CACING PARASITIK
PADA INSANG IKAN KEMBUNG (Decapterus spp.)
SKRIPSI
NOVA JULIANTI EMELINA
FAKULTAS KEDOKTERAN HEWAN INSTITUT PERTANIAN BOGOR
BOGOR 2008
ABSTRAK
NOVA JULIANTI EMELINA. Cacing Parasitik Pada Insang Ikan kembung (Decapterus spp.). Dibimbing oleh RISA TIURIA dan ADHI RACHMAT HARYADI.
Penelitian ini bertujuan untuk melihat adanya dan mengidentifikasi jenis cacing parasitik yang terdapat pada insang ikan kembung (Decapterus spp.) yang diambil dari tempat pelelangan ikan di daerah Jakarta. Delapan belas ekor ikan kembung digunakan dalam penelitian ini. Cacing parasitik diisolasi dari bagian insang, diwarnai dengan menggunakan pewarnaan minyak cengkeh dan KOH untuk nematoda serta Semichon’s Acetocarmin untuk trematoda. Pengamatan organ dalam setiap cacing yang berhasil diisolasi dilakukan dengan menggunakan mikroskop elektron, sedangkan pengukuran dilakukan menggunakan video mikro meter untuk mengukur panjang dan lebar tubuh cacing. Pengukuran ini bertujuan untuk mempermudah tahap identifikasi setiap cacing parasitik yang ditemukan. Seluruh data yang berhasil dikumpulkan dianalisa secara deskriptif. Hasil penelitian membuktikan adanya cacing parasitik pada insang ikan kembung yang berasal dari genus Pseudosteringophorus, Lecithocladium, Digenea A, Digenea B, Anisakis, Nematoda 1, Nematoda 2 serta serkaria digenea.
Kata kunci: Insang, Ikan Kembung, Digenea, dan Nematoda.
CACING PARASITIK
PADA INSANG IKAN KEMBUNG (Decapterus spp.)
NOVA JULIANTI EMELINA
Skripsi
Sebagai syarat untuk memperoleh gelar Sarjana Kedokteran Hewan pada
Fakultas Kedokteran Hewan
FAKULTAS KEDOKTERAN HEWAN INSTITUT PERTANIAN BOGOR
LEMBAR PENGESAHAN
Judul Penelitian : Cacing Parasitik pada Insang Ikan Kembung (Decapterus spp.) Nama : Nova Julianti Emelina
NRP : B04104163
Disetujui,
Pembimbing I Pembimbing II
Dr. Drh. Risa Tiuria, MS Adhi Rachmat Haryadi,Bsc.Msi NIP. 131 690 352
Mengetahui, Wakil Dekan
Fakultas Kedokteran Hewan IPB
Dr. Nastiti Kusumorini NIP. 131 669 942
KATA PENGANTAR
Puji dan syukur kepada Tuhan Yesus Kristus oleh karena berkat dan penyertaanNya yang luar biasa, penulis dapat menyelesaikan penelitian dengan baik serta dengan karunia dan kebaikanNya, penulis dapat menyelesaikan skripsi yang merupakan syarat kelulusan Sarjana Kedokteran Hewan. Skripsi ini disusun berdasarkan hasil penelitian cacing parasitik pada insang ikan kembung (Decapterus spp.) dan pada kesempatan ini penulis juga ingin menyampaikan terima kasih yang sebesar-besarnya kepada:
1 Dr. Drh. Risa Tiuria, MS selaku pembimbing pertama dan Adhi Rachmat Sudrajat Haryadi, Bsc.Msi, selaku pembimbing kedua, atas bimbingan, pengarahan dan masukan serta waktunya selama penelitian hingga selesainya skripsi ini.
2 Kedua orang tua, ”Papi” dan ”Mami” tersayang, orang tua yang luar biasa dan terbaik bagi penulis, untuk segala cinta kasih, waktu, tenaga dan doanya juga yang selalu memberikan dukungan baik moral maupun material kepada penulis.
3 Seseorang yang sangat berarti bagi penulis, Bang Junarto Halomoan Simamora Debataraja, Msi yang telah memberi banyak dukungan, doa, perhatian, dan bantuan mulai dari penelitian hingga proses penyusunan skripsi ini.
4 Dr. Drh. R. Hary Soehartono, MpappSc. PhD, selaku dosen pembimbing akademik.
5 Rekan sepenelitian: Ina, Linong, Sio, Vonti, Uya, Asri, Mones, Reni, Debi, Ari, Arios, Ivan, dan Dwi atas kerjasama, bantuan dan dukungannya. 6 Seluruh staf dan pegawai laboratorium Helmintologi (Pak Eman dan Bu Irawati) yang telah banyak membantu mulai dari awal hingga selesainya penelitian ini dengan lancar.
7 Tim “Sahabat Selamanya”: Linca, Tuti, dan Maria, yang selama ini memberi semangat, keceriaan, doa dan selalu ada bagi penulis dalam susah maupun senang.
8 Saudariku Cia atas doa, nasehat, kritikan, masukan dan dukungannya bagi penulis.
9 Kak Jani dan Kak Bosko, yang selama ini sudah menjadi kakak yang baik buat penulis, memberi semangat, doa, nasehat dan masukan yang sangat dibutuhkan penulis dalam menyelesaikan tugas akhirnya..
10 Kelompok PA: Kak Rade, Yanti, Othe, Irma, dan Ancel juga anak-anak PD 14 angkatan 2004 untuk nasehat, motivasi serta doanya.
11 Rekan-rekan seperjuangan Asteroidea 41: Ai, Vonte, Ina, Uya, Lina, Sio, Tongki, Teteg dan semuanya, atas semangat, kebersamaan dan kerjasamanya selama ini.
12 Keluarga besar Persekutuan Fakultas Kedokteran Hewan IPB. 13 Keluarga besar Persekutuan Mahasiswa Kristen IPB.
14 Pihak-pihak lain yang tidak dapat disebutkan satu persatu yang telah banyak membantu keberhasilan penulis.
Penulis menyadari banyaknya kekurangan dalam penulisan skripsi ini, oleh karena itu kritik dan saran dari pembaca sangat penulis harapkan dan penulis berharap agar skripsi ini dapat bermanfaat bagi banyak pihak termasuk penulis pribadi.
Bogor, Agustus 2008
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 3 Juli 1986 dan merupakan anak tunggal dari pasangan Bapak John Fardinanto Tumpal Sinaga dan Ibu Tumiar br. Nainggolan.
