona hambat terendah terhadap miselium Fusarium sp. 4 dengan diameter sebesar 0,25 cm. Zona hambat tersebut menunjukkan kemampuan bakteri kitinolitik membentuk zona bening di sekitar koloni fungi. Menurut Fakhrullah (2008), isolat bakteri kitinolitik dengan kode BK07, BK08, BK09, dan Tgk 20 memiliki kemampuan menghambat miselium fungi Ganoderma boninense dan Fusarium oxysporum.
Struktur dinding sel fungi tidak hanya terdiri dari kitin, namun adanya β -1,3-glukan yang berikatan dengan kitin dalam satu matriks amorf yang merupakan stuktur penyusun dinding sel jamur juga. Pengendalian hayati jamur dengan menggunakan bakteri kitinolitik didasarkan pada kemampuan mikroorganisme untuk menghasilkan kitinase. Dinding sel Fusarium sp. tersusun atas 39% kitin, 29% glukan, 7% protein dan 6% lemak, Kandungan kitin pada dinding sel jamur Fusarium sp. ini akan memicu pembentukan enzim degradatif (Webster & Weber, 2007). Kitin merupakan komponen struktural dari sebagian besar dinding sel cendawan (Singh et al., 1999). Kitinase menghidrolisis kitin, sedangkan β-1,3-glukanase menghidrolisis β -1,3-glukan, keduanya dapat melisiskan sel jamur (EI-Katatny et al, 2000). Beberapa bakteri tanah seperti: Pseudomonas (Wang et al., 1997), Streptomyces (Okazaki et al., 1995, Tsujibo et al., 1995), Arthrobacter (Okazaki et al., 1999), Aeromonas (Ueda et al., 1996), Enterobacter (Chernin et al., 1995), dilaporkan memiliki aktivitas kitinolitik, yakni mampu menguraikan kitin.
Peranan kitinase dalam pertahanan tanaman terhadap serangan patogen terjadi melalui dua tahap, yaitu: (a), menghambat pertumbuhan cendawan dengan secara langsung menghidrolisis dinding miselia cendawan dan (b) melalui pengelupasan elisitor endogen oleh aktivitas kitinase yang kemudian memicu reaksi ketahanan sistemik pada inang. Bakteri kitinolitik sering kali menghasilkan berbagai kitinase,
walaupun degradasinya belum diketahui secara menyeluruh (Oku, 1994). Koloidal
Aeromonas caviae (Inbar & Chet, 1991), E. agglomerans (Chernin et al., 1998) dan B. cereus (Pleban et al., 1997). Bakteri dapat menghidrolisis kitin setelah 72-96 jam yang ditumbuhkan pada media campuran agar minimal yang dicampur dengan koloidal kitin sebagai sumber karbon (Chernin et al., 1995).
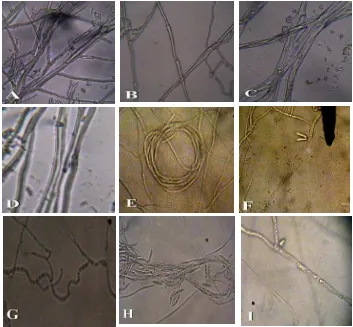
Gambar 3. Uji antagonis bakteri kitinolitik terhadap beberapa Fusarium sp. pada hari ke- 9. A. Zona hambat bakteri PP01 sp. 2 terhadap Fusarium sp. 1. B. Zona hambat bakteri KM013 sp. 2 terhadap Fusarium sp. 2, C. Zona hambat bakteri KM013 sp. 3 terhadap Fusarium sp. 3, D. Zona hambat bakteri BB01 sp. 2 terhadap Fusarium sp. 4, a.Koloni bakteri kitinolitik, b. koloni fungi patogen, c. zona hidrolisis kitin.
3. 3. Perbandingan Hifa Normal dengan Hifa Abnormal Fusarium sp.
Adanya senyawa metabolit lainnya selain kitinase juga biasa menjadi salah
satu faktor perbedaan daya hambat isolat kitinolitik terhadap pertumbuhan miselium
fungi patogen tanaman. Mekanisme kerja dari agen pengendalian hayati yang bersifat
antifungal pada umumnya dalam menghambat pertumbuhan (antagonis) terhadap
pertumbuhan miselium jamur patogen digolongkan sebagai persaingan zat makanan,
parasitisme, dan antibiosis (Johansson, 2003). Kitinase merupakan sekelompok enzim
yang umumnya dihasilkan oleh bakteri, disamping kitinase dalam sistem kitinolitik
mikroorganisme terdapat protein yang memiliki kemampuan antifungal. Protein
tersebut adalah protein pengikat kitin yang dihasilkan selama proses perombakan
kitin. Menurut Yurnaliza (2001), kehadiran protein pengikat kitin dalam system
kitinolitik S. olivaceoviridis mengefisiensikan peran enzim kitinase dari S.
olivaceoviridis dalam mendegradasi kitin dari dinding sel fungi.
