RESPONS SUPEROVULASI DENGAN
PENYUNTIKAN TUNGGAL FSH DALAM RUANG
EPIDURAL BERBASIS PERTUMBUHAN GELOMBANG
FOLIKEL PADA SAPI PERANAKAN ONGOLE
MUHAMMAD IMRON
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa disertasi berjudul Respons Superovulasi Dengan Teknik Penyuntikan Tunggal Hormon FSH dalam Ruang Epidural pada Sapi Peranakan Ongole adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
MUHAMMAD IMRON. Respons Superovulasi dengan Penyuntikan Tunggal FSH dalam Ruang Epidural Berbasis Pertumbuhan Gelombang Folikel pada Sapi Peranakan Ongole. Dibimbing oleh MOHAMAD AGUS SETIADI, IMAN SUPRIATNA dan AMROZI.
Serangkaian penelitian dilakukan untuk menguji respons superovulasi dengan teknik penyuntikan tunggal hormon FSH dalam ruang epidural berdasarkan pada pertumbuhan gelombang folikel pada sapi peranakan ongole. Penelitian tahap pertama dilakukan untuk mendapat informasi dinamika ovarium pada sapi PO sebagai respon terhadap penyuntikan gonadorelin (GnRH) dalam protokol Ovsynch berbasis preparat progesteron intravaginal. Hasil penelitian ini digunakan untuk protokol sinkronisasi ovulasi pada dua penelitian tahap berikutnya. Sapi dara (n=18) dan induk (n=12) dibagi secara acak menjadi tiga group. Group satu (CP) diberi perlakuan dengan memasukkan Cuemate kedalam vagina pada hari ke 0 – 7 dan disuntik prostaglandin pada hari ke-7. Kelompok dua (CPG) sama dengan kelompok satu, tapi dengan penambahan penyuntikan GnRH pada hari ke-9. Kelompok tiga (GCPGP) sama dengan kelompok dua, tapi dengan diawali penyuntikan GnRH pada hari ke-0. Pemeriksaan USG dilakukan pada hari ke 0 – 4, 7 sampai ovulasi dan 7 hari setelah ovulasi. Persentase sinkronisasi ovulasi meningkat sangat nyata berturut-turut pada CP, CPG and GCPG pada dara (16, 50 dan 85 %) maupun induk (0, 60 dan 100 %), P <0.01 pada hari ke-11. Diameter folikel ovulatori tidak berbeda nyata antara perlakuan CP, CPG dan GCPG pada dara (11.9+0.5, 11.9+0.5 dan 12.1+0.6 mm) maupun pada induk (11.7+0.4, 11.8+0.7, 11.1+0.6 mm). Penelitian ini menyimpulkan bahwa protokol GCPG meningkatkan persentase sinkronisasi ovulasi baik pada dara maupun induk tanpa mempengaruhi diameter folikel ovulatori maupun korpus luteum.
menggunakan 6 ekor sapi PO yang sudah diketetahui pola gelombangnya pada satu IOI berikutnya. Hasil pengamatan pertumbuhan gelombang dan jumlah folikel dalam penelitian ini memiliki nilai repeatabilitas yang tinggi pada individu yang sama, yaitu beruturut-turut 0,88 dan 0,93.
Penelitian ketiga dilakukan berdasarkan metode sinkronisasi ovulasi paling optimal dari hasil penelitian tahap satu dan data dinamika pertumbuhan folikel pada penelitian tahap dua untuk melakukan superovulasi pada sapi PO. Pelaksanaan superovulasi secara konvensional umumnya dilakukan dengan penyuntikan Follicle Stimulating Hormone (FSH) dua kali sehari selama 4 hari sehingga memerlukan alokasi petugas teknis secara khusus dalam pelaksanaannya. Tujuan penelitian tahap tiga adalah untuk mengkaji respon superovulasi menggunakan penyuntikan dosis tunggal hormon FSH ke dalam ruang epidural pada sapi Peranakan Ongole (PO). Penelitian tahap tiga terdiri dari 2 bagian, yaitu penelitian 1 dan 2. Penelitian 1 membandingkan perlakuan kombinasi penyuntikan tunggal hormon FSH ke dalam ruang epidural plus intramuskular (perlakuan kombinasi epi+i.m.) dengan penyuntikan FSH dua kali sehari secara intramuskular selama 4 hari (perlakuan intramuskular), menggunakan dosis total masing-masing 400 mg FSH. Respon superovulasi dari kelompok perlakuan epi+i.m (n=4) tidak berbeda nyata dengan kelompok perlakuan intramuskular (n=4) pada semua hasil pengamatan. Penelitian 2 membandingkan penggunaan dua konsentrasi FSH (280 mg dan 160 mg) yang disuntikkan dalam dosis tunggal ke dalam ruang epidural. Sebagai kontrol digunakan data perlakuan epi+im dari penelitian 1. Kelompok perlakuan FSH 280 mg (n=4) menghasilkan total koleksi dan embrio layak transfer (9,0+2,65 dan 3,33+2,52), berbeda nyata dengan kelompok perlakuan 160 mg (n=4) (2,0+1,26 dan 0,00), P<0,05, tetapi tidak berbeda nyata dengan kontrol (9,33+5,68 dan 3,67+3,21). Disimpulkan bahwa supersovulasi dengan penyuntikan FSH dengan dosis tunggal ke dalam ruang epidural menghasilkan embrio layak transfer yang setara dengan penyuntikan dua kali sehari berturut-turut selama 4 hari. Kata kunci: Sapi PO, Superovulasi, penyuntikan tunggal, ruang epidural
SUMMARY
MUHAMMAD IMRON. Superovulatori Respons of single epidural injection of FSH based on follicular wave emergence in Peranakan Ongole Cattle. Supervised by MOHAMAD AGUS SETIADI, IMAN SUPRIATNA and AMROZI.
ovulation. Percentage of ovulation synchronization increased significantly between CP, CPG and GCPG respectively both in heifer (16, 50 and 85, respectively) and cow (0, 60 and 100 %, respectively), P<0.01 at day 11. Preovulatori follicle diameter between CP, CPG and GCPG treatments were not different significantly both in heifer (11.9+0.5, 11.9+0.5 and 12.1+0.6 mm, respectively) and cow (11.7+0.4, 11.8+0.7, 11.1+0.6 mm, respectively). This study concluded that GCPG protocol increased the synchrony of ovulation rate both in cows and heifer, without affected the follicle preovulatory and CL diameter.
The second study was conducted to investigate the ovarian dynamics and its repeatability during the estrous cycle in PO cattle. Follicular growth and atresia during estrous cycle were evaluated using a portable ultrasound device. In the early study, ovarian dynamics observations conducted on 9 PO cows trough ultrasound scanning once every single day to map the patterns of follicular wave development for one interovulatori interval ( IOI ). The result of observations indicated that PO cattle had 3 (66%) and 4(34%) follicular waves pattern. The first wave of 3 and 4 wave pattern emerged on day -0,4+0,9 and 1,4+1,1 respectively, relative to ovulation occurance. The second wave of 3 and 4 wave pattern emerged on day 9,8+1,5 and 7,4+1,9 respectively, relative to ovulation occurance. The pattern of three follicular wave has a longer follicle dominance duration (11,6+1,5 day) in the first wave of estrous cycle, compared with 4 wave pattern (10+2,92 and 7+1.00 day respectively). The growth rate of dominant follicle (FD) were not different significantly between the patterns of 3 and 4 follicular waves (0,87+0,23 and 0,25+ 0.94 mm/day). The growth rate of FD candidate also were not different significantly compared with its subdominant follicles (FS) until deviation stage occurred. Similarly, ovulatory follicle diameter between 3 and 4 waves patterns also not significant different, 12,24+12,34 and 12,30+12,23 mm respectively. Diameter CL also did not differ significantly between 3 and 4 follicular wave pattern, 18,94+0,47 and 19,44+0,87 mm, respectively. Observations of wave patterns repeatability were performed using 6 PO cows which already known for its wave pattern on the next consecutive IOI. The result of this study indicate that follicular wave pattern and number of emergence follicles has high repeatability value in the same individual, 0,88 and 0,91 respectively.
0,00) P <0,05, but not significantly different compared to control (9,33+5,68 and 3,67+3,21). In summary, injection of single dose of FSH into epidural space resulted in a comparable transferable embryo to the traditional twice daily intramuscular injection of FSH over 4 days.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
RESPONS SUPEROVULASI DENGAN
PENYUNTIKAN TUNGGAL FSH DALAM RUANG
EPIDURAL BERBASIS PERTUMBUHAN GELOMBANG
FOLIKEL PADA SAPI PERANAKAN ONGOLE
MUHAMMAD IMRON
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada Program Studi Biologi Reproduksi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup: Prof drh Bambang Purwantara, M.Sc, Ph.D Dr Ir Syahruddin Said, M.Agr.Sc
Judul Disertasi : Respons Superovulasi dengan Teknik Penyuntikan Tunggal Hormon FSH dalam Ruang Epidural Berbasis Pertumbuhan Gelombang Folikel pada Sapi Peranakan Ongole
Nama : Muhammad Imron NIM : B 362120051
Disetujui oleh Komisi Pembimbing
Prof Dr drh Mohamad Agus Setiadi Ketua
Prof Dr drh Iman Supriatna Anggota
Dr drh Amrozi Anggota Diketahui oleh
Ketua Program Studi Biologi Reproduksi
Prof Dr drh Mohamad Agus Setiadi
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
Tanggal Ujian: 4 Pebruari 2016
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga penulis mampu menyelesaikan disertasi ini dengan judul “Respons superovulasi dengan penyuntikan tunggal FSH dalam ruang epidural berbasis pertumbuhan gelombang folikel pada sapi Peranakan Ongole”.
Terima kasih dan penghargaan yang tak terhingga penulis ucapkan kepada Prof Dr Mohamad Agus Setiadi baik selaku Ketua Komisi Pembimbing maupun sebagai Ketua Program Studi Biologi Reproduksi, Prof Dr drh Iman Supriatna dan Dr drh Amrozi selaku anggota komisi pembimbing yang telah banyak meluangkan waktu memberikan koreksi dan saran mulai dari penyusunan proposal, perencanaan dan pelaksanaan penelitian serta penyusunan disertasi ini.
