BIOEKOLOGI CACING TANAH
Pheretima darnleiensis
ANDY DARMAWAN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Bioekologi Cacing Tanah Pheretima darnleiensis adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada Perguruan Tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir tesis ini.
Bogor, Juli 2010
ABSTRACT
ANDY DARMAWAN. Bioecology of Earthworm Pheretima darnleiensis. Supervised by RIKA RAFFIUDIN and TRI HERU WIDARTO.
Pheretima darnleiensis is a native earthworm in Southeast Asia, India, Japan, and Indonesia, however there is lack of study on P. darnleiensis. Determining the characteristics of P. darnleiensis and the ecology surrounding its habitat is needed in order to culture the earthworm. Hence, this study was aimed to study the structure of several organs and morphological characters that can be used to identify P. darnleiensis and its habitat condition. Earthworms were collected in a plot of 100 x 100 x 20 cm in 10 locations and were identified. The structure of P. darnleiensis organs was studied histologically using paraffin. Pheretima darnleiensis did not have diverticula and stalked gland on copulatory pouches, nephridia occurred on bithecate spermatheca, and the first spermathecal pores were at segment 4/5. Based on this study, the highest population of P. darnleiensis was found in an area containing abundance of other earthworm species at 15 individuals/m2, plant cover at 25.00%, soil arthropods at 16.32 individuals/kg fresh soil, abiotic component cover at 78.50%, soil temperature at 27.42 ± 1.01 oC, soil pH at 5.77 ± 0.53, soil moisture at 19.83 ± 9.92%, organic carbon at 3.39%, N at 0.29%, P at 286.80 ppm, K at 500.00 ppm, sand at 28.76%, silt at 38.74%, and clay at 32.50%. Silt fraction could be considered as one of factors influencing P. darnleiensis abundance.
RINGKASAN
ANDY DARMAWAN. Bioekologi Cacing Tanah Pheretima darnleiensis. Dibimbing oleh RIKA RAFFIUDIN dan TRI HERU WIDARTO.
Cacing tanah merupakan hewan yang memiliki banyak manfaat, misalnya dapat digunakan sebagai obat, agen dekomposisi, atau bahan makanan. Penelitian mengenai cacing tanah dan cara membudidayakannya di Indonesia selama ini banyak menggunakan cacing Eropa seperti Eisenia dan Lumbricus, sementara itu Pheretima darnleiensis sebagai cacing lokal Indonesia belum banyak diteliti. Untuk membudidayakan P. darnleiensis, perlu diketahui karakter yang menjadi ciri identifikasi dan ekologinya. Oleh karena itu, perlu dilakukan penelitian mengenai bioekologi P. darnleiensis. Aspek yang dipelajari meliputi karakter morfologi yang dapat menjadi ciri identifikasi P. darnleiensis, serta kondisi lingkungan tempat ditemukannya P. darnleiensis.
Sampling dilakukan pada sepuluh lokasi di sekitar kampus IPB Darmaga Kabupaten Bogor dan Baranangsiang Kotamadya Bogor. Plot berukuran sekitar 100 x 100 cm dengan kedalaman 20 cm dibuat pada tiap lokasi. Seluruh cacing dikoleksi dengan cara menggali menggunakan sekop.
Identifikasi P. darnleiensis dilakukan dengan mengamati karakter eksternal dan internalnya. Karakter eksternal diamati secara langsung ataupun dengan mikroskop stereo. Organ-organ internal yang diamati adalah sekum, spermateka, dan kantong kopulasi. Organ tersebut diamati dengan mikroskop stereo.
Pengamatan histologi P. darnleiensis dilakukan dengan metode sayatan menggunakan parafin. Jaringan P. darnleiensis difiksasi dengan FAAC, diwarnai dengan haematoksilin-eosin, lalu ditutup dengan entellan. Identifikasi dan pembuatan preparat jaringan P. darnleiensis dilakukan di Laboratorium Mikroteknik bagian Fungsi dan Perilaku Hewan Departemen Biologi FMIPA IPB. Ekologi P. darnleiensis dipelajari dengan mengamati kondisi biotik maupun abiotik lingkungannya. Kondisi biotik yang dipelajari meliputi jumlah cacing yang ditemukan bersama dengan P. darnleiensis, persen tutupan tumbuhan, dan jumlah artropoda tanah yang ditemukan pada lokasi sampling. Seluruh cacing yang dikoleksi diawetkan dalam alkohol 70% untuk diidentifikasi hingga tingkat genus, kecuali cacing Pheretima diidentifikasi hingga tingkat spesies. Persen tutupan tumbuhan ditentukan dengan menghitung luas lokasi sampling pada tiap lokasi yang tertutup tumbuhan. Seluruh tumbuhan yang terdapat di lokasi pengamatan diidentifikasi hingga tingkat ordo. Jumlah artropoda pada tiap lokasi dihitung dalam 250 gram tanah segar menggunakan saluran Tullgren. Artropoda yang dikoleksi diidentifikasi hingga tingkat ordo.
Analisis karbon organik, nitrogen, fosfat, kalium, dan tekstur tanah dilakukan di Laboratorium Departemen Ilmu Tanah dan Sumber Daya Lahan Fakultas Pertanian IPB. Karbon organik dianalisis dengan metode Walkley-Black, nitrogen dengan metode Kjeldahl, fosfat dan kalium dengan metode spektrofotometri, dan tekstur dengan metode pipet.
Analisis komponen utama dilakukan untuk menentukan variabel yang berpengaruh terhadap keberadaan P. darnleiensis. Hubungan antarvariabel dianalisis dengan korelasi Pearson. Analisis data dilakukan dengan program R 2.4.0.
Cacing yang dikoleksi dari sepuluh lokasi terdiri atas empat genus, yaitu Amynthas, Metaphire, Pheretima, dan Pontoscolex. Genus Pheretima memiliki tubuh silindris dengan warna keseluruhan tubuh gelap dengan bagian dorsal lebih gelap daripada bagian ventral, banyak seta tersebar per segmen (sekitar 40 pada segmen 13), sekum pada segmen 27, terdapat kantong kopulasi, dan memiliki nefridia pada spermateka. Sementara itu, spesies P. darnleiensis memiliki kantong kopulasi tanpa divertikula dan kelenjar bertangkai, posisi spermateka bithecate, dan lubang spermateka pertama pada segmen 4/5.
Klitelum P. darnleiensis memiliki lapisan kelenjar mukus, kelenjar pensekresi kokon, dan kelenjar pensekresi albumin. Faring bagian dalam P. darnleiensis tersusun atas epitel kolumnar bersilia, pada lambungnya terdapat lapisan kutikula yang tebal, dan tiflosol P. darnleiensis berbentuk lurus. Pembuluh darah dorsal P. darnleiensis lebih tebal daripada pembuluh darah ventralnya. Ganglion serebral P. darnleiensis terdiri atas serabut saraf dan sel saraf, pada tali sarafnya terdapat serabut kasar. Sperma dewasa ditemukan pada spermateka P. darnleiensis, sedangkan pada vesikula seminalis ditemukan sperma dewasa dan muda serta Monocystis. Prostat P. darnleiensis berbentuk racemose.
Pheretima darnleiensis dikoleksi pada lima dari sepuluh lokasi pengamatan, yaitu di lokasi 3, 4, 5, 8, dan 9 sebanyak masing-masing satu individu kecuali pada lokasi 5 sebanyak tujuh individu. Kondisi lingkungan lokasi yang terdapat P. darnleiensis dibandingkan dengan lokasi tanpa P. darnleiensis (1, 2, 6, 7, dan 10) menggunakan uji t. Nilai-nilai variabel antara lokasi dengan P. darnleiensis dan lokasi tanpa P. darnleiensis tidak berbeda signifikan.
Jumlah P. darnleiensis paling banyak pada lokasi 5, yaitu sebanyak tujuh individu. Pada lokasi ini dikoleksi cacing lain sebanyak 15 individu/m2, yaitu Amynthas, Metaphire, Pontoscolex, dan cacing juvenil berturut-turut sebanyak 6, 4, 1, dan 4 individu. Lokasi 5 juga tertutup tumbuhan sebanyak 25.00% (Glumiflorae 22.00% dan Spathiflorae 3.00%). Artropoda yang terdapat pada lokasi 5 dalam tiap kilogram tanah segar antara lain Colembola, Hymenoptera (Fomicidae), Diplura, Isopoda, Parasitiformes, dan Acariformes, masing-masing sebanyak 2.04 individu kecuali Hymenoptera sebanyak 6.12 individu. Tutupan komponen abiotik pada lokasi 5 sebesar 78.50%, temperatur tanah 27.42 ± 1.01
o
Biplot analisis komponen utama variabel dan lokasi penelitian menunjukkan P. darnleiensis berada dekat dengan lokasi 5. Hal ini sejalan dengan kelimpahan P. darnleiensis yang paling tinggi pada lokasi 5. Variabel yang terletak dekat dengan lokasi 5 adalah tutupan tumbuhan dan fraksi debu. Tutupan tumbuhan dan kelimpahan P. darnleiensis memiliki nilai korelasi tidak signifikan sebesar 0.14 sedangkan fraksi debu dan kelimpahan P. darnleiensis memiliki nilai korelasi signifikan sebesar 0.82.
Berdasarkan hasil penelitian ini, karakter yang dapat menjadi ciri identifikasi P. darnleiensis adalah memiliki tubuh silindris dengan warna keseluruhan tubuh gelap dengan bagian dorsal lebih gelap daripada bagian ventral dan banyak seta tersebar per segmen (sekitar 40 pada segmen 13). Sekum P. darnleiensis terletak pada segmen 27. Pada dasar prostat P. darnleiensis terdapat kantong kopulasi tanpa divertikula dan kelenjar bertangkai. Pada spermateka terdapat nefridia, posisi spermateka bithecate, dan lubang spermateka pertama pada segmen 4/5.
Berdasarkan penelitian ini, P. darnleiensis paling banyak dikoleksi pada kondisi lingkungan terdapat 15 individu cacing lain/m2, tutupan tumbuhan 25.00%, artropoda tanah 16.32 individu/kg tanah segar, tutupan komponen abiotik 78.50%, temperatur tanah 27.42 ± 1.01 oC, pH tanah 5.77 ± 0.53, kelembaban tanah 19.83 ± 9.92%, C organik = 3.39%, N = 0.29%, P = 286.80 ppm, K = 500.00 ppm, fraksi pasir 28.76%, debu 38.74%, dan liat 32.50%. Fraksi tanah debu merupakan komponen yang berpengaruh terhadap kelimpahan P. darnleiensis.
