PERTUMBUHAN ISOLAT BAKTERI RUMEN PENCERNA
SERAT DAN KARAKTERISTIK FERMENTASINYA
DALAM MEDIA YANG DISUPLEMENTASI
MINERAL
SKRIPSI
RIZKINIA GUNARSIH
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNAKAN
i
RINGKASAN
RIZKINIA GUNARSIH. D24062825. 2010. Pertumbuhan Isolat Bakteri Rumen Pencerna Serat dan Karakteristik Fermentasinya dalam Media yang Disuplementasi Mineral. Skripsi. Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor.
Pembimbing Utama : Prof. Dr. Ir. Toto Toharmat, M. AgrSc. Pembimbing Anggota : Dr. Ir. Dwierra Evvyernie A, MS., MSc.
Mikroba rumen sangat penting dalam proses pencernaan pakan dan penyediaan protein pada ruminansia. Pertumbuhan mikroba dalam rumen sangat tergantung pada ketersediaan, baik jumlah maupun kualitas nutrien dalam pakan. Laju aktivitas selulolitik mikroba rumen ternak kerbau (43,2%/hari) lebih tinggi dibandingkan dengan ternak sapi (6,3%/hari). Ketersediaan mineral dalam pakan ruminansia sangat mempengaruhi pertumbuhan mikroba dan induk semangnya. Upaya meningkatkan produksi ternak sering dilakukan melalui peningkatan pertumbuhan dan aktifitas mikroba rumen melalui penambahan mineral. Penelitian ini bertujuan untuk menyeleksi isolat bakteri rumen kerbau yang mampu hidup dalam berbagai jenis media yang mengandung berbagai jenis mineral berkonsentrasi tinggi dan mengkaji kemampuan fermentabilitas isolat bakteri tersebut di dalam rumen secara in vitro dengan substrat serat yang berbeda.
Penelitian ini terdiri dari 3 percobaan. Percobaan 1 dilakukan pemilihan isolat bakteri berdasarkan produksi bahan kering sel bakteri dan data dari penelitian sebelumnya dari 12 isolat bakteri menjadi 6 isolat bakteri terpilih. Percobaan 2 yaitu pertumbuhan keenam isolat bakteri terpilih sebagai perlakuan di dalam calf starter bermineral Co, Cu, Zn, dan Mn tinggi dengan mengukur produksi bahan kering sel bakteri, dan pH. Percobaan 3 dilakukan evaluasi pengaruh penambahan mineral organik sebagai produk Isolat terhadap fermentabilitas In Vitro Jerami padi (JP) dan rumput gajah (RG) dengan 4 perlakuan (JP=jerami padi, JP+OR=jerami padi+mineral organik, JP+SU=jerami padi+susu, dan JP+MIX=jerami padi+premix), begitu pula dengan rumput gajah. Alat dan bahan yang digunakan dalam penelitian ini yaitu tabung reaksi, sumbat karet, isolasi panviks, label, cawan Conway, tabung fermentor, pipet, magnetic stirrer, tabung destilasi, labu Erlenmeyer, tabung hungate, botol film, gelas piala, autoclave, penangas air, termos, shaker water bath, oven 1050C, cawan porselen, eksikator, timbangan, tabung gas CO2, botol selai, buret, Co-klorida, Cu-sulfat, susu, pakan starter, 12 isolat bakteri, brain heart infusion (BHI), larutan McDougall, rumput gajah, jerami padi, cairan rumen, asam borat, larutan HgCl2, larutan Na2CO3 jenuh, larutan H2SO4, vaselin, dan gas CO2. Rancangan percobaan yang digunakan untuk ketiga percobaan ialah sama yaitu Rancangan Acak Lengkap (RAL) sebanyak 3 ulangan.
ii
sel bakteri berkisar 0,5–5,50 mg/ml. Lain halnya dengan pertumbuhan isolat bakteri pada media bermineral Co, isolat bakteri pencerna serat tersebut memiliki kemampuan yang berbeda (P<0.05) pada media BHI bermineral Cu. Kadar pH dalam media BHI menunjukkan hasil yang sama. Kisaran pH yang dihasilkan sebesar 5,00-6,87. Jumlah bakteri dalam media BHI menunjukkan hasil yang berbeda (P<0,05) pada mineral Co, Cu, dan Zn, sedangkan pada mineral Mn tidak berbeda nyata. Keenam isolat bakteri dalam calf starter memiliki respon yang tidak berbeda nyata (P>0,05) terhadap penambahan mineral Co, Cu, Zn, dan Mn tinggi dalam produksi BK sel isolat bakteri. Nilai pH dalam media calf starter perlakuan menunjukkan perbedaan yang nyata pada mineral Co. Kisaran pH dalam media calf starter berkisar antara 7,30-8,93. Jumlah bakteri dalam media calf starter menunjukkan hasil yang tidak berbeda nyata. Hal ini dapat dikarenakan keenam isolat bakteri memiliki kemampuan yang sama, baik dalam memanfaatkan nutrien pakan maupun daya adaptasi terhadap media tersebut.
Konsentrasi NH3 pada jerami padi baik yang diberikan tanpa penambahan mineral organik maupun dengan penambahan mineral organik didapatkan hasil yang berbeda (P<0,05). Jerami padi dengan penambahan mineral organik (JP+OR) menunjukkan nilai konsentrasi NH3 lebih tinggi dibandingkan dengan yang lain. Kisaran NH3 yang dihasilnya adalah 4-23 mM. Konsentrasi NH3 pada substrat rumput gajah menunjukkan hasil yang berbeda nyata setelah 30 menit diinkubasi. Konsentrasi NH3 pada rumput gajah dengan penambahan mineral organik memiliki nilai yang lebih besar. Kisaran konsentrasi NH3 yang dihasilkan adalah 2-27 mM. Konsentrasi VFA pada jerami padi dengan atau tanpa penambahan mineral organik menunjukkan hasil yang sama. Hal ini menggambarkan bahwa penambahan mineral organik pada substrat jerami padi tidak berpengaruh terhadap fermentabilitas pakan karena secara keseluruhan nilai VFA rendah. Kisaran nilai konsentrasi VFA yaitu 43-120 mM. Konsentrasi VFA pada rumput gajah menunjukkan hasil yang berbeda nyata setelah diinkubasi selama 4 jam. Kisaran VFA yang dihasilkan adalah 57-221 mM. Konsentrasi NH3 dan VFA rumput gajah lebih tinggi dibandingkan dengan jerami padi. Hal tersebut dapat disebabkan oleh kualitas rumput gajah yang lebih baik dibandingkan jerami padi. Isolat bakteri menunjukkan kemampuan adaptasi yang bervariasi terhadap kadar mineral Co, Cu, Zn, dan Mn yang tinggi. Isolat bakteri yang mampu tumbuh dengan baik dalam media BHI ialah isolat B, C, E, F, G, dan L. Keenam isolat bakteri terpilih lebih mampu tumbuh baik di dalam media susu bermineral tinggi daripada media calf starter bermineral tinggi. Isolat bakteri yang digunakan dapat berperan sebagai carrier mineral konsentrasi tinggi (Co, Cu, Zn, dan Mn). Penambahan mineral organik produk isolat berpotensi meningkatkan aktifitas mikroba rumen berdasarkan konsentrasi NH3 dan VFA yang diperoleh.
iii
ABSTRACT
Growth of Cellulolitic Rumen Bacteria Isolates and Their Fermentative Characteristics in Media Supplemented with Mineral
Gunarsih, R., T. Toharmat, and D. Evvyernie
Rumen microbes have important roles in digestion of feed components in ruminant. The activities of microbes can be increased by dietary minerals supplementation. However, the response of cellulolitic bacteria on high concentration of mineral and the role of cellulolitic bacteria in utilization of inorganic minerals is unclear. This experiment aimed to evaluate the capability of sellulolitic bacteria isolates to adapt in media containing high cobalt (Co), cuprum (Cu), zinc (Zn), or manganese (Mn) content. Three experiments were conducted: (1) Identification of adaptation capability of bacteria isolates in media containing 18.33 ppm Co; 5.39 ppm Cu; 18.38 ppm Zn; or 1176.5 ppm Mn; (2) Incubation of six bacteria isolates in milk or calf starter as medium containing 3.74 ppm Co, 1.09 ppm Cu; 3.68 ppm Zn; or 235.3 ppm Mn; (3) Evaluation of in vitro fermentability of rice straw and elephant grass in media contaning organic mineral produced from incubation of bacteria isolates in media supplemented Co, Cu, Zn, or Mn. The treatments were: (1) JP (rice straw), JP+OR (rice straw + organic minerals), JP+SU (rice straw + milk), and JP+MIX (rice straw + premix), (2) RG (elephant grass), RG+OR (elephant grass + organic minerals), RG+SU (elephant grass + milk), and RG+MIX (elephant grass + premix). Data observed were the dry cell of bacteria, pH, bacteria population, NH3 and volatile fatty acids concentration. The result showed that the bacteria isolates varied in their adaptability to media supplemented with Co, Cu, Zn, and Mn content. Milk was the best media to incubate sellulolitic bacteria isolates. Addition of organic mineral into the media of rice straw and elephant grass increased VFA and NH3 concentration. The conclusion was that the bacteria isolates were capable to adap to different concentration of minerals and convert them to organic minerals or bio-mineral that might increase rumen microbes activities and improved fermentability of feed incubated in the rumen liquid.
iv
PERTUMBUHAN ISOLAT BAKTERI RUMEN PENCERNA
SERAT DAN KARAKTERISTIK FERMENTASINYA
DALAM MEDIA YANG DISUPLEMENTASI
MINERAL
RIZKINIA GUNARSIH D24062825
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNAKAN
v
Judul : Pertumbuhan Isolat Bakteri Rumen Pencerna Serat dan Karakteristik Fermentasinya dalam Media yang Disuplementasi Mineral
Nama : Rizkinia Gunarsih
NIM : D24062825
Menyetujui,
Pembimbing Utama, Pembimbing Anggota,
(Prof. Dr. Ir. Toto Toharmat, M.AgrSc) (Dr. Ir. Dwierra Evvyernie A, MS., MSc.) NIP: 19590902 198303 1 003 NIP: 19610602 198603 2 001
Mengetahui: Ketua Departemen,
Ilmu Nutrisi dan Teknologi Pakan
(Dr. Ir. Idat Galih Permana, M.Sc.Agr.) NIP: 19670506 1991031 001
vi
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 18 Agustus 1988 dari pasangan Bapak Gunawan dan Ibu Rohmah. Penulis merupakan anak ketiga dari tiga bersaudara. Penulis mengawali pendidikan dasarnya di Sekolah Dasar Bekasi Jaya Indah 2 Bekasi Timur pada tahun 1994 dan diselesaikan pada tahun 2000. Pendidikan lanjutan pertama dimulai oleh penulis pada tahun 2000 dan diselesaikan pada tahun 2003 di Sekolah Lanjutan Tingkat Pertama Negeri (SLTPN) 1 Bekasi. Penulis kemudian melanjutkan ke Sekolah Menengah Atas (SMA) PGRI 1 Bekasi pada tahun 2003 dan lulus pada tahun 2006.
vii
KATA PENGANTAR
Alhamdulillahirabbil’aalamiin. Puji dan syukur penulis panjatkan kehadirat Allah SWT atas segala karunia dan rahmat-Nya sehingga penulisan skripsi ini dapat diselesaikan. Penelitian ini mengambil tema mengenai daya adaptasi mikroba rumen pencerna serat terhadap berbagai media yang berkadar mineral tinggi. Penelitian ini berjudul: Pertumbuhan Isolat Bakteri Rumen Pencerna Serat dan Karakteristik Fermentasinya dalam Media yang Disuplementasi Mineral. Skripsi ini ditulis berdasarkan hasil penelitian yang penulis lakukan mulai bulan Agustus - November 2009 bertempat di Laboratorium Ilmu Nutrisi Ternak Perah, Departemen Ilmu
Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor.