Penulis mengucapkan terima kasih dan penghargaan kepada Prof drh Bambang Purwantara, M.Sc, Ph.D dan Dr Ir Syahruddin Said, M.Agr.Sc sebagai penguji luar komisi yang telah banyak memberikan masukan untuk meningkatkan kualitas penulisan disertasi ini. Penghargaan juga penulis sampaikan kepada seluruh staf pengajar dan karyawan Program Studi Biologi Reproduksi atas bantuan dan dukungannya selama penulis menempuh pendidikan S3. Terima kasih juga kepada Teh Yanti yang telah banyak memberikan pelayanan administrasi di Program Studi BRP dengan sangat baik.
Ucapan terima kasih juga penulis sampaikan kepada Kementerian Pertanian Republik Indonesia, cq. Kepala Pusat Pendidikan, Standardisasi, dan Sertifikasi Profesi Pertanian, Badan Penyuluhan dan Pengembangan Sumberdaya Manusia Pertanian, SDM Pertanian yang telah memberikan kesempatan Tugas Belajar Program Doktor Dalam Negeri kepada penulis untuk melanjutkan studi S3 di IPB. Penulis ucapkan terima kasih kepada Kepala BET Cipelang yang banyak memberikan fasilitas untuk penelitian dan seluruh rekan kerja di BET Cipelang yang banyak membantu pelaksanaan kegiatan penelitian di lapangan.
Terima kasih juga penulis sampaikan kepada teman-teman S3 seperjuangan, Pak Soni, Pak Hasbi, Bu Sri dan Bang Saor, yang banyak memberikan warna dan menghabiskan waktu bersama baik offline maupun online selama menempuh studi di IPB. Tanpa kalian, mungkin studi ini berasa seperti sayur kurang garam.
Akhirnya, penulis mengucapkan terima kasih yang tak terhingga dan setulus-tulusnya kepada Ramanda (alm.) Rosyidi dan ibunda Hj. Farida yang telah memberikan seluruh usahanya untuk memberikan dasar pendidikan terbaik untuk penulis. Terima kasih kepada kedua kakakku, Iyu Hj. Raudah Srisuhastiningsih dan Mbak dra. Mudafiatun Isriah, S.Pd, M.Pd serta seluruh keluarga besar di Lumajang, atas segala doa dan kasih sayangnya. Terima kasih juga untuk keluarga kecilku tercinta di Bogor, istriku Erni Supriatna, S.Si dan anak-anakku Hibban Syafie Rosyidi dan Athaya Nayla Zahra atas cinta, pengertian dan pengorbanan waktunya yang banyak tersita selama penulis melaksanakan pendidikan di IPB. Demi kalianlah Abi saat ini mampu berdiri didepan forum terhormat ini. Akhirya, semoga segala apa yang sudah penulis kerjakan dapat menjadi sumbangsih pengembangan ilmu pengetahuan di Indonesia.
DAFTAR ISI
RINGKASAN iv
DAFTAR ISI xiii
DAFTAR TABEL xiv
DAFTAR GAMBAR xiv
DAFTAR LAMPIRAN xv
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 3
Tujuan Penelitian 5
Manfaat Penelitian 5
2 DINAMIKA OVARIA PADA SAPI PERANAKAN ONGOLE (PO) SETELAH PENYUNTIKAN GnRH DALAM PROTOKOL OVSYNCH
BERBASIS PREPARAT PROGESTERON 7
Pendahuluan 7
Materi dan Metode 8
Hasil dan Pembahasan 10
Simpulan dan Saran 17
Daftar Pustaka 17
3 DINAMIKA OVARIA DAN REPEATABILITAS PERTUMBUHAN
GELOMBANG FOLIKEL PADA SAPI PERANAKAN ONGOLE (PO) 21
Pendahuluan 21
Materi dan Metoda 22
Hasil dan Pembahasan 23
Simpulan dan Saran 29
Daftar Pustaka 29
4 RESPON SUPEROVULASI SAPI PERANAKAN ONGOLE MELALUI PENYUNTIKAN TUNGGAL FSH KE DALAM RUANG EPIDURAL 35
Pendahuluan 35
Materi dan Metoda 36
Hasil dan Pembahasan 39
Simpulan dan Saran 48
Daftar Pustaka 48
5 PEMBAHASAN UMUM 53
Simpulan dan Saran 56
Daftar Pustaka 57
DAFTAR TABEL
1 Dinamika ovaria pada kelompok dengan perlakuan penyuntikan GnRH pertama (Perlakuan GCPG) dan tanpa penyuntikan GnRH pertama
(perlakuan CP dan CPG) pada dara dan induk 11
2 Pengamatan tanda-tanda berahi dan dinamika ovaria pada sapi PO dara dan induk yang diberi perlakuan CP, CPG dan GCPG 15 3 Karakteristik dinamika folikel dengan pola 3 dan 4 gelombang folikel
pada satu IOI 25
4 Karakteristik dinamika folikel dan corpus luteum (CL) pada sapi PO
yang memiliki 3 dan 4 gelombang folikel 26
5 Nilai beberapa karakteristik pengamatan dinamika folikel pada sapi PO
28 6 Respon superovulasi menggunakan perlakuan penyuntikan FSH
berturut-turut selama 4 hari sebanyak dua kali sehari secara i.m. (perlakuan intramuskular) dan penyuntikan tunggal FSH via epidural
plus intramuskular (perlakuan epi+i.m.) 40
7 Respon superovulasi dengan metode epi 70%, epi 40% dan kombinasi
epi+i.m (kontrol) pada sapi PO 45
8 Jumlah sapi yang mengalami ovulasi setelah penyuntikan prostaglandin dalam perlakuan CP, CPG dan GCPG pada sapi PO
induk dan dara 53
DAFTAR GAMBAR
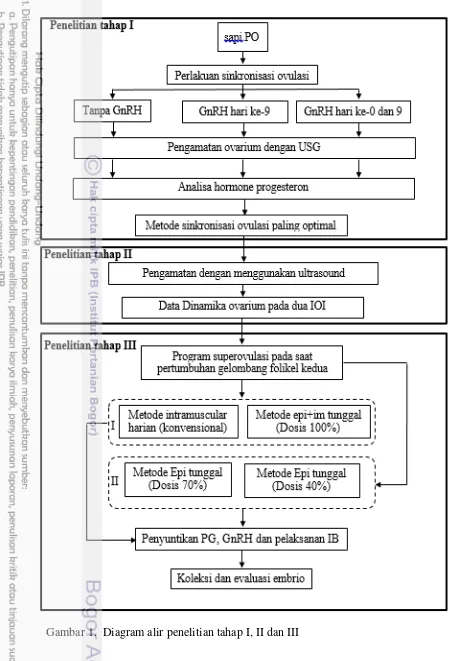
1 Alur penelitian tahap 1 4
2 Skema jenis perlakuan pada kelompok CP, CPG and GCPG. PGF2α (PG) diberikan sesaat setelah pelepasan Cuemate 9 3 Perubahan rataan diameter folikel dominan mulai hari ke-0 sampai 10
pada kelompok ovulasi dan perlakuan CP, CPG, GCPG 12 4 Distribusi persentase ovulasi setelah penyuntikan PGF2α dengan
interval waktu tertentu pada perlakuan CP, CPG dan GCPG untuk sapi
dara (A) dan sapi induk (B) 13
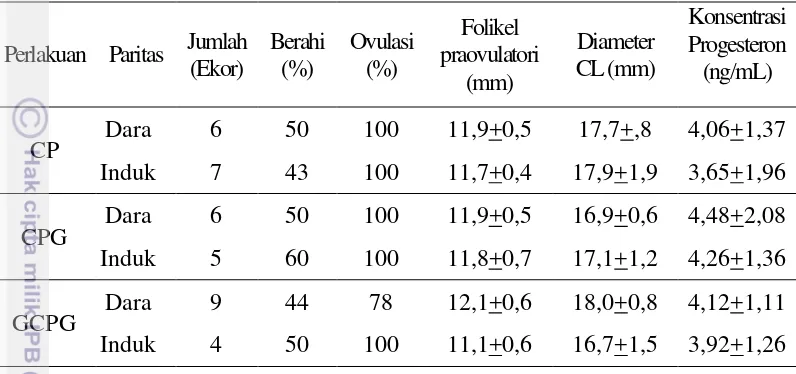
5 Pola perkembangan gelombang folikel dan CL pada sapi PO 24 6 Skema umum metode superovulasi dan koleksi embrio 37 7 Beberapa fase perkembangan dan kualitas embrio pada sapi PO 41 8 Pengamatan dinamika ovaria dengan menggunakan USG pada sapi PO
yang diberi perlakuan kombinasi epi+i.m. 42
9 Respon dinamika ovaria pada sapi PO dengan perlakuan epi 70%, epi
40% dan kombinasi epi+i.m. (kontrol) 43
regresi (H8). Terlihat ada CL assesories. F. Korpus luteum tunggal
(H11) 46
11 Saran untuk protokol IB fixed time pada sapi PO dengan menggunakan
protokol sinkronisasi ovulasi GCPG 54
DAFTAR LAMPIRAN
1 Sertifikat persetujuan etik hewan 61
Latar Belakang
Sapi Ongol merupakan sapi asli dari daerah Andra Prades, pesisir India. Sapi ini di India berfungsi sebagai sapi potong dan juga memproduksi susu dimana pada puncak laktasinya dapat menghasilkan 8-10 liter susu/hari (Gaur et al. 2002). Sebagai sapi asli yang berkembang di wilayah tropis, sapi Ongol tahan terhadap temperatur tropis yang panas, tahan terhadap parasit eksternal dan serangga, menunjukkan respon baik pada pakan yang berkualitas namun juga tahan terhadap pakan berkualitas rendah di lapangan (Behl et al. 2010). Secara umum sapi ongol berwarna putih, dan terdapat sedikit warna abu-abu pada bagian punggung dan belakang (Gaur et al. 2002).
Sapi Ongol diintroduksi pertama kali ke Indonesia pada tahun 1906 dan mulai dikembangbiakkan di pulau Sumba, yang kemudian dikenal dengan nama sapi Sumba Ongol (SO). Selanjutnya mulai tahun 1915 keturunan sapi ini disebarkan ke beberapa wilayah Indonesia bagian barat, terutama di pulau Jawa. Kebijakan Ongolisasi diterapkan oleh pemerintah Belanda pada tahun 1936 yang menyatakan bahwa sapi Jawa betina harus dikawinkan dengan pejantan SO dan sapi Jawa jantan harus dikebiri (Hardjosubroto 2002). Hasil persilangan antara sapi Jawa yang kecil dengan sapi Ongol yang lebih besar menghasilkan sapi dengan ukuran yang relatif lebih besar dibandingkan ukuran sapi Jawa, berwarna putih dan berpunuk atau yang saat ini dikenal dengan sapi Perananakan Ongol (PO) (Diwyanto dan Inounu 2009). Saat ini sapi PO sudah berkembang secara alami menjadi suatu bangsa sapi yang stabil dengan karakteristik morfologi yang mudah dikenali. Sapi PO juga menunjukkan keunggulan sapi tropis yaitu daya adaptasi iklim tropis yang tinggi, tahan terhadap panas, tahan terhadap gangguan parasit seperti gigitan nyamuk dan caplak, disamping itu juga menunjukkan toleransi yang baik terhadap pakan yang mengandung serat kasar tinggi (Astuti 2004). Pada beberapa daerah, sapi PO dipelihara dengan tujuan ganda disamping sebagai sapi potong juga untuk sapi kerja.