Hak Cipta milik IPB, tahun 2010
Hak cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tidak merugikan kepentingan yang wajar IPB
BIOEKOLOGI CACING TANAH
Pheretima darnleiensis
ANDY DARMAWAN
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Biosains Hewan
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PRAKATA
Puji dan syukur penulis panjatkan kepada Tuhan atas berkat-Nya sehingga tulisan ini dapat diselesaikan. Terima kasih penulis ucapkan kepada Dr. Rika Raffiudin, M.Si. dan Ir. Tri Heru Widarto, M.Sc. selaku pembimbing, dan juga Dr. Ir. Sulistijorini, M.Si. selaku penguji atas sarannya. Penulis juga mengucapkan terima kasih kepada seluruh staf pengajar Biosains Hewan Departemen Biologi FMIPA IPB atas diskusi dan dukungannya, juga kepada Pak Hari Nugroho selaku staf LIPI Cibinong atas bimbingannya. Terima kasih juga penulis ucapkan kepada Mbak Tini, Mbak Ani, Pak Nunu, Pak Heri, dan Pak Adi selaku laboran bagian Fungsi dan Perilaku Hewan Biologi FMIPA IPB, Pak Suparman selaku laboran bagian Taksonomi Tumbuhan, Bu Iko di departeman tanah, seluruh teman, dan orang tua penulis atas dukungannya. Semoga karya ilmiah ini bermanfaat.
Bogor, Juli 2010
RIWAYAT HIDUP
DAFTAR ISI
Halaman
DAFTAR ISI ... xiii
DAFTAR TABEL ... xiv
DAFTAR GAMBAR ... xv
1 PENDAHULUAN Latar Belakang Penelitian ... 1
Tujuan Penelitian ... 2
Manfaat Penelitian ... 2
2 KARAKTER MORFOLOGI DAN HISTOLOGI Pheretima darnleiensis Pendahuluan ... 3
Bahan dan Metode ... 4
Hasil ... 6
Pembahasan ... 14
Simpulan ... 21
3 EKOLOGI Pheretima darnleiensis Pendahuluan ... 22
Bahan dan Metode ... 24
Hasil ... 27
Pembahasan ... 35
Simpulan ... 39
4 PEMBAHASAN UMUM ... 40
5 SIMPULAN DAN SARAN ... 42
DAFTAR PUSTAKA ... 43
DAFTAR TABEL
Halaman
1 Jumlah cacing yang dikoleksi dari lokasi koleksi empat genus cacing ... 7
2 Perbandingan warna, aktivitas, dan panjang cacing yang dikoleksi ... 7
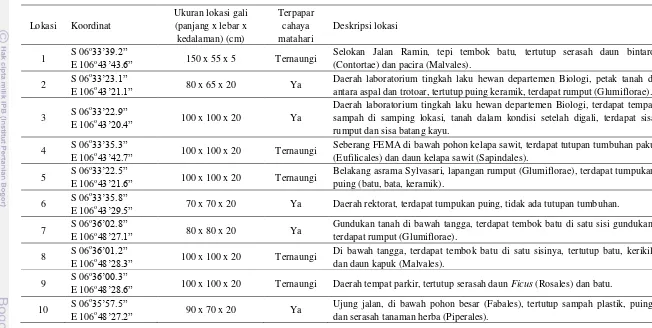
3 Koordinat dan deskripsi lokasi koleksi empat genus cacing di kampus IPB Darmaga Kabupaten Bogor dan kampus IPB Baranangsiang Kotamadya Bogor ... 28
4 Karakter biotik dan abiotik lokasi koleksi empat genus cacing ... 29
5 Tumbuhan yang terdapat di lokasi koleksi empat genus cacing ... 30
6 Artropoda tanah yang terdapat di lokasi koleksi empat genus cacing ... 31
7 Kisaran temperatur, pH, dan kelembaban tanah lokasi koleksi empat genus cacing ... 31
DAFTAR GAMBAR
Halaman
1 Empat genus cacing yang dikoleksi ... 7
2 Karakter eksternal P. darnleiensis ... 8
3 Karakter internal P. darnleiensis... 9
4 Sekum, kantong kopulasi, spermateka, dan gambaran skematik spermateka dengan nefridia P. darnleiensis ... 9
5 Fotomikrograf dinding tubuh P. darnleiensis ... 10
6 Faring dan lambung P. darnleiensis... 11
7 Fotomikrograf saluran pencernaan P. darnleiensis ... 11
8 Fotomikrograf sistem sirkulasi P. darnleiensis ... 12
9 Fotomikrograf nefridia P. darnleiensis ... 12
10 Fotomikrograf sistem saraf P. darnleiensis ... 13
11 Fotomikrograf organ reproduksi P. darnleiensis ... 13
12 Diagramatik sayatan melintang Lumbricus ... 19
13 Saluran Tullgren untuk mengekstraksi artropoda tanah ... 26
14 Biplot analisis komponen utama variabel dan lokasi penelitian ... 33
15 Gambar skematik lokasi lubang jantan Pontoscolex corethrurus ... 48
16 Lokasi lubang jantan P. darnleiensis ... 48
17 Gambar skematik seta Pontoscolex corethrurus ... 48
18 Gambar skematik seta P. darnleiensis ... 49
19 Kantong kopulasi P. darnleiensis dan gambar skematiknya ... 49
20 Prostat Amynthas dan gambar skematiknya ... 49
21 Spermateka P. darnleiensis dan gambar skematiknya ... 49
1 PENDAHULUAN
Latar Belakang Penelitian
Cacing tanah merupakan cacing terrestrial yang tersebar luas di bumi, kecuali pada lingkungan ekstrim seperti gurun, salju, dan es (Stephenson 1930). Cacing tanah secara umum memiliki banyak manfaat, diantaranya adalah menyuburkan tanah dan dapat digunakan sebagai indikator pencemaran tanah (Edwards 2004), bahan makanan (Simanjuntak & Walujo 1982), obat (Mihara et al. 1992; Hong et al. 2007), atau agen dekomposisi (Ndegwa et al. 2000; Arancon
et al. 2003; Aira et al. 2007). Selain itu, keberadaan cacing dapat meningkatkan
kandungan auksin dan sitokinin dalam tanah (Krishnamoorthy & Vajranabhaiah 1986). Kasting cacing banyak mengandung unsur hara yang siap tersedia bagi tanaman dan aktivitas menggali cacing dapat memperbaiki tata udara tanah sehingga infiltrasi udara menjadi lebih baik dan mudah ditembus akar (Hardjowigeno 2007). Pheretima darnleiensis juga diperkirakan memiliki potensi seperti cacing tanah pada umumnya, namun penelitian mengenai manfaat P. darnleiensis belum pernah dilakukan.
Pheretima endemik di Asia Tenggara, termasuk India Timur dan Jepang
(Stephenson 1930). Sims & Easton (1972) mengoleksi spesies P. darnleiensis di Borneo. Stephenson (1930) mengelompokkan P. darnleiensis ke dalam ordo Oligochaeta, subordo Neooligochaeta, famili Megascolecidae, subfamili Megascolecinae, genus Pheretima, spesies P. darnleiensis.
Pheretima darnleiensis sebagai cacing lokal di Indonesia belum banyak
diteliti. Budidaya cacing di Indonesia umumnya menggunakan cacing Eropa Lumbricus dan Eisenia. Untuk membudidayakan P. darnleiensis, diperlukan data
Tujuan Penelitian
Tujuan penelitian ini adalah mempelajari (1) karakter morfologi dan histologi yang dapat menjadi ciri identifikasi P. darnleiensis, serta (2) kondisi lingkungan tempat ditemukannya P. darnleiensis. Hasil penelitian ini diharapkan dapat menggali potensi P. darnleiensis untuk dimanfaatkan dan dibudidayakan.
Manfaat Penelitian
2 KARAKTER MORFOLOGI DAN HISTOLOGI
Pheretima darnleiensis
PENDAHULUAN
Dalam membudidayakan cacing tanah diperlukan penentuan spesies cacing yang dipelihara. Oleh karena itu, cara untuk mengidentifikasi cacing tanah penting untuk diketahui. Untuk mengidentifikasi cacing tanpa mematikannya dapat dilakukan dengan mengamati karakter eksternalnya, seperti bentuk tubuh, warna tubuh, posisi seta, dan posisi lubang genital, namun identifikasi cacing tanah secara pasti hingga famili hanya dapat dilakukan dengan melakukan pembedahan dan mengamati karakter morfologi yang menjadi ciri identifikasi, seperti sekum, kantong kopulasi, dan spermateka (Sims & Easton 1972).
di segmen 8/9 dan 9/10. Jantung berpasangan di segmen 11–13 dan satu di segmen 10. Tubula nefridial tidak ada (Ishizuka 1999).
Penelitian mengenai histologi P. darnleiensis belum pernah dilakukan. Penelitian mengenai histologi cacing tanah pernah dilakukan oleh Coggeshall (1966) dan Hama (1959, 1960). Coggeshall (1966) mempelajari lapisan epidermis Lumbricus terrestris dan menyatakan lapisan tersebut terdapat di bawah lapisan
kutikula dan memiliki ketebalan 50–70 µm. Hama (1959) menyatakan serabut kasar Eisenia foetida diselubungi lapisan Schwann, sedangkan Hama (1960) mempelajari pembuluh darah pada Eisenia foetida dan mendapati adanya lapisan otot sikular dan longitudinal pada pembuluh darah dorsal dan ventral.
Penentuan spesies P. darnleiensis dalam membudidayakannya sangat penting, akan tetapi belum ada penelitian mengenai karakter morfologi dan histologi P. darnleiensis, maka penelitian ini perlu dilakukan. Penelitian ini bertujuan mempelajari karakter morfologi dan histologi yang dapat menjadi ciri identifikasi P. darnleiensis. Data hasil penelitian ini diharapkan dapat digunakan untuk melakukan identifikasi terhadap P. darnleiensis.
BAHAN DAN METODE
Waktu dan Tempat Penelitian
Penelitian mengenai karakter morfologi dan histologi P. darnleiensis dilakukan dari bulan Oktober 2009–Januari 2010. Identifikasi dan pengamatan histologi P. darnleiensis dilakukan di Laboratorium Mikroteknik bagian Fungsi Hayati dan Perilaku Hewan, Departemen Biologi FMIPA IPB.