Tujuan dari penelitian ini adalah untuk menyeleksi isolat bakteri rumen kerbau yang mampu hidup dalam berbagai jenis media yang mengandung berbagai jenis mineral berkonsentrasi tinggi dan mengkaji kemampuan isolat bakteri tersebut di dalam memfermentasi komponen serat pakan in vitro. Isolat ini selanjutnya diharapkan dapat dijadikan kultur yang digunakan sebagai inokulan atau probiotik pada pedet atau ruminansia muda.
Penulis dengan rendah hati mengucapkan terima kasih kepada seluruh pihak yang telah membantu penulis sehingga penulisan skripsi ini dapat terselesaikan dengan baik. Semoga skripsi ini dapat bermanfaat khususnya bagi penulis sendiri dan untuk pembacanya.
Bogor, Agustus 2010
ix
Prosedur ... 16
Persiapan Sampel ... 16
Metode ... 16
Percobaan Tahap 1 ... 16
Pembuatan Media BHI Bermineral Tinggi ... 17
Peubah yang Diamati ... 17
Percobaan Tahap 2 ... 18
Pembuatan Media Calf Starter ... 18
Inokulasi Isolat Bakteri ... 18
Peubah yang Diamati ... 18
Percobaan Tahap 3 ... 19
Pembuatan Mineral Organik dengan Susu ... 19
Isolat Bakteri yang Digunakan ... 20
Pengukuran NH3 ... 20
Populasi Isolat Bakteri dalam Media BHI ... 24
Percobaan Tahap 2 ... 27
Produksi Bahan Kering Sel Bakteri dan Nilai pH Media Calf Starter ... 28
Populasi Isolat Bakteri dalam Media Calf Starter ... 28
x 4. Jumlah Bakteri Rumen (x 108/ml) pada Sapi dan Kerbau yang
Diberikan Pakan Berserat Tinggi ... 9 5. Produksi Bahan Kering (BK) Sel Isolat Bakteri pada Media
BHI yang Disuplementasi Mineral (mg/ml) ... 23 6. Nilai pH Media BHI yang Disuplementasi Mineral Setelah
Dinkubaskan Tiga Hari dengan 12 Isolat Bakteri ... 24 7. Jumlah Bakteri Berdasarkan Nilai Absorbansi (600 nm) dalam
Media BHI yang Disuplementasi Mineral ... 25 8. Populasi Bakteri Total (108 cfu/ml) dari Isolat Bakteri dalam
Media BHI yang Disuplementasi Mineral ... 26 9. Produksi Bahan Kering (BK) Sel Isolat Bakteri dalam Media
Calf Starter yang Disuplementasi Mineral (mg/ml) ... 27 10. Nilai pH dalam Media Calf Starter yang Disuplementasi
Mineral ... 28 11. Jumlah Bakteri dalam Media Calf Starter yang Disuplementasi
Berdasarkan Nilai Absorbansi (600 nm) ... 29 12. Populasi Bakteri Total (108 cfu/ml) dari Isolat Bakteri dalam
Media Calf Starter yang Disuplementasi Mineral ... 29 13. Konsentrasi NH3 pada Jerami Padi yang Difermentasi pada
Media Tanpa atau Ditambah Mineral Organik (mM) ... 31 14. Konsentrasi NH3 pada Rumput Gajah yang Difermentasi pada
Media Tanpa atau Ditambah Mineral Organik (mM) ... 31 15. Konsentrasi VFApada Jerami Padi yang Difermentasi pada
Media Tanpa atau Ditambah Mineral Organik (mM) ... 33 16. Konsentrasi VFA pada Rumput Gajah yang Difermentasi pada
Media Tanpa atau Ditambah Mineral Organik (mM) ... 33
xi DAFTAR GAMBAR
Nomor Halaman
xii DAFTAR LAMPIRAN
Nomor
1. ANOVA Kadar pH dalam Media Susu Bermineral Kobalt (Co) ... 44
2. Uji Kontras Ortogonal Kadar pH dalam Media Susu Bermineral Kobalt (Co) ... 44
3. Grafik Kadar pH dalam Media BHI ... 44
4. Grafik Produksi Bahan Kering Sel Isolat Bakteri dalam Media BHI 45
5. Grafik Jumlah Bakteri dalam Media BHI ... 45
6. Grafik Kadar pH dalam Media Calf Starter ... 45
7. Grafik Produksi Bahan Kering Sel Isolat Bakteri dalam Media Calf Starter ... ... 46
8. Jumlah Bakteri dalam Media Calf Starter ... 46
9. Grafik Konsentrasi NH3 pada Substrat Jerami Padi ... 46
10. Grafik Konsentrasi NH3 pada Substrat Rumput Gajah ... 47
11. Grafik Konsentrasi VFA pada Substrat Jerami Padi ... 47
12. Grafik Konsentrasi VFA pada Substrat Rumput Gajah ... 48
1
PENDAHULUAN
Latar Belakang
Mikroba rumen sangat penting dalam pencernaan pakan dan penyediaan
protein pada ruminansia. Pertumbuhan mikroba dalam rumen sangat tergantung pada
ketersediaan baik jumlah maupun kualitas pakan. Keberadaan mikroba khususnya
bakteri selulolitik dalam rumen memungkinkan ternak ruminansia mampu
memanfaatkan pakan berkadar serat kasar tinggi sebagai komponen utama pakannya.
Laju aktivitas selulolitik mikroba pada rumen ternak kerbau (43,2%/hari) lebih tinggi
dibandingkan dengan ternak sapi (6,3%/hari) (Suryahadi et al., 1996). Ketersediaan mineral termasuk salah satu faktor yang mempengaruhi pertumbuhan mikroba.
Upaya meningkatkan aktifitas rumen telah dilakukan dengan penambahan mineral
(Supriyati et al., 2000).
Unsur mineral merupakan salah satu komponen yang sangat dibutuhkan oleh
mahkluk hidup selain karbohidrat, protein, lemak, dan vitamin dalam tubuh. Mineral
berdasarkan jumlah penggunaannya terdiri dari dua jenis, yaitu mineral makro dan
mineral mikro. Mineral makro merupakan jenis mineral yang dapat digunakan dalam
jumlah besar, sedangkan mineral mikro merupakan mineral yang dapat digunakan
dalam jumlah sedikit tetapi memiliki peran yang penting baik dalam proses
pertumbuhan maupun metabolisme tubuh ternak. Pemberian mineral mikro yang
berlebih dapat menyebabkan keracunan pada ternak, sebaliknya apabila defisiensi
dalam tubuh ternak dapat mengakibatkan kelainan. Pada ternak ruminansia kelebihan
atau defisiensi nutrien pakan akan mempengaruhi perkembangan mikroba rumen.
Oleh sebab itu, tingkat pengaruh dan daya tahan mikroba rumen terhadap pemberian
mineral mikro yang mengandung konsentrasi tinggi sangat diperlukan.
Unsur kobalt (Co) merupakan mineral esensial untuk pertumbuhan hewan,
dan merupakan bagian dari molekul vitamin B12 (Davis dan Mertz 1987; Arifin,
2008). Vitamin B12 dibutuhkan untuk metabolisme propionat dan pertumbuhan
mikroorganisme (McDowell et al., 1993), serta berperan dalam meningkatkan
kecernaan selulosa secara in vitro (Allen, 1986; Kincaid et al., 2003).
Tembaga (Cu) merupakan salah satu mineral yang sering dilaporkan defisien
pada ternak ruminansia (Underwood, 1971). Hal ini dapat disebabkan oleh adanya
2
mineral tersebut. Seng (Zn) merupakan komponen penting pada struktur dan fungsi
membran sel, berperan sebagai antioksidan, melindungi tubuh dari serangan lipid
peroksidase, dan berperan dalam sintesis dan transkripsi protein, serta berperan
dalam menstabilkan struktur protein, seperti insulin, alkohol dehidrogenase hati,
alkalin fosfat, dan superoksida dismutase (Brown et al., 2002). Mangan (Mn) adalah
mineral mikro esensial bagi ternak, terdapat terutama di hati, kulit, otot dan tulang
(Anggorodi, 1994).
Suplementasi mineral sudah umum dilakukan oleh peternak. Mineral yang
digunakan sebagai suplemen umumnya dalam bentuk anorganik. Informasi mengenai
kemampuan mikroba rumen dalam mengkonversi mineral anorganik ke dalam
bentuk organik dan perannya dalam penyerapan mineral masih terbatas. Oleh sebab
itu, diperlukan adanya suatu kajian yang dapat menjelaskan peran bakteri selulolitik
dalam memanfaatkan mineral organik dan pengaruh mineral tersebut terhadap
aktivitas mikroba di dalam rumen.
Tujuan
Penelitian ini bertujuan untuk menyeleksi isolat bakteri rumen kerbau yang
mampu hidup dalam berbagai jenis media yang mengandung berbagai jenis mineral
berkonsentrasi tinggi dan mengkaji kemampuan isolat bakteri tersebut di dalam
3
TINJAUAN PUSTAKA
Mineral Cobalt (Co)
Mineral Co dengan bobot atom 58,9 termasuk dalam golongan VIII A pada
tabel periodik. Cobalt merupakan mineral mikro esensial dalam pakan ruminansia
untuk proses produksi vitamin B12 oleh mikroba rumen untuk memenuhi kebutuhan
bakteri pada ternak ruminansia dan induk semangnya (McDowell et al., 1993). Cobalt diserap dan diangkut melalui darah ke berbagai jaringan tubuh (Underwood
dan Suttle, 1999; Kincaid et al., 2003) yang dibutuhkan untuk metabolisme propionate dan pertumbuhan mikroorganisme (McDowell et al., 1993).
Pada ternak ruminansia, efisiensi produksi vitamin B12 (cobalamin) dari
mineral Co rendah, hanya sebesar 3 %; akan tetapi, efisiensi dapat meningkat sampai
13 % ketika konsumsi Co tinggi (Smith dan Marston, 1970; Kincaid et al., 2003). Liver mengandung Co dengan konsentrasi tinggi karena merupakan tempat
penyimpanan utama unsur tersebut (Underwood dan Suttle, 1999; Kincaid et al., 2003).