Pada penelitian yang lain, Matondang dan Rusdiana (2013) juga melaporkan masalah yang sering muncul dalam usaha ternak sapi PO adalah kawin berulang (S/C > 2) dan rendahnya angka kebuntingan (< 60%) sehingga jarak beranak menjadi panjang (> 18 bulan). Sapi PO memiliki masa tidak bunting panjang salah satunya disebabkan oleh kesulitan deteksi berahi karena sapi PO sering tidak menunjukkan tanda-tanda berahi yang jelas sehingga pelaksanaan IB menjadi tidak tepat yang mengakibatkan interval beranak menjadi panjang (Nuryadi dan Wahjuningsih 2011). Karena itulah diperlukan protokol sinkronisasi berahi/ovulasi yang tepat untuk mengoptimalkan pelaksanaan IB pada sapi PO tanpa harus memperhatikan tanda-tanda berahi.
2
(lokal) menjadi bangsa pejantannya (Putro 2009). Masyarakat menyukai upgrading dengan sapi bangsa B. taurus karena turunan hasil persilangannya memiliki kecepatan pertumbuhan yang lebih baik dibandingkan dengan sapi lokal. Demikian pula hal ini juga terjadi pada banyak sapi PO yang dikawin silang dengan sapi B. taurus terutama di pulau Jawa sehingga dilaporkan menekan populasi sapi PO (Astuti 2004). Dilain pihak, jika proporsi genetik sapi persilangan dari darah B. taurus semakin meningkat dibandingkan proporsi genetik sapi induk aslinya (lokal), dikhawatirkan akan terjadi penurunan daya adaptasinya terhadap iklim tropis yang panas. Selain itu peningkatan komposisi genetik sapi B. taurus juga mempengaruhi kemampuan reproduksi sapi terutama jika peternak tidak mampu menyediakan pakan atau manajemen yang sesuai dengan kebutuhan sapinya (Putro 2009). Untuk itu pemerintah berupaya untuk meningkatkan budidaya dan pembibitan sapi PO antara lain menggunakan teknologi superovulasi dan embrio transfer.
Tujuan utama dari program superovulasi adalah untuk mendapatkan jumlah embrio layak transfer yang maksimal dan memiliki kemungkinan yang tinggi untuk menghasilkan kebuntingan (Bo dan Mapletoft 2014). Umumnya program superovulasi dilakukan dengan penyuntikan hormon FSH dua kali sehari (metode konvensional) selama 4-5 hari (Mapletoft et al. 2012) untuk mempertahankan konsentrasi FSH yang cukup untuk memicu superstimulasi ovarium. Perlakuan ini kemudian diikuti dengan penyuntikan hormon PGF2α untuk memicu luteolisis dan dilanjutkan dengan IB setelah sapi menunjukkan tanda-tanda berahi. Penyuntikan FSH yang beruntun ini memerlukan alokasi penyediaan teknisi yang ditugaskan khusus dalam pelaksanaannya dan juga dapat menyebabkan peningkatan cekaman pada sapi donor.
Beberapa penelitian telah dilakukan untuk mengurangi frekuensi penyuntikan FSH antara lain dengan mencampur FSH dengan media pembawa dan disuntikkan dengan dosis tunggal secara intramuskular dan dilaporkan menghasilkan respon superovulasi yang sebanding dengan metode superovulasi konvensional (Suzuki et al. 1994, Kimura et al. 2007, Tribulo et al. 2012). Teknik lain yang dilaporkan menghasilkan respond superovulasi yaitu penyuntikan tunggal hormon FSH dibawah kulit (Hiraizumi et al. 2015). Keberhasilan teknik ini dipengaruhi oleh lokasi penyuntikan dan Nilai Kondisi Tubuh (NKT) sehingga diduga ketebalan lemak subkutan merupakan faktor yang mempengaruhi tingkat kecepatan penyerapan FSH kedalam peredaran darah.
Sejalan dengan dugaan ini, dalam bidang veteriner ruang epidural banyak digunakan untuk lokasi penyuntikan obat anastesi dengan tujuan memperpanjang durasi efeknya (Grubb et al. 2002). Ruang epidural adalah ruang diantara lapisan terluar dan terdalam medulla spinalis pada kanal vertebral yang dikelilingi oleh jaringan berlemak dalam bentuk semi liquid, limfatik, arteri dan beberapa jaringan lain (Moore & Dalley 2006). Diduga bahwa penggunaan ruang epidural sebagai posisi penyuntikan FSH dengan dosis tunggal dapat memicu respon superovulasi karena adanya jaringan berlemak didalamnya.
bahwa perlakuan superovulasi akan menghasilkan respon yang optimal jika dilakukan pada saat terjadi pertumbuhan folikel. Perlakuan gonadotropin yang diberikan sebelum atau setelah terjadinya gelombang pertumbuhan folikel akan mengurangi respon superovulasi secara signifikan (Adam et al. 2008). Untuk itu informasi tentang dinamika gelombang folikel pada sapi donor menjadi urgen untuk menghasilkan respon superovulasi yang lebih optimal.
Namun sampai saat ini masih sedikit informasi tentang dinamika gelombang pertumbuhan folikel terutama pada sapi PO. Bo et al. (2003) menyatakan fisiologi reproduksi sapi B. indicus tidak identik dengan B. taurus antara lain dalam hal panjang siklus berahi, waktu berahi, tingkah laku berahi, karakteristik folikel dominan dan korpus luteum (CL). Walaupun Adam et al. (2008) menyatakan mayoritas bangsa sapi menunjukkan pola dua dan tiga gelombang folikel dalam satu siklus berahi, peneliti lain juga melaporkan adanya pola satu, empat dan lima gelombang folikel (Bleach et al. 2004, Viana et al. 2000). Pola gelombang yang berbeda akan menyebabkan waktu terjadinya pertumbuhan gelombang folikel juga berbeda. Karena itulah diharapkan pengamatan dinamika ovarium pada sapi PO dapat menjadi data dasar dan referensi dalam program superovulasi yang dimulai pada saat terjadi pertumbuhan gelombang folikel. Dengan demikian diharapkan program superovulasi berbasis pertumbuhan folikel akan menghasilkan embrio layak transfer yang maksimal pada sapi PO.
Perumusan Masalah
Peningkatan populasi ternak bangsa persilangan dengan Bos taurus telah menekan populasi ternak lokal, termasuk bangsa sapi PO. Untuk itu perlu dilakukan percepatan pembibitan sapi PO terutama yang memiliki performans (genetik) unggul melalui penerapan program superovulasi dan embrio transfer. Untuk mencapai tujuan tersebut akan dilakukan serangkaian penelitian tentang pemanfaatan informasi dinamika ovaria dalam program superovulasi. Penelitian ini terdiri dari tiga tahap yaitu dengan judul:
1. Tahap I: Dinamika ovaria pada sapi Peranakan Ongole (PO) setelah penyuntikan GnRH dalam protokol Ovsynch berbasis preparat progesteron
2. Tahap II: Dinamika ovarium dan repeatabilitas pertumbuhan gelombang folikel pada sapi Peranakan Ongole (PO)
3. Tahap III: Respon superovulasi sapi Peranakan Ongole melalui penyuntikan tunggal FSH ke dalam ruang epidural
Keseluruhan rangkaian penelitian ini telah mendapat sertifikat persetujuan etik hewan dari Komisi Etik Hewan, Fakultas Kedokteran Hewan, Institut Pertanian Bogor (Lampiran 1).
4
Tujuan Penelitian
Secara umum penelitian ini dilakukan untuk mendapatkan metode superovulasi yang efisien berbasis pertumbuhan gelombang folikel pada sapi PO dalam rangka menghasilkan embrio layak transfer,.
Secara khusus penelitian ini dilakukan untuk:
1. Menguji efek GnRH terhadap dinamika ovaria dalam protokol sinkronisasi ovulasi yang berbasis preparat progesteron intravaginal.
2. Menerangkan karakteristik pola dinamika ovarium dan repeatabilitas gelombang folikel
3. Menguji efek penyuntikan tunggal hormon FSH ke dalam ruang epidural dengan tujuan superovulasi untuk memperoleh embrio layak transfer.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat menghasilkan:
1. Metode yang optimal untuk menghasilkan sinkronisasi ovulasi folikel yang lebih seragam.
2. Data dasar pola dinamika ovarium sapi PO yang dapat menjadi dasar aplikasi bioteknologi reproduksi lebih lanjut.
3. Metode baru teknik penyuntikan FSH untuk tujuan superovulasi yang lebih efektif dan menghasikan embrio layak transfer yang optimal.
Tingkat kebaruan
2
DINAMIKA OVARIA PADA SAPI PERANAKAN ONGOLE
(PO) SETELAH PENYUNTIKAN GNRH DALAM
PROTOKOL OVSYNCH BERBASIS PREPARAT
PROGESTERON
Pendahuluan
Sapi Ongole (B. indicus) adalah bangsa sapi yang beradaptasi sangat baik untuk berkembang di daerah dengan iklim tropis. Sapi ini tahan terhadap temperatur tropis yang panas, tahan terhadap parasit eksternal dan serangga, menunjukkan respon baik pada pakan yang berkualitas namun juga tahan terhadap pakan berkualitas rendah di lapangan. Secara umum sapi ongol berwarna putih, dan terdapat sedikit warna abu-abu pada bagian punggung dan belakang (Gaur et al. 2002). Namun sapi Ongole juga dilaporkan memiliki periode masa berahi yang lebih pendek dibandingkan dengan sapi bangsa B. taurus, sering tidak menunjukkan tanda-tanda berahi yang jelas dan kebanyakan kejadian berahi ini terjadi di waktu malam hari (Behl et al. 2010). Kelemahan dalam menunjukkan tanda-tanda berahi ini menjadi penyebab utama kesulitan penerapan teknologi reproduksi seperti inseminasi buatan (IB) pada sapi bangsa B. indicus (Sartori dan Baros 2011, Acevedo et al. 2007).