Penulis menyelesaikan pendidikan di Taman Kanak-Kanak dari tahun 1990-1992 di TK Tiranus, Jakarta dan TK Santa Clara, Depok, Sekolah Dasar Katolik (SDK) Slamet Riyadi II Jakarta pada tahun 1998, dilanjutkan ke Sekolah Lanjutan Tingkat Pertama Katolik (SLTPK) Slamet Riyadi Jakarta pada tahun 2001. Tahun 2004 penulis lulus dari Sekolah Menengah Umum Negeri (SMUN) 14 Jakarta dan pada tahun yang sama penulis berhasil lulus seleksi masuk IPB melalui jalur Saringan Penerimaan Mahasiswa Baru (SPMB) pada Fakultas Kedokteran Hewan.
Semasa aktif sebagai mahasiswa, penulis tercatat sebagai pengurus besar Persekutuan Fakultas Kedokteran Pewan, pengurus Himpro HKSA divisi pendidikan, anggota Paduan Suara FKH Gita Klinika, anggota Persekutuan Mahasiswa Kristen IPB, anggota Komisi Pelayanan Khusus PMK IPB. Penulis juga aktif dalam kepanitiaan organisasi lainnya, asisten praktikum Endoparasitik pada tahun 2008 serta menjadi staf pengajar salah satu bimbingan belajar di Bogor.
DAFTAR ISI Halaman DAFTAR GAMBAR……… ix DAFTAR TABEL………..….. x PENDAHULUAN….……….. 1 Latar Belakang………. 1 Tujuan………... 2 TINJAUAN PUSTAKA………... 3
Ikan kembung (Decapterus spp.)... 3
Trematoda……….…... 4
Digenea………... 5
Nematoda... 11
BAHAN DAN METODE PENELITIAN……….….. 18
Waktu dan Tempat Penelitian……… 18
Bahan dan Alat………... 18
Metode Penelitian………... 18
HASIL DAN PEMBAHASAN ……..………..…………..……....…... 21
Jenis Cacing Parasitik yang Ditemukan……… 21
Digenea………. 22 Genus Pseudosteringophorus……… 22 Genus Lecithocladium………... 26 Digenea A………. 31 Digenea B………. 31 Serkaria Digenea………... 32 Genus Anisakis……….. 34
KESIMPULAN DAN SARAN……….. 42
DAFTAR GAMBAR
No. Halaman
1. Ikan Kembung (Decapterus spp.)... 3
2. Helicometra spp. suatu digenea opecoelid pada ikan... 6
3. Morfologi umum trematoda digenea ………... 8
4. Siklus hidup trematoda digenea dalam menginfeksi ikan…………...… 8
5. Berbagi stadia perkembangan trematoda digenea………...…... 10
6. Morfologi umum cacing nematoda ……….……….…. 13
7. Siklus hidup langsung cacing nematoda ………...…....……… 14
8. Siklus hidup tidak langsung cacing nematoda………... 15
9. Siklus hidup tidak langsung ………...….………...……….. 16
10.Ikan sebagai induk semang paratenik……...………... 16
11.Larva nematoda Camallanus spp. di lingkungan perairan... 17
12.Genus Pseudosteringophosrus…...……….……… 24
13.Struktur batil isap oral dan ventral Pseudosteringophorus………. 25
14.Lecithocladium scombri dan Lecithocladium megalapsis ………... 29
15.Genus Lechitocladium dari insang kembung 10………….……..…..… 30
16.Struktur vitelaria dan ovarium Lecithocladium 1………....… 30
17.Preparat cacing, telur dan vitelaria digenea A dari insang kembung 1... 31
18.Preparat cacing digenea B dari insang kembung 7……….…… 32
19.Struktur telur, uterus dan vitelaria digenea B………... 32
20.Gambaran umum serkaria digenea ……….… 33
21.Preparat serkaria digenea dari insang kembung 2. ………. 33
22.Preparat serkaria digenea dari insang kembung 3 ……..………….….. 34
23.Preparat serkaria digenea dari insang kembung 4 ………...…………... 34
24.Struktur mulut Anisakis simplex…....………..…… 35
25.Ujung anterior famili Anisakidae stadium ketiga... 36
26.Siklus hidup Anisakis... 37
27.Ujung anterior yang memperlihatkan struktur kepala dan kutikula….. 38
28.Ujung posterior yang memperlihatkan struktur mukron... 39
DAFTAR TABEL
NO Halaman 1. Jenis, ukuran, dan jumlah cacing parasitic yang ditemukan ………….. 21 2. Perbandingan morfometrik dan ciri morfologis……….. 23
PENDAHULUAN
Latar Belakang
Indonesia sebagai salah satu negara yang sedang berkembang memiliki jumlah penduduk sekitar 242 juta (Anonim 2008). Tingginya jumlah penduduk Indonesia menuntut tingginya tingkat ketersediaan pangan untuk memenuhi kebutuhan hidup masyarakatnya. Indonesia adalah negara yang sangat kaya dengan berbagai hasil alam dan bahan pangan yang melimpah ruah, dari yang bersifat nabati hingga hewani termasuk ikan. Ikan sebagai salah satu bahan pangan asal hewan, mengandung berbagai macam zat gizi seperti protein, lemak, vitamin dan mineral. Beberapa tahun belakangan ini, masyarakat Indonesia mulai melirik ikan sebagai salah satu bahan pangan penting untuk mencukupi kebutuhan gizinya terutama protein, selain dari daging, telur dan susu. Kebutuhan masyarakat Indonesia terhadap ikan saat ini minimal 20 kg/jiwa/tahun (Jangkaru 2002) dan jumlah ini akan terus meningkat seiring peningkatan taraf hidup dan kesadaran masyarakat Indonesia akan pentingnya nilai gizi ikan.