BAB 4
KESIMPULAN DAN SARAN
4.1 Kesimpulan
Dari hasil penelitian tentang kemampuan bakteri kitinolitik dalam menghambat pertumbuhan Fusarium sp. tanaman kopi dapat diambil kesimpulan sebagai berikut: 1. Diperoleh sebanyak 4 jenis Fusarium yang diduga patogen pada tanaman kopi
yaitu, Fusarium sp. 1, Fusarium sp. 2, Fusarium sp. 3 dan Fusarium sp. 4 yang memiliki ciri-ciri berbeda.
2. Kemampuan tertinggi dalam menghambat pertumbuhan koloni fungi adalah isolat BK15 dengan diameter zona hambat sebesar 2,18 cm sedangkan isolat BK09 memiliki kemampuan terendah dalam menghambat pertumbuhan fungi dengan diameter zona hambat sebesar 0,25 cm.
3. Pengamatan hifa Fusarium sp. setelah uji antagonis mengalami keabnormalan antara lain, hifa menggulung, melilit, keriting, lisis dan patah.
4.2 Saran
DAFTAR PUSTAKA
Agrios, G. N. 1988. Plant Pathology. Professor and Chairman Dept. of Plant Pathology University of Florida. Academic Press Inc. Guinesville. hlm: 359. Brown, J. & Ogle, H. 1997. Fungal Disease and Their Control. In J. Brown and H.
Ogle (eds.). Plant Pathogens and Plant Diseases. The University of New England Printery, Armidal.
Burkhead, K. D., Schisler, D. A., & Slininger, P. J. 1994. Pyrrolnitrin Production by Biological Control Agent Pseudomonas cepacia B37w in Culture and in Colonized Wounds of Potatoes. Appl. Environ. Microbiol. 60 (6): 2031-2039.
Chernin, L. S., Michael K. W., Jacquelyn M. T., Shoshan H., Barrie W. B., Cheat. W., & Gordon S. A. B., Stewart. 1998. Chitinolytic Activity in Chromobacterium violaceum. J. Bacteriol. 65: 25-37.
Danarti, S & Najiyati. 1999. Budidaya Tanaman Kopi. Jakarta. Penebar Swadaya. hlm. 29.
Datta, K., Z.K. Nicola, N. Baisakh, N. Oliva & S.K. Datta (2000). Agrobacterium mediated engineering for sheath blight resistance of indica rice cultivars from different ecosystems. Theor. Appl. Genet. 100: 832-839.
Djafaruddin. 2008. Dasar-dasar Pengendalian Penyakit Tanaman. Edisi Pertama. Cetakan keempat. Jakarta. Bumi Aksara. hlm. 43-49.
El-katatny MH. Somicth W, Robra K-H,El-katatny MS, & Gubizt GM. 2000. Production of Chitinase dan β-1,3-glukanase by Tricoderma harzyanum for Control of The Phytophatogenic Fungus Scelerotium rolfsii. Food Technol. Biotechnol. 38: 173-180.
Fakhrullah, 2008. Identifikasi Bakteri Kitinolitik dari Tanah Hutan Tangkahan Taman Nasional Gunung Leuser dan Sifat Antagonisnya Terhadap Beberapa Jamur Patogen Tanaman. Skripsi. USU.
Gandjar, I., Robert, A. S., Karin, T., Oetari, A. & Santoso, I. 1999. Pengenalan Kapang Tropik Umum. Yayasan Obor Indoneia. Depok. hlm. 66.
Gohel, V., Singh, A., Vimal, M., Ashwini, D &Chatpar, H. S. 2003. Bioprospecting and Antifungal Potential of Chitinolytic Microorganism. African J. Biotechnol. 5: 54-72.
Hebbar, K. P., Atkinson, D., Tucker, W., & Dart, P. J. 1992. Suppression of Fusarium moniliforme by Maize Root Associated Pseudomonas cepacia. Soil Biol. Biochem. 24 (10): 1009-1020.