Sapi Ongole berasal dari India dan mulai didatangkan ke Indonesia (pulau Sumba, Nusa Tenggara Timur) sekitar awal abad ke-19. Sapi ini banyak dikembangbiakkan di pulau Jawa dan dikawinsilangkan dengan sapi lokal Jawa untuk menghasilkan sapi persilangan yang saat ini dikenal dengan sapi Peranakan Ongole (PO) (Diwyanto 2008). Saat ini sapi PO sudah berkembang secara alami menjadi suatu bangsa sapi yang stabil, dengan karakteristik morfologi yang mudah dikenali. Nuryadi dan Wahjuningsih (2011) melaporkan bahwa sapi PO memiliki masa tidak bunting panjang yang disebabkan oleh masalah deteksi berahi dan menyebabkan interval beranak menjadi panjang. Karena itulah diperlukan protokol sinkronisasi berahi yang tepat untuk mengoptimalkan deteksi berahi pada sapi PO. Protokol sinkronisasi berahi yang banyak berkembang saat ini umumnya melakukan control terhadap pertumbuhan gelombang folikel dan menghasilkan sinkronisasi ovulasi (protokol Ovsych) melalui kombinasi gonadotropin releasing hormone (GnRH), implantasi preparat progesteron dan penyuntikan PGF2α dalam rangka pelaksanaan program IB pada satu waktu (fixed time artificial insemination, FTAI) (Sahu et al. 2014, Kasimanickam et al. 2014). Penyuntikan hormon GnRH dengan dosis tunggal akan menyebabkan pelepasan hormon LH yang memicu sinkronisasi ovulasi atau luteinisasi pada kebanyakan folikel dominan. Sebagai konsekwensi, sebuah gelombang folikel baru akan terbentuk dalam 2 atau 3 hari setelah penyuntikan GnRH (Patterson et al. 2003). Sementara itu, suplementasi preparat progesteron diantara penyuntikan GnRH dan PGF2α akan meningkatkan efek sinkronisasi dengan mencegah terjadinya berahi dini, LH surge dan ovulasi pada sapi tanpa mempengaruhi waktu pelaksanaan program IB (Sá Filho et al. 2010, Whittier et al. 2010, Peterson et al. 2011).
8
yang baik pada sapi B. taurus (Azevedo et al. 2014, Kim et al. 2005) dan B. indicus (Martins et al. 2014, Meneghetti et al. 2009). Namun bagaimanapun penelitian lebih lanjut masih diperlukan untuk menguji apakah protokol tersebut dapat diterapkan dan menghasilkan sinkronisasi ovulasi yang optimal pada sapi PO. Penelitian ini bertujuan untuk mendapatkan informasi pengaruh GnRH terhadap dinamika ovaria meliputi pertumbuhan gelombang folikular, tingkat ovulasi, diameter folikel praovulasi dan CL pada protokol Ovsynch yang berbasis preparat progesteron intravaginal.
Materi dan Metode Materi Penelitian
Penelitian ini menggunakan sapi PO induk sebanyak 12 ekor dan dara sebanyak 18 ekor. Sapi PO induk berumur antara 5 – 7 tahun dengan bobot badan antara 375 – 450 kg, sedangkan sapi PO dara berumur antara 2.5 – 3.5 tahun dengan bobot badan 325 – 375 kg. Nilai Kondisi Tubuh (NKT) sapi PO antara 2.7 – 3.2 dengan basis penilaian 0 – 5 dan memiliki organ reproduksi yang normal sesuai pemeriksaan per rektal. Sapi-sapi ini dipelihara didalam kandang, diberi pakan hijauan harian sebanyak 30 – 40 kg/ekor dan ditambah dengan 2 – 3 kg/ekor konsentrat. Sapi yang mendapat perlakuan berbeda, diberikan istirahat selama 2 bulan sebelum diberikan perlakuan yang baru untuk mengeliminasi pengaruh perlakuan sebelumnya.
Perlakuan
Sapi PO pada masing-masing kelompok umur dibagi menjadi tiga kelompok secara acak. Pada hari ke-0 semua kelompok sapi diberi perlakuan implantasi preparat progesteron (Cuemate, mengandung 1.56 mg progesteron dalam dua pod silicon, Bioniche Animal Health, Asia Pty, Australia) selama 7 hari. Cuemate dimasukkan kedalam vagina menggunakan aplikator yang diberi pelumas. Selanjutnya pada hari ke-7 hormon prostaglandin diberikan sebanyak 2 ml secara intramuskular (i.m.) (Prostavet C, 5 mg of etiproston per 2 ml larutan; Virbac Animal Health, Prancis) sesaat setelah pelepasan Cuemate. Kelompok Cuemate-PGF2α-GnRH (CPG) diberikan 100 µg hormon GnRH (Fertagyl, mengandung 1000 µg gonadorelin diacetate tetrahydrate dalam larutan 20 ml, Intervet Schering-Plough Animal Health, Jerman) secara i.m. pada hari ke-9. Pada kelompok GnRH-Cuemate-PGF2α-GnRH (GCPG) diberikan perlakuan penyuntikan 100µg GnRH secara i.m. pada hari ke-0 dan 9. Sedangkan kelompok Cuemate-PGF2α (CP) tidak mendapatkan tambahan perlakuan yang lain (Gambar 2).
Pengamatan ovaria
Gambar 2. Skema jenis perlakuan pada kelompok CP, CPG and GCPG. PGF2α (PG) diberikan sesaat setelah pelepasan Cuemate. Pengamatan dengan USG (U) dilakukan setiap hari mulai H0 – H4, H7 sampai terjadi ovulasi (Ov) dan 7 hari setelah ovulasi (CL).
Analisa Hormon Progesteron
Analisa hormon progesteron untuk menilai fungsionalitas CL diukur menggunakan teknik enzyme linked immunosorbent assay (ELISA). Sampel darah dikoleksi dari vena koksigeal pada hari ke-7 setelah ovulasi, ditampung dalam plain vacutainer dan dibiarkan selama 24 jam untuk menjadi menggumpal. Serum darah dipisahkan dengan sentrifugasi pada kecepatan 500G selama 10 menit pada suhu ruang dan hasilnya dibekukan pada suhu -20° C sampai dilakukan analisa hormon.
Analisa Statistik
10
Pengaruh umur sapi pada masing-masing perlakuan diuji menggunakan Independent-Sample T Test. Awal pertumbuhan gelombang folikel, perbedaan rataan pada pengamatan diameter folikel, diameter CL dan konsentrasi progesteron diantara perlakuan dianalisa menggunakan ANOVA satu arah dan dilanjutkan dengan uji Tukey menggunakan post hoc jika terdapat perbedaan yang nyata pada ANOVA dengan P-value 0,05 dianggap sebagai nyata berbeda. Hubungan antara diameter folikel ovulatori dan diameter CL diuji menggunakan Pearson correlation test. Sedangkan proporsi sapi yang mengalami ovulasi dan yang memiliki gelombang pertumbuhan folikel baru dibandingkan menggunakan uji Chi kuadrat.
Hasil dan Pembahasan
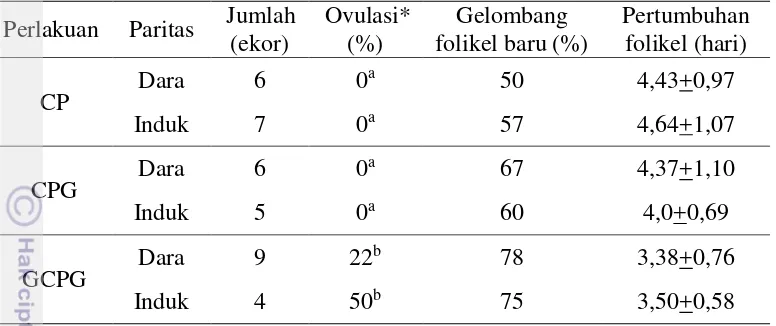
Penelitian ini menunjukkan bahwa penyuntikan GnRH pada hari ke-0 (perlakuan GCPG) menyebabkan ovulasi baik pada dara maupun induk sebesar 22 dan 50%, sementara itu tidak ada kejadian ovulasi yang teramati pada perlakuan CP dan CPG, yaitu perlakuan tanpa pemberian GnRH pada hari ke-0 (Tabel 1). Penelitan lain melaporkan penyuntikan GnRH tanpa memperhatikan siklus berahi menghasilkan tingkat ovulasi sebesar 27 – 45 % (Sahu et al. 2014, Giordano et al. 2013, Lima et al. 2011). Diameter minimum folikel yang mengalami ovulasi sebagai respon terhadap perlakuan GnRH pada hari ke-0 (GnRH pertama) dalam penelitian ini adalah 9 mm, lebih kecil dari hasil penelitian Sartori et al. (2001) pada sapi Nellor (10 mm). Mungkin perbedaan ini dipengaruhi oleh bangsa sapi yang digunakan dan teknik pengukuran folikel yang berbeda.
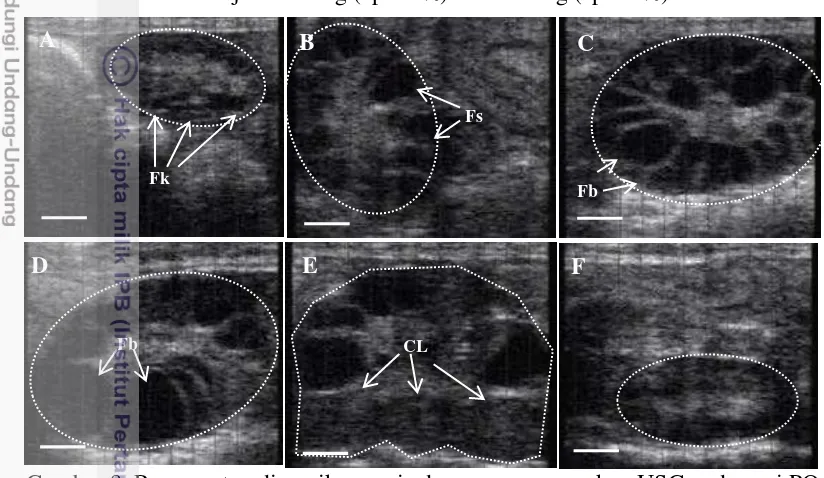
Tabel 1. Dinamika ovaria pada kelompok dengan perlakuan penyuntikan GnRH pertama (Perlakuan GCPG) dan tanpa penyuntikan GnRH pertama (perlakuan CP dan CPG) pada dara dan induk.