Mineral Tembaga (Cu)
Mineral Cu adalah salah satu mineral yang sering dilaporkan defisiensi pada
ternak ruminansia (Underwood, 1971). Defisiensi sekunder mineral mikro sering
dialami oleh ternak ruminansia walaupun ternak diberi suplemen mineral dalam
jumlah yang mencukupi kebutuhan (Kardaya et al., 2001). Di Australia, defisiensi Cu dikenal dengan nama “enzootic ataxia”, yang ditandai bahwa ternak mengalami
gangguan keseimbangan. Gangguan ini disebabkan pastura pada padang
penggembalaanya rendah kandungan Cu-nya, gejala kekurangan ini dapat dicegah
dengan pemberian garam Cu (McDowell et al., 1993). Sedangkan keracunan Cu yang kronis akan menyebabkan nekrosis pada sel hati, penyakit kuning, kehilangan
nafsu makan, dan kematian yang diawali dengan gejala koma (McDowell et al., 1993).
Fungsi biologis Cu antara lain berikatan dengan seruloplasmin dan dismutase
peroksida (Sutardi, 2001). Seruloplasmin berfungsi sebagai antioksidan, peredam
radikal bebas oksigen yang dihasilkan fagosit pada peradangan, sedangkan dismutase
4 menjadi peroksida hydrogen dan oksigen (Harmon dan Torre, 1997). Unsur Cu
diabsorbsi kurang baik oleh ruminansia (Kardaya, 2000). Hubungan Cu dengan
mineral essensial, antara lain bersifat sinergis terhadap unsur P, Fe, dan Co, serta
bersifat antagonistik terhadap unsur Ca, S, Zn, dan Mo (Georgievskii et al., 1982). Asosiasi Cu dengan protein atau asam amino dalam bentuk chelate atau kompleks
meningkatkan ketersediaan Cu (Kardaya et al., 2001). Kompleks-mineral organik makin diminati sebagai sumber mineral pada ternak karena potensi ketersediaan
biologisnya yang lebih tinggi daripada sumber mineral anorganik (Trihartanti, 2002).
Mineral Seng (Zn)
Unsur Zn mempunyai fungsi penting pada struktur dan membran sel, sebagai
antioksidan, dan melindungi tubuh dari serangan lipid peroksidase. Seng berperan
dalam regulasi gen terkait sintesis dan transkripsi protein. Pada suhu tinggi, hewan
banyak mengeluarkan keringat dan seng dapat hilang bersama keringat sehingga
perlu penambahan (Ahmed et al., 2002). Pemberian mineral Zn dapat memacu
pertumbuhan mikroba rumen (Muhtarudin dan Liman, 2006). Seng sebagai
metalloenzim yang melibatkan banyak enzim antara lain polimerase DNA, peptidase
karboksi A dan B dan posfatase alkalin. Aktivitas enzim-enzim tersebut akan
terganggu apabila terjadi defisiensi Zn. Di negara maju, suplementasi Zn dan Cu
digunakan untuk mengatasi mastitis pada sapi perah. Penyerapan Zn secara langsung
merupakan refleksi kebutuhan fisiologis akan Zn, baik pada anak maupun induk sapi
(Miller, 1970). Tingkat absorpsi Seng oleh ternak monogastrik dewasa adalah rendah
(7-15% dari konsumsi) berbanding terbalik dengan kandungannya dalam pakan.
Ternak ruminansia mengabsorbsi 20-40% (Tarmidi, 2008). Seng pakan dan pada
ternak muda absorpsinya lebih tinggi. Jika ruminansia muda mendapatkan ransum
dengan kandungan Zn yang sangat kurang, maka kadar Zn beberapa jaringan akan
turun, perubahan kadar jaringan tersebut berbeda-beda. Jika kekurangan sangat besar
maka akan terjadi penurunan Zn rambut, tulang, hati, paru-paru, ginjal, limpa,
pankreas dan plasma darah (Miller, 1970). McDonald et al. (1981) menyatakan bahwa kelebihan Zn kemungkinan dapat menyebabkan defisiensi mineral lain seperti
5
Mineral Mangan (Mn)
Mangan adalah zat mineral mikro esensial bagi unggas, terdapat terutama di
hati juga dialat-alat lainnya dan kulit, otot-otot serta tulang (Anggorodi, 1994).
Mangan diserap terutama di usus halus (Parakkasi, 1983) dan sebagian besar di
deodenum pada hewan monogastrik dan poligastrik (Georgievskii et al., 1982). Penyerapan mangan dari pakan adalah sangat rendah, rata-rata 2–5% dari intake pada
unggas, sedangkan pada ruminansia dewasa adalah 10–18%.
Mangan sulfat, mangan khlorida, mangan karbonat, kalium permanganat dan
mangan dioksida merupakan sumber mangan bagi ayam. Mangan sulfat merupakan
bentuk yang paling sering digunakan untuk melengkapi ransum unggas (Anggorodi,
1994).
Tabel 1. Dosis Maksimum dan Toksisitas Mineral untuk Mikroflora Rumen
Mineral Komponen Konsentrasi Maksimum
(µg/ml)
Sumber: Georgievskii et al. (1982)
Ransum Pemula (Calf Starter)
Pemberian hijauan atau konsentrat untuk pedet harus dilakukan secara
bertahap. Hal ini disebabkan adanya kebiasaan anak sapi yang lebih menyukai
makanan cair. Makanan padat yang diberikan pada pedet sering dikenal dengan
sebutan calf starter (ransum pemula). Ransum pemula yang diberikan biasanya berupa campuran dari berbagai jenis bahan pakan berenergi dan protein tinggi
(Parakkasi, 1999). Ransum pemula dapat terdiri dari jagung giling 38 65%, polar
28,98%, bungkil kedelai 28,98%, dan mineral mix 3,39% (Perdhanayuda, 2010).
Berdasarkan formulasi ransum tersebut, pedet sudah mendapatkan asupan pakan
yang mengandung energi dan protein tinggi serta mineral yang cukup. Ransum
6 dapat tumbuh baik dan sehat dibandingkan dengan pemberian pakan untuk ternak
dewasa.
Kualitas calf starter lebih baik dari kualitas konsentrat untuk sapi sapi dewasa dan dewasa. Konsentrat untuk sapi dewasa biasanya tersusun dari berbagai bahan
pakan biji–bijian dan hasil ikutan dari pengolahan hasil pertanian maupun industri.
Pemberian konsentrat dimaksudkan untuk mempercepat pertumbuhan sapi.
Pemberian pakan penguat berupa konsentrat harus memperhitungkan nilai
ekonomisnya. Pemberian konsentrat yang terlalu tinggi dapat menyebabkan kerugian
bila tidak diiringi peningkatan pertumbuhan yang sesuai (Parakkasi, 1999).
Kebutuhan pedet dengan bobot badan 100 kg (NRC, 2001), yaitu konsumsi
BK 3,1 kg/hari (2,5–3% BB); PK 16,3%; TDN 62,9%; Ca 25 g/hari dan P 11 g/hari.
Penelitian Veira et al. (1980), melaporkan kadar protein yang diberikan pada pedet adalah 9,9%-16,2%. Pemberian protein 9,9% menghasilkan keuntungan bobot hidup
sebesar 0,70 kg/hari dan pemberian protein 16,2% sebesar 1,22 kg/hari.
Susu
Susu segar adalah Susu adalah produk yang dihasilkan oleh glandula mamae
dan merupakan nutrisi bagi anaknya untuk mendapatkan imunitas pasif (Lestari, 2006).
Komponen terbesar dari susu adalah air dan sisanya terdiri dari lemak, dan bahan
kering tanpa lemak. Komponen lemak terbagi atas trigliserida dan komponen yang
larut dalam lemak. Bahan kering tanpa lemak terdiri atas vitamin B dan C, substansi
nitrogen, laktosa dan mineral. Selain itu, substansi nitrogen yang disebut pula
sebagai protein terbagi atas kasein, protein whey, dan nitrogen non protein. Susu
yang dihasilkan pada awal periode laktasi mempunyai kandungan bahan kering yang
tinggi. Susu diproduksi oleh glandula mammae dari kumpulan sel-sel epithelial sekretori
yang spesifik. Sel-sel ini membentuk struktur yang disebut alveoli. Sel-sel alveoli
dikelilingi oleh sel-sel kontraktil yang disebutt sel-sel myoepithelial (Lestari, 2006).
Sel-sel berkontraksi sebagai respon dari hormone yang dikeluarkan oleh kelenjar pituitary
7 Tabel 2. Komposisi Nutrien Susu Sapi
Komponen Kandungan (%)
Serat Kasar Pakan dan Perannya
Menurut Sofyan et al. (2000), berdasarkan analisis Van Soest, bahan pakan dapat digolongkan menjadi bahan bermanfaat yaitu komponen isi sel seperti gula,
pati, pektin, non protein nitrogen (NPN), protein, lemak, mineral dan vitamin, serta
bahan yang agak sulit dimanfaatkan yang berupa dinding sel. Menurut Sutardi
(1980), dinding sel dapat dibagi menjadi fraksi yang larut dan tidak larut. Fraksi yang
larut sebagian besar terdiri atas hemiselulosa dan sedikit protein dinding sel. Fraksi
yang tidak larut adalah lignoselulosa yang lazim disebut Acid Detergent Fiber
(ADF), dan dari ADF dapat diperoleh selulosa dan lignin.
Serat kasar adalah fraksi dari karbohidrat yang tidak larut dalam basa dan
asam encer setelah pendidihan masing–masing 30 menit (Sofyan et al., 2000). Kandungan serat kasar yang tinggi akan menghambat gerak laju digesta di dalam alat
pencernaan (Winugroho et al., 1983). Kandungan serat kasar tersebut menyebabkan daya cerna karbohidrat maupun nutrien lainnya menjadi turun (Parakkasi, 1999).
Menurut Tillman et al. (1982), hasil akhir dari proses pencernaan golongan hemiselulosa dan selulosa berupa asam asetat, asam propionate dan asam butirat.
Fungsi hemiselulosa dan selulosa dalam saluran pencernaan tidak spesifik, tetapi
penting dalam meningkatkan gerak peristaltik pada pencernaan hewan golongan non
ruminansia, juga merupakan sumber energi dari mikroorganisme dalam lambung dan
sebagai bahan pengisi lambung. Golongan lignin tidak dapat dicerna dan tidak
memiliki hasil akhir dari proses pencernaan serta keberadaannya dapat menghambat
8
Jerami Padi
Jerami padi merupakan bagian dari batang tumbuhan tanpa akar yang
tertinggal setelah dipanen butir buahnya (Shiddieqy, 2005). Jerami padi pada
umumnya masih dipandang sebagai limbah pertanian yang kurang berguna dibanding
hijauan berkualitas rendah lainnya karena adanya faktor pembatas yaitu rendahnya
kandungan nutrien serta koefisien cernanya. Hogan dan Leche (1981), melaporkan
bahwa komponen jerami padi yang dapat dicerna secara in vitro hanya 45–50%. Jerami padi memiliki kandungan nutrisi yang rendah. Komposisi nutrien jerami padi
dapat dilihat pada Tabel 3.