Alat dan Bahan
Alat yang digunakan untuk melakukan identifikasi P. darnleiensis adalah mikroskop stereo, gunting bedah, jarum, pinset, lilin, dan cawan petri. Bahan yang digunakan untuk melakukan identifikasi P. darnleiensis adalah alkohol 70% dan akuades.
5
preparat, kaca tutup, mikroskop, dan fotomikroskop. Bahan yang digunakan dalam proses pembuatan sayatan P. darnleiensis adalah FAAC, alkohol seri (30, 50, 70, 80, 95, dan 100%), xilol, parafin, albumin, akuades, haematoksilin Ehrlich, eosin 1%, air, dan entellan.
Identifikasi P. darnleiensis
Seluruh cacing yang didapatkan dari lokasi penelitian diawetkan dalam alkohol 70%. Cacing diidentifikasi dengan kunci identifikasi Sims & Easton (1972). Karakter morfologi yang diamati meliputi klitelum, prostomium, seta, lubang jantan, lubang betina, lubang spermateka, lubang dorsal, sekum, prostat, dan spermateka.
Pembuatan Preparat untuk Pengamatan Histologi P. darnleiensis
Metode Pembuatan Preparat dan Organ P. darnleiensis yang
Dipelajari. Seluruh penelitian ini menggunakan cacing dewasa kecuali untuk pengamatan dinding tubuh yang menggunakan cacing dewasa dan muda. Metode yang digunakan untuk mempelajari histologi P. darnleiensis adalah metode sayatan modifikasi Gunarso (1989). Metode ini menghasilkan sayatan seri yang saling berhubungan. Sayatan yang dibuat adalah dinding tubuh (tanpa klitelum dan dengan klitelum), organ pencernaan (faring, lambung, dan intestinum), organ sirkulasi (pembuluh darah dorsal dan ventral), organ ekskresi (nefridia), organ saraf (ganglion serebral dan tali saraf), dan organ reproduksi (spermateka, vesikula seminalis, dan prostat). Seluruh sayatan tersebut dibuat dengan menggunakan P. darnleiensis yang telah memiliki klitelum.
Proses Pembuatan Preparat P. darnleiensis. Pertama cacing dibersihkan
alkohol:xilol dengan perbandingan 1:1 selama 60 menit, xilol I selama 60 menit, dan xilol II selama 10 menit. Penjernihan dengan xilol II dilakukan di dalam oven dengan temperatur 60 oC. Setelah penjernihan organ cacing diinfiltrasi dengan parafin cair yang memiliki titik lebur 59 oC selama 3 x 45 menit di dalam oven dengan temperatur 60 oC. Organ yang telah selesai diinfiltrasi dicetak ke dalam cetakan yang terbuat dari kertas kalender. Blok parafin disayat melintang dengan mikrotom putar setebal 6 μm. Pita hasil sayatan dilekatkan ke kaca preparat dengan mengoleskan kaca preparat dengan satu tetes larutan albumin:gliserin dengan perbandingan 1:1, lalu dipanaskan di atas hot plate dengan temperatur 40
o
C selama 24 jam. Parafin dihilangkan dari jaringan dengan merendam preparat dalam tiga tabung xilol yang terpisah masing-masing selama 5, 5, dan 10 menit. Selanjutnya adalah tahap dealkoholisasi dengan mencelupkan preparat berisi sayatan jaringan ke dalam alkohol 100, 95, 80, dan 70%. Jaringan diwarnai dengan haematoksilin Ehrlich selama 1 menit lalu dibilas dengan air untuk menghilangkan kelebihan haematoksilin Ehrlich. Pewarnaan berikutnya dengan merendam jaringan dalam eosin selama 10 menit lalu dibilas dengan air untuk menghilangkan kelebihan eosin. Setelah diwarnai jaringan didehidrasi dengan mencelupkan preparat ke dalam alkohol 30, 50, 70, 80, 95, dan 100%. Penjernihan dilakukan dengan merendam preparat dalam xilol I selama 10 menit dan xilol II 10 menit. Preparat ditutup dengan kaca tutup mengunakan entellan. Hasil sayatan difoto dengan menggunakan fotomikroskop (Olympus).
HASIL
Identifikasi P. darnleiensis
Cacing yang ditemukan di lokasi penelitian sebanyak empat genus, yaitu Amynthas (Gambar 1a), Metaphire (Gambar 1b), Pheretima (Gambar 1c), dan
Pontoscolex (Gambar 1d) (Tabel 1). Cacing genus Pheretima memiliki warna
7
(a) (b)
(c) (d)
Gambar 1 Empat genus cacing yang dikoleksi: (a) Amynthas, (b) Metaphire, (c) Pheretima, dan (d) Pontoscolex. Garis skala = 5 cm.
Tabel 1 Jumlah cacing yang dikoleksi dari lokasi koleksi empat genus cacing
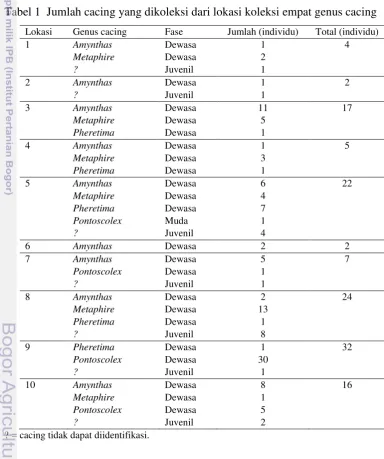
Lokasi Genus cacing Fase Jumlah (individu) Total (individu)
Tabel 2 Perbandingan warna dorsal, aktivitas, dan panjang cacing yang dikoleksi
Genus cacing Warna tubuh Aktivitas Panjang (cm)
Amynthas Gelap (+++) Aktif (++) 15–20
Metaphire Gelap (+++) Aktif (++) 15–20
Pheretima Sangat gelap (++++) Sangat aktif (+++) 10–15
Pontoscolex Pucat (+) Pasif (+) 5–15
(a) (b)
(c) (d)
(e) (f) (g)
(h) (i) (j)
9
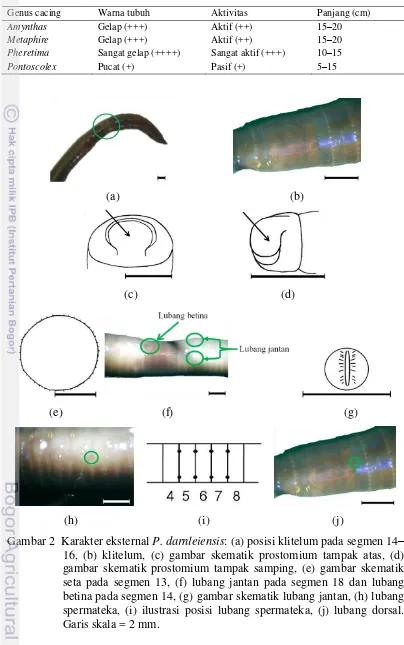
Karakter eksternal P. darnleiensis yang diamati adalah klitelum berbentuk cincin pada segmen 14–16 (Gambar 2a, 2b), prostomium epilobus dengan dasar terbuka (Gambar 2c, 2d), jumlah seta sekitar 40 pada segmen 13 (Gambar 2e). Lubang betina satu di bagian ventral tengah pada segmen 14 (Gambar 2f), lubang jantan sepasang di bagian ventral tepi pada segmen 18 (Gambar 2f, 2g), lubang spermateka 4 pasang di bagian ventral tepi pada segmen 4/5, 5/6, 6/7, 7/8 (Gambar 2h, 2i). Lubang dorsal satu di bagian tengah segmen dan lubang dorsal pertama di segmen 12/13 (Gambar 2j).
Gambar 3 Karakter internal P. darnleiensis. Garis skala = 2 mm.
(a) (b)
(c) (d)
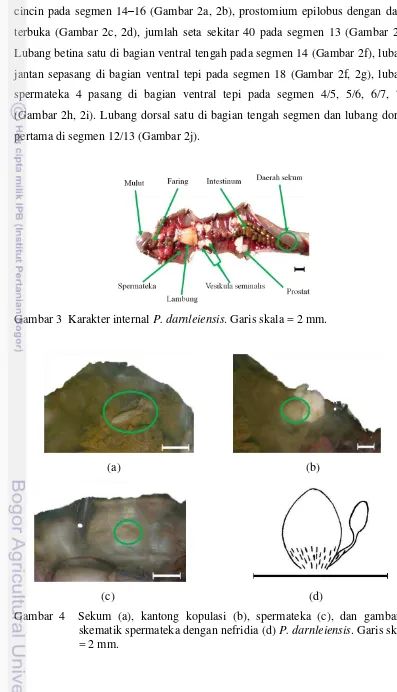
Untuk mengamati karakter internal, P. darnleiensis dibedah pada bagian dorsal dari bagain anterior ke posterior (Gambar 3). Karakter internal P. darnleiensis yang berfungsi sebagai ciri identifikasi adalah terdapat sekum pada
segmen 27 (Gambar 4a). Kantong kopulasi tanpa divertikula dan kelenjar bertangkai (Gambar 4b), spermateka dengan nefridia (Gambar 4c, 4d), bithecate, dan lubang spermateka pertama pada segmen 4/5.
Histologi P. darnleiensis
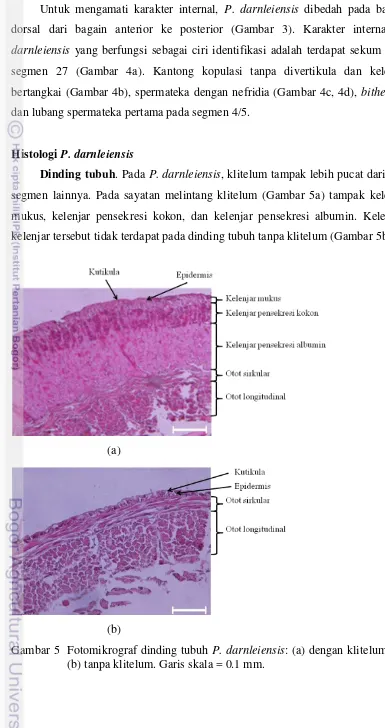
Dinding tubuh. Pada P. darnleiensis, klitelum tampak lebih pucat daripada segmen lainnya. Pada sayatan melintang klitelum (Gambar 5a) tampak kelenjar mukus, kelenjar pensekresi kokon, dan kelenjar pensekresi albumin. Kelenjar-kelenjar tersebut tidak terdapat pada dinding tubuh tanpa klitelum (Gambar 5b).