Perlakuan Paritas Jumlah
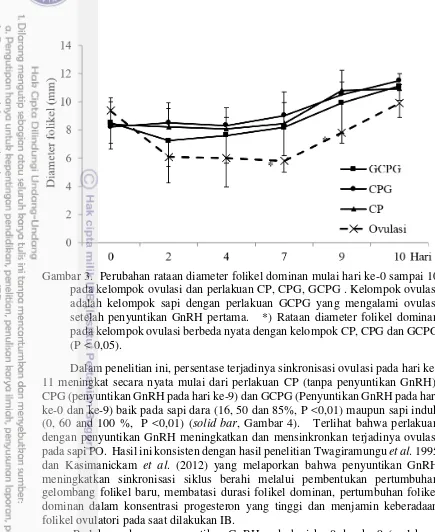
C:Cuemate, P:PGF2α, G:GnRH. Penyuntikan GnRH pertama dilakukan pada hari ke-0. Nilai dengan superscript berbeda pada kolom yang sama berbeda nyata (P<0,05). *) Kejadian ovulasi yang teramati pada awal perlakuan (hari ke-2 dan 3) Perubahan rataan diameter folikel dominan tidak berbeda nyata antara perlakuan CP, CPG dan GCPG mulai hari ke-0 sampai ke-10 (Gambar 3). Tetapi jika sapi yang mengalami ovulasi sebagai respon terhadap GnRH pertama pada perlakuan GCPG dikelompokkan sebagai satu grup (kelompok ovulasi), maka rataan diameter kelompok ovulasi akan berbeda nyata (P<0,05) terutama pada hari ke-7 dan ke-9, dibandingkan dengan perlakuan CP, CPG dan GCPG (Gambar 3).
12
Gambar 3. Perubahan rataan diameter folikel dominan mulai hari ke-0 sampai 10 pada kelompok ovulasi dan perlakuan CP, CPG, GCPG . Kelompok ovulasi adalah kelompok sapi dengan perlakuan GCPG yang mengalami ovulasi setelah penyuntikan GnRH pertama. *) Rataan diameter folikel dominan pada kelompok ovulasi berbeda nyata dengan kelompok CP, CPG dan GCPG (P < 0,05).
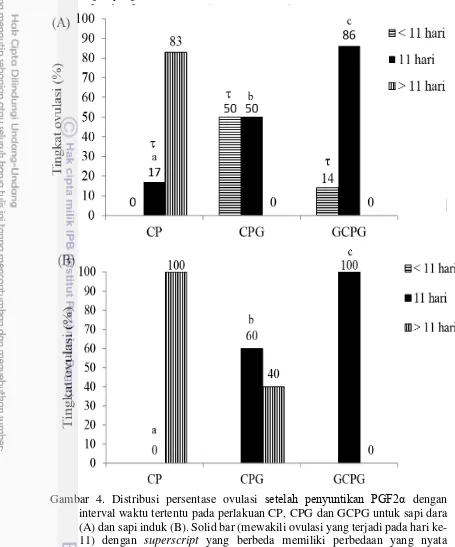
Dalam penelitian ini, persentase terjadinya sinkronisasi ovulasi pada hari ke-11 meningkat secara nyata mulai dari perlakuan CP (tanpa penyuntikan GnRH), CPG (penyuntikan GnRH pada hari ke-9) dan GCPG (Penyuntikan GnRH pada hari ke-0 dan ke-9) baik pada sapi dara (16, 50 dan 85%, P <0,01) maupun sapi induk (0, 60 and 100 %, P <0,01) (solid bar, Gambar 4). Terlihat bahwa perlakuan dengan penyuntikan GnRH meningkatkan dan mensinkronkan terjadinya ovulasi pada sapi PO. Hasil ini konsisten dengan hasil penelitian Twagiramungu et al. 1995 dan Kasimanickam et al. (2012) yang melaporkan bahwa penyuntikan GnRH meningkatkan sinkronisasi siklus berahi melalui pembentukan pertumbuhan gelombang folikel baru, membatasi durasi folikel dominan, pertumbuhan folikel dominan dalam konsentrasi progesteron yang tinggi dan menjamin keberadaan folikel ovulatori pada saat dilakukan IB.
Ovsynch menggunakan penyuntikan dua kali GnRH masih menghasilkan tingkat kebuntingan yang memuaskan.
Gambar 4. Distribusi persentase ovulasi setelah penyuntikan PGF2α dengan interval waktu tertentu pada perlakuan CP, CPG dan GCPG untuk sapi dara (A) dan sapi induk (B). Solid bar (mewakili ovulasi yang terjadi pada hari ke-11) dengan superscript yang berbeda memiliki perbedaan yang nyata (P<0,05). : menunjukkan ovulasi terjadi lebih awal pada sapi dara dibandingkan dengan sapi induk.
14
dibandingkan sapi dara. Sementara itu, Bo et al. (2003) melaporkan bahwa 28.9% sapi dara B. indicus yang diberi perlakuan Ovsynch menunjukkan tanda tanda berahi lebih awal setelah penyuntikan GnRH kedua. Dalam penelitian ini, rata-rata jumlah folikel besar (lebih dari 6 mm) yang teramati dibagian kortex ovaria pada sebagaian besar hasil pengamatan harian pada sapi dara cenderung lebih banyak dibandingkan sapi induk baik pada perlakuan CP, CPG dan GCPG. Diduga folikel pada sapi dara cenderung mengalami tumbuh lebih cepat dibandingkan pertumbuhan folikel pada sapi induk sehingga folikel mengalami ovulasi lebih cepat. Untuk itu diperlukan penelitian lebih lanjut untuk mengkonfirmasi dugaan tersebut.
Sebagian besar sapi dara dan induk dalam penelitian ini terlihat memiliki sebuah CL atau berada dalam fase luteal pada saat dilakukan pemasangan preparat progesteron di hari ke-0 pada perlakuan CP (83% dan 100 %), CPG (67% dan 100%) dan GCPG (78% dan 75%), walaupun tidak diketahui umur dan kapabilitas CL ini dalam memproduksi hormon progesteron. Perlakuan implantasi preparat progesteron eksternal yang dimulai pada saat sapi berada dalam fase luteal akan memperkuat pengaruh progesteron endogen yang diproduksi oleh sel-sel luteal dan mencegah terjadinya ovulasi (Kim et al. 2005, Whittier et al. 2010). .
Jika perlakuan implantasi preparat progesteron dimulai pada saat sapi dalam fase folikuler (tidak ada CL yang terlihat pada ovaria), maka pelepasan progesteron dari preparat progesteron intravaginal akan menciptakan lingkungan seperti dalam kondisi fase mid-luteal pada sapi dara maupun induk yang berfungsi untuk mencegah berahi dan ovulasi dini seperti yang dilaporkan oleh Bo et al. 2003. Sales et al. (2015) juga melaporkan konsentrasi plasma progesteron pada sapi yang diovariektomi akan meningkat seperti fase mid-luteal (5-7 ng/mL) setelah implantasi preparat progesteron (CIDR) dalam waktu 24 jam dan kemudian menurun secara bertahap. Plasma progesteron yang diproduksi oleh CL mencapai puncaknya antara hari ke-10 sampai 14 setelah ovulasi dan kemudian menurun setelah CL mengalami luteolisis (Adam et al. 2008)
Ketepatan sinkronisasi ovulasi sangat dipengaruhi oleh kemampuan kontrol terhadap pertumbuhan CL dan folikel. Penyuntikan PGF2α mungkin akan menginduksi terjadinya luteolisis, tetapi tidak serta merta akan menyebabkan folikel mengalami ovulasi secara bersamaan (Akbarabadi et al. 2014). Waktu terjadinya ovulasi dipengaruhi oleh diameter folikel dominan pada saat dilakukan penyuntikan PGF2α (Wiltbank dan Pursley 2014), karena folikel dominan akan terus tumbuh dan mengalami ovulasi pada saat folikel mencapai ukuran ovulatori, walaupun CL sudah mengalami luteolisis sebagai respon terhadap penyuntikan PGF2α. Hal ini mungkin menjelaskan waktu ovulasi pada perlakuan CP terjadi lebih tidak sinkron dibandingkan perlakuan CPG dan GCPG karena pada perlakuan CP tidak ada kontrol pertumbuhan folikular dan ovulasi melalui penyuntikan GnRH sebagaimana halnya pada perlakuan CPG dan GCPG.
berahi karena tanda-tandanya memang sulit diamati pada sapi bangsa B. indicus. Dalam hal ini, sepertinya sapi PO menunjukkan tanda-tanda berahi pada malam hari atau mengalami berahi diam (silent heat) karena ada sedikit jejak lendir yang mengering teramati di pagi hari pada beberapa sapi PO.
Tabel 2. Pengamatan tanda-tanda berahi dan dinamika ovaria pada sapi PO dara dan induk yang diberi perlakuan CP, CPG dan GCPG.
Perlakuan Paritas Jumlah
Alasan lain yang mungkin untuk menjelaskan kurang teramatinya tanda-tanda berahi pada sapi, selain karena pengaruh bangsa sapi adalah pengaruh dari suhu lingkungan yang rendah. Penelitian ini dilakukan di lokasi dengan ketinggian sekitar 900 m diatas permukaan laut dengan suhu rata-rata antara 16 - 26oC. Suhu rendah mungkin dapat menjadi faktor cekaman pada sapi PO sehubungan dengan sapi ini dikembangkan dan sudah beradaptasi di iklim tropis yang panas dan menyebabkan gejala berahi tidak terlihat pada sebagian sapi PO. Pada penelitian lain, Sartori dan Barros (2011) melaporkan 50% populasi sapi dara tidak teramati adanya berahi yang mungkin disebabkan oleh stress karena pengaruh observasi transrektal dengan USG yang berlebihan dan penurunan suhu lingkungan terutama dimalam hari. Faktor cekaman dapat mempengaruhi fungsi-fungsi reproduksi melalui aksi pada hipotalamus dengan mengganggu pelepasan LH yang diinduksi oleh GnRH dan hal ini mengakibatkan pengurangan produksi estradiol yang menyebabkan perlambatan pertumbuhan folikel (Etim et al. 2013). Mekanisme ini mungkin menjelaskan kurangnya tingkat pengamatan gejala berahi sapi PO dalam penelitian walaupun sampai saat ini belum ada penelitian yang melaporkan pengaruh cekaman dingin terhadap fungsi reproduksi pada sapi PO atau sapi bangsa B. indicus yang lain.