Tabel 3. Komposisi Nutrien Jerami Padi
Komponen Selly (1994) Hanafi (2008)
Bahan Kering (%) 89,41 71,2
Bahan Organik (%) 88,31 -
Serat Kasar (%) - 40,45
Lignin (%) 3,74 -
TDN (%) - 56,46
Silika (%) 20,49 -
Protein Kasar (%) 8,63 5,47
Rumput Gajah
Berdasarkan taksonominya, rumput gajah digolongkan ke dalam division
Spermatophita, subdivisio Angiospermae, kelas Monocotyledonea, ordo Glumifora,
family Gramineae, subfamili Panicodea, genus Pennisetum dan spesies Pennisetum purpureum. Rumput gajah mengandung protein kasar 8,44% (Sugiarto, 2002). Menurut Sofyan et al. (2000), rumput gajah umumnya mengandung bahan kering (BK) yang rendah yaitu 12–18%. Serat kasar berkisar dari 26–40,5%, bahan ekstrak
tanpa nitrogen (BETN) sekitar 30,4–49,8% dengan kandungan lemak kasar 1,0–
9
Mikroba Rumen
Mikroba rumen yang bersifat anaerob adalah penting dalam proses fermentasi
rumen, karena dapat melakukan berbagai reaksi dan interaksi dengan makanan yang
dikonsumsi ternak, untuk menghasilkan nutrien yang dapat diserap dan selanjutnya
dapat dimanfaatkan oleh tubuh ternak. Selain sifatnya yang anaerob, mikroba rumen
juga memerlukan kondisi pH 5,7-7,3 dan suhu 38-41oC (Hoover dan Miller, 1992).
Jenis mikroba penting yang menghuni rumen adalah bakteri, protozoa dan fungi
(Preston dan Leng, 1987). Bakteri merupakan penghuni terbesar dalam rumen yaitu
1010-1012/ml cairan rumen, sedangkan populasi protozoa 105-106/ml cairan rumen
(Ogimoto dan Imai, 1981).
Sutardi (1977) menyatakan bahwa adanya bakteri dan protozoa yang hidup
dalam rumen menyebabkan ruminansia dapat mencerna bahan pakan yang
mengandung serat kasar tinggi. Berbagai jenis mikroorganisme yang masing-masing
memiliki produk fermentasi antara dan produk fermentasi akhir yang
bermacam-macam menyebabkan kehidupan di dalam rumen menjadi kompleks. Interaksi yang
luas antara mikroorganisme di dalam rumen adalah interaksi yang bersifat
ketergantungan, saling menguntungkan dan kompetitif.
Tabel 4. Jumlah Bakteri Rumen (x 108/ml) pada Sapi dan Kerbau yang Diberi Pakan Berserat Tinggi
Tabel 4 memperlihatkan bahwa jumlah total bakteri pada kerbau (18,45 x
108/ml) lebih besar dibandingkan dengan jumlah total bakteri pada sapi (11,62 x
108/ml). Hal tersebut menunjukkan bahwa bakteri selulolitik 2-3 kali lipat lebih besar
pada kerbau dibandingkan sapi. Persentase bakteri selulolitik pada sapi sebesar
22,2% dan pada kerbau 37,2% dari total bakteri. Pada percobaan in vitro pada berbagai kondisi menunjukkan bahwa pemecahan selulosa terjadi lebih awal pada
10
Bakteri Selulolitik
Guedon (2002) menyatakan bahwa mikroorganisme selulolitik berperan
penting dalam biosfer. Beberapa spesies bakteri hidup pada kondisi temperatur,
tekanan dan pH yang ekstrim. Habitatnya termasuk tanah, rawa, sungai, danau,
sedimen air laut, kayu, kapas, lumpur, silase, kompos, bahan sayuran yang
membusuk, tempat sumber air panas dan tempat sumber asam maupun sumber
alkalin.
Menurut Beguin dan Aubert (1992), bakteri selulolitik juga terdapat dalam
usus herbivora vertebrata. Selain itu, bakteri selulolitik bersifat anaerob yang
bersimbiosis dalam menghancurkan makanan. Secara fisiologi bakteri selulolitik
dapat dibagi menjadi beberapa kelompok yaitu: (1) fermentasi anaerob, tipe gram
positif (Clostridium, Ruminococcus dan Caldicellulosiruptor) tetapi juga mengandung sedikit spesies gram negatif yang secara genetik masih berhubungan
dekat dengan keluarga Clostridium (Butyrivibrio dan Acetivibrio) dan yang tidak (Fibrobacter), (2) bakteri aerob gram positif (Cellulomonas dan Thermobifida) dan (3) bakteri aerob yang dapat bergerak (Cytophaga dan Sporocytophaga) (Lynd et al., 2002).
Pertumbuhan Bakteri
Pertumbuhan ialah pertambahan teratur semua komponen suatu
mikroorganisme. Pada waktu inkubasi setiap sel induk berbagi diri dengan
pembelahan biner dalam waktu 20-30 menit menjadi dua sel anak (Hadioetomo,
1985). Hobson (1988) menyatakan bahwa waktu penggandaan populasi bakteri
adalah 21-27 menit. Waktu generasi pada setiap bakteri tidak sama, ada yang hanya
memerlukan 20 menit bahkan ada yang memerlukan sampai berjam-jam atau
berhari-hari. Waktu generasi adalah waktu yang diperlukan oleh mikroorganisme
untuk meningkatkan jumlah sel menjadi dua kali lipat jumlah semula (Sumarsih,
2003).
Pertumbuhan umum digunakan untuk bakteri dan mikroorganisme lain dan
biasanya mengacu pada perubahan di dalam hasil panen sel (pertambahan total massa
sel) dan bukan perubahan individu organisme. Pertumbuhan merupakan pertambahan
jumlah atau massa melebihi yang ada di dalam inokulum asalnya. Selama fase
11 pertambahan komponen selular yang lain seperti DNA, RNA dan protein (Pelczar
dan Chan, 1986).
Pertumbuhan mikroba dapat diukur berdasarkan konsentrasi sel (jumlah sel
persatuan isi biakan) atau densitas sel (berat kering dari sel-sel persatuan sel biakan)
dan jumlah sel hidup biasanya dianggap sebagai ukuran konsentrasi sel. Pengukuran
jumlah bakteri dapat menggunakan teknik absorbsi cahaya. Absorbansi cahaya dari
suatu biakan dengan cara fotoelektris dan menghubungkan jumlah mikroba hidup
dengan ukuran-ukuran optik dalam suatu kurva standar, dengan kurva standar ini
semua hasil pengukuran optik dapat diubah menjadi konsentrasi sel. Bila bakteri
diinokulasikan ke dalam medium baru, pembiakan tidak segera terjadi tetapi ada
periode penyesuaian pada lingkungan yang dikenal dengan pertumbuhan adaptif.
Kemudian akan memperbanyak diri (replikasi) dengan laju yang konstan, sehingga
akan diperoleh kurva pertumbuhan. Pada kurva pertumbuhan dikenal beberapa fase
pertumbuhan yaitu dengan fase lag, fase eksponensial, fase stasioner dan fase kematian. Fase pertumbuhan bakteri secara umum dapat dilihat pada Gambar 1.
Gambar1. Kurva Pertumbuhan Bakteri (Cann, 2007)
Sumber : Cann (2007)
Selama fase lag terjadi peningkatan ukuran sel, pada waktu fase ini sel belum
12 menyesuaikan diri dalam medium baru. Fase eksponensial terjadi pertumbuhan
seimbang, sel membelah dengan kecepatan yang tetap dan maksimal. Pertumbuhan
yang paling cepat terjadi pada fase eksponensial ini. Fase stasioner ditandai dengan
penurunan kecepatan pertumbuhan (pembelahan bakteri berkurang), terjadi karena
penumpukan limbah metabolisme, racun, kekurangan nutrien, dan perubahan kondisi
pada lingkungan. Pertumbuhan sel yang hidup masih lebih banyak daripada jumlah
sel yang mati. Fase kematian ditandai dengan jumlah sel yang mati lebih banyak
daripada sel yang hidup karena nutrien semakin menurun (bahkan habis), energi
cadangan di dalam sel juga habis dan terkumpulnya produk limbah (Tarigan, 1988).
Kecernaan Pakan
Nilai kecernaan adalah persentase bahan pakan yang dapat dicerna dan
diserap oleh saluran pencernaan, jika dinyatakan dalam persen maka disebut
koefisien cerna. Kecernaan nutrien merupakan salah satu ukuran dalam menentukan
kualitas suatu bahan pakan. Kecernaan dapat diukur dengan teknik fermentasi in vitro (Tilley dan Terry, 1963).
Faktor–faktor yang berpengaruh terhadap nilai kecernaan yaitu pakan, ternak
dan lingkungan. Perlakuan terhadap pakan (pengolahan, penyimpanan, dan cara
pemberian), jenis, jumlah, dan komposisi pakan yang diberikan pada ternak. Umur
ternak, kemampuan mikroba rumen mencerna pakan, jenis hewan serta variasi hewan
turut menentukan nilai kecernaan. Kondisi lingkungan yang berpengaruh terhadap
nilai kecernaan adalah derajat keasaman (pH), suhu dan konsentrasi udara baik itu
secara aerob atau anaerob (Anggorodi, 1994).
Produksi Amonia
Protein mengalami hidrolisa menjadi oligopeptida oleh enzim proteolisis
yang dihasilkan oleh mikroba di dalam rumen yang selanjutnya akan didegradasi dan
menghasilkan amonia. Besarnya protein yang lolos dari degradasi rumen berkisar
20–80% (Sutardi, 1977). Amonia digunakan oleh bakteri untuk membentuk protein
tubuhnya selanjutnya protein mikroba akan dicerna dan diserap serta dikatabolisasi
dalam tubuh induk semang.
Produksi amonia rumen dipengaruhi oleh pH rumen, kelarutan bahan pakan,
13 Sutardi (1977), untuk memperkecil degradasi protein dalam rumen adalah sebagai
berikut: (1) pemberian air (mempercepat laju pergerakan isi rumen), (2) pemberian
garam (hewan haus sehingga banyak minum), (3) menurunkan daya larut protein, (4)
penggilingan sampai halus, lalu dijadikan pellet (meningkatkan laju pergerakan
digesta), (5) penambahan bahan kimia dan (6) pembungkusan protein dengan kapsul.