(a)
(b)
11
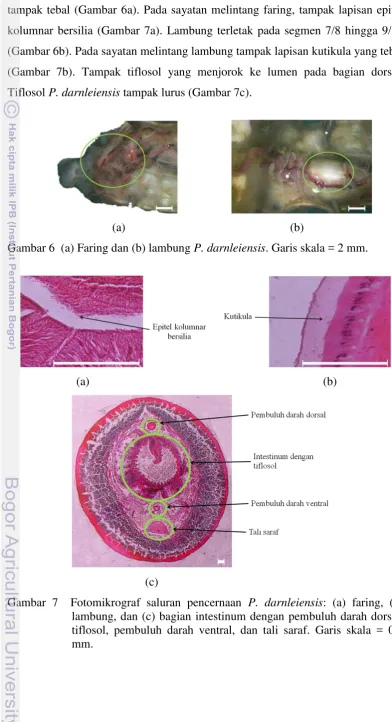
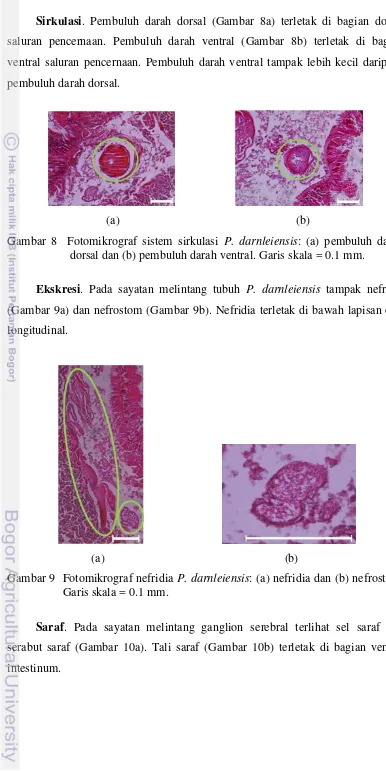
Pencernaan. Faring P. darnleiensis terletak di belakang rongga mulut dan tampak tebal (Gambar 6a). Pada sayatan melintang faring, tampak lapisan epitel kolumnar bersilia (Gambar 7a). Lambung terletak pada segmen 7/8 hingga 9/10 (Gambar 6b). Pada sayatan melintang lambung tampak lapisan kutikula yang tebal (Gambar 7b). Tampak tiflosol yang menjorok ke lumen pada bagian dorsal. Tiflosol P. darnleiensis tampak lurus (Gambar 7c).
(a) (b)
Gambar 6 (a) Faring dan (b) lambung P. darnleiensis. Garis skala = 2 mm.
(a) (b)
(c)
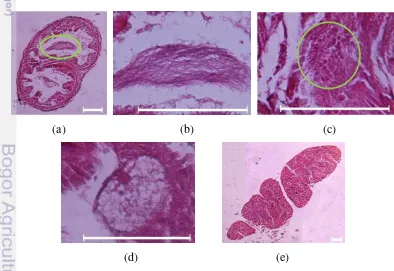
Sirkulasi. Pembuluh darah dorsal (Gambar 8a) terletak di bagian dorsal saluran pencernaan. Pembuluh darah ventral (Gambar 8b) terletak di bagian ventral saluran pencernaan. Pembuluh darah ventral tampak lebih kecil daripada pembuluh darah dorsal.
(a) (b)
Gambar 8 Fotomikrograf sistem sirkulasi P. darnleiensis: (a) pembuluh darah dorsal dan (b) pembuluh darah ventral. Garis skala = 0.1 mm.
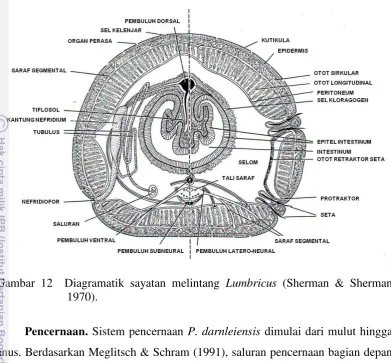
Ekskresi. Pada sayatan melintang tubuh P. darnleiensis tampak nefridia (Gambar 9a) dan nefrostom (Gambar 9b). Nefridia terletak di bawah lapisan otot longitudinal.
(a) (b)
Gambar 9 Fotomikrograf nefridia P. darnleiensis: (a) nefridia dan (b) nefrostom. Garis skala = 0.1 mm.
13
(a) (b)
Gambar 10 Fotomikrograf sistem saraf P. darnleiensis: (a) ganglion serebral dan (b) tali saraf. Garis skala = 0.1 mm.
Reproduksi. Spermateka P. darnleiensis memiliki divertikula dan nefridia. Pada sayatan melintang spermateka P. darnleiensis (Gambar 11a), tampak kumpulan sperma yang tersimpan di dalamnya (Gambar 11b). Pada preparat histologi vesikula seminalis P. darnleiensis tampak kumpulan sperma (Gambar 11c) dan tampak pula tropozoit Monocystis (Gambar 11d). Prostat Pheretima berbentuk racemose. Pada sayatan melintang prostat P. darnleiensis, tidak tampak batas yang jelas antarsel (Gambar 11e).
(a) (b) (c)
(d) (e)
PEMBAHASAN
Identifikasi P. darnleiensis
Seluruh cacing yang dikoleksi dari lokasi pengamatan diawetkan dengan merendamnya di alkohol 70% untuk diidentifikasi. Proses identifikasi cacing dapat dilakukan berdasarkan karakter eksternal dan internal. Cacing juvenil tidak dapat diidentifikasi karena organ seksual sebagai ciri identifikasi (misalnya klitelum, spermateka, dan prostat) belum berkembang.
Amynthas, Metaphire, dan Pheretima termasuk ke dalam famili
Megascolecidae. Famili Megascolecidae secara umum memiliki ciri seta dengan ujung lancip dan susunan seta lumbricine atau perichaetine. Klitelum dimulai pada segmen 15 atau di depannya. Lubang jantan satu pasang, biasanya pada segmen 17 atau 18, jarang pada segmen 19. Lubang betina berpasangan atau satu di bagian tengah pada segmen 14. Kantong esofagus biasanya ada. Testis dua pasang pada segmen 10 dan 11, atau satu pasang pada segmen 10 atau 11 saja. Prostat biasanya ada, satu atau dua pasang. Ovari satu pasang pada segmen 13 (Stephenson 1930).
Pontoscolex termasuk dalam famili Glossoscolecidae. Ciri umum famili Glossoscolecidae adalah seta umumnya berujung tunggal, jarang berujung ganda, biasanya memiliki ornamen, umumnya berjumlah lebih dari delapan per segmen. Lubang dorsal tidak ada, lubang nuchal jarang ada. Klitelum biasanya dimulai di sebelah segmen 14. Lubang jantan berada pada daerah klitelum, biasanya di bagian anterior daerah klitelum atau di depan klitelum, jarang di belakangnya. Lambung umumnya satu, jarang lebih dari satu, terletak di depan segmen testis, sering terdapat satu lambung rudimenter di bagian belakang esofagus di sebelah segmen ovari. Meganefridial, jarang terdapat dua pasang nefridia per segmen. Bagian ectal vas deferens biasanya simpel, sering dengan muscular apparatus (bursa propulsoria atau kantong kopulasi), jarang dengan kelenjar prostat. Seta penis tidak ada, seta kopulatori sering ada (Stephenson 1930).
15
cacing memiliki lubang jantan setelah daerah klitelum, cacing termasuk famili Megascolecidae.
Cacing yang dikoleksi dalam penelitian ini yang termasuk famili Glossoscolecidae hanya satu genus, yaitu Pontoscolex. Pontoscolex memiliki jumlah seta sebanyak delapan tiap segmen dan lubang jantan pada segmen 23. Ciri ini membedakannya dari Amynthas, Metaphire, dan Pheretima yang memiliki jumlah seta lebih dari delapan tiap segmen dan lubang jantan pada segmen 18.
Dalam penelitan ini, cacing yang termasuk famili Megascolecidae adalah Amynthas, Metaphire, dan Pheretima. Penentuan genus cacing tersebut dilakukan
melalui pembedahan untuk mengamati kantong kopulasi pada ujung saluran prostat di segmen 18. Jika cacing memiliki kantong kopulasi, kemungkinan cacing tersebut adalah Metaphire atau Pheretima, sedangkan cacing yang tidak memiliki kantong kopulasi adalah Amynthas. Selanjutnya, untuk membedakan genus Metaphire dan Pheretima dilakukan pengamatan terhadap nefridia pada
spermateka yang terdapat di sekitar segmen 5–9. Cacing yang tidak memiliki nefridia pada spermateka adalah Metaphire dan cacing yang memiliki nefridia pada spermateka adalah Pheretima (Lampiran 1).
Cacing genus Pheretima diidentifikasi lebih lanjut hingga tingkat spesies. Dalam penelitian ini hanya ditemukan satu spesies Pheretima yaitu P. darnleiensis. Spesies ini memiliki ciri kantong kopulasi tanpa divertikula dan
kelenjar bertangkai, posisi spermateka bithecate, dan lubang spermateka pertama pada segmen 4/5.
Karakter eksternal dapat digunakan untuk mengidentifikasi P. darnleiensis, namun untuk memastikan spesiesnya, perlu dilakukan pembedahan untuk mengamati karakter internalnya yaitu sekum, kantong kopulasi, dan spermateka. Karakter eksternal yang dapat menjadi ciri identifikasi P. darnleiensis adalah klitelum, prostomium, seta, lubang jantan, lubang betina, lubang spermateka, dan lubang dorsal.
famili Megascolecidae dan juga dimiliki oleh Amynthas dan Metaphire (Stephenson 1930). Klitelum ini memanjang sepanjang tiga segmen dari segmen 14 hingga 16 dan berwarna agak pucat dibandingkan dengan segmen tubuh lainnya. Cacing yang masih muda tidak memiliki klitelum dan pendugaan spesiesnya harus dilakukan dengan mengamati karakter lain.
Prostomium. Prostomium P. darnleiensis berbentuk epilobus dengan dasar terbuka. Ciri ini juga dimiliki oleh Amynthas dan Metaphire. Prostomium merupakan cuping yang terletak paling anterior (Edwards & Lofty 1972). Prostomium terletak di depan mulut dan dapat berfungsi sebagai organ perasa lingkungan serta membantu menggenggam substrat (Blakemore 2002). Selain itu, prostomium merupakan karakter morfologi yang dapat menjadi ciri identifikasi (Sims & Easton 1972). Prostomium melekat pada segmen pertama, yaitu peristomium (Blakemore 2002). Prostomium terkadang sulit diamati pada cacing berukuran kecil.