16
cekaman dingin (15–250C) dibandingkan dengan cekaman panas (3–6oC) sehingga cekaman dingin menyebabkan masalah yang lebih sedikit dan bisa lebih ditolerensi oleh hewan dibandingkan dengan cekaman panas.
Dalam pengamatan yang lain, dua sapi dara pada perlakuan GCPG mengalami ovulasi setelah penyuntikan GnRH pertama pada hari ke-3, namun tidak mengalami ovulasi dalam penyuntikan GnRH kedua pada hari ke-9. Konfirmasi dengan ultrasound menunjukkan bahwa sapi dara ini memiliki CL asesoris sebagai perkembangan lebih lanjut dari folikel yang mengalami ovulasi sebagai respon penyuntikan GnRH pertama. Diduga CL asesoris ini tidak mengalami luteolisis setelah penyuntikan PGF2α pada ehari ke-7 yang disebabkan oleh umur CL tersebut yang masih muda (4 hari). Hasil ini sependapat dengan Kasimanickam et al. (2014) yang menyatakan bahwa ovulasi yang terjadi setelah penyuntikan GnRH pertama akan membentuk CL yang mungkin tidak mengalami regresi dengan satu kali penyuntikan PGF2α. Sel-sel luteal besar yang berasal dari sel-sel granulosa lebih responsive terhadap PGF2α dan mengakibatkan luteolisis dibandingkan dengan sel-sel luteal kecil (Peterson et al. 2011). Pada penelitian ini umur CL asesoris masih muda sehingga mungkin lebih banyak mengandung sel luteal kecil dibandingkan dengan sel luteal besar sehingga perlakuan PGF2α tidak merusak fungsi sel luteal untuk memproduksi hormon progesteron. Data ini menyarankan bahwa penyuntikan hormon PGF2α pada sapi dara mungkin lebih baik dilakukan pada hari berikutnya (hari ke-8) untuk memberikan kesempatan sel luteal besar bertumbuh dan respon terhadap perlakuan PGF2α. Penelitian lebih lanjut perlu dilakukan untuk mengkonfirmasi dugaan ini.
Diameter folikel ovulatori antara perlakuan CP, CPG dan GCPG dalam penelitian ini tidak berbeda nyata pada sapi dara (11,9+0,5, 11,9+0,5 dan 12,1+0,6 mm) dan sapi induk (11,7+0,4, 11,8+0,7, 11,1+0,6 mm) (Tabel 2). Hasil ini konsisten dengan Sahu et al. (2014) yang melaporkan folikel ovulatori memiliki diameter yang tidak berbeda nyata antara perlakuan tanpa penyuntikan GnRH pertama maupun perlakuan yang mendapatkan penyuntikan GnRH dalam protokol Ovsynch berbasis CIDR pada sapi perah. Tetapi diameter folikel yang berkembang pada perlakuan sinkronisasi dengan menggunakan preparat progesteron (Tabel 2), memiliki rataan diameter yang sangat nyata lebih kecil (P<0,01) jika dibandingkan dengan folikel ovalatori yang berkembang secara alami tanpa perlakuan preparat progresteron (Tabel 4, pada penelitian tentang repeatabilitas gelombang folikel), dengan rataan diameter folikel berturut-turut 11,23+0,53 dan 12,07+0,54 mm. Hal ini terjadi karena tambahan pelepasan progesteron dari Cuemate menekan pertumbuhan folikel. Hasil ini konsisten dengan penelitian lain yang melaporkan program sinkronisasi ovulasi berbasis CIDR baru menghasilkan diameter folikel ovulatori yang lebih kecil dibandingkan dengan menggunakan CIDR bekas pakai dengan tidak mempengaruhi tingkat ovulasinya (Junior et al. 2010, Dias et al. 2009).
efek heterosis menyebabkan perbedaan ukuran dari CL, dibandingkan dengan sapi Ongol galur murni. Hal lain yang mungkin menyebabkan perbedaan hasil adalah perbedaan jenis ultrasound, teknik pemindaian dan teknik pengukuran ovarium.
Diameter folikel ovulatori dalam penelitian ini memiliki korelasi positif dengan diameter CL (R2=0,45, P<0,01), sesuai dengan hasil penelitian sebelumnya yang dilakukan oleh Dadarwal et al. 2013 dan Perez et al. 2009. Field et al. (2012) menyatakan setiap peningkatan diameter folikel sebesar 1 mm in follicle size akan meningkatkan berat CL sebesar 1.5 g pada hari ke 10 setelah ovulasi. Lebih lanjut analisa konsentrasi progesteron darah pada hari ke-7 setelah ovulasi dalam penelitian ini juga tidak berbeda nyata antara perlakuan CP, CPG dan GCPG pada sapi dara (4,06+1,37, 4,48+2.08, dan 4,12+1,11 ng/mL) dan sapi induk (3,65+1,96, 4,26+1,36 dan 3,92+1,26 ng/mL) (Tabel 2). Dalam penelitian lain Sahu et al. (2014) melaporkan konsentrasi progesteron darah sapi Nellor pada hari ke-7 setelah ovulasi adalah 3 – 5 ng/ml.
Penelitian ini menunjukkan bahwa terjadinya ovulasi setelah penyuntikan GnRH pertama pada hari ke-0 menghasilkan pertumbuhan gelombang folikel yang tersinkronisasi dan meningkatkan keseragaman ukuran folikel ovulatori pada saat penyuntikan PGF2α. Selain itu, perlakuan GnRH kedua pada hari ke-9 menginduksi terjadinya keseragaman tingkat ovulasi lebih baik dibandingkan perlakuan tanpa GnRH. Karena itu menjadi jelas bahwa penyuntikan GnRH pertama dan kedua pada protokol Ovsynch berbasis implantasi preparat progesteron selama 7 hari sangat bermanfaat untuk menghasilkan sinkronisasi pertumbuhan gelombang folikel dan meningkatkan keseragaman terjadinya ovulasi baik pada sapi PO dara maupun sapi induk.
Simpulan dan saran
Penelitian ini menyimpulkan bahwa protokol GnRH-Cuemate-Prostaglandin-GnRH (GCPG) meningkatkan sinkronisasi terjadinya ovulasi baik pada sapi dara maupun sapi induk lebih baik dibandingkan metode Cuemate-Prostaglandin (CP) dan Cuemate-Prostaglandin-GnRH (CPG). Penelitian lebih lanjut mungkin diperlukan untuk menguji efektifitas metode sinkronisasi ovulasi GCPG sampai pada hasil perolehan kebuntingan di lapangan.
Daftar Pustaka
Acevedo N, Carlos SG, Angel P, Agustín O. 2007. Dynamics in sexually active groups of Zebu cattle (Bos indicus) comparing two procedures for estrus induction. J Vet Behav. 2:5-9.
Adams GP, Jaiswal R, Singh J Malhi P. 2008. Progress in understanding ovarian follicular dynamics in cattle. Theriogenology 69:72–80
Adriani, Rosadi B, Depison. 2009. The Application of Follicle Stimulating Hormone and Pregnant Mare Serum Gonadotrophin Hormone for Superovulation in Brahman Cross Cattle. Med Pet 32(2):163-170
18
insemination during the warm or cold periods of the year. Theriogenology. 82: 509–516
Azevedo C, Maia I, Canada N, Simões J. 2014. Comparison of fertility, regular returns-to-estrus, and calving interval between Ovsynch and CO-synch+CIDR protocols in dairy cows. Theriogenology. 82 :910–914
Behl R, Behl J, Joshi BK. 2010. Heat tolerance mechanisms in cattle—status in zebu cattle: A review. Indian J Anim Sci.80 (9): 891–897
Bianca W. 1976. The significance of meteorology in Animal Production. Int J. Biometeorol. 20(2), 139–156
Bó GA, Baruselli PS, Mart MF. 2003. Pattern and manipulation of follicular development in Bos indicus cattle. Anim Reprod Sci.78:307–326
Bo GA, Adams GP, Pierson RA, Mapletoft RJ. 1995. Exogenous control of follicular wave emergence in cattle. Theriogenology. 43:31–40
Dadarwal D, Mapletoft RJ, Adams GP, Pfeifer LFM, Creelman C, Singh J. 2013. Effect of progesterone concentration and duration of proestrus on fertility in beef cattle after fixed-time artificial insemination. Theriogenology. 79: 859–866 Diwyanto K. 2008. Pemanfaatan sumber daya lokal dan Inovasi teknologi dalam
mendukung Pengembangan sapi potong di indonesia. Pengembangan Inovasi Pertanian 1(3): 173-188.
Dias CC, Wechsler FS, Day ML, Vasconcelos JLM. 2009. Progesteron concentration, exogenous eCG and timing of prostaglanding treatment affect fertility in postpuberal Nelore heifers. Theriogenology 72: 378-85.
Etim NN, Offiong EEA, Udo MD, Williams MD, Evans EI. 2013. Physiological relationship between stress and reproductive efficiency. Agric Biol J N Am. 4(6):600-604.
Fields SD, Gebhart KL, Perry BL, Gonda MG, Wright CL, Bott RC, Perry GA. 2012. Influence of standing estrus before an injection of GnRH during a beef cattle fixed-time AI protocol on LH release, subsequent concentrations of progesterone, and steriodogenic enzyme expression. Dom Anim Endo. 42: 11– 19
Giordano JO, Wiltbank MC, Fricke PM, Bas S, Pawlisch R, Guenther JN, Nascimento AB. 2013. Effect of increasing GnRH and PGF2α dose during Double-Ovsynch on ovulatory response, luteal regression, and fertility of lactating dairy cows. Theriogenology. 80:773–783
Gaur GK, Kaushik SN, Garg RC. 2002. Ongole cattle status in India. Animal Genetic Resources Information. 32: 27-33
Galina CS, Oriheula A. 2007. The Detection of estrus incattle raised under tropical conditions: What we know and what we need to know. Horm Behav. 52(1): 32-38.
Schneider JE. 2004. Energy balance and reproduction. Physiol Behav. 81:289– 317
Junior CI, Sa Filho OG, R. F. G. Perez, F. H. S. Aono, M. L. Day & J. L. M. Vasconcelos. 2010. Reproductive Performance of prepubertal Bos indicus herifer after progesteron-based treatments. Theriogenology. 74:903-911. Kasimanickam RK, Firth P, Schuenemann GM, Whitlock BK, Gay JM, Mooe DA,
Kasimanickam Rk, Asay M, Firth P, Whittier WD, Hall JB. 2012. Artificial insemination at 56 h after intravaginal progesterone device removal improved AI pregnancy rate in beef heifers synchronized with fiveday CO-Synch controlled internal drug release (CIDR) protocol. Theriogenology. 77:1624–31. Keskin A, Yilmazbas-Mecitoglua G, Gumena A, Karakayaa E, Daricib R, Okutc H.