Produksi VFA (Volatile Fatty Acid)
Mikroba juga membutuhkan rantai karbon selain amonia untuk
pertumbuhannya dan ini dapat disuplai dari asam lemak terbang atau VFA yang
merupakan hasil fermentasi karbohidrat. Volatile Fatty Acid (VFA) merupakan sumber energi bagi ruminansia yang diproduksi bila karbohidrat ransum mengalami
fermentasi dalam rumen. Oleh sebab itu konsentrasi asam lemak terbang (VFA)
dalam cairan rumen dapat digunakan sebagai tolak ukur fermentabilitas pakan dan
sangat erat kaitannya dengan aktifitas dan populasi mikroba rumen (Hartati, 1998).
Sebagian besar VFA tersebut diserap langsung melalui dinding rumen; sebagian
kecil asetat dan propionat serta sebagian besar butirat termetabolisme dalam dinding
rumen (Parakkasi, 1999).
Laju pertumbuhan mikroba dalam rumen sangat tergantung kepada
ketersediaan karbohidrat. Laju pencernaan karbohidrat merupakan salah satu faktor
penentu produksi protein mikroba rumen. Selain sebagai sumber kerangka karbon,
karbohidrat adalah sumber energy untuk mikroba, dalam bentuk ATP (Adenosin Tri
Phosphate). Pertumbuhan mikroba rumen proporsional terhadap jumlah ATP yang
dihasilkan dari katabolisme sumber energy (Erwanto, 1995). Konsentrasi VFA dalam
rumen tergantung pada: 1) laju produksi dalam rumen; 2) penyerapan dalam rumen;
3) laju pengaliran dari rumen ke abomasum; 4) pelarutan oleh saliva; 5) laju
pengosongan rumen dan 6) perubahan menjadi metabolit lain oleh mikroba rumen
(Sutardi, 1977).
Peran Suplementasi
Suplementasi dapat dipandang sebagai langkah yang strategis dalam
mengatasi permasalahan nutrisi ternak, karena selain akan mampu mengatasi
masalah defisiensi juga akan dapat meningkatkan kapasitas mencerna dari hewan,
14 bila dirancang dengan baik, suplementasi lebih mudah diterapkan dibandingkan
dengan cara–cara pengolahan pakan lainnya, karena tidak membutuhkan tambahan
waktu kerja dan beban energi ekstra bagi petani (Suryahadi et al., 2002).
Salah satu persyaratan agar suplementasi tersebut dapat berhasil adalah
adanya informasi tentang: (1) status mineral ternak yang dapat diduga melalui kadar
mineral dalam pakannya dan pada organ tubuhnya (plasma darah). Informasi ini
telah dapat diperoleh atas dasar kajian terdahulu (Suryahadi, 1990) dan (2) tingkat
kebutuhan mineral. Dari berbagai penelitian terdahulu diperkirakan bahwa kebutuhan
mineral ternak sapi perah di Indonesia adalah berkisar antara 1,5–2,0 kali dari yang
15
MATERI DAN METODE
Lokasi dan Waktu
Penelitian dilaksanakan dari bulan Agustus sampai dengan November 2009.
Semua kegiatan dikonsentrasikan di Laboratorium Nutrisi Ternak Perah dan
Laboratorium Biokimia, Fisiologi dan Mikrobiologi Nutrisi, Departemen Ilmu
Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor.
Materi
Alat
Alat yang digunakan pada penelitian adalah sentrifuge, cawan Conway,
penangas air, gelas piala, eksikator, botol selai, label, magnetic stirrer, botol film, tabung destilasi, tabung gas CO2, oven dengan suhu 1050C, oven 600C, tabung
fermentor, gelas ukur, labu Erlenmeyer, tabung reaksi, tutup karet, isolasi panfiks,
plastik tahan panas, spoit, buret, pipet mikro, pipet volumetrik, bulp, pH meter,
sprayer, timbangan digital, autoclave, shaker bath, vortex, spektrofotometer UV 200 RS, sarung tangan dan alumunium foil.
Bahan
Bahan yang digunakan pada penelitian antara lain: cairan rumen, jerami padi,
rumput gajah, aquades, medium BHI (brain heart infusion), glukosa, celubiosa,
cystein-HCl, resazurin, hemin, aquadest, larutan McDougall, asam borat, larutan
HgCl2, larutan Na2CO3 jenuh, larutan H2SO4, susu steril, calf starter, 12 isolat
bakteri, vaselin, gas CO2, dan 4 jenis sumber mineral (CoCl2.6H2O, CuSO4. 5H2O,
ZnSO4. 7H2O, MnSO4. H2O).
Rancangan Percobaan
Rancangan Percobaan
Percobaan 1 dalam penelitian ini menggunakan rancangan acak lengkap
(RAL) dengan 3 ulangan, sedangkan percobaan 2 dan 3 menggunakan 2 ulangan.
Model matematik yang digunakan dalam analisa statistik adalah:
16 Keterangan: Yij = Nilai pengamatan pada ulangan ke-j dan perlakuan ke-i, µ = Nilai
rataan umum, τi = Pengaruh perlakuan ke-i, εij = Error perlakuan ke-i
dan ulangan ke-j.
Data yang diperoleh dianalisis ragam (Analysis of Variance, ANOVA)
berdasarkan Steel dan Torrie (1993). Selanjutnya, jika perlakuan berbeda nyata
maka dilakukan uji kontras ortogonal.
Prosedur
Penelitian ini dilakukan dalam tiga tahap percobaan. Tahap pertama
dilakukan pemilihan isolat bakteri dari 12 isolat bakteri menjadi 6 isolat bakteri
berdasarkan produksi bahan kering (BK) sel isolat bakteri, nilai CMC-ase, dan
jumlah bakteri yang dihasilkan pada penelitian sebelumnya (Astuti, 2010). Tahap
kedua dilakukan kajian pertumbuhan 6 isolat bakteri terpilih di dalam media calf starter yang disuplementasi mineral (Co, Cu, Zn, dan Mn) berkonsentrasi tinggi dengan peubah yang diukur adalah produksi bahan kering (BK) sel isolat bakteri,
nilai pH, dan jumlah bakteri. Penelitian tahap ketiga dilakukan untuk menguji
fermentabilitas in vitro menggunakan konsorsium 6 isolat bakteri terpilih dalam media terbaik hasil pengujian tahap kedua.
Persiapan Sampel
Pembuatan larutan mineral Co, Cu, Zn, atau Mn dilakukan pertama kali
sebelum pembuatan media BHI. Konsentrasi mineral dalam media diperhitungkan
agar mencapai 75% dari batas toksik (Georgievskii et al., 1982) untuk pertumbuhan bakteri rumen. Perhitungan dan penimbangan dilakukan dengan mempetimbangkan
jenis sumber mineral, memperhitungkan bobot molekul dan bobot atom
masing-masing mineral. Senyawa sumber mineral tersebut dilarutkan dalam 1 liter aquades
untuk dijadikan larutan stok mineral.
Metode
Percobaan Tahap 1: Kajian Adatasi Bakteri Terhadap Suplementasi Mineral
Penelitian ini bertujuan menseleksi 12 isolat bakteri rumen koleksi
Laboratorium Nutrisi Ternak Perah, Departemen Ilmu Nutrisi dan Teknologi Pakan,
17 ke dalam tabung reaksi yang berisi media BHI yang telah tercampur dengan salah
satu mineral (Co, Cu, Zn, atau Mn). Setiap kombinasi perlakuan isolat dengan jenis
mineral dilakukan dalam 3 ulangan. Isi tabung yang mengandung BHI tersebut telah
dikondisikan anaerob. Isolat bakteri dalam tabung diinkubasikan selama 3 hari. Pada
akhir inkubasi jumlah produksi bahan kering (BK) sel dan jumlah bakteri dari semua
isolat yang telah ditumbuhkan di dalam media BHI diukur.
Data produksi bahan kering sel bakteri digunakan untuk mengetahui daya
adaptasi isolat bakteri terhadap kadar beberapa mineral di dalam media BHI.
Berdasarkan data tersebut dan data CMC-ase dari penelitian sebelumnya (Astuti,
2010) dipilih sebanyak 6 isolat bakteri untuk ditumbuhkan dalam media calf starter
(kajian kedua).
Pembuatan Media BHI Bermineral Tinggi
Media BHI yang digunakan dalam kajian tahap ini mengandung Co, Cu, Zn
dan Mn tinggi. Bahan yang digunakan dalam pembuatan media BHI adalah BHI 7,4
g; cystein HCl 0,1 g; pati 0,1 g; glukosa 0,1 g; selebiosa 0,1 g; resazurin 1 ml; dan
hemin 0,6 ml. Semua bahan dilarutkan menjadi 200 ml menggunakan aquades yang
telah mengandung Co 18,33 ppm atau 75% dari kadar toksik Co. Sebanyak 200 ml
larutan BHI dibuat dengan prosedur yang sama untuk mendapatkan larutan yang
mengandung Cu 5,39 ppm atau 75% dari kadar toksik Cu, begitu pula dengan
mineral Zn dan Mn yaitu dengan konsentrasi masing-masing 18,38 ppm dan 1176,5
ppm. Pelarutan komponen penyusun BHI dilakukan dengan pemanasan. Setiap
larutan kemudian dialiri dengan gas CO2 selama 20 menit dan larutan dipipet 5 ml
untuk dimasukkan ke dalam tabung reaksi. Jumlah tabung reaksi yang digunakan
adalah 36 buah untuk setiap perlakuan mineral. Seluruh tabung reaksi ditutup
menggunakan sumbat karet dan disterilisasi dengan menggunakan autoclave dengan
suhu 1210C selama 15 menit.
Peubah yang Diamati
Peubah yang diamati pada penelitian ini yaitu: produksi bahan kering (BK)
18
Percobaan Tahap 2: Adaptasi Isolat Bakteri Terpilih dalam Media Calf Starter
Percobaan tahap kedua dilakukan dengan menumbuhkan 6 isolat bakteri
terpilih di dalam media calf starter. Setiap isolat bakteri dari enam isolat terpilih disuntikan ke dalam tabung reaksi yang yang telah berisi calf starter yang telah tercampur dengan salah satu mineral (Co, Cu, Zn, atau Mn). Setiap kombinasi jenis
isolat bakteri dan mineral dilakukan dalam 2 ulangan. Media yang telah diinokulasi
isolat bakteri tersebut diinkubasi selama 3 hari kemudian. Peubah yang diukur adalah
nilai pH media, produksi BK sel isolat bakteri, dan jumlah bakteri.
Pembuatan Media Calf Starter
Calf starter ditimbang sebesar 0,05 g dan dimasukan ke dalam tabung rekasi. Bahan pakan dalam tabung reaksi tersebut ditambah larutan McDougall sebanyak 5
ml dan larutan mineral Co, Cu, Zn, atau Mn sehingga konsentrasi masing-masing
mineral mencapai 3,74 ppm (Co); 1,09 ppm (Cu); 3,68 ppm (Zn); atau 235,3 ppm
(Mn). Jumlah tabung reaksi yang berisi calf starter yang disuplementasi mineral Co, Cu, Zn, atau Mn yang digunakan untuk setiap isolat masing-masing 12 tabung.