Inbar, J. & I. Chet. 1991. Evidence That Chitinase Produced by Aeromonas caviae is Involved in The Biological Control of Soil-Borne Plant Pathogens by The Bacterium Soil Biol. Biochem. 23: 973-978.
Istikorini, Y. 2002. Pengendalian Penyakit Tumbuhan secara Hayati yang Ekologis dan Berkelanjutan. (Online). http://tumoutou.net/702_05123/yunik_istikori. htm. Diakses tanggal 23 Juni2009.
Johansson, P. M. 2003. Biocontrol of Fusarium in Wheat-Introducing bacteria to a System of Complex Interactions. Thesis Doctoral. Plant Pathology and Biocontrol Unit. Uppsala, Swedia: Swedish University of Agricultural Scienses.
King, E. B. & Parke, J. L. 1993. Biocontrol of Aphanomyces Root Rot and Pythium Damping-off by Pseudomonas cepacia AMMD of Four Pea Cultivars. Plant Disease. 77: 1185-1188.
Mahfud, M.C., E. Korlina, A. Budijono, M, Soleh & A. Surjadi. 1998. Uji Aplikasi Datta, K., Z.K. Nicola, N. Baisakh, N. Oliva & S.K. Datta 2000. Agrobacterium mediated engineering for sheath blight resistance of indica rice cultivars from different ecosystems. Theor. Appl. Genet. 100: 832-839.
Martorejo, T., Sumardiyono, C. & Astuti, E. H. 2001. Kajian Pengendalian Hayati Penyakit Kapang Hijau pada Buah jeruk dengan Trichoderma sp. Proseding Seminar PFI. IPB. Bogor. hlm. 354-356.
Miller, A. S., Randall C. R. & Richard M. R. 1986. Fusarium and Verticillium Wilts of Tomato, Potato, Pepper, and Eggplant.The Ohio State University.
Nigam, N & Mukerji K,G. 1988. Biological control-concept and practice. Florida .CRC Press. hlm. 52.
Nugroho, T. J., Muhammad, A., Cipta, G., Wahyuningsih, Andi, D., Silvera, D., Yessy, S. 2003. Isolasi dan Karakterisasi sebagian Kitinase Trichoderma viridae TNJ63. J. Natur Indones. 5: 101-106.
Okazaki, K., F. Kato, N. Watanabe, S. Yasuda, Y. Masui & S. Hayakawa. 1995. Purification and properties of two chitinases from Streptomyces sp. J-13-3. Biosci. Biotech. Biochem. 59: 1586-1587.
Okeniyi, M. O. 2007. The Current Status of Vaskular wilt disease of Coffee Tracheomycosis in Nigeria. African Crop Science Conference Proceedings. 8: 833-834.
Patonah, S. 2008. Penggunaan Bakteri Kitinolitik untuk Menghambat Penyakit Layu Fusarium pada Benih Cabai Merah (Capsicum annuum L.). Skripsi. USU. Medan. hlm. 2.
Pitt, J. I & Hocking, A. D. 1997. Fungi and Food Spoilage. Second edition. Cambridge. Great Britain at the University Press.
Pleban, S., L. Chernin, & I. Chet. 1997. Chitinolytic Activity of Endophitic Strain of Bacillus cereus. Lett. Appl. Microbiol. 25: 284-288.
Pudjihartati, E., Siswanto, Satrias, I. & Sudarsono. 2006. Aktivitas Enzim Kitinase pada Kacang Tanah yang Sehat dan yang Terinfeksi Sclerotium rolfsii. Unit Penelitian Bioteknologi Perkebunan. Bogor. hlm. 37.
Rajarathanam, S., Shashriekha, M. N. J. & Bano, Z. 1998. Biodegradative and Biosynthetic Capacities of Mushrooms. Present and Future Strategies. Crit. Rev. Biotech. 18: 91-23.
Rao, S. N. 1994. Mikroorganisme Tanah dan Pertumbuhan Tanaman. Edisi kedua. Jakarta. Universitas Indonesia. hlm. 13.
Semangun, H. 1996. Pengantar Ilmu Penyakit Tumbuhan. Yogyakarta. Gadjah Mada University Press. hlm. 49.
, H. 2000. Penyakit-penyakit Tanaman Hortikultura di Indonesia. Gadjah Mada University Press. Yogyakarta. hlm. 59.
, H. 2007. Penyakit-penyakit Tanaman Hortikultura di Indonesia. Gadjah Mada University Press. Yogyakarta. hlm. 97.