Seta. Seta berfungsi sebagai alat gerak cacing. Pheretima darnleiensis memiliki tipe seta perichaetine, yaitu tersebar baik pada bagian ventral maupun dorsal dalam tiap segmen kecuali pada bagian peristomium dan pigidium (Blakemore 2002). Jumlah seta pada P. darnleiensis sekitar 40 (dihitung pada segmen 13).
Lubang jantan. Lubang jantan sering terletak pada tonjolan yang biasa disebut kantong kopulasi (Blakemore 2002). Pada kantong kopulasi terdapat ujung vas deferens dan kantong kopulasi dapat membalik keluar membentuk penis sementara pada saat cacing berkopulasi (Stephenson 1930). Pheretima darnleiensis memiliki kantong kopulasi pada bagian lubang jantannya. Lubang
jantan P. darnleiensis pada segmen 14 kontak dengan lubang spermateka pasangannya pada segmen 4/5–7/8 saat berkopulasi. Cairan seminal ditransfer ke spermateka cacing pasangan melalui lubang jantan (Edwards & Lofty 1972). Lubang jantan P. darnleiensis tidak memiliki divertikula dan kelenjar bertangkai. Lubang ini dapat dilihat tanpa bantuan mikroskop.
17
pada tiap spesies cacing. Pada Pheretima, dalam satu kokon dapat terdiri atas tiga sel telur. Sperma akan dimasukkan ke sel telur saat melewati lubang spermateka. Fertilisasi terjadi secara eksternal. Kokon akan dilepaskan melalui bagian kepala cacing (Edwards & Lofty 1972). Lubang betina P. darnleiensis dapat dilihat tanpa bantuan mikroskop, namun lubang ini akan menjadi sulit dilihat pada cacing yang telah lama diawetkan.
Lubang spermateka. Lubang spermateka pada segmen 4/5–7/8 menghubungkan spermateka yang terletak pada segmen 5–8 dengan lingkungan luar tubuh. Sperma cacing pasangan dimasukkan melalui lubang saat berkopulasi. Divertikula pada spermateka berfungsi untuk menyimpan sperma yang diterima saat kopulasi (Stephenson 1930). Lubang spermateka P. darnleiensis terkadang sulit dilihat. Hal ini dapat disebabkan oleh kontraksi cacing saat pembiusan.
Lubang dorsal. Lubang dorsal menghubungkan rongga tubuh dengan lingkungan luar (Stephenson 1930). Lubang dorsal juga merupakan tempat mensekresikan cairan selom untuk menjaga kelembaban tubuh cacing (Blakemore 2002). Lubang dorsal P. darnleiensis terletak di seluruh segmen tubuh kecuali pada bagian anterior dari segmen 1–11.
Sekum. Sekum merupakan divertikula pada intestinum (Stephenson 1930). Sekum P. darnleiensis terletak pada segmen 27 dan memanjang hingga segmen 24. Hal ini didukung oleh Sims & Easton (1972) yang menyatakan sekum P. darnleiensis terletak di segmen 27 dan memanjang hingga segmen anatara 23–25. Fungsi sekum belum banyak diketahui, diduga sekum befungsi sebagai tempat mikroorganisme simbion (Blakemore 2002).
Kantong kopulasi. Kantong kopulasi terletak di ujung saluran prostat yang kontak dengan lingkungan luar. Secara eksternal, keberadaan kantong kopulasi dapat dilihat dari bentuk lubang jantan. Jika terdapat kantong kopulasi, lubang jantan tampak agak menonjol atau memiliki lubang agak lebar.
Spermateka. Spermateka hanya dapat diamati dengan membedah cacing. Selain bentuk spermateka, keberadaan nefridia juga menjadi ciri pembeda. Nefridia terletak pada dasar spermateka dan tampak seperti serabut.
Borneo. Sims & Easton (1972) menemukan P. darnleiensis dengan panjang 7.5– 17 cm, klitelum pada segmen 14–16, dan jumlah seta 12–35 pada segmen 7 dan 38–45 pada segmen 20. Lubang dorsal pertama pada segmen 12/13, terkadang pada segmen 11/12. Lubang jantan sepasang dengan kantong kopulasi pada segmen 18. Lubang betina satu pada segmen 14. Lubang spermateka berpasangan, besar, lateral pada segmen (4/5), 5/6, 6/7, 7/8, 8/9.
Histologi P. darnleiensis
Pengamatan histologi organ P. darnleiensis diawali dengan melakukan pembedahan. Sayatan dari anterior (sekitar segmen 3) hingga ke posterior (melewati segmen 30) dibuat pada bagian dorsal P. darnleiensis (Gambar 3). Hal ini dilakukan karena organ-organ P. darnleiensis melekat pada bagian ventral sehingga pembedahan pada bagian dorsal tidak merusak organ tersebut.
19
Gambar 12 Diagramatik sayatan melintang Lumbricus (Sherman & Sherman 1970).
Sirkulasi. Pembuluh darah dorsal mengalirkan darah dari posterior menuju anterior dan melalui jantung mengalirkan darah ke pembuluh darah ventral. Pembuluh dorsal terletak berdekatan pada bagian dorsal saluran pencernaan. Pembuluh darah ventral mengalirkan darah dari anterior menuju posterior dan menyalurkan darah ke berbagai organ tubuh cacing (Edwards & Lofty 1972).
Ekskresi. Nefridia merupakan organ eksresi Oligochaeta (Stephenson 1930). Cairan selom masuk melalui nefrostom dan disaring dalam tubulus. Pada Pheretima, nefridia terbuka kembali ke saluran pencernaan sehingga hasil
ekskresi P. darnleiensis dibuang lewat anus (Edwards & Lofty 1972).
Saraf. Ganglion serebral berkaitan dengan bagian anterior, yaitu prostomium yang merupakan organ sensori. Ganglion serebral dihubungkan oleh saraf sirkumfaringeal ke tali saraf yang terletak memanjang di bagian ventral (Edwards & Lofty 1972). Serabut kasar terletak pada bagian dorsal tali saraf. Serabut ini berfungsi memfasilitasi penghantaran impuls (Stephenson 1930).
21
(Reinhart & Dollahon 2003). Prostat Pheretima berbentuk racemose (Blakemore 2002). Prostat mensekresikan semen bagi sperma (Stephenson 1930).
SIMPULAN
P. darnleiensis memiliki tubuh silindris dengan warna keseluruhan tubuh
gelap dengan bagian dorsal lebih gelap daripada bagian ventral dan banyak seta tersebar per segmen (sekitar 40 pada segmen 13). Sekum terletak pada segmen 27. Pada dasar prostat terdapat kantong kopulasi tanpa divertikula dan kelenjar bertangkai. Pada spermateka terdapat nefridia, posisi spermateka bithecate, dan lubang spermateka pertama pada segmen 4/5.
Klitelum P. darnleiensis memiliki lapisan kelenjar mukus, kelenjar pensekresi kokon, dan kelenjar pensekresi albumin. Faring bagian dalam P. darnleiensis memiliki epitel kolumnar bersilia, pada lambungnya terdapat lapisan
3 EKOLOGI
Pheretima darnleiensis
PENDAHULUAN
Persebaran cacing tanah dipengaruhi kondisi biotik maupun abiotik lingkungannya (Gonzalez et al. 2007). Faktor biotik yang mempengaruhi persebaran cacing tanah misalnya keberadaan cacing lain (Baker et al. 2002), tumbuhan (Nadeem et al. 2007), dan artropoda (Gonzalez et al. 2003). Baker et al. (2002) menyatakan keberadaan cacing Aporrectodea longa dan A. caliginosa
dapat menurunkan kelimpahan dan biomassa cacing A. trapezoides, Microscolex dubius, M. phosphoreus, Spenceriella macleayi, dan Heteroporodrilus sp., namun Monroy et al. (2003) tidak menemukan bukti bahwa introduksi A. longa menurunkan kelimpahan A. caliginosa. Tumbuhan dapat berfungsi sebagai kanopi bagi cacing tanah. Nadeem et al. (2007) memperoleh lebih banyak cacing pada lokasi ternaungi tumbuhan. Gonzalez et al. (2003) memperoleh artropoda seperti Acarina dan Colembola pada lokasi penelitiannya. Migge-Kleian et al. (2006) menyatakan adanya hubungan antara keberadaan artropoda dan cacing.
23
63.6%, pH 7.2, C total 0.72%, dan N total 0.22% dan Yeates (1991) menemukan cacing pada lokasi dengan kelembaban tanah 31% dan karbon organik 4.46%.
Tekstur tanah menunjukkan kekasaran tanah dari fraksi tanah halus dan dapat dibedakan menjadi pasir (ukuran 2000–50 µm), debu (50–2 µm), dan liat (< 2 µm) (Hardjowigeno 2007). Tanah lempung berpasir memiliki kelimpahan cacing yang tinggi (Edwards & Lofty 1972). Tian et al. (2000) menemukan rata-rata 150 individu cacing (dominan Hyperiodrilus africanus dan Eudrilus eugeniae) pada kondisi tanah dengan tekstur lempung berpasir.
Pada umumnya, cacing tanah bersarang di tanah lembab dan melakukan eksplorasi untuk makan dan kawin pada malam hari (Hickman et al. 1984). Kedalaman bersarang cacing berkisar antara 5–240 cm dan pada musim panas cacing bersarang di tempat yang lebih dalam yang temperaturnya lebih konstan (Stephenson 1930). Smith et al. (2008) lebih banyak menemukan cacing tanah pada kedalaman 0–25 cm. Cacing epigeik dapat ditemukan beberapa sentimeter dari permukaan tanah, sedangkan cacing endogeik dapat ditemukan di kedalaman 10–15 cm atau lebih dalam lagi (Edwards & Bohlen 1996). Pheretima darnleiensis memakan bahan organik membusuk dan hidup di kedalaman
beberapa sentimeter dari permukaan tanah sehingga termasuk kategori epigeik. Densitas cacing dalam suatu lokasi berbeda-beda. Mergia (2007) menemukan empat jenis cacing (Eiseniella, Dendrobaena, Allelobophora, dan Aporrectodea) dengan rata-rata 3.5 individu/m2 pada lahan pertanian. Kelimpahan cacing di Jaguapita, Brazil, berkisar 3–739 individu/m2 di daerah berumput, 0–29 individu/m2 di daerah pertanian, dan 3–13 individu/m2 di daerah hutan. Rata-rata jumlah cacing yang ditemukan di lokasi tersebut cenderung lebih tinggi pada kedalaman 0–30 cm terutama pada musim hujan (Nunes et al. 2006).