Sebelum isolat dimasukan ke dalam tabung, tabung reaksi bersama media di
dalamnya disterilisasi dengan menggunakan autoclave pada suhu 1210C selama 20 menit.
Inokulasi Isolat Bakteri ke dalam Media Calf Starter Bermineral
Biakan bakteri yang dihasilkan dari penelitian sebelumnya atau penelitian
tahap 1, diambil sebanyak 0,1 ml dari stok bakteri dengan menggunakan spoit, lalu
bakteri tersebut disuntikan ke dalam masing-masing tabung reaksi yang berisi media
calf starter bermineral tinggi. Jumlah dan teknik memasukan isolat bakteri ke dalam tabung reaksi yang berisi media calf starter, sama seperti yang dilakukan pada media BHI. Setelah itu, media yang sudah mengandung calf starter bermineral tinggi diinokulasi isolat bakteri diinkubasi selama 3 hari dalam shaker water bath pada suhu 390C.
Peubah yang Diamati
Peubah yang diamati pada penelitian ini yaitu: pH media, produksi bahan
kering (BK) sel isolat bakteri (mg/ml), dan jumlah bakteri (cfu/ml). Pengukuran pH
19 diangkat dari shaker water bath untuk diukur nilai pHnya. Masing-masing tabung reaksi diambil sebesar 1 ml larutan untuk diukur besarnya pH dengan menggunakan
alat pH meter. Pengukuran BK sel bakteri adalah sebagai berikut: bahan kering sel
bakteri diukur dengan cara media yang sudah tercampur bakteri diambil sebanyak 1
ml lalu dimasukkan ke dalam tabung eppendorf untuk disentrifuse selama 10 menit dengan kecepatan 7000 rpm. Setelah itu, supernatan dibuang dan endapan bersama
wadahnya dimasukkan ke dalam oven 600C selama 1 hari, kemudian dimasukkan ke
dalam oven 1050C. Perhitungan populasi bakteri dilakukan dengan mengamati
pertumbuhan bakteri dengan mengukur optical density (OD) media menggunakan spektrofotometer pada panjang gelombang 600 nm.
Percobaan Tahap 3: Kajian Fermentabilitas Jerami Padi dan Rumput Gajah in vitro menggunakan Konsorsium Enam Isolat Bakteri Terpilih.
Perlakuan dalam percobaan 3 ini terdiri atas 4 perlakuan dengan 2 ulangan
dimana pada percobaan dengan penambahan mineral organik diberikan penambahan
6 isolat bakteri yang terpilih pada percobaan 2. Adapun perlakuan tersebut adalah
sebagai berikut: (1) Media Rumput Gajah dengan perlakuan sebagai berikut: RG+OR
= Rumput Gajah + Mineral Organik; RG+SU = Rumput Gajah + Susu; RG =
Rumput Gajah; RG+MIX = Rumput Gajah + Premix. Mineral organik mengandung
Co (0,00098 ppm), Cu (0,097 ppm), Zn (0,396 ppm), Mn (0,393 ppm), Cr (10 ppm),
susu dan bakteri. (2) Media Jerami Padi dengan perlakuan sebagai berikut: JP+OR =
Jerami Padi + Mineral Organik; JP+SU = Jerami Padi+ Susu; JP = Jerami Padi;
JP+MIX = Jerami Padi + Premix. Mineral organik mengandung Co (0,00098
ppm), Cu (0,097 ppm), Zn (0,396 ppm), Mn (0,393 ppm), Cr (10 ppm), susu dan
bakteri.
Pembuatan Mineral Organik dengan Media Susu
Empat buah botol 250 ml diisi susu masing-masing sebanyak 200 ml.
Kemudian ke dalam susu ditambahkan 2 ml larutan mineral Co, Cu, Zn, atau Mn
sehingga masing-masing media susu mengandung 3,74 ppm (Co); 1,09 ppm (Cu);
3,68 ppm (Zn), atau 235,3 ppm (Mn). Media susu bermineral dalam botol
20 media susu bermineral tersebut diinokulasi dengan campuran 6 isolat bakteri yang
selanjutnya diinkubasi selama 3 hari.
Isolat Bakteri yang Digunakan
Isolat bakteri yang digunakan pada tahap ini ialah campuran keenam isolat
bakteri (B, C, E, F, G, dan L) yang ditumbuhkan dalam media susu. Hal tersebut
disebabkan oleh kemampuan hidup isolat bakteri dalam media susu lebih baik
dibandingkan dengan media yang lain.
Pengukuran NH3
Konsentrasi NH3 diukur dengan menggunakan teknik Mikrodifusi Conway
(Conway, 1958). Bibir cawan Conway dan tutupnya diolesi dengan vaselin,
kemudian supernatan yang dihasilkan dari pencernaan fermentatif diambil sebanyak
1 ml dan ditempatkan pada salah satu ruang sekat cawan dan larutan Na2CO3 jenuh
ditempatkan pada ruang sekat yang lain. Larutan asam borat sebanyak 1 ml
berindikator ditempatkan dalam cawan kecil yang terletak di tengah cawan Conway.
Selanjutnya cawan Conway ditutup rapat agar udara tidak dapat masuk. Supernatan
dan larutan Na2CO3 jenuh dicampur hingga merata dengan cara
menggoyang-goyangkan cawan dan memiringkannya. Setelah itu, cawan dibiarkan selama 24 jam
pada suhu kamar, dan setelah 24 jam cawan dibuka. Pada bagian asam borat
selanjutnya dititrasi dengan larutan H2SO4 0,005 N sampai terjadi perubahan warna
biru ke warna asam borat (merah jambu). Konsentrasi NH3 dihitung dengan rumus :
N NH3 (mM) = ml H2SO4 x N H2SO4 x 1000
g sampel x BK sampel
Pengukuran Volatile Fatty Acid (VFA)
Konsentrasi VFA diukur dengan menggunakan teknik destilasi uap (Steam
destilation) (General Laboratory Procedure, 1966). Lima mililiter supernatan (berasal
dari tabung yang sama dengan supernatan untuk analisa NH3) dimasukkan ke dalam
tabung destilasi, lalu ditambahkan 1 ml H2SO4 15%. Dinding tabung dibilas dengan
aquadest dan secepatnya ditutup dengan sumbat karet yang telah dihubungkan
dengan pipa destilasi berdiameter ±0,5 cm. Kemudian ujung pipa yang lain
21 labu didih yang telah berisi air mendidih tanpa menyentuh permukaan air tersebut.
Uap air panas akan mendesak VFA dan akan terkondensasi di dalam pendingin.
Hasil destilasi ditampung dengan labu Erlenmeyer 500 ml yang telah diisi 5 ml
NaOH 0,5 N. Proses destilasi selesai pada saat jumlah destilat yang tertampung
mencapai 300 ml. Destilat yang tertampung ditambah indikator phenolphtalein (PP)
sebanyak 2-3 tetes, lalu dititrasi dengan HCl 0,5 N sampai terjadi perubahan dari
warna merah jambu menjadi tidak berwarna (bening). Perhitungan Produksi VFA
total adalah sebagai berikut:
VFA total = (volume titran blanko-volume titran sampel) x N HCl x 1000/5 mM
22
HASIL DAN PEMBAHASAN
Percobaan Tahap 1: Kajian Adaptasi Isolat Bakteri Terhadap Suplementasi Mineral
Produksi Bahan Kering Sel Bakteri dan Perubahan Nilai pH Media
Tabel 5 menunjukkan variasi produksi BK sel bakteri dalam media yang
disuplemntasi Co, Cu, Zn, dan Mn. Produksi BK sel pada media BHI yang
disuplementasi Co, Zn, dan Mn tidak berbeda nyata antar isolat bakteri dengan nilai
rataan sebesar 1,66; 2,30; dan 2,42 mg/ml. Hal ini menunjukkan bahwa semua isolat
bakteri yang berjumlah 12 mempunyai kemampuan yang sama, baik dalam
memanfaatkan nutrien pakan maupun daya adaptasi terhadap media BHI yang
disuplementasi Co, Cu, Zn, dan Mn. Produksi BK sel bakteri berkisar 0,50–5,50
mg/ml. Lain halnya dengan pertumbuhan isolat bakteri pada media bermineral Co,
Zn, dan Mn, isolat bakteri pencerna serat tersebut memiliki kemampuan yang
berbeda (P<0,05) pada media BHI bermineral Cu. Hal tersebut menunjukkan bahwa
di dalam media bermineral Cu isolat bakteri mempunyai kemampuan adaptasi yang
berbeda dalam memanfaatkan nutrien didalamnya.
Penetapan isolat bakteri yang digunakan dalam kajian berikutnya tidak hanya
mempertimbangkan nilai rataan BK sel (Tabel 5), tetapi juga mempertimbangkan
hasil penelitian pada kajian isolat yang sama dengan mengukur aktifitas CMC-ase
dan jumlah bakteri (Astuti, 2010). Isolat bakteri yang dianggap paling adaptif
terhadap penambahan mineral dalam media dan terpilih adalah B, C, E, F, G dan L.
Keenam isolat bakteri dapat memanfaatkan nutrien dalam media sehingga dapat
tumbuh dengan baik walaupun kadar Co, Cu, Zn, dan Mn dalam media tersebut
ditingkatkan. Selain itu berdasarkan kajian CMC-ase, bakteri tersebut dapat hidup
lebih baik dalam media dengan serat kasar tinggi.