Singh, P., Shin, Y.C., Park, C. S. & Chung, Y. R. 1999. Biological Control of Fusarium Wilt of Cucumber by Chitinolytic Bacteria. The American Phytopatholl. Soc. 89: 92-99.
Siswoputranto, P. S. 1978. Perkembangan Teh, Kopi, Coklat Internasional. Jakarta: Gramedia. hlm. 149-150.
Soesanto, L. 2008. Pengantar Pengendalian Hayati Penyakit Tanaman. Edisi Pertama Jakarta. PT RajaGrafindo Persada. hlm. 167.
Suryanto D., Patonah, S., Munir E., 2010. Control of Fusarium Wilt of Chili With
Chitinolytic Bacteria. Hayati Journal of Bioscienses. 17: 5-8.
Suryanto, D & Munir E. 2006. Potensi Pemanfaatan Isolat Bakteri Kitinolitik untuk Pengendalian Hayati Jamur. Prosiding seminar hasil-hasil penelitian USU. Medan. hlm. 15-25.
Ekstrak Botani. Makalah Falsafah Sains Program Pasca Sarjana/S3. IPB. Bogor.
Tsujibo, H., H. Endo, K. Miyamoto & Y. Inamori. 1995. Expression in E. coli of a gene encoding a thermostable chitinase from Streptomyces thermoviolaceus OPC-520. Bioscience, Biotechnology, and Biochemistry. 59: 145-146.
Ueda, M., M. Shiro, T. Kawaguchi &M. Arai. 1996. Expression of chitinase III gen of Aeromonas 10S-24 in E. coli. Bioscience, Biotechnology, and Biochemistry. 60: 1195-1197.
Upadhyay, R. S. & Jayaswal, R. K. 1992. Pseudomonas cepacia Causes Mycelial Deformities and Inhibition of Conidiation in Phythopathogenic Fungi. Curr. Microbiol. 24 (4): 181-187.
Wang, S.L., S.H. Chiou & W.T. Chang. 1997. Production of chitinase from shellfish waste by Pseudomonas aeruginosa K-187. Proceeding of the National Science Council of R.O.C. 21: 71-78.
Wang, S., Wu J., Rao P., Ng TB & Ye X. 2005. A Chitinase with Antifungal Activity from the Mung bean. Protein. Expr. Pufif. 40: 230-236.
Webster, J. & Weber, R. W. S. 2007. Introduction of Fungi. New York. Cambridge University Press.
Yurnaliza, 2001. Kajian Peran Aktinomisetes Khitinolitik dalam Pengendalian Jamur Patogen Fusarium oxysporum Skala Laboratorium. Tesis. Pasca Sarjana/S2. Yogyakarta. Indonesia: Universitas Gadjah Mada.
Yusriadi. 2004. Pengendalian Biologi (Biocontrol) Penyakit Tular tanah Kacang tanah dengan Pseudomonas (Ralstonia) fluorescens BSK8. Jurnal Kalimantan Scientiae 64 (12): 78-84.
Lampiran 1: Pembuatan koloidal khitin dengan cara hidrolisis parsial (Rodriquez-Kabana et al., 1983 ).
Dihaluskan
Dilarutkan dalam 180 ml 10N HCl sambil diaduk dengan pengaduk magnetik selama 1,5-2 jam.
Dituang ke dalam erlenmeyer yang berisi 2 liter air.
Dibiarkan semalam kemudian bagian yang bening dipisahkan dari endapannya.
Dicuci sampai pH suspensi khitin antara 5,5-6
Diambil 10 ml kemudian dikeringkan pada suhu 80oC. Dihitung berat keringnya.
20 g Crab shell chitin
Suspensi Kitin
Hasil
Lampiran 2 . Komposisi Medium MGMC Padat
K2HPO4 0,7 g
KH2PO4 0,3 g
MgSO4.7H2O 0,5 g
FeSO4.7H2O 0,01 g
ZnSO4 0,001g
MnCl2 0,001g
Koloidal kitin 12,5% (h/v) 72,7 ml
Agar 20 g
pH 6,8
Cara Pembuatan
Lampiran 3. Foto Tanaman Kopi yang Terserang Fusarium
Lampiran 3. Foto Isolasi Awal
Keterangan : A. Ranting Bagian Pucuk Kopi B. Daun isolat awal 1 Kopi C. Daun isolat awal 2 Kopi D. Buah Kopi
A B