Koleksi cacing tanah dapat dilakukan dengan menggunakan aliran listrik (Butt & Lowe 2004), mostar (Nuutinen et al. 2001), bola lampu (Tisdall & McKenzie 1999), dan suara dengan frekuensi 97.3 ± 11.7 Hz (Mitra et al. 2009). Metode menggali dan menyortir dengan tangan merupakan metode yang paling sederhana dan umum dilakukan (Jimenez et al. 2000).
pertumbuhannya jika dibudidayakan dalam kondisi yang tidak cocok. Oleh karena itu, perlu diketahui kondisi lingkungan tempat ditemukannya P. darnleiensis untuk membuat habitat yang mendukung pertumbuhannya dalam proses budidaya. Tujuan penelitian ini adalah mempelajari kondisi lingkungan biotik dan abiotik tempat ditemukannya P. darnleiensis. Faktor biotik yang dipelajari adalah keberadaan cacing lain, tumbuhan, dan artropoda di lokasi tempat P. darnleiensis dikoleksi. Sedangkan faktor abiotik yang dipelajari adalah tutupan komponen abiotik (seperti batu, bata, dan plastik), temperatur tanah, pH tanah, kelembaban tanah, kandungan C organik, N, P, K, dan tekstur tanah di lokasi tempat P. darnleiensis dikoleksi.
BAHAN DAN METODE
Waktu dan Tempat Penelitian
Penelitian kondisi tempat ditemukannya P. darnleiensis berlangsung dari bulan Februari 2010–April 2010. Koleksi sampel dilakukan di lingkungan kampus IPB Darmaga Kabupaten Bogor dan kampus IPB Baranangsiang Kotamadya Bogor. Analisis tanah dilakukan di Laboratorium Departemen Ilmu Tanah dan Sumber Daya Lahan Fakultas Pertanian IPB.
Alat dan Bahan
Alat yang digunakan untuk mempelajari habitat P. darnleiensis adalah sekop, meteran, termometer tanah, soil pH & moisture tester (Takemura Electric Works, Ltd), oven, dan saluran Tullgren. Bahan yang digunakan untuk mempelajari habitat P. darnleiensis adalah alkohol 70%.
Pengamatan Faktor Biotik
Koleksi cacing tanah. Sebagai cacing epigeik, P. darnleiensis dapat ditemukan pada lapisan atas tanah. Maka, kedalaman lokasi sampling konsisten sedalam 20 cm kecuali pada lokasi 1 karena kedalaman selokan hanya 5 cm. Pheretima darnleiensis dikoleksi dengan menggali petak berukuran sekitar 55–
25
(modifikasi metode Cannavacciuolo et al. 1998; Margerie et al. 2001). Penggalian dilakukan di 10 lokasi, 6 lokasi di Kampus IPB Darmaga Kabupaten Bogor dan 4 lokasi di Kampus IPB Baranangsiang Kotamadya Bogor. Seluruh cacing yang dikoleksi dari lokasi penelitian diawetkan dalam alkohol 70% untuk diidentifikasi dengan kunci identifikasi Sims & Easton (1972).
Penghitungan persen tutupan tumbuhan lokasi pengamatan. Penghitungan persen tutupan tumbuhan dilakukan dengan mengukur lokasi pengamatan yang tertutup oleh tumbuhan dengan menggunakan meteran. Persen tutupan dihitung dengan rumus:
% tutupan tumbuhan = luas tutupan tumbuhan x 100% luas lokasi pengamatan
Tumbuhan yang dikoleksi dari lokasi pengamatan dibuat herbarium untuk diidentifikasi. Pembuatan herbarium dilakukan dengan merentangkan dan menjepit tumbuhan dengan kertas koran selama satu hari, lalu disimpan dalam oven 45 oC selama satu hari. Identifikasi tumbuhan hingga tingkat ordo dilakukan dengan bertanya kepada peneliti bagian taksonomi tumbuhan dan membandingkannya dengan buku identifikasi Naiola (1986) dan Wiegand (1999).
Gambar 13 Saluran Tullgren untuk mengekstraksi artropoda tanah.
Pengamatan Faktor Abiotik
Penghitungan persen tutupan komponen abiotik lokasi pengamatan. Penghitungan persen tutupan komponen abiotik (seperti puing, plastik, dan batu) dilakukan dengan mengukur lokasi pengamatan yang tertutup oleh komponen abiotik dengan menggunakan meteran. Persen tutupan komponen abiotik dihitung dengan rumus:
% tutupan komponen abiotik = luas tutupan komponen abiotik x 100% luas lokasi pengamatan
Pengukuran temperatur, pH, dan kelembaban tanah. Temperatur tanah diukur dengan menggunakan termometer tanah. Pengukuran pH dan kelembaban tanah dilakukan dengan menggunakan soil pH & moisture tester (Takemura Electric Works, Ltd). Dalam satu lokasi pengamatan, pengukuran dilakukan di lima titik yaitu pada empat titik sudut dan satu di tengah lokasi pengamatan (Kulmatiski et al. 2004).
Analisis tanah. Sampel tanah diambil di lima titik (empat titik sudut dan satu di tengah) masing-masing sebanyak 200 gram dengan menggunakan sekop dan diaduk. Analisis tanah dilakukan di Laboratorium Departemen Ilmu Tanah dan Sumber Daya Lahan Fakultas Pertanian IPB, meliputi kandungan C organik, N, P, K, dan teksturnya. Kandungan C organik dianalisis dengan metode Walkley-Black, N dengan metode Kjeldahl, sedangkan P dan K dengan spektrofotometri
27
terlebih dahulu dengan penyaringan, lalu fraksi debu dan liat dipisahkan dengan menggunakan pipet untuk diukur bobotnya (Gee & Bauder 1986).
Analisis Data
Analisis komponen utama dilakukan untuk menentukan variabel yang mempengaruhi keberadaan P. darnleiensis pada suatu lokasi (Everitt & Hothorn 2006). Korelasi Pearson dihitung untuk melihat hubungan antara kelimpahan P. darnleiensis dan variabel lingkungan (Irianto 2004). Analisis data dilakukan
dengan menggunakan program R 2.4.0 (Everitt & Hothorn 2006).
HASIL
Pengambilan sampel dilakukan pada sepuluh lokasi. Lokasi 1–6 terdapat di kampus IPB Darmaga Kabupaten Bogor dan lokasi 7–10 terdapat di kampus IPB Baranangsiang Kotamadya Bogor. Sebanyak lima lokasi ternaungi tumbuhan dan lima lokasi terpapar cahaya matahari (Tabel 3).
Pheretima darnleiensis dikoleksi pada lima lokasi yaitu lokasi 3, 4, 5, 8, dan
9 (Tabel 4). Cacing yang mendominasi lokasi 3 adalah Amynthas (Tabel 1). Lokasi ini tertutup seluruhnya oleh serasah Glumiflorae (Tabel 5). Hymenoptera mendominasi lokasi ini (Tabel 6). Tidak ada komponen abiotik yang menutupi lokasi tersebut dan kandungan kalium pada lokasi ini paling tinggi dibandingkan lokasi lainya (Tabel 4). Artropoda yang terdapat di lokasi 4 hanya Isoptera (Tabel 6). Lokasi tersebut tidak ditutupi oleh komponen abiotik (Tabel 4). Pada lokasi 5 dikoleksi seluruh genus cacing (Amynthas, Metaphire, Pheretima, dan Pontoscolex) dan P. darnleiensis paling banyak dikoleksi di lokasi ini (Tabel 1).
28
Tabel 3 Koordinat dan deskripsi lokasi koleksi empat genus cacing di kampus IPB Darmaga Kabupaten Bogor (lokasi 1–6) dan kampus IPB Baranangsiang Kotamadya Bogor (lokasi 7–10)
Lokasi Koordinat
Daerah laboratorium tingkah laku hewan departemen Biologi, petak tanah di antara aspal dan trotoar, tertutup puing keramik, terdapat rumput (Glumiflorae).
3 S 06
o33’22.9”
E 106o43’20.4” 100 x 100 x 20 Ya
Daerah laboratorium tingkah laku hewan departemen Biologi, terdapat tempat sampah di samping lokasi, tanah dalam kondisi setelah digali, terdapat sisa rumput dan sisa batang kayu.
4 S 06
o33’35.3”
E 106o43’42.7” 100 x 100 x 20 Ternaungi
Seberang FEMA di bawah pohon kelapa sawit, terdapat tutupan tumbuhan paku (Eufilicales) dan daun kelapa sawit (Sapindales).
5 S 06
o33’22.5”
E 106o43’21.6” 100 x 100 x 20 Ternaungi
Belakang asrama Sylvasari, lapangan rumput (Glumiflorae), terdapat tumpukan puing (batu, bata, keramik).
6 S 06
o33’35.8”
E 106o43’29.5” 70 x 70 x 20 Ya Daerah rektorat, terdapat tumpukan puing, tidak ada tutupan tumbuhan.
7 S 06
o36’02.8”
E 106o48’27.1” 80 x 80 x 20 Ya
Gundukan tanah di bawah tangga, terdapat tembok batu di satu sisi gundukan, terdapat rumput (Glumiflorae).
E 106o48’28.6” 100 x 100 x 20 Ternaungi Daerah tempat parkir, tertutup serasah daun Ficus (Rosales) dan batu.