Hasil kajian produksi bahan kering sel bakteri menunjukkan bahwa isolat
bakteri mampu memanfaatkan media dengan baik walaupun konsentrasi Co, Cu, Zn,
dan Mn ditingkatkan. Hal ini menunjukkan bahwa unsur Co, Cu, Zn, dan Mn yang
ditambahkan ke dalam media tumbuh tidak menghentikan metabolisme dan
pertumbuhan bakteri. Pengaruh penambahan kadar Co, Cu, Zn, dan Mn terhadap
pertumbuhan bervariasi antar isolat. Pengaruh Cu diperkirakan lebih besar dari pada
23 Tabel 5. Produksi Bahan Kering (BK) Sel Isolat Bakteri pada Media BHI yang
Disuplementasi Mineral (mg/ml)
Isolat Mineral Rataan
Co Cu Zn Mn
A 0,50±0,71 2,45±1,20b 0,70±0,99 2,65±0,07 1,58±0,74
B 2,25±0,78 0,75±1,06d 0,50±0,71 5,75±6,01 2,31±8,56
C 0,65±0,92 3,60±0,14a 2,95±0,64 2,35±1,34 2,39±0,76
D 2,15±0,92 2,35±1,06b 1,95±0,78 0,90±0,00 1,84±0,69
E 1,40±1,98 3,75±0,21a 2,45±0,21 2,00±0,71 2,40±0,78
F 3,00±0,85 0,95±0,35d 2,80±0,57 1,95±0,92 2,18±0,67
G 2,10±1,70 0,70±0,14d 1,85±0,21 2,20±2,69 1,71±1,19
H 2,05±0,35 1,75±0,21c 2,25±1,63 2,60±1,56 2,16±0,94
I 1,40±0,71 3,40±0,71a 0,95±0,64 2,25±0,07 2,00±0,53
J 2,00±0,99 2,25±0,64c 2,95±1,20 2,35±0,64 2,39±0,87
K 0,75±1,06 2,90±0,00b 2,85±0,07 1,65±0,92 2,04±0,51
L 1,65±1,91 2,25±0,07c 5,50±5,94 2,40±0,85 2,95±2,19
Rataan 1,66±1,07 2,26± 0,48 2,30±1,13 2,42±1,96
Keterangan: A,B,C,….,L = kode isolat bakteri; Nilai dengan superskrip yang berbeda pada setiap kolom menunjukkan perbedaan (P<0,05)
Mikroorganisme selulolitik berperan penting dalam biosfer. Beberapa spesies
bakteri hidup pada kondisi temperatur, tekanan dan pH yang ekstrim (Guedon et al., 2002). Dalam penelitian ini, dilakukan pengukuran pH pada setiap media yang telah
diinokulasi isolat bakteri dan berkadar mineral tinggi setelah proses inkubasi selama
3 hari. Nilai pH media cukup bervariasi, namun masih berada dalam kisaran pH yang
dapat ditolerir bakteri rumen. Hal ini menunjukkan bahwa isolat bakteri tersebut aktif
memanfaatkan nutrien media tersebut dan menghasilkan asam yang menurunkan pH
media. Keseragaman nilai pH menunjukkan bahwa bakteri yang dikaji mempunyai
24 Tabel 6. Nilai pH Media BHI yang Disuplementasi Mineral Setelah Dinkubasikan
tiga hari dengan 12 Isolat Bakteri
Isolat Mineral Rataan
Co Cu Zn Mn
Keterangan : A,B,C,…,L = kode isolat bakteri
Berdasarkan nilai pada Tabel 6, terlihat bahwa nilai pH dalam media BHI
tidak memiliki perbedaan yang nyata. Kisaran pH yang dihasilkan adalah sebesar
5,00-6,87. Menurut Sanchez et al. (2007), jenis pakan dalam fermentasi mikroba rumen mempengaruhi besarnya pH tersebut yaitu sebesar 5,5-6,4. Hal tersebut
menunjukkan bahwa 12 isolat bakteri yang digunakan mampu memanfaatkan media
dan memproduksi asam yang menurunkan pH media meskipun media telah
disuplementasi mineral dan berhenti pada tingkat produksi asam tertentu sehingga
kondisi pH media tidak menurun lebih jauh.
Populasi Isolat Bakteri dalam Media
Pertumbuhan mikroba rumen merupakan fungsi dari pemanfaatan jumlah
nutrien dan senyawa yang dihasilkannya dalam rumen. Produksi nutrien dan energi
dalam cairan rumen sangat tergantung pada interaksi yang kompleks antara substrat
yang difermentasi dan jenis mikroorganisme yang terlibat. Keseluruhan hasil
fermentasi dalam rumen merupakan wujud saling ketergantungan diantara
25 media ditandai dengan timbulnya kekeruhan (Wulandari et al., 2005). Pertumbuhan bakteri dapat diamati melalui pengukuran dengan turbidimeter, dimana pertumbuhan
bakteri yang dibiakkan sebanding dengan tingkat kekeruhan (Suyasa, 2007).
Pengukuran populasi bakteri dalam penelitian ini menggunakan metode
turbidimetri. Metode ini berlandaskan pada kenyataan bahwa suatu populasi sel
dalam medium cair akan menahan cahaya yang sebanding dengan total masanya atau
konsentrasi sel dalam biakan. Kekeruhan biakan bakteri dalam penggunaan
turbidimetri dikorelasikan dengan beberapa metode penentuan lain seperti penentuan
jumlah mikroba dengan metode penaburan. Setiap pengenceran yang telah diukur
jumlah OD nya dapat dihitung jumlah mikrobanya masing-masing berdasarkan
jumlah mikroba yang telah diperoleh dari metode penaburan tersebut (Muchtadi dan
Laksmi, 1980).
Tabel 7. Jumlah Bakteri Berdasarkan Nilai Absorbansi (600 nm) dalam Media BHI yang Disuplementasi Mineral
Isolat Mineral Rataan
Co Cu Zn Mn
A 0,311±0,064c 0,372±0,275b 0,331±0,129c 0,351±0,271 0,341±0,185
B 0,293±0,040c 0,298±0,039b 0,324±0,036c 0,348±0,010 0,316±0,031
C 0,238±0,019c 0,685±0,402a 0,283±0,057c 0,364±0,049 0,393±0,132
D 0,192±0,045d 0,194±0,037c 0,251±0,104c 0,202±0,006 0,209±0,048
E 0,262±0,003c 0,408±0,169a 0,324±0,025c 0,365±0,043 0,340±0,060
F 0,344±0,028b 0,401±0,035a 0,384±0,040b 0,404±0,082 0,383±0,046
G 0,218±0,302c 0,095±0,046c 0,052±0,070d 0,517±0,496 0,221±0,329
H 0,085±0,075d 0,191±0,123c 0,144±0,087d 0,249±0,112 0,167±0,099
I 0,497±0,049a 0,515±0,024a 0,471±0,016a 0,634±0,164 0,529±0,063
J 0,425±0,045b 0,380±0,026b 0,612±0,296a 0,471±0,067 0,472±0,109
K 0,451±0,010b 0,461±0,086a 0,408±0,031b 0,564±0,041 0,471±0,042
L 0,563±0,209a 0,562±0,218a 0,490±0,045a 0,463±0,027 0,520±0,125
Rataan 0,323±0,074 0,380±0,123 0,340±0,078 0,411±0,114
26 Tabel 8. Populasi Bakteri Total (108 CFU/ml) dari Isolat Bakteri dalam Media BHI
yang Disuplementasi Mineral
Isolat Mineral Rataan
Co Cu Zn Mn
A 7,33±1,90c 9,10±8,30b 7,92±3,90c 8,53±8,10 8,22±5,55
B 6,80±1,20c 6,95±1,20b 7,71±1,10c 8,43±0,30 7,47±0,95
C 5,13±0,57c 18,60±0,12a 6,48±1,70c 8,92±1,50 9,78±0,97
D 3,76±1,40d 3,83±1,10c 5,53±3,10c 4,05±0,18 4,29±1,45
E 5,87±0,96c 10,20±5,10a 7,71±0,76c 8,95±1,30 8,18±2,03
F 8,33±0,84b 10,00±1,10a 9,53±1,20b 10,11±2,50 8,75±1,41
G 4,53±9,10c 0,85±1,40c 0,00±2,10d 13,52±0,15 4,73±3,19
H 0,55±2,30d 3,72±3,70c 2,33±2,60d 5,46±3,40 3,02±3,00
I 12,90±1,50a 13,50±0,72a 12,13±0,49a 17,02±4,90 13,89±1,90
J 10,80±1,40b 9,39±0,78b 16,36±8,90a 12,12±2,00 12,17±3,27
K 11,50±0,30b 11,80±2,60a 10,25±0,93b 14,91±1,20 12,12±1,26
L 14,90±6,30a 14,90±6,50a 12,70±1,40a 11,90±0,82 13,60±3,76
Rataan 7,70±2,31 9,40±2,72 8,22±2,35 10,33±2,20
Keterangan : A,B,C,…,L = kode isolat bakteri, nilai dengan superskrip yang berbeda pada setiap kolom menunjukkan pengaruh yang nyata diantara perlakuan
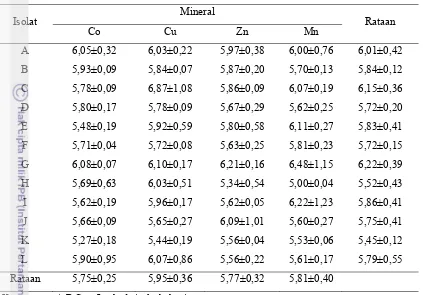
Berdasarkan hasil pada Tabel 7 dan 8, terlihat bahwa jumlah isolat bakteri di
dalam media BHI bermineral Co, Cu, dan Zn berbeda nyata (P<0,05). Hal tersebut
dapat disebabkan oleh masing-masing isolat bakteri yang memiliki kemampuan
tumbuh berbeda dalam media BHI bermineral tinggi tersebut. Lain halnya dengan
ketiga mineral tersebut, populasi isolat bakteri dalam media BHI bermineral Mn
menunjukkan hasil yang tidak berbeda nyata. Hal tersebut menunjukkan bahwa
ke-12 isolat bakteri tersebut memiliki kemampuan tumbuh yang sama dalam media BHI
bermineral Mn tinggi. Populasi tertinggi dari media BHI bermineral Co, Cu, dan Zn
ialah isolat bakteri I dan L, sedangkan dalam media BHI bermineral Mn ialah isolat
27
Percobaan Tahap 2: Adaptasi Isolat Bakteri Terpilih dalam Media Calf Starter
Produksi Bahan Kering Sel Bakteri dan Nilai pH Media
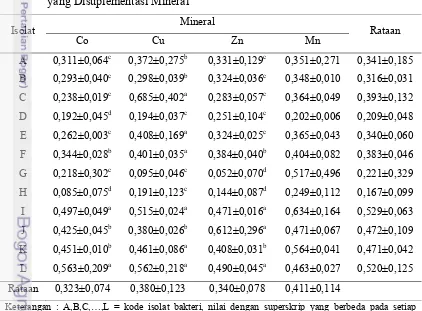
Produksi bahan kering (BK) sel bakteri dalam media calf starter yang berkadar Co, Cu, Zn, dan Mn tinggi ditunjukkan dalam Tabel 9. Pertumbuhan isolat
bakteri dalam substrat calf starter sangat rendah. Produksi bahan kering sel bakteri berkisar antara 0,05-5,70 mg/ml. Hasil tersebut menggambarkan bahwa isolat bakteri
tidak dapat berkembang dengan baik dalam subtrat bahan pakan yang ditambah Co,
Cu, Zn, dan Mn. Isolat B sangat sensitif terhadap penambahan kadar Cu, sedangkan
isolat C pertumbuhannya kurang baik dibandingkan dengan yang lainnya. Hal
tersebut menunjukkan bahwa isolat bakteri tidak dapat tumbuh baik dalam media calf starter. Produksi bahan kering sel bakteri baik pada media mengandung Co, Cu, Zn, dan Mn tinggi tidak berbeda antar bakteri. Semua isolat bakteri menunjukkan
pertumbuhan yang sama rendahnya pada substrat pakan yang ditambah Co, Cu, Zn,
dan Mn.