2010. Effect of hCG vs. GnRH at the beginning of the Ovsynch on first ovulation and conception rates in cyclic lactating dairy cows. Theriogenology. 74 :602–607.
Kim UH, Suh GH, Nam HW, Kang HG, Kim IH. 2005. Follicular wave emergence, luteal function and synchrony of ovulation following GnRH or estradiol benzoate in a CIDR-treated, lactating Holstein cows. Theriogenology. 63: 260– 268
Lee SM, Rahman MS, Kwon WS, Chung HJ, Yang BS, Pang MG. 2013. Efficacy of four synchronization protocols on the estrus behavior and conception in native Korean cattle (Hanwoo). Theriogenology. 80:855–861.
Lima FS, Ayres H, Favoreto MG, Bisinotto RS, LF Greco, Ribeiro ES. 2011. Effects of gonadotropin-releasing hormone at initiation of the 5-d timed artificial insemination (AI) program and timing of induction of ovulation relative to AI on ovarian dynamics and fertility of dairy heifers. J Dairy Sci. 94:4997–5004. Martins T, Peres RFG, Rodrigues ADP, Pohler KG, Pereira MHC, Day ML,
Vasconcelos JLM. 2014. Effect of progesterone concentrations, follicle diameter, timing of artificial insemination, and ovulatory stimulus on pregnancy rate to synchronized artificial insemination in postpubertal Nellore heifers. Theriogenology. 81: 446–453
Meneghetti M, Sa´ Filho OG, Peres RFG, Lamb GC, Vasconcelos JLM. 2009. Fixed-time artificial insemination with estradiol and progesterone for Bos indicus cows I: Basis for development of protocols. Theriogenology. 72: 179– 189
Nardone A, Ronchi B, Lacetera N, Bernabucci U. 2006. Climatic Effects on Productive Traits in Livestock. Vet Res Com. 30(Suppl. 1):75–81
Nuryadi, Wahjuningsih S. 2011. Penampilan reproduksi sapi peranakan ongole dan peranakan limousin di Kabupaten Malang. J Ternak Tropika 12(1): 76-81, 2011
Patterson DJ, Kojima FN, Smith MF. 2003. A review of methods to synchronize estrus in replacement beef heifers and postpartum cows. J Anim Sci. 81:166-177. Perez RFG, Júnior G, Sá Filho OG, Nogueira GP, Vasconcelos JLM. 2009.
Strategies to improve fertility in Bos indicus postpubertal heifers and nonlactating cows submitted to fixed-time artificial insemination. Theriogenology. 72:681–9.
Peterson C, Alkar A, Smith S, Kerr S, Hall JB, Moore D. 2011. Effects of one versus two doses of prostaglandin F2alpha on AI pregnancy rates in a 5-day, progesterone-based, CO-Synch protocol in crossbred beef heifers. Theriogenology. 75:1536–42.
Ramana KV, Rao KV, Supriya K, Rajanna N. 2013. Postpartum ovarian follicular dynamics and estrus activity in lactating Ongole cows. J Res Angrau. 41(1): 51-55
20
GnRH+progesterone program on synchronization of ovulation in pasture-based dairy heifers. Theriogenology. 82:643–651
Sá Filho MF, Ayres H, Ferreira RM, Marques M, Reis EL, Silva RC, Rodrigues CA, Madureira EH, Bó GA, Baruselli PS. 2010. eCG and GnRH enhance fertility in a norgestomet-based, timed AI protocol in suckled Nelore (Bos indicus) cows. Theriogenology 73: 651–658
Sales JNS,Carvalho JBP, Crepaldi GA, Soares JG, Girotto RW, Maio JRG, Souza JC, Baruselli PS. 2015. Effect of circulating progesterone concentration during synchronization for fixed-time artificial insemination on ovulation and fertility in Bos indicus (Nelore) beef cows . Theriogenology. 83: 1093–1100
Sartori R, Barros CM. 2011. Reproductive cycles in Bos indicus cattle. Anim Reprod Sci. 124: 244–250
Sartori R, Fricke M, Ferreira JC, Ginther OJ, Wiltbank MC. 2001. Follicular deviation and acquisition of ovulatory capacity in bovine follicles. Biol Reprod 65:1403–1409
Souza AH, Ayres H, Ferreira RM, Wiltbank MC. 2008. A new presynchronization system (Double-Ovysnch) increases fertility at first postpartum timed AI in lactating dairy cows. Theriogenology. 70:208 –15.
Twagiramungu H, Guilbault LA, Dufour JJ. 1995. Synchronization of ovarian follicular waves with a gonadotropin-releasing hormone agonist to increase the precision of estrus in cattle: a review. J Anim Sci.73:3141–51.
Whittier WD, Kasimanickam RK, Currin JF, Schramm HH, Vlcek M. 2010. Effect of timing of second prostaglandin F 2 alpha administration in a 5-day, progesterone-based CO-Synch protocol on AI pregnancy rates in beef cows. Theriogenology. 74:1002–9.
3
DINAMIKA OVARIA DAN REPEATABILITAS
PERTUMBUHAN GELOMBANG FOLIKEL PADA SAPI
PERANAKAN ONGOLE (PO)
Pendahuluan
Pertumbuhan folikel pada ovarium secara umum terdiri dari dua kelompok folikel yang berbeda yaitu kelompok folikel tidak bertumbuh (non growing pool) yaitu folikel primordial dan kelompok folikel yang bertumbuh (growing pool) terdiri dari folikel primer, sekunder dan tersier (Kanitz et al. 2003). Perubahan pertumbuhan folikel primordial keluar dari kelompok tidak bertumbuh dan mengalami perubahan transisi menjadi folikel primer berlangsung sepanjang siklus reproduksi. Oosit mengalami peningkatan diameter dan sel-sel pragranulosa berubah berbentuk kuboid dan lebih lanjut berproliferasi membentuk satu lapis sel-sel kuboid dipermukaan oosit yang berkembang (Fortune et al. 2000). Selama proses rekrutment folikel menjadi folikel yang bertumbuh, kelompok sel-sel teka mengorganisasi diri menjadi lapisan khusus disekeliling folikel dan membuat interaksi yang kuat dengan sel-sel granulosa. Pada fase ini oosit mencapai ukuran finalnya dan berkembang mencapai tahap full developmental competence dan disebut folikel primer (Fair et al. 1997, Parrott dan Skinner 2000). Namun mekanisme yang memicu pertumbuhan folikel primordial menjadi primer ini masih kurang dapat dimengerti (Kanitz et al. 2003). Perubahan ekspresi gen diduga terlibat dalam rekrutment, seleksi, dominasi dan atresia pada folikel (Beg et al. 2002).
Fase pertumbuhan cepat terjadi dalam waktu 5-7 hari dan fase ini dikenal dengan gelombang pertumbuhan folikel. Penelitian dengan menggunakan ultrasound untuk memonitor populasi folikel dengan kategori diameter tertentu atau folikel yang diidentifikasi secara individual menyimpulkan bahwa pertumbuhan folikel pada sapi terbentuk seperti gelombang (Adam 1999). Gelombang folikel dicirikan dengan pertumbuhan sekelompok folikel kecil dengan diameter 3-4 mm (Adam 1999, Taylor dan Rajamahendran 1991).
Pola pertumbuhan gelombang folikel ini dapat diamati pada periode prapubertal (Melvin et al. 1999), selama periode kebuntingan (Taylor dan Rajamahendran 1991), setelah beranak (Murphy et al. 1990) dan selama siklus berahi (Roche et al. 1999). Pertumbuhan gelombang folikel didahului dengan peningkatan konsentrasi serum FSH dan diikuti dengan pertumbuhan 8-41 folikel kecil secara tiba-tiba dalam waktu 2-3 hari (Ginther et al. 1997). Satu folikel akan terpilih untuk terus tumbuh menjadi folikel dominan (FD) sedangkan folikel lain yang lebih kecil disebut folikel subordinat (FS) dan akan mengalami atresia. Jika terjadi luteolisis pada saat FD dalam fase pertumbuhan, maka folikel akan mengalami proses pematangan final dan dilanjutkan dengan ovulasi. Jika tidak ada luteloisis selama FD sedang tumbuh, maka FD juga akan mengalami atresia (Vasenna et al. 2003, Jaiswal et al. 2004, Adam et al. 2008).
22
dipengaruhi oleh mekanisme sistemik (endokrin) dibandingkan dengan chanel lokal. Hanya satu folikel dominan yang akan terpilih secara acak dari sepasang ovarium dan posisi folikel dominan tersebut bisa pada ipsilateral atau kontraleteral dengan folikel subordinate terbesarnya. Sisi CL atau folikel dominan dari gelombang folikel sebelumnya tidak mempengaruhi sisi folikel ovulatori (Adam et al. 2008).
Mayoritas bangsa sapi menunjukkan pola dua dan tiga gelombang folikel dalam satu siklus berahi (Adam et al. 2008) walaupun terkadang dilaporkan juga adanya pola satu, empat dan lima gelombang folikel (Bleach et al. 2004, Viana et al. 2000). Dinamika folikel merupakan salah satu subyek penelitian yang banyak dilakukan pada sapi bangsa eropa (B. taurus) namun sangat terbatas informasi yang diperoleh dari sapi zebu (B. indicus). Karakteristik reproduksi sapi B. indicus juga tidak identik dengan B. taurus antara lain dalam hal panjang siklus berahi, waktu berahi, tingkah laku berahi, pertumbuhan folikel dominan dan CL, sudah direview oleh Bó et al. (2003). Pengetahuan tentang perbedaan ini sangat penting untuk menetapkan manajemen reproduksi yang lebih baik dan pengembangan teknologi untuk peningkatan performan reproduksi untuk sapi B. indicus terutama pada sapi PO.
Penelitian ini dilakukan untuk mendapatkan informasi karakteristik dinamika ovarium selama siklus berahi dan tingkat repeatabilitas pola pertumbuhan gelombang folikel pada sapi Peranakan Ongol (PO).
Materi dan Metoda Materi Penelitian
Penelitian ini menggunakan sapi PO dewasa sebanyak 9 ekor yang berumur antara 5 sampai 7 tahun dengan berat badan antara 375 sampai 450 kg. Semua sapi yang digunakan memiliki Nilai Kondisi Tubuh (NKT) antara 2,7 sampai 3,2 dalam skala penilaian 1 sampai 5, dengan siklus birahi yang normal. Pakan diberikan sebanyak 30 sampai 40 kg rumput per ekor per hari dan ditambah dengan 2 sampai 3 kg pakan konsentrat per ekor per hari.