10 S 06
o35’57.5”
E 106o48’27.2” 90 x 70 x 20 Ya
29
29
Tabel 4 Karakter biotik dan abiotik lokasi koleksi empat genus cacing
LOK CACING PHERE TUMB HEWAN ABIO TEMP pH rH C N P K PASIR DEBU LIAT
1 4.85 0.00* 100.00* 145.24* 0.00* 26.82 5.88 28.61 5.18 0.47 240.60 200.00 22.68 13.95* 63.37
2 3.85 0.00* 29.00 32.00 100.00 30.23 5.91 21.00 2.07 0.18 213.50 390.00 27.71 6.46* 65.83
3 16.00 1.00* 100.00* 23.18 0.00* 31.71 5.59 25.17 2.47 0.23 414.20 1050.00* 20.20 7.68* 72.12
4 4.00 1.00* 35.00 39.58 0.00* 26.67 5.35 24.98 1.35 0.14 223.00 600.00 10.50 16.24* 73.26
5 15.00 7.00 25.00 16.32 78.50 27.42 5.77 19.83 3.39 0.29 286.80 500.00 28.76 38.74 32.50
6 4.08 0.00* 0.00* 58.33 100.00 31.45 5.37 32.25 3.03 0.26 254.90 892.00 55.33 9.95* 34.72
7 10.94 0.00* 44.00 48.84 55.96 29.68 5.93 19.75 4.22 0.44 457.50 600.00 75.07* 6.67* 18.26
8 23.00 1.00* 48.00 35.00 49.75 26.44 6.05 10.17 2.71 0.29 436.50 750.00 63.11 13.80* 23.09
9 31.00 1.00* 44.00 172.13* 46.88 26.27 6.33 6.67 3.83 0.36 482.70 700.00 58.48 12.63* 28.89
10 25.40 0.00* 29.73 22.21 64.42 26.57 6.22 9.92 4.39 0.45 390.30 750.00 66.89 10.53* 22.58
MEAN 13.81 1.10 45.47 59.28 49.55 28.33 5.84 19.84 3.26 0.31 340.00 643.00 42.87 13.67 43.46
SD 10.00 2.13 31.75 54.24 38.82 2.20 0.33 8.51 1.17 0.12 106.02 244.32 23.14 9.41 22.36
MIN 3.85 0.00 0.00 16.32 0.00 26.27 5.35 6.67 1.35 0.14 213.50 200.00 10.50 6.46 18.26
MAX 31.00 7.00 100.00 172.13 100.00 31.71 6.33 32.25 5.18 0.47 482.70 1050.00 75.07 38.74 73.26
(*) = nilai lebih tinggi atau lebih rendah daripada nilai pada lokasi 5. Daerah diarsir merupakan lokasi P. darnleiensis paling banyak ditemukan. LOK = lokasi
pengambilan sampel, CACING = jumlah cacing selain P. darnleiensis/m2, PHERE = jumlah P. darnleiensis/m2, TUMB = % rata-rata tutupan tumbuhan, HEWAN =
Tabel 5 Tumbuhan yang terdapat di lokasi koleksi empat genus cacing
Lokasi Ordo Tumbuhan Jumlah Tutupan (%) Total tutupan (%)
1 Contortae + Malvales 100.00 100.00
Daerah diarsir merupakan lokasi P. darnleiensis paling banyak ditemukan.
Sebanyak tujuh individu P. darnleiensis dikoleksi di lokasi 5 dan masing-masing satu individu P. darnleiensis pada lokasi 3, 4, 8, dan 9. Berdasarkan perhitungan standar deviasi, jumlah P. darnleiensis pada lokasi 5 secara signifikan paling tinggi dibandingkan dengan lokasi lainnya (Tabel 4). Pada lokasi 5 dikoleksi tujuh individu P. darnleiensis dan lima belas individu cacing selain P. darnleiensis (Tabel 1). Glumiflorae mendominasi tutupan tumbuhan di lokasi 5
31
Tabel 6 Artropoda tanah yang terdapat di lokasi koleksi empat genus cacing
Lokasi Ordo Artropoda Tanah Jumlah
(individu/kg tanah segar )
Tabel 7 Kisaran temperatur, pH, dan kelembaban tanah lokasi koleksi empat
Daerah diarsir merupakan lokasi P. darnleiensis paling banyak ditemukan.
Lokasi yang terdapat P. darnleiensis dibandingkan dengan lokasi tanpa P. darnleiensis dengan uji t. Hasil analisis uji t antara lokasi yang terdapat P.
darnleiensis dan tanpa P. darnleiensis menunjukkan nilai P dari perbandingan
kelimpahan cacing selain P. darnleiensis = 0.23, tutupan tumbuhan = 0.65, jumlah artropoda tanah = 0.91, tutupan komponen abiotik = 0.26, temperatur tanah = 0.40, pH tanah = 0.72, kelembaban tanah = 0.39, C organik = 0.34, N = 0.20, P = 0.43, K = 0.35, fraksi pasir = 0.39, fraksi debu = 0.20, dan fraksi liat = 0.74. Nilai-nilai tersebut menunjukkan variabel antara lokasi dengan P. darnleiensis dan tanpa P. darnleiensis tidak berbeda signifikan (P > 0.05). Seluruh nilai variabel pada lokasi yang ternaungi tumbuhan juga dibandingkan dengan lokasi yang terpapar cahaya matahari dengan uji t. Hasil analisis uji t antara lokasi ternaungi tumbuhan dan terpapar cahaya matahari menunjukkan nilai P dari perbandingan kelimpahan cacing selain P. darnleiensis = 0.61, kelimpahan P. darnleiensis = 0.23, tutupan tumbuhan = 0.65, jumlah artropoda tanah = 0.24, tutupan komponen abiotik = 0.26, temperatur tanah = 0.02, pH tanah = 0.75, kelembaban tanah = 0.54, C organik = 0.95, N = 0.98, P = 0.87, K = 0.25, fraksi pasir = 0.43, fraksi debu = 0.09, dan fraksi liat = 0.92. Hasil tersebut menunjukkan hanya temperatur yang berbeda signifikan (P < 0.05). Lokasi yang ternaungi tumbuhan umumnya memiliki temperatur lebih rendah daripada lokasi yang terpapar cahaya matahari (Tabel 7).
33
kelimpahan P. darnleiensis, tutupan tumbuhan, dan fraksi debu berada dekat dengan lokasi 5 (Gambar 14). Korelasi antara kelimpahan P. darnleiensis dan tutupan tumbuhan sebesar 0.14 (P = 0.69), sedangkan korelasi antara kelimpahan P. darnleiensis dan fraksi debu sebesar 0.82 (P < 0.05) (Tabel 8).
34
Tabel 8 Nilai korelasi log antarvariabel pada sepuluh lokasi koleksi empat genus cacing
CACING PHERE TUMB HEWAN ABIO TEMP pH rH C N P K PASIR DEBU LIAT
CACING 0.33 0.32 0.74 0.47 0.28 0.03 0.00 0.25 0.16 0.00 0.19 0.11 0.65 0.06
PHERE 0.35 0.69 0.27 0.98 0.46 0.81 0.72 0.62 0.55 0.85 0.73 0.46 0.00 1.00
TUMB 0.35 0.14 0.93 0.13 0.28 0.21 0.39 0.81 0.64 0.39 0.36 0.43 0.99 0.60
HEWAN -0.12 -0.39 0.03 0.63 0.50 0.53 0.74 0.32 0.33 0.91 0.29 0.73 0.70 0.94
ABIO 0.26 0.01 -0.51 -0.17 0.88 0.24 0.26 0.49 0.55 0.56 0.55 0.01 0.93 0.02
TEMP -0.38 -0.26 -0.38 -0.24 0.06 0.14 0.06 0.62 0.44 0.73 0.38 0.95 0.08 0.49
pH 0.69 -0.09 0.43 0.23 0.41 -0.50 0.00 0.07 0.05 0.08 0.70 0.06 0.84 0.10
rH -0.82 -0.13 -0.31 -0.12 -0.39 0.62 -0.85 0.48 0.33 0.03 0.50 0.11 0.83 0.08
C 0.40 -0.18 0.09 0.35 0.25 -0.18 0.59 -0.25 0.00 0.24 0.45 0.06 0.94 0.10
N 0.48 -0.22 0.17 0.34 0.22 -0.27 0.64 -0.35 0.98 0.12 0.56 0.04 0.98 0.05
P 0.88 0.07 0.31 0.04 0.21 -0.13 0.59 -0.68 0.41 0.52 0.11 0.05 0.62 0.03
K 0.45 0.12 -0.32 -0.37 0.22 0.31 -0.14 -0.24 -0.27 -0.21 0.54 0.39 0.60 0.38
PASIR 0.53 -0.27 -0.28 0.12 0.76 -0.02 0.61 -0.53 0.62 0.66 0.64 0.30 0.49 0.00
DEBU 0.16 0.82 0.00 -0.14 -0.03 -0.58 -0.08 -0.08 0.03 -0.01 -0.18 -0.19 -0.25 0.91
LIAT -0.61 0.00 0.19 0.03 -0.70 0.25 -0.55 0.57 -0.55 -0.63 -0.67 -0.31 -0.90 -0.04
35
PEMBAHASAN
Perbedaan Lokasi dengan P. darnleiensis dan Lokasi Tanpa P. darnleiensis
Pheretima darnleiensis dikoleksi di lokasi 3, 4, 5, 8, dan 9 sebanyak
masing-masing satu individu kecuali di lokasi 5 sebanyak tujuh individu. Banyaknya P. darnleiensis yang dikoleksi di lokasi 5 (seluruhnya telah memiliki klitelum) menunjukkan kondisi lingkungan pada lokasi tersebut cocok bagi P. darnleiensis. Sedangkan pada lokasi 3, 4, 8, dan 9, P. darnleiensis hanya dikoleksi
sebanyak satu individu pada masing-masing lokasi. Pheretima darnleiensis yang dikoleksi pada lokasi-lokasi tersebut kemungkinan tidak mendiami lokasi tersebut. Perbedaan lokasi yang terdapat P. darnleiensis dengan yang tidak dianalisis dengan uji t. Seluruh nilai variabel tidak berbeda signifikan. Berdasarkan hasil analisis tersebut, kondisi lingkungan tempat P. darnleiensis dikoleksi tidak berbeda signifikan dengan lingkungan tempat dikoleksinya cacing selain P. darnleiensis (Amynthas, Metaphire, Pontoscolex, dan cacing juvenil). Hal ini
sesuai dengan Sims & Easton (1972) yang menyatakan bahwa cacing genus Pheretima belum diketahui memiliki kondisi habitat yang khusus.
Keberadaan P. darnleiensis pada lokasi 3, 4, 5, 8, dan 9 mungkin berkaitan dengan naungan. Nadeem et al. (2007) menyatakan bahwa cacing lebih banyak ditemukan di bawah kanopi. Lokasi 4, 5, 8, dan 9 ternaungi sehingga temperatur lebih stabil (P = 0.02). Walaupun demikian, pH tanah dan kelembaban tanah tidak dipengaruhi naungan (P = 0.75 dan 0.54). Variabel lainnya juga tidak berbeda signifikan. Pada lokasi yang ternaungi tumbuhan, jumlah cacing selain P. darnleiensis yang dikoleksi lebih banyak daripada lokasi yang terpapar cahaya
matahari, namun tidak signifikan (P = 0.61). Jumlah P. darnleiensis yang dikoleksi pada lokasi ternaungi tumbuhan dengan lokasi terpapar cahaya matahari tidak berbeda signifikan (P = 0.23). Hal ini disebabkan oleh tingginya jumlah P. darnleiensis yang dikoleksi pada lokasi 5 sehingga membuat standar deviasi pada
lokasi terpapar menjadi tinggi. Dengan demikian, keberadaan P. darnleiensis dalam penelitian ini tidak berkaitan dengan naungan.