Tabel 9. Produksi Bahan Kering (BK) Sel Isolat Bakteri dalam Media Calf Starter
yang Disuplementasi Mineral (mg/ml)
Isolat Mineral Rataan
Co Cu Zn Mn
B 2,20±0,90 0,05±0,07 0,90±0,07 2,60±0,30 1,44±0,34
C 0,30±0,30 0,40±0,60 5,70±6,20 3,90±3,70 2,58±2,70
E 1,30±1,80 0,80±1,00 1,10±0,20 1,70±2,10 1,23±1,28
F 1,90±0,10 0,40±0,07 1,30±0,28 1,30±15,80 1,23±4,06
G 2,20±2,40 2,20±0,60 2,60±1,27 0,60±0,80 1,90±1,27
L 2,10±2,80 0,80±0,80 0,90±0,60 1,10±0,00 1,23±1,05
Rataan 1,67±1,38 0,78±0,52 2,08±1,44 1,87±3,78
Keterangan : B,C,E,F,G, dan L = kode isolat bakteri
Isolat bakteri pencerna serat diperkirakan tidak mampu memanfaatkan
substrat berupa calf starter dengan baik. Penambahan unsur Co, Cu, Zn, dan Mn ke dalam media diperkirakan menganggu mekanisme penggunaan nutrien substrat oleh
bakteri. Produksi dan aktivitas enzim pencerna komponen calf starter terganggu dengan penambahan Co, Cu, Zn, dan Mn dalam lingkungannya. Hal ini
28 yang lebih tinggi dalam media sehingga calf starter tidak dapat digunakan sebagai media untuk mengkonversi unsur Co, Cu, Zn, dan Mn inorganik menjadi Co, Cu, Zn,
dan Mn organik oleh isolat bakteri.
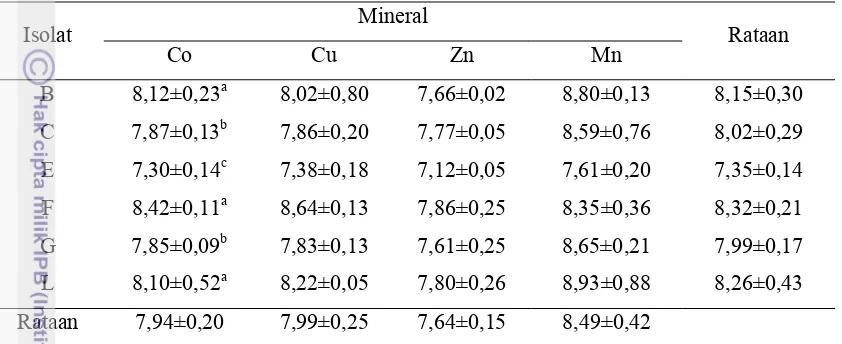
Tabel 10. Nilai pH dalam Media Calf Starter yang Disuplementasi Mineral
Isolat Mineral Rataan
Rataan 7,94±0,20 7,99±0,25 7,64±0,15 8,49±0,42
Keterangan : B,C,E,F,G, dan L = kode isolat bakteri, nilai dengan superskrip yang berbeda pada setiap kolom menunjukkan pengaruh yang nyata diantara perlakuan
Berdasarkan Tabel 10, terlihat bahwa yang memberikan pengaruh berbeda
diantara perlakuan ialah pada media calf starter bermineral cobalt (Co). Hal tersebut menunjukkan bahwa keenam isolat bakteri memiliki respon yang berbeda pada
media calf starter bermineral Co meskipun secara keseluruhan nilai pH yang dihasilkan cenderung sama yaitu dalam keadaan basa. Keadaan basa dalam media
calf starter dapat disebabkan oleh kualitas calf starter yang rendah dan banyaknya kation dalam media tersebut yang menyebabkan aktifitas bakteri kurang maksimal,
sehingga kemampuan bakteri untuk tumbuh juga terbatas. Kisaran pH yang
dihasilkan berkisar antara 7,30-8,93. Nilai pH pada keenam isolat bakteri dalam
media calf starter bermineral Cu, Zn, dan Mn menunjukkan hasil yang tidak berbeda. Hal tersebut mengindikasikan bahwa respon keenam isolat bakteri tersebut sama
diantara ketiga mineral.
Populasi Isolat Bakteri dalam Media
Berdasarkan hasil pada Tabel 11 dan 12 menunjukkan bahwa populasi isolat
bakteri dalam media calf starter bermineral Co, Cu, Zn, dan Mn tidak berbeda nyata. Hal tersebut mengindikasikan bahwa kemampuan hidup dan adaptasi keenam isolat
29 Meskipun demikian, jumlah isolat bakteri terbesar terdapat pada media calf starter
bermineral Mn dengan rataan nilai jumlah bakteri sebesar 0,517 cfu/ml atau populasi
total bakteri sebesar 13,49 (108 cfu/ml).
Tabel 11. Jumlah Bakteri dalam Media Calf Starter yang Disuplementasi
Berdasarkan Nilai Absorbansi (600 nm)
Isolat Mineral Rataan
Co Cu Zn Mn
B 0,260±0,004 0,400±0,030 0,386±0,009 0,495±0,071 0,385±0,029
C 0,369±0,049 0,339±0,045 0,424±0,065 0,504±0,132 0,409±0,073
E 0,313±0,056 0,276±0,013 0,306±0,093 0,332±0,085 0,307±0,062
F 0,302±0,081 0,305±0,004 0,403±0,004 0,563±0,185 0,393±0,069
G 0,375±0,061 0,365±0,112 0,365±0,007 0,545±0,088 0,413±0,067
L 0,356±0,098 0,382±0,055 0,421±0,027 0,660±0,110 0,455±0,073
Rataan 0,330±0,058 0,345±0,043 0,384±0,034 0,517±0,112
Keterangan : B,C,E,F,G, dan L = kode isolat bakteri
Tabel 12. Populasi Bakteri Total (108 CFU/ml) dari Isolat Bakteri dalam Media Calf Starter yang Disuplementasi Mineral
Isolat Mineral Rataan
Co Cu Zn Mn
B 5,80±1,30 9,99±9,10 9,58±0,85 12,80±2,14 9,54±3,35
C 9,06±1,46 8,16±1,34 10,70±1,95 13,10±3,95 10,26±2,18
E 7,38±1,68 6,27±4,03 7,18±2,80 7,96±2,55 7,20±2,77
F 7,05±2,44 7,14±1,06 10,10±1,06 14,90±5,56 9,80±2,53
G 9,25±1,82 8,94±3,37 8,95±2,12 14,40±2,63 10,39±2,49
L 8,68±2,93 9,48±1,65 10,60±8,06 17,80±3,31 11,64±3,99
Rataan 7,87±1,94 8,33±3,43 9,52±2,81 13,49±3,36
Keterangan : B,C,E,F,G, dan L = kode isolat bakteri
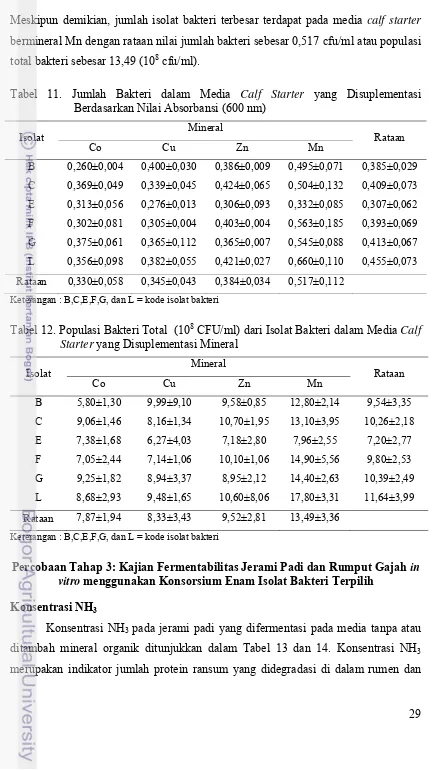
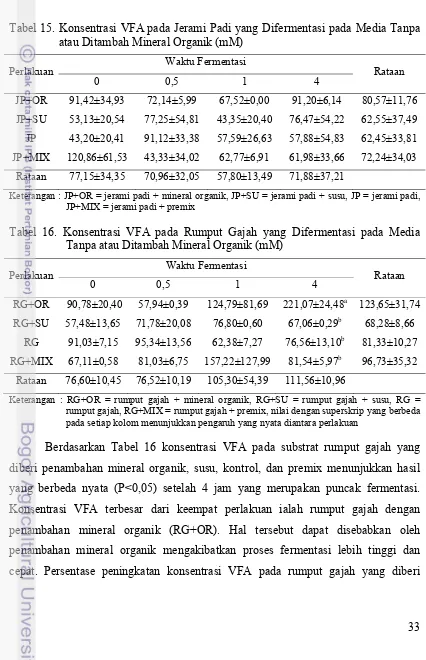
Percobaan Tahap 3: Kajian Fermentabilitas Jerami Padi dan Rumput Gajah in vitro menggunakan Konsorsium Enam Isolat Bakteri Terpilih
Konsentrasi NH3
Konsentrasi NH3 pada jerami padi yang difermentasi pada media tanpa atau
ditambah mineral organik ditunjukkan dalam Tabel 13 dan 14. Konsentrasi NH3
30 nilainya sangat dipengaruhi oleh kemampuan mikroba rumen dalam mendegradasi
protein ransum (Prihandono, 2001).
Protein mengalami hidrolisa menjadi oligopeptida bahkan asam amino oleh
enzim proteolisis yang dihasilkan oleh mikroba di dalam rumen (Sutardi, 1977).
Besarnya protein yang lolos dari degradasi rumen berkisar 20–80% (Sutardi, 1977).
Amonia digunakan oleh bakteri untuk membentuk protein tubuhnya selanjutnya
protein mikroba akan dicerna dan diserap serta dikatabolisasi sebagai salah satu
masukan induk semang.
Data pada Tabel 13 menunjukkan bahwa konsentrasi NH3 meningkat setelah
inkubasi 0,5 jam, kecuali pada substrat jerami padi yang ditambah campuran mineral
inorganik. Konsentrasi NH3 berbeda antar bahan dan waktu fermentasi. Peningkatan
kadar NH3 diperkirakan akibat adanya degradasi protein. Substrat yang mengandung
susu baik JP+OR maupun JP+SU, NH3 diperkirakan berasal dari protein susu yang
mengalami degradasi, sedangkan NH3 dalam subtrat jerami padi saja atau JP, NH3
dapat berasal dari degradasi protein bakteri. Penurunan ammonia pada JP+Mix
diperkirakan akibat pemanfaatan NH3 yang efisien dalam subtrat yang mendapat
suplementasi mineral. Kadar NH3 dalam perlakuan JP+OR meningkat pada jam ke 4.
Hal ini menggambarkan bahwa penambahan mineral organik cenderung
meningkatkan degradasi protein oleh mikroba rumen. Persentase peningkatan
konsentrasi NH3 pada jerami padi yang diberi penambahan mineral organik
dihasilkan sebanyak 31,33%. Konsentrasi NH3 dengan penambahan mineral
inorganik justru menurun sebesar 48,67% dibandingkan dengan kontrol (JP).
Menurut Sutardi (1977) kadar NH3 yang mendukung perkembangan mikroorganisme