Ikan sama seperi makhluk hidup lainnya, tidak pernah lepas dari ancaman berbagai jenis penyakit dan salah satu penyebab penyakit tersebut adalah parasit. Menurut Noble & Noble (1989), parasit adalah organisme yang hidupnya dapat menyesuaikan diri dengan inangnya namun merugikan bagi organisme yang ditempatinya. Berbagai jenis parasit telah diketahui, baik yang bersifat endoparasit maupun ektoparasit dan salah satu contoh dari parasit tersebut adalah cacing. Cacing sebagai salah satu parasit yang sering ditemukan pada ikan, memegang peranan besar bagi kesehatan dan kesejahteraan hewan dan manusia karena salah satu aspek penting dari parasitologi ikan ialah masalah higiene pangan. Faktanya, beberapa larva dan cacing dewasa golongan trematoda, nematoda dan cestoda dapat menimbulkan berbagai resiko kesehatan bagi sistem pencernaan manusia serta menghasilkan enzim yang akan merusak tekstur dan kualitas daging ikan (Buchmann & Bresciani 2001). Ikan kembung sebagai salah satu ikan air laut dari genus Decapterus yang umum dikonsumsi manusia, pada insangnya dapat ditemukan berbagai jenis cacing parasitik baik dalam bentuk larva maupun dewasa. Berbagai kerugian dapat ditimbulkan akibat kehadiran cacing parasitik ini
seperti kerugian ekonomi, kesehatan dan ekologi. Oleh karenanya diperlukan pemahaman dan pengendalian terhadap cacing parasitik dan penyakit yang ditimbulkannya terutama yang berasal dari ikan untuk dapat mengembangkan berbagai produk asal ikan terutama untuk konsumsi manusia (Yamaguti 1958). Pemahaman dan pengendalian terhadap cacing parasitik ini didahului oleh proses identifikasi. Identifikasi cacing parasitik dalam penelitian ini menggunakan pewarnaan khusus yang bersifat semi permanen dengan minyak cengkeh (Eugenol aromatika) dan kalium hidroksida (KOH)10% untuk nematoda, sedangakan untuk trematoda digunakan pewarnaan yang bersifat permanen menggunakan Semichon’s Acetocarmin (Neta 2006).
Tujuan Penelitian
Penelitian ini bertujuan untuk mengetahui adanya serta mengidentifikasi jenis cacing parasitik yang terdapat pada insang ikan kembung (Decapterus spp.).
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat memberikan informasi akan adanya cacing parasitik pada insang ikan kembung. Selain itu, penelitian ini juga diharapkan dapat memberikan manfaat akademik dan praktis. Dari sisi akademik sebagai temuan cacing parasitik pada insang ikan kembung khususnya yang berada di perairan Indonesia bagian barat. Dari sisi praktis, hasil penelitian ini diharapkan dapat menjadi informasi tambahan dalam daftar parasit ikan di perairan Indonesia sehingga dapat dijadikan acuan dalam program pengendalian penyakit zoonosis yang berasal dari ikan konsumsi.
TINJAUAN PUSTAKA
Ikan Kembung (Decapterus spp.)
Klasifikasi ikan kembung menurut Saanin (1968) adalah sebagai berikut: kingdom: Animalia; filum: Chordata; subfilum: Vertebrata; superkelas: Osteichtyes; kelas: Actinopterygii; subkelas: Neopterygii; infrakelas: Teleostei; superordo: Acanthopterygii; ordo: Perciformes; subordo: Percoidei; famili: Carangidae; genus: Decapterus; spesies: Decapterus spp. (Gambar 1)
Gambar 1. Ikan kembung (Decapterus spp.)(Anonim 2008)
Ikan kembung, memiliki nama lain Round Scad, Cigar Minnow, Hardtail, Cigarfish, Chuparaco, merupakan ikan laut genus Decapterus dari famili Carangidae (Allen 2000). Jenis ikan ini sangat diminati masyarakat Indonesia sebagai ikan konsumsi karena selain memiliki rasa yang enak, ikan ini juga memiliki kandungan gizi yang sangat tinggi terutama protein dan asam lemak tak jenuh yang dapat menurunkan kadar kolesterol jahat dalam tubuh, juga vitamin dan mineral. Ikan ini juga tergolong ikan yang sangat ekonomis sehingga dapat dijangkau oleh masyarakat Indonesia dari berbagai kalangan.
Ikan kembung mempunyai panjang tubuh yang tidak lebih dari 30 cm (rata-rata 15-20 cm) dengan berat terbesar yang pernah dilaporkan 300 gram (Vaniz et al. 1990). Morfologi ikan ini terdiri atas 9 spina dorsal, 3 spina anal, dan 24 tulang vertebrae. Tubuhnya langsing, mengkilat, berwarna hijau keabu-abuan pada bagian dorsal dan abu-abu putih di bagian bawahnya. Pada ikan dewasa terdapat satu sampai tiga bintik hitam kecil pada lengkung lateral tubuhnya. Moncongnya lebih panjang daripada diameter mata. Rahang atas dilengkapi
gigi-gigi kecil dan terletak anterior. Garis lateral melengkung halus di sebelah dorsal bawah dengan 30-34 skutum yang kuat. Ikan yang termasuk genus Decapterus
mudah dibedakan dengan 26 genus lainnya dalam famili Carangidae dengan adanya tanda khusus, yaitu sebuah finlet yang terdapat di belakang sirip punggung dan sirip dubur (Saanin 1968). Vaniz et al. (1990) menyatakan bahwa ikan ini sama sekali tidak beracun dan berbahaya baik melalui kontak secara langsung ataupun jika dimakan.
Ikan kembung memiliki kekeluargaan yang sama dengan ikan Selar (Selar crumenthalmops), Makarel (Decapterus macarellus) dan ikan ekor merah atau Redtail Scad (Decapterus kurruides). Ikan kembung sama seperti ikan tuna, tongkol, ataupun ikan ekor kuning, tergolong pelagis yaitu ikan yang biasa berenang-renang bebas dan memiliki kemampuan renang yang cepat serta jauh. Berdasarkan Moyle & Cech (1988), ikan pelagis umumnya berukuran kecil, bentuk mulut superior, kepala berbentuk pipih datar dengan mata lebar dan sirip punggung berada di bagian belakang badan. Morfologi ikan ini sesuai untuk menangkap plankton, insekta atau ikan-ikan kecil yang hidup di dekat permukaan air. Ikan kembung biasanya hidup di perairan laut terutama daerah karang, pada kedalaman 0-100 m dari permukaan air laut dengan iklim tropis maupun subtropis (Cuvier 1829). Makanan ikan ini adalah plankton invertebrata, terutama dari golongan copepoda namun juga memakan invertebrata dari golongan gastropoda, pteropoda dan ostrakoda.
Trematoda
Trematoda (cacing daun) atau yang sering disebut cacing pipih, merupakan kelas dari filum Plathyhelminthes dan seluruh anggotanya bersifat parasiter. Beberapa trematoda hidup pada permukaan tubuh inang definitifnya dan beberapa lainnya di dalam tubuh sebagai parasit internal (Noble & Noble 1989).