Pheretima darnleiensis dikoleksi di lokasi 5 sebanyak tujuh individu.
standar deviasi. Hasil analisis menunjukkan bahwa nilai-nilai variabel pada lokasi 5 tidak berbeda dengan lokasi lainnya kecuali pada komponen debu. Pada lokasi 5 komponen debu mencapai 38.74% dan paling tinggi dibanding lokasi lainnya. Fraksi tanah debu dapat menyimpan air lebih baik daripada fraksi tanah pasir dan memiliki porositas yang lebih baik daripada tanah liat (Hardjowigeno 2007). Berkaitan dengan hal tersebut, P. darnleiensis banyak dikoleksi di lingkungan dengan air dan udara tanah yang cukup.
Korelasi antara Variabel Lingkungan dan Kelimpahan P. darnleiensis
Dalam penelitian ini, korelasi antara P. darnleiensis dan cacing lain (Amynthas, Metaphire, Pontoscolex, dan cacing juvenil) sebesar 0.33 dan tidak signifikan (Tabel 8). Dengan demikian, kelimpahan P. darnleiensis tidak dipengaruhi keberadaan cacing lain. Hal ini dapat disebabkan oleh rendahnya kelimpahan P. darnleiensis sehingga peluang untuk menemukan P. darnleiensis lebih tinggi pada lokasi yang banyak terdapat cacing. Selain itu, kondisi habitat P. darnleiensis juga mirip dengan kondisi habitat cacing lain.
Gonzalez et al. (1996) menyatakan bahwa jenis tumbuhan tidak berpengaruh terhadap densitas cacing tanah yang ditemukannya, namun hendaknya kesimpulan tersebut tidak digeneralisasi terhadap semua jenis tumbuhan. Sarlo (2006) menyatakan adanya hubungan antara biomassa cacing yang ditemukan dan jenis tumbuhan. Hal ini mungkin disebabkan oleh perbedaan kandungan nutrisi dari serasah tumbuhan tersebut. Nadeem et al. (2007) menyatakan bahwa cacing lebih banyak ditemukan di lokasi tumbuhan jambu biji dibandingkan dengan mangga, kelapa sawit, murbei, atau jeruk. Dalam penelitian ini, tumbuhan yang terdapat di lokasi penelitian sangat beragam sehingga sulit untuk menyimpulkan hubungan antara jenis tumbuhan dan kelimpahan P. darnleiensis. Tumbuhan memiliki nilai korelasi tidak signifikan dengan
kelimpahan P. darnleiensis (P = 0.14) (Tabel 8).
37
(terutama mikroartropoda dan enchytraeids) dalam suatu lokasi bergantung kondisi kelimpahan cacing tanah di lokasi tersebut sebelumnya. Kelimpahan artropoda dalam penelitian ini berkorelasi negatif dengan kelimpahan P. darnleiensis (-0.38) dan cacing selain P. darnleiensis (-0.11). Korelasi antara
artropoda dan kelimpahan P. darnleiensis maupun cacing selain P. darnleiensis tidak signifikan (P > 0.05) (Tabel 8). Dalam jangka panjang, kelimpahan cacing tanah dapat menurunkan kelimpahan mikroartropoda tanah akibat kompetisi sumber daya (Migge-Kleian et al. 2006).
Komponen abiotik dapat berfungsi sebagai mulsa. Mulsa merupakan material yang kontak dengan permukaan tanah. Mulsa dapat dapat mengurangi energi air hujan, menyaring, dan mereduksi kecepatan aliran permukaan sehingga menurunkan kapasitas transpor, meningkatkan infiltrasi air ke dalam tanah, dan mencegah erosi tanah baik oleh tumbukan air hujan ataupun karena aliran permukaan (Singer & Munns 2006). Katsvairo et al. (2007) menyatakan densitas cacing tanah berkorelasi positif dengan infiltrasi air. Mulsa juga meningkatkan proliferasi tanah dan dapat meningkatkan populasi cacing epigeik (Blanco-Canqui & Lal 2007). Komponen abiotik dalam penelitian ini dapat dikatakan tidak memiliki korelasi dengan kelimpahan P. darnleiensis dengan nilai korelasi 0.01 (Tabel 8). Interaksi antara P. darnleiensis dan variabel lain memiliki korelasi yang lebih tinggi daripada dengan mulsa.
Kandungan C organik, N, P, dan K pada lokasi 5 dapat dikatakan tidak berbeda dengan lokasi lainnya (Tabel 4). Menurut Hardjowigeno (2007), jumlah cacing berkaitan dengan bahan organik yang terkandung di dalamnya. Semakin banyak bahan organik, semakin gelap warna tanah, dan semakin tinggi kelimpahan cacing. Estevez et al. (1996) dan Karmegam & Daniel (2007) memperoleh korelasi positif antara kelimpahan cacing yang ditemukan dan karbon organik. Ivask et al. (2006) memperoleh cacing sebanyak 47–107 individu/m2 pada lingkungan dengan kandungan bahan organik = 3.45–4.16%, N = 0.158– 0.207%, P terlarut = 11.19–16.91 mg/100 g tanah kering, dan K+ = 17.24–21.32 mg/100 g tanah kering di lokasi penelitiannya, namun penelitian tersebut dilakukan di daerah Estonia sehingga mungkin saja terdapat perbedaan pengaruh temperatur dengan kelimpahan cacing yang diperoleh. Dalam penelitian ini, korelasi antara P. darnleiensis dan C organik = -0.18 serta cacing dan C organik = 0.40, keduanya tidak signifikan (Tabel 8). Menurut Mainoo (2008), kelimpahan cacing tanah tidak selalu berkorelasi positif dengan C organik. Ortiz-Ceballos & Fragoso (2004) juga memperoleh nilai korelasi negatif antara jumlah cacing yang diperoleh (Balanteodrilus pearsei dan Larsonidrilus orbiculatus) dengan bahan organik pada kedalaman 0–10 cm. Dalam penelitian ini, kemungkinan C organik telah habis dimakan cacing selain P. darnleiensis saat dilakukan sampling sehingga P. darnleiensis pergi mencari tempat baru yang memiliki kandungan C organik tinggi. Hal ini dapat mengakibatkan korelasi negatif P. darnleiensis dengan kandungan C organik. Dalam penelitian ini, korelasi antara kelimpahan P. darnleiensis dan N, P, serta K juga tidak signifikan (Tabel 8).
Pada lokasi 5, komponen debu memiliki nilai 38.74% dan berkorelasi positif signifikan dengan kelimpahan P. darnleiensis. Sedangkan korelasi debu dengan cacing lain sebesar 0.16 dan tidak signifikan (Tabel 8). Hal ini bertentangan dengan Ivask et al. (2006) yang menyatakan kelembaban tanah lebih mempengaruhi kelimpahan cacing dibandingkan dengan jenis tanah. Sama halnya dengan Rossi et al. (1997) yang tidak menemukan asosiasi antara kelimpahan Polypheretima elongata dan fraksi pasir, debu, maupun liat. Perbedaan jumlah
39
Variabel yang Mempengaruhi Kelimpahan P. darnleiensis
Biplot hasil analisis komponen utama (Gambar 14) menunjukkan variabel yang terletak berdekatan dengan lokasi 5 adalah tutupan tumbuhan, fraksi debu, dan kelimpahan P. darnleiensis. Variabel ini memiliki kontribusi terhadap lokasi 5 dan mungkin memiliki pengaruh terhadap kelimpahan P. darnleiensis. Sementara berdasarkan analisis korelasi, kelimpahan P. darnleiensis dengan tutupan tumbuhan memiliki nilai korelasi sebesar 0.14 dan kelimpahan P. darnleiensis dengan fraksi debu sebesar 0.82. Tumbuhan memiliki korelasi positif
tidak signifikan terhadap kelimpahan P. darnleiensis (P = 0.69). Hanya korelasi P. darnleiensis dengan debu yang memiliki nilai signifikan (P < 0.05). Sementara
itu, korelasi antara kelimpahan cacing selain P. darnleiensis dan fraksi debu sebesar 0.16 dan tidak signifikan (P = 0.65) (Tabel 8). Berdasarkan hasil analisis, dapat dikatakan bahwa semakin tinggi fraksi debu, maka semakin banyak P. darnleiensis yang ditemukan, namun semakin tinggi fraksi debu, belum tentu
jumlah cacing (selain P. darnleiensis) yang ditemukan semakin banyak. Dengan demikian, fraksi debu merupakan komponen yang berpengaruh terhadap kelimpahan P. darnleiensis.
SIMPULAN
4 PEMBAHASAN UMUM
Identifikasi cacing tanah dapat dilakukan dengan mengamati karakter eksternal. Karakter eksternal P. darnleiensis yang dapat menjadi ciri identifikasi adalah tubuh berbentuk silindris dan seta tersebar pada bagian ventral maupun dorsal. Ciri ini membedakan P. darnleiensis dari genus Planapheretima yang memiliki tubuh pipih dan seta tersebar di bagian ventral (Sims & Easton 1972). Warna tubuh P. darnleiensis sangat gelap. Warna ini dapat membedakannya dari cacing lain yang memiliki warna pucat seperti Pontoscolex. Gerakan P. darnleiensis juga sangat aktif dan ciri ini dapat membedakannya dari cacing yang
bergerak kurang aktif seperti Pontoscolex, namun untuk membedakan P. darnleiensis dengan cacing lain yang memiliki warna dan gerakan mirip, misalnya
Amynthas dan Metaphire, perlu dilakukan pembedahan untuk mengamati karakter
internalnya.
Karakter internal yang dapat menjadi ciri identifikasi adalah sekum, kantong kopulasi, dan spermateka. Keberadaan sekum pada segmen 27 membedakan P. darnleiensis dengan genus Ephemitra, Archipheretima, dan Metapheretima.
Keberadaan kantong kopulasi membedakan P. darnleiensis dengan Amynthas dan kantong kopulasi tanpa divertikula serta kelenjar bertangkai membedakannya dengan Parapheretima. Nefridia yang terdapat pada spermateka membedakan P. darnleiensis dengan Metaphire dan lubang spermateka pertama pada segmen 4/5
menjadi ciri spesies P. darnleiensis (Sims & Easton 1972).