The Origin of Madura Cattle Based on Mitochondrial DNA
Teks penuh
Gambar


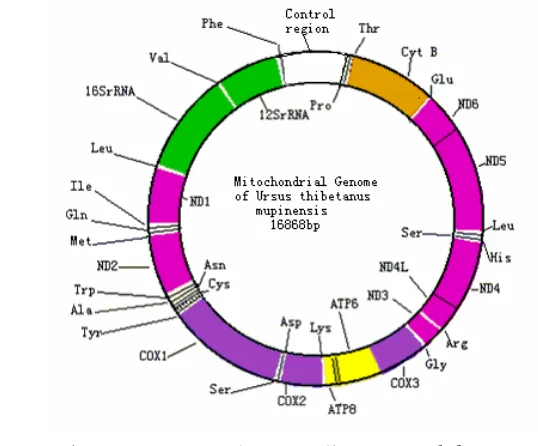
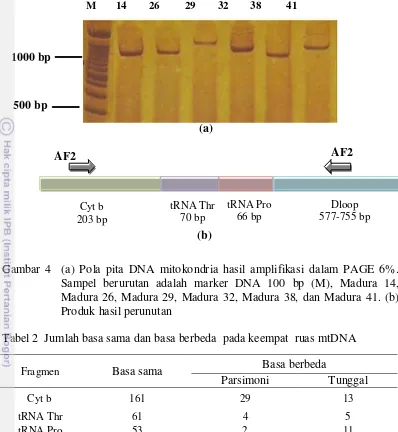
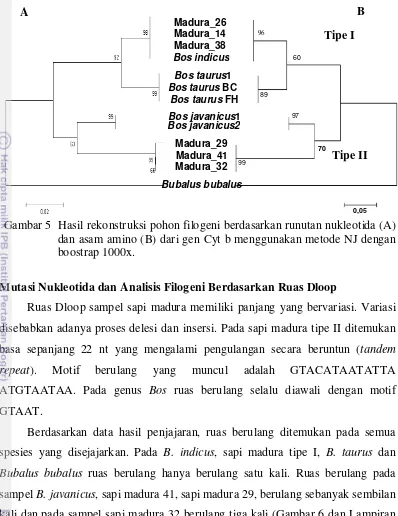



Dokumen terkait
Sampel darah sapi yang digunakan adalah sapi lokal Indonesia yaitu sapi Bali, Madura, Pesisir dan Katingan, sebanyak 80 sampel dengan masing-masing bangsa sapi
Sampel darah sapi yang digunakan adalah sapi lokal Indonesia yaitu sapi Bali, Madura, Pesisir dan Katingan, sebanyak 80 sampel dengan masing-masing bangsa sapi
Hasil analisis regresi berganda bobot hidup dan BCS terhadap total IB sapi Madura betina yang tidak disilangkan diperoleh koefisien determinasi 0,031, ini berarti 3,1% variasi
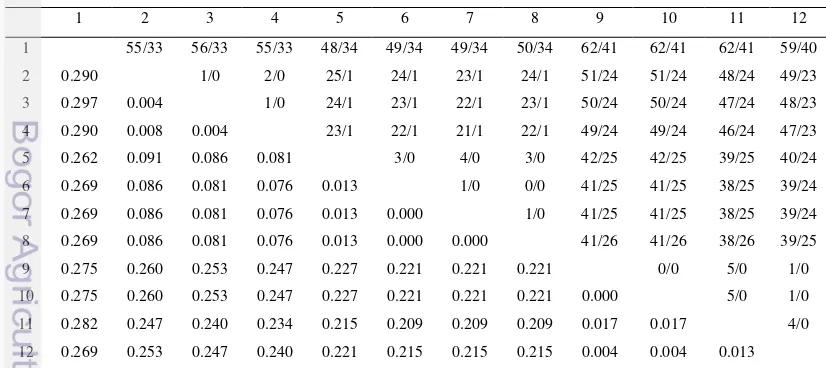
Sapi Madura, Jawa dan Peranakan Ongole diklasifikasikan sebagai Bos indicus, sedangkan sapi Bali sangat nyata terpisah dari dua kelompok sapi India maupun Eropa.. Analisis
Nilai Most Probable Producing Ability (MPPA) dapat digunakan sebagai dasar pedoman seleksi induk sapi Madura yang berkualitas berdasarkan berat sapih dan
Hasil keturunan memiliki sifat unggul sapi Madura yaitu adaptasi panas, adaptasi terhadap parasit, dan tahan terhadap pakan jelek serta memiliki sifat unggul sapi
Initially we compare cytochrome b sequences cyt b between nine Asian and five African elephants and use the fossil-based age of their separation ;5 million years ago to obtain a rate of
By applying HindIII restriction enzyme, the study showed that polymorphism genetic variation was found in ND-5 fragments of mitochondrial DNA in Bengali cattle Ongole Grade and Madura