Morfologi Umum Trematoda
Trematoda tidak mempunyai rongga tubuh dan seluruh organ berada di dalam rongga parenkim. Tubuh cacing ini berbentuk seperti daun, simetris bilateral, pipih dorsoventral dan tidak bersegmen. Trematoda memiliki dua alat
penghisap, satu mengelilingi mulut atau yang sering disebut sebagai batil isap oral dan yang lain berada di dekat pertengahan tubuh atau ujung posterior yang disebut batil isap ventral atau asetabulum karena bentuknya mirip mangkuk cuka (Levine 1990). Dyková & Lom (2008) menyatakan ada beberapa spesies trematoda yang tidak memiliki salah satu atau bahkan kedua alat penghisap ini. Alat penghisap oral dan ventral merupakan karakter spesifik dari cacing yang berasal dari kelas trematoda (Barnes 1963)
Dinding luar tubuh cacing daun adalah tegumen yang kadang-kadang mengandung duri atau sisik (Levine 1990). Menurut Noble (1989), ciri khas cacing pipih terletak pada sistem protonefridial atau sistem ekskretorisnya. Sistem ekskretoris trematoda terdiri atas flame cells, dihubungkan oleh tubulus yang kemudian bersatu menjadi duktus yang lebih besar. Duktus ini kemudian bermuara secara bebas ke luar tubuh atau bergabung dahulu menjadi suatu kandung kencing yang bermuara pada atau dekat ujung posterior tubuh cacing. Trematoda memiliki mulut dan saluran pencernaan, namun tidak memliki anus. Lubang mulut bersambung dengan faring berotot yang panjangnya berbeda-beda tergantung spesies dan diteruskan dengan usus yang bercabang-cabang (Grabda 1991). Kecuali famili Schistosomatidae yang khusus ditemukan pada hewan darat, sistem reproduksi trematoda ialah hermafrodit (Soulsby 1982), namun sebuah kecenderungan pemisahan jenis kelamin juga ditemukan pada subkelas
Didymozoidae (Grabda 1991). Grabda (1991) membagi kelas trematoda menjadi menjadi tiga subkelas berdasarkan anatomi dan siklus hidupnya, yaitu: subkelas
Aspidogastrea, Digenea dan Didymozoida.
Digenea
Digeneasida atau trematoda digenetik merupakan cacing pipih parasitik yang paling umum ditemukan pada hewan dan menyebabkan infeksi asimptomatik pada ikan (Noga 1996). Cacing digenea pada umumnya bersifat endoparasit yang dapat ditemukan pada organ dalam ikan seperti usus, pembuluh darah atau terbungkus kiste di jaringan tubuh (Moller & Anders 1986), namun beberapa jenis digenea bersifat ektoparasit dan dapat ditemukan pada permukaan insang, operkulum dan rongga mulut. Pardede 2002 menyatakan bahwa investasi
cacing trematoda digenea pada insang akan mengakibatkan terjadinya pembengkakan dan kerusakan.
Jenis dan Klasifikasi Digenea
Klasifikasi trematoda digenea pada ikan dapat diuraikan sebagai berikut (Rudolphi 1808), kingdom: Animalia; filum: Platyhelminthes; kelas: Trematoda; subkelas: Digenea; ordo: Allocreadiinae; famili: Allocreadiidae; genus:
Helicometra ; spesies: Helicometra spp. (Gambar 2).
Gambar 2. Helicometra spp., suatu digenea opecoelid pada ikan (Anonim 2008)
Morfologi Digenea
Parasit ini ditandai dengan batil isap muskuler berbentuk mangkok, biasanya tanpa kait atau organ-organ tambahan lain untuk berpegangan, dengan lubang-lubang genital yang biasanya bermuara di permukaan ventral antara batil-batil isap serta sebuah lubang ekskretoris posterior (Noble & Noble 1989). Bentuk klasik trematoda digenea adalah seperti daun oval yang tebal. Meskipun bentuk khas cacing ini ditandai dengan adanya dua buah batil isap pada bagian mulut dan pada bagian ventral, namun ada beberapa diantaranya yang hanya memiliki sebuah batil isap dan ada pula yang sama sekali tidak memiliki. Monostoma adalah cacing daun dengan satu batil isap sedangkan distoma memiliki dua batil isap (Noble 1989). Menurut Grabda (1991), tubuh trematoda digenea ditutupi oleh suatu kantung “dermomuscular” dengan lapisan tegumen yang tipis namun elastis di permukaannya. Di bawah tegumen terdapat tiga lapisan otot: sirkular, diagonal dan longitudinal. Kontraksi dari ketiga otot inilah yang menyebabkan pergerakan tubuh cacing digenea.
Morfologi umum cacing digenea (Gambar 3) dapat diuraikan sebagai berikut: sistem saraf, terdiri dari ganglion kepala serta trunkus anterior dan posterior; organ sensorik yang sangat minimalis dan kadang-kadang terdapat bintik mata. Terkadang cacing digenea memiliki sistem limfatik namun tanpa sistem peredaran darah (Yamaguti 1958); sistem pencernaan, dimulai dari mulut yang berlanjut ke faring yang berotot. Faring membuka ke esofagus yang panjangnya berbeda-beda bergantung spesies. Esofagus terbagi menjadi dua sekum yang bercabang-cabang; sistem ekskresi, terdiri atas protonefridia dan kanalikuli yang terbagi menjadi dua kantong ekskretoris lateral yang simetris. Kantong ekskretoris trematoda digenea dapat berbentuk I-, Y- atau V- (Dawes 1956) dan menurut Grabda (1991) sistem ekskretoris ini merupakan salah satu kunci penting dalam identifikasi trematoda digenea. Sistem reproduksi digenea terbagi atas sistem reproduksi jantan dan betina. Sistem reproduksi jantan pada trematoda digenea berupa sepasang testis namun beberapa cacing memiliki testis tunggal atau banyak. Vas efferen menghubungkan testis dengan vas defferens yang kemudian bersambung dengan duktus ejakulatoris. Duktus ejakulatoris terbuka dekat dengan lubang uterus. Suatu bagian yang membesar di ujung distal sistem reproduksi jantan, memperlihatkan suatu alat yang berguna menyalurkan sperma ke alat kelamin betina cacing lain atau juga ke alat kelamin betina cacing yang sama. Bagian ini dinamakan sirus (analog dengan penis pada hewan tingkat tinggi). Noble & Noble (1989) menyatakan bahwa saluran Laurer mungkin merupakan duktus yang mereduksi sesuai dengan kanal vagina pada cacing pita dan apabila kanal tersebut berfungsi sebagai organ penyimpan sperma, maka akan terdapat sebuah alat penerima yang disebut reseptakulum seminis. Sistem reproduksi betina terdiri atas ovarium tunggal, oviduk sebagai penghubung antara ovarium dan ootip dimana fertilisasi akan berlangsung. Ootip akan terhubung dengan kelenjar vitellin dimana sekresi dari kelenjar ini berperan dalam pembentukan kerabang telur dan penyediaan bahan makanan bagi embrio. Telur yang sudah dibuahi di ootip, akan melewati uterus yang akan menyalurkannya keluar melalui lubang genital yang terletak di anterior tubuh (Grabda 1991). Ootip dikelilingi masa sel-sel kelenjar kecil, disebut kelenjar Mehlis (Noble & Noble 1989).