Sinkronisasi Ovulasi
Pengamatan dinamika ovaria
Pengamatan dinamika ovaria dilakukan satu hari sekali dengan waktu dan operator yang sama mengunakan ultrasound portabel (EasyScan Lite, Inggris) yang dilengkapi dengan monitor goggle dan probe dengan frekuensi dinamis 4 – 8 Mhz. Unit utama ultrasound dihubungkan dengan komputer dan data pengamatan direkam dalam format video. Pengukuran diameter folikel dan CL dilakukan menggunakan Microsoft image tool. Pengamatan harian menggunakan ultrasound dimulai sejak sapi PO diberikan suntikan PGF2α. Satu interovulatory interval (IOI) didefinisikan sebagai periode waktu antara satu ovulasi dengan ovulasi berikutnya. Ovulasi pertama yang diamati setelah perlakuan sinkronisasi ovulasi dianggap sebagai hari ke-0 dalam satu IOI.
Pada tahap awal, pengamatan dilakukan untuk mendapatkan karakteristik dinamika ovaria dan memetakan pola gelombang dalam satu IOI. Sapi yang digunakan dalam penelitian tahap awal sebanyak 9 ekor. Sedangkan untuk mendapatkan data repeatabilitas pertumbuhan gelombang folikel, pengamatan dinamika ovaria dilanjutkan pada satu IOI berikutnya tanpa jeda menggunakan 6 ekor sapi yang telah diketahui pola jumlah gelombangnya, terdiri dari 3 ekor pola 3 gelombang dan 3 ekor pola 4 gelombang.
Prosedur Analisa Data
Satu gelombang folikel didefinisikan sebagai durasi pertumbuhan folikel mulai diameter sekitar 4 mm sampai folikel dominan kembali mengecil keukuran semula. Folikel yang mencapai ukuran diameter terbesar dari gelombang yang sama dianggap sebagai folikel dominan (FD), sedangkan folikel yang lebih kecil sebagai folikel subordinat (FS). Fase pertumbuhan folikel adalah periode folikel terdeteksi pada diameter 4 mm sampai hari saat diameter folikel tersebut tidak bertambah besar. Fase regresi dimulai pada saat folikel terdeteksi mengalami pengurangan diameter sampai folikel mulai sulit terdeteksi secara individual. Fase statis didefinisikan sebagai periode antara hari terakhir fase pertumbuhan dan hari pertama fase regresi. Data kemudian dikelompokkan berdasarkan pola jumlah gelombang folikel yang teramati dalam satu IOI, yaitu kelompok pola 3 dan 4 gelombang folikel dan diuji dengan Independent-Sample T Test. Repeatabilitas (nilai antara 0 sampai 1) didefinisikan sebagai proporsi ragam total yang dapat berkontribusi terhadap ragam individu, dihitung dengan rumus 2 individu / (2 individu + 2 galat) (Burns et al. 2005). Komponen dari nilai ragam untuk repeatabilitas diestimasi menggunakan ANOVA dengan faktor tunggal. Persentase terjadinya ovulasi folikel merupakan jumlah kejadian ovulasi pada ovarium kiri atau kanan dibagi dengan total kejadian ovulasi folikel yang teramati. Persentase terjadinya gelombang folikel pada ovarium merupakan jumlah gelombang folikel yang terjadi pada ovarium kanan atau kiri dibagi dengan jumlah total gelombang folikel yang teramati.
Hasil dan Pembahasan
24
2 gelombang yang teramati dalam penelitian ini. Hal ini berbeda dengan Adam et al. (2008) yang melaporkan bahwa pada sapi mayoritas sapi dewasa memiliki pola 2 gelombang folikel. Melia et al. (2014) juga melaporkan bahwa sapi PO menunjukkan pola 3 gelombang folikel pada 6 ekor sapi yang diamati. Pada penelitian lain Viana et al. (2000) melaporkan hanya 1 dari 15 ekor (6%) sapi Gyr (B. indicus) memiliki pola 2 gelombang folikel, sedangkan 60% dan 26.7% berturut-turut memiliki pola 3 dan 4 gelombang folikel.
Jumlah gelombang dalam satu siklus dilaporkan tidak dipengaruhi oleh bangsa sapi (Adam et al. 2008), namun Bó et al. (2003) melaporkan terdapat peningkatan pola 3 gelombang pada sapi dengan kualitas nutrisi yang buruk dan stress karena panas. Pada sapi perah tidak ada perbedaan terkait proporsi pola gelombang antara sapi muda dan dewasa (Wolfenson et al. 2004). Tetapi dalam penelitian lain Adam et al. (2008) menyatakan mayoritas sapi dara (65%) pada sapi Nellor (B. indicus) menunjukkan pola 3 gelombang sedangkan mayoritas sapi induk (83%) menunjukkan pola 2 gelombang. Selanjutnya Satheshkumar et al. (2015) melaporkan pola 3 gelombang lebih banyak terjadi pada musim dingin dibandingkan pola 2 gelombang, dan sebaliknya terjadi pada musim panas.
Pertumbuhan gelombang folikel pertama mulai terjadi hari ke -0,4+0,9 dan 1,4+1,1 relatif terhadap terjadinya ovulasi pada sapi yang memiliki pola gelombang 3 dan 4. Gelombang folikel berikutnya mulai tumbuh pada 8,1+1,5 dan 7,4+1,9 untuk pola gelombang 3 dan 4 (Tabel 3). Pola 3 gelombang memiliki durasi dominan yang paling panjang (11,6+1,5 hari) pada gelombang pertama dan lebih pendek pada pola 4 gelombang yaitu 10,0+2,9. Hasil ini sesuai dengan Adam et al. (2008) yang menyatakan terdapat korelasi yang tinggi antara durasi dominan dengan pola jumlah gelombangnya, yaitu semakin pendek durasi dominan akan menambah jumlah gelombang dalam satu IOI. Dalam penelitian lain Jaiswal et al. (2009) menyatakan durasi dominan pada gelombang pertama dalam IOI dapat digunakan untuk memprediksi pola jumlah gelombang.
Tabel 3. Karakteristik per gelombang folikel dominan pada sapi PO dengan pola 3 dan 4 gelombang folikel pada satu IOI.
Pengamatan Gelombang Folikel
I II III VI
Onset folikel (hari ke):
3 gelombang -0,4+0,9 8,1+1,5 15+2,1 -
4 gelombang 1,4+1,1 7,4+1,9 12,2+1,5 16,2+3,2 Durasi dominan (hari)
3 gelombang 11,6+1,5 11+2,3 6,8+1,3 -
4 gelombang 10+2,9 8,8+1,6 7,7+1,5 6,6+0,9
Diameter FD (mm)
3 gelombang 10,05+0,43 9,26+0,94 12,24+0,71a* - 4 gelombang 9,42+0,44 8,84+1,15 9,1+1,44b 12,30+0,22 Huruf superscript yang berbeda pada kolom dan kriteria yang sama, berbeda nyata pada P<0.05. *: Diameter folikel ovulatori
Diameter folikel dominan masing-masing gelombang folikel pada pola 3 dan 4 tidak berbeda nyata walaupun terdapat kecenderungan diameter folikel dominan semakin mengecil dengan bertambahnya jumlah gelombang dalam satu IOI (Tabel 3). Viana et al. (2000) menyatakan penurunan diameter folikel dominan dan durasi dominannya diasosiasikan dengan peningkatan jumlah gelombang dalam satu siklus berahi. Dalam penelitian lain Bó et al. (2003) melaporkan banyaknya jumlah sapi B. indicus yang memiliki pola 4 gelombang (16%) dibandingkan dengan B. taurus (0%) diduga merupakan konsekuensi diameter folikel dominan yang lebih kecil dan durasi dominan yang lebih pendek pada B. indicus.
26
estradiol pada gelombang pertama IOI meregulasi waktu munculnya reseptor oxytosin dalam endometrium yang akhirnya mengatur waktu produksi prostaglandin untuk memicu luteolisis. Sehingga ada dugaan bahwa ukuran diameter folikel dominan pada gelombang pertama IOI mempengaruhi pola jumlah gelombang folikel yang terbentuk (Adam et al. 2008, Jaiswal et al. 2009). Namun Boer et al. (2011) menyatakan mekanisme terbentuknya pola gelombang masih belum jelas dan diduga melibatkan regulasi pertumbuhan folikel yang lebih komplek.
Tabel 4. Karakteristik dinamika folikel dominan, folikel sub dominan, dan corpus luteum (CL) pada sapi PO yang memiliki 3 dan 4 gelombang folikel.
Karakteristik 3 gelombang 4 gelombang
Folikel Dominan
Jumlah IOI 7 5
Panjang IOI (hari) 22,0+1,6 22,8+2,9
Kecepatan pertumbuhan (mm/hari) 0,87+0,23 0,94+0,25
Fase statis (hari) 3,7+2,0 2,7+1,9
Kecepatan atresia (mm/hari) 0,83+0,21 O,91+0,31 Pertumbuhan folikel sampai ovulasi (hari) 7,6+0,6 7,4+1,1 Diameter folikel ovulatori (mm) 12,24+0,71 12,30+0,22
Folikel Sub Dominan
Diameter saat deviasi (mm) 6,25+0,37 6,44+0,44
Diameter maksimal (mm) 6,48+0,80 6,92+0,57
Kecepatan pertumbuhan (mm/hari) 0,83+0,25 0,72+0,33 Kecepatan atresia (mm/hari) 0,69+0,15 0,8+0,29
Korpus Luteum (CL)
Diameter CL hari ke-10 (mm) 18,94+0,47 19,44+0,87 Dalam penelitian ini, kecepatan pertumbuhan antara folikel dominan (FD) tidak berbeda nyata antara pola 3 dan 4 gelombang (0,87+0,23, 0,94+0,25 mm/hari) (Tabel 4). Demikian pula kecepatan pertumbuhan FD tidak berbeda nyata dengan folikel subdominannya (FS) sampai terjadi deviasi folikel. Hasil ini sesuai dengan Adam et al. (2008) yang menyatakan bahwa sekelompok folikel pada awal pertumbuhan folikel memiliki kecepatan pertumbuhan yang sama sampai terpilih satu folikel untuk terus tumbuh dan menjadi FD sedangkan yang lain mengalami atresia.