Gambar 3. Morfologi umum trematoda digenea (Anonim 2008)
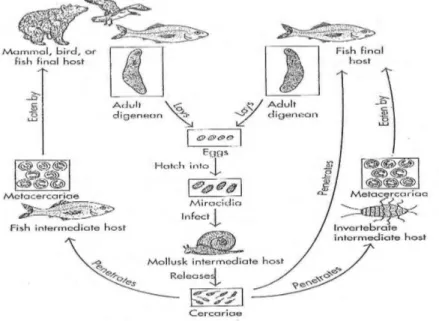
Siklus Hidup Digenea
Trematoda digenetik mempunyai siklus hidup yang kompleks (Gambar 4), melalui beberapa bentuk dan membutuhkan satu atau lebih induk semang antara.
Menurut Barnes (1963), berbeda dari trematoda monogenea, siklus hidup trematoda digenea dapat melibatkan dua sampai empat induk semang dimana induk semang bagi cacing dewasa disebut sebagai induk semang akhir (final host) atau yang lebih dikenal sebagai induk semang definitif dan satu sampai tiga induk semang bagi berbagai tahap perkembangan larva disebut sebagai induk semang antara (intermediate host). Digenea dewasa memproduksi telur ber”operkulum” berbentuk oval (Gambar 5A) dalam jumlah besar yang kemudian dilepaskan oleh induk semang akhir (final host). Telur kemudian menetas menjadi mirasidium bersilia yang akan berenang bebas di lingkungan perairan dan kemudian menginfeksi molluska (terutama siput) sebagai induk semang antara (Noga 1996). Mirasidium (Gambar 5B) dilengkapi dengan kelenjar penembus, sistem ekskretoris, sel-sel germinal, dan beberapa diantaranya, memiliki bintik mata (Noble & Noble 1989). Dengan tarikan kemotaktik atau dengan cara lain, mirasidium ini mendapatkan siput, tiram, kopepoda atau molluska lain. Walaupun demikian banyak mirasidium mendapatkan induk semang antaranya secara kebetulan, meskipun tropisme-tropisme, misalnya fototropisme, mungkin mengarahkan mereka ke tempat berkumpulnya induk semang antara (Noble & Noble 1989). Di dalam tubuh siput, mirasidium berkembang menjadi sebuah kantung yang dinamakan sporokista (Gambar 5C). Sel-sel germinal yang melapisi dinding bagian dalam sporokista berkembang menjadi redia (Gambar 5D) yang masak dan kemudian akan keluar melalui lubang peranakan atau dengan pecahnya sporokista. Menurut Noble (1989), sporokista atau redia dapat dianggap sebagai kantung atau kista germinal. Tiap-tiap sel germinal di dalam suatu redia berkembang menjadi satu serkaria (Gambar 5E) yang masak dan kemudian akan keluar melalui lubang peranakan atau dengan pecahnya redia. Serkaria akan berpenetrasi ke dalam tubuh ikan sebagai inang antara kedua (Noga 1996). Berdasarkan kemampuannya untuk mencari dan menginfeksi induk semangnya, maka stadium larva serkaria dapat dikatakan sebagai stadium infektif dari siklus hidup cacing trematoda digenea. Setelah mencapai target jaringan pada tubuh ikan, serkaria berdiferensiasi menjadi metaserkaria (Gambar 5F). Ketika ikan yang mengandung metaserkaria dimakan oleh induk semang definitifnya, metaserkaria segera berdiferensiasi menjadi dewasa (Noga 1996).
Nematoda
Nematoda sering disebut cacing ”gilig”, merupakan kelas tersendiri dari filum Nemathelmintes. Nematoda merupakan parasit yang sering dijumpai pada ikan (Hirschhorn 1989), dimana ikan dapat bertindak sebagai induk semang antara maupun induk semang definitif.
Noga (1996) menyatakan bahwa ikan laut biasanya terinfeksi oleh nematoda yang berasal dari golongan Ascaridoidoiea (Contracecum, Pseudoterranova, Anisakis, Cotracaecum), Camallanoidea (Camallanus, Culcullanus), Dracunculoidea (Philonema, Philometra), dan Spiruroidea (Metabronema, Ascarophis). Sebagian besar camallanoids, dracunculoids, dan spiruroids memiliki dua induk semang dalam siklus hidupnya dimana ikan bertindak sebagai induk semang definitif. Spirocamallanus dapat bersifat patogen bagi ikan laut tropis (Rychlinski & Deardorff 1982). Truttaedactitras, Philonema
dan Philometra umum menyerang populasi ikan di alam dan menimbulkan berbagai kerusakan pada organ tubuh ikan (Dick et al. 1987). Parasit dari kelas nematoda juga dapat bersifat zoonosis atau menginfeksi manusia yang mengkonsumsinya, misalnya Anisakis yang menyebabkan anisakiasis (Pardede 2000).
Morfologi Umum Nematoda
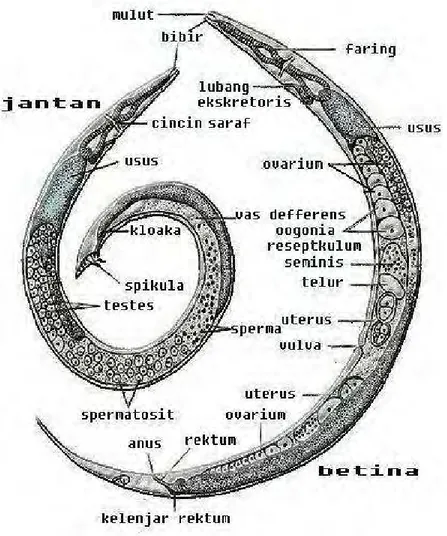
Menurut Buchmann & Bresciani (2001), cacing ini berbentuk panjang, ramping, silindris, tidak bersegmen, dengan kedua ujung meruncing, mempunyai mulut serta anus (saluran pencernaan yang lengkap) serta memiliki rongga tubuh semu yang disebut ”pseudoselom” (Gambar 6) (Noble 1989). Tubuhnya tertutup oleh suatu kantung dermomuskular yang terdiri dari tiga lapisan: kutikula, hipodermis dan otot (Grabda 1991). Penutup luar tubuh (external covering) nematoda berupa suatu lapisan non-selular yang disebut kutikula. Lapisan ini sering disebut sebagai lapisan kitin namun memiliki struktur kimia berbeda dari lapisan kitin eksoskleton arthropoda (Whitlock 1960).
Menurut Grabda (1991), sistem pencernaan nematoda dimulai dengan mulut yang dikelilingi oleh bibir (labia), biasanya terdapat tiga buah bibir (satu terletak dorsal dan dua pada ventro-lateral), yang dapat ditemukan pada spesies
Anisakis spp. Pada spesies lain (Contracaecum spp.), terdapat struktur di antara bibir yang disebut interlabia dan ada pula beberapa spesies yang sama sekali tidak memiliki bibir (Philometra spp.). Pada Camallanus spp. mulut terbuka masuk ke dalam kapsul bukal dengan kutikula yang tebal. Mulut berlanjut ke esofagus yang memiliki dinding otot tebal. Pada beberapa spesies, esofagus dan usus dipisahkan oleh kelenjar ventrikulus. Pola hubungan antara esofagus-ventrikulus inilah yang menjadi kunci karakteristik taksonomi untuk mengidentifikasi spesies dari famili Anisakidae. Usus merupakan sebuah tabung lurus yang dibentuk oleh selapis sel-sel usus dan jumlahnya sudah tetap pada masing-masing spesies. Usus berlanjut ke rektum dan akan berakhir pada anus. Pada cacing jantan, vas defferens juga akan bermuara di rektum yang pada beberapa kasus dapat bertindak sebagai kloaka (Grabda 1991).
Menurut Kusumamihardja (1989), nematoda tidak mempunyai sistem peredaran darah. Rongga tubuhnya mengandung haemoglobin, glukosa, protein, garam dan vitamin, yang kesemuanya memenuhi fungsi darah. Sistem sarafnya terdiri atas cincin saraf sekeliling esofagus dan enam saraf posterior yang keluar dari cincin tersebut. Alat indranya terletak pada bibir, daerah serviks, sekitar anus dan sekitar alat kelamin. Sistem ekskresi terdiri atas dua saluran lateral. Kedua saluran tersebut dihubungkan dengan sebuah saluran di daerah esofagus dan saluran yang menghubungkannya dengan lubang pembuangan di daerah ventral daerah esofagus. Bentuk spesifik dari sistem ekskresi ini berbeda-beda pada setiap grup nematoda; dapat berupa bentuk ”H” atau ”U”. Lokasi lubang ekskresi pada nematoda sangat penting dalam sistem taksonomi (Grabda 1991)
Nematoda tidak memiliki sistem pernafasan dan alat kelamin terpisah antara cacing jantan dan betina (Buchmann & Bresciani 2001). Cheng (1973) menyatakan bahwa secara umum nematoda jantan dapat dibedakan dari nematoda betina dari ukuran dan bentuk tubuhnya dimana nematoda jantan memiliki ukuran yang lebih kecil, ujung posterior yang melingkar dan adanya bursa serta beberapa struktur reproduksi tambahan lainnya yang tidak dimiliki oleh nematoda betina.
Sistem reproduksi jantan terdiri atas testis tunggal, satu atau dua buah spikula sebagai alat kopulasi dan kantung gubernakulum pada beberapa spesies (Cheng 1973). Testes akan berlanjut menjadi vas defferens yang merupakan ujung
yang kuat dan berotot serta berfungsi sebagai duktus ejakulatoris. Vas defferens akan bermuara di rektum yang berperan sebagai kloaka (Grabda 1991). Rektum yang dilapisi oleh kutikula akan berakhir pada anus. Menurut Cheng (1973), bentuk, ukuran dan jumlah dari spikula kopulatoris dan struktur alat bantu kopulasi lainnya, merupakan unsur yang sangat penting dalam menentukan taksonomi cacing nematoda.
Sistem reproduksi betina terdiri atas dua buah ovarium yang terletak di anterior dan posterior tubuh. Beberapa spesies nematoda hanya memiliki sebuah ovarium yang dapat terletak di bagian anterior atau posterior, oviduk yang berlanjut pada ujung proksimal ovarium, dua buah uterus yang masing-masing berhubungan langsung dengan kedua ovarium serta vagina (Cheng 1973).
Siklus Hidup Nematoda
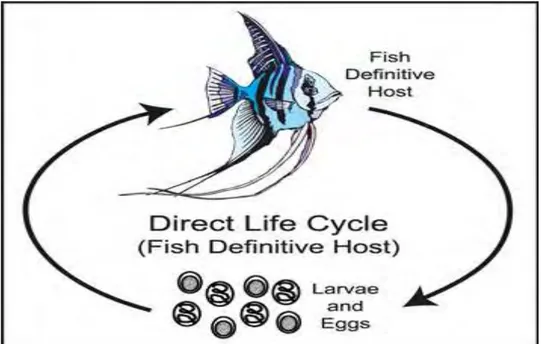
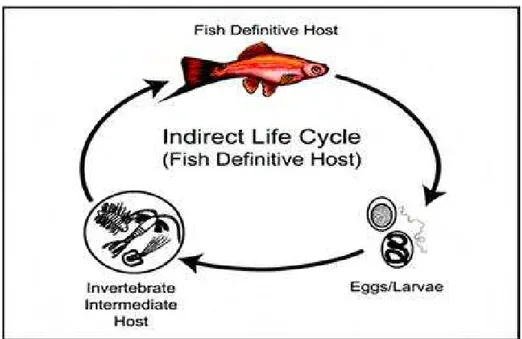
Nematoda memiliki siklus hidup yang rumit, berbeda-beda tergantung pada spesies (Yanong 2008) dan melibatkan inang antara invertebrata. Organisme yang mengandung stadium dewasa kelamin dari cacing nematoda ini dikenal sebagai induk semang definitif, sedangkan organisme yang hanya dibutuhkan untuk melengkapi siklus hidup cacing ini tetapi tidak mengandung stadium dewasa kelamin cacing dikenal sebagai induk semang antara (Yanong 2008). Noga (1996) menyatakan bahwa ikan merupakan induk semang antara sekaligus induk semang definitif bagi perkembangan cacing nematoda. Secara umum, di dalam tubuh ikan, cacing nematoda memiliki lima stadia dalam siklus hidupnya yang dipisahkan oleh empat kali pergantian kulit (moulting) (Buchmann & Bresciani 2001). Yanong (2008) membagi siklus hidup nematoda menjadi dua kategori utama, yaitu siklus hidup langsung dan tidak langsung. Siklus hidup langsung (Gambar 7), dimana ikan bertindak sebagai induk semang definitif bagi nematoda dan tidak diperlukan induk semang antara sehingga infeksi dapat langsung disebarkan secara langsung dari satu ikan ke ikan lain melalui telur atau larva infektif yang termakan.
Jika nematoda memiliki siklus hidup tidak langsung (Gambar 8), telur atau larva akan dikeluarkan ke dalam air dan selama proses perkembangannya, larva yang belum dewasa ini setidaknya akan melewati dua organisme yang berbeda yang salah satunya adalah ikan.
Gambar 8. Siklus hidup tidak langsung cacing nematoda (Yanong 2008)
Menurut Yanong (2008), siklus hidup tidak langsung nematoda dapat dibagi lagi menjadi dua subkategori dimana ikan berperan sebagai induk semang definitif atau sebagai induk semang antara (Gambar 9). Siklus hidup tidak langsung dimana ikan merupakan induk semang definitif, apabila cacing nematoda memasuki organisme lain diluar ikan yang pada umumnya adalah invertebrata air seperti golongan kopepoda atau larva insekta yang di dalamnya akan terjadi tahap perkembangan sebelum cacing dimakan oleh ikan. Ketika dimakan oleh ikan yang tepat, cacing akan mencapai kematangan kelamin dan akan bereproduksi di dalam ikan. Siklus hidup tidak langsung dimana ikan merupakan induk semang antara terjadi apabila nematoda hanya menggunakan ikan sebagai perantara sebelum memasuki induk semang definitifnya yaitu organisme lain pemakan ikan seperti ikan yang lebih besar, burung atau mamalia air lainnya.
Gambar 9. Siklus hidup tidak langsung dimana ikan sebagai induk semang antara. Telur atau larva nematoda (a) memasuki induk semang antara invertebrata (b) atau induk semang antara ikan (c) sebelum dimakan oleh atau memasuki tubuh induk semang akhir (definitif), dapat berupa ikan, burung atau mamalia pemakan ikan (Yanong 2008)
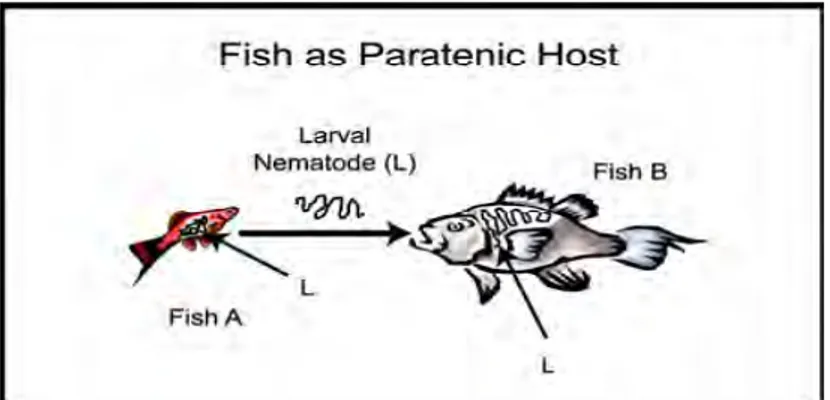
Beberapa cacing nematoda memiliki kemampuan bertahan hidup pada organisme alternatif atau yang lebih dikenal sebagai induk semang paratenik (Gambar 10) (Yanong 2008). Induk semang ini tidak dibutuhkan untuk melengkapi siklus hidup cacing tetapi dapat mengandung stadium infektif nematoda dan menjadi sumber infeksi bagi organisme lain. Induk semang paratenik dapat berupa ikan, cacing atau organisme air lainnya yang memakan telur atau larva infektif nematoda.
Yanong (2008) menyatakan bahwa nematoda dapat menginfeksi berbagai spesies ikan baik ikan air tawar maupun ikan laut dimana dalam jumlah kecil sering ditemukan pada ikan yang sehat dan tidak menunjukkan gejala penyakit yang khas namun ikan yang terinfeksi akan mengalami penurunan produktifitas. Dalam lingkungan perairan, ikan dapat terinfeksi nematoda jika memakan ”makanan” hidup yang mengandung larva infektif nematoda atau jika ikan tersebut berperan sebagai inang antara atau yang membawa larva infektif nematoda (Gambar 11) yang pada akhirnya nematoda dapat ditularkan secara langsung dari satu ikan ke ikan yang lain (Yanong 2008).
Gambar 11. Larva nematoda Camallanus spp. di lingkungan perairan (Yanong 2008)
Nematoda dewasa biasanya ditemukan dalam saluran pencernaan ikan, meskipun demikian, bergantung pada spesies nematoda dan spesies ikan yang diinfeksinya, stadium dewasa maupun stadium lainnya dari cacing nematoda dapat ditemukan hampir di seluruh bagian dari tubuh ikan termasuk pada organ dalam, gelembung renang, kulit, otot, maupun insang (Yanong 2008).
BAHAN DAN METODE PENELITIAN
Waktu dan Tempat Penelitian
Isolasi cacing parasitik dilakukan pada bulan Juli 2007 dari insang ikan kembung yang diambil dari pasar ikan Muara Angke (Jakarta Utara). Pewarnaan serta identifikasi dilakukan mulai dari bulan Desember 2007 hingga Juli 2008, bertempat di Laboratorium Helmintologi Bagian Parasitologi dan Entomologi Kesehatan, Departemen Ilmu Penyakit Hewan dan Kesehatan Masyarakat Veteriner Fakultas Kedokteran Institut Pertanian Bogor.
Bahan dan alat Penelitian
Bahan dan alat yang digunakan dalam penelitian ini adalah sampel ikan kembung, NaCl fisiologis 0,85%, alkohol bertingkat, alkohol absolut, ethanol 70%, kalium hidroksida 10%, minyak cengkeh, pewarna Semichon’s Acetocarmine, entelan, xylol, seperangkat alat bedah (dissecting kit), timbangan kue, cawan petri, pinset, kait, pipet tetes, gunting, botol kaca, spidol, label nama, gelas objek dan kaca penutup, kaca pembesar, mikroskop cahaya, mikroskop stereo, dan video mikrometer.
Metode Penelitian 1. Pengambilan Sampel
Sampel ikan kembung diambil dari pasar ikan Muara Angke, Jakarta Utara sebanyak 18 ekor, dengan berat badan rata-rata 200 gram.
2. Teknik Parasitologi
Ikan kembung yang sudah dalam keadaan mati (kira-kira 24 jam), diambil insangnya lalu dipindahkan ke dalam cawan petri yang berisi larutan NaCl fisiologis. Dengan menggunakan mikroskop stereo, cacing dikoleksi dengan menyisir setiap filamen insang. Cacing yang ditemukan direlaksasi dalam cawan petri berisi larutan NaCl fisiologis dan selanjutnya difiksasi dalam etanol 70% sebelum diwarnai.
3. Pewarnaan
Ada dua jenis teknik pewarnaan yang digunakan dalam penelitian ini yaitu pewarnaan permanen untuk trematoda dan pewarnaan semi permanen untuk nematoda. Kedua jenis pewarnaan melewati beberapa tahap sebagai berikut:
Pewarnaan Permanen
Pewarnaan permanen atau pewarnaan Semichon’s Acetocarmine digunakan untuk mengidentifikasi cacing pipih (golongan trematoda). Pewarnaan ini dilakukan dengan merendam spesimen dalam larutan Semichon’s Acetocarmine selama 15-20 menit sampai warna terserap dan spesimen menjadi berwarna merah cerah lalu spesimen dibilas etanol 70% dan kemudian direndam dalam larutan asam alkohol (99 bagian etanol 70% dicampur dengan 1 bagian HCl). Setelah itu dilakukan dehidratasi spesimen menggunakan alkohol bertingkat (70%, 85%, 95%, 100%) dengan merendamnya selama 5 menit pada tiap-tiap konsentrasi alkohol, lalu dilanjutkan perendaman dengan xylol sampai spesimen terlihat tembus pandang. Terakhir, spesimen di-mounting dengan entelan sebagai media fiksasi (Soulsby 1982).
Pewarnaan Semi Permanen
Pewarnaan semi permanen menggunakan KOH dan minyak cengkeh digunakan untuk nematoda yang memiliki struktur berbeda dari cacing pipih, dengan prosedur sebagai berikut (Neta & Khairunnisa 2007): penipisan atau penghilangan lapisan kutikula cacing dilakukan dengan merendam spesimen dalam KOH 10% selama 1-3 menit sampai kutikula / lapisan luar spesimen terlihat tembus pandang, kemudian cacing tersebut dipindahkan ke dalam minyak cengkeh selama kurang-lebih 30 detik hingga 1 menit sampai organ-organ tubuh terlihat berwarna lebih jelas. Selanjutnya cacing dimasukkan ke dalam alkohol bertingkat (70%, 85%, 95%) masing-masing selama 15 sampai 30 detik untuk dehidrasi dan terakhir cacing di-mounting dengan entelan sebagai media fiksasi.
4. Pengamatan dan Pengukuran
Pengamatan dilakukan dengan menggunakan mikroskop cahaya terhadap organ dalam cacing sedangkan pengukuran dilakukan dengan menggunakan video mikrometer untuk mengukur panjang dan lebar tubuh cacing. Pengukuran ini bertujuan untuk memudahkan tahap identifikasi.
5. Identifikasi cacing parasitik
Identifikasi cacing parasitik golongan trematoda mengacu pada Dawes (1956), Yamaguti (1958), Winterbourn (1973), dan Noble & Noble (1989), sedangkan identifikasi cacing parasitik golongan nematoda mengacu pada Hirschmann (1960), Cheng (1973), (Meyers 1975), Noble & Noble (1989), Grabda (1991), Buchmann & Bresciani (2001), Dixon (2006), Nuchjangreed et al. (2006), dan Yanong (2008).
6. Analisis statistik
Analisis statistik yang digunakan untuk pengolahan data penelitian ini adalah analisis deskriptif.
HASIL DAN PEMBAHASAN
Jenis Cacing Parasitik yang Ditemukan
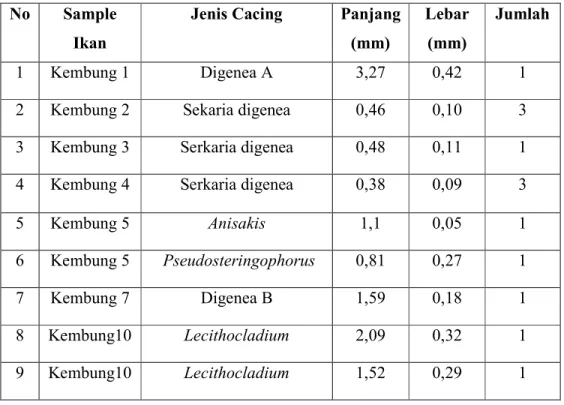
Hasil identifikasi cacing parasitik yang ditemukan pada insang ikan kembung berasal dari filum Plathyhelminthes, subkelas Digenea, dan dari filum Nemathelminthes, kelas Nematoda. Dari 18 sampel ikan yang dipakai, tujuh ikan positif terinfeksi cacing parasitik. Jumlah cacing yang ditemukan berjumlah 13 dan terdapat 6 jenis cacing berbeda dengan perincian sebagai berikut: 1 cacing termasuk dalam filum Nemathelminthes, kelas Nematoda, genus Anisakis; 12 cacing berasal dari kelas digenea: satu cacing genus Pseudosteringophorus, dua cacing genus Lecithocladium, tujuh cacing masih dalam bentuk larva serkaria dan dua cacing hanya dapat teridentifikasi sebagai digenea dewasa. Ukuran, jenis, dan jumlah cacing parasitik yang ditemukan, disajikan dalam tabel 1.
Tabel 1. Jenis, ukuran dan jumlah cacing parasitik yang ditemukan No Sample
Ikan
Jenis Cacing Panjang (mm)
Lebar (mm)
Jumlah
1 Kembung 1 Digenea A 3,27 0,42 1
2 Kembung 2 Sekaria digenea 0,46 0,10 3
3 Kembung 3 Serkaria digenea 0,48 0,11 1
4 Kembung 4 Serkaria digenea 0,38 0,09 3
5 Kembung 5 Anisakis 1,1 0,05 1
6 Kembung 5 Pseudosteringophorus 0,81 0,27 1
7 Kembung 7 Digenea B 1,59 0,18 1
8 Kembung10 Lecithocladium 2,09 0,32 1 9 Kembung10 Lecithocladium 1,52 0,29 1
Digenea
Identifikasi terhadap cacing trematoda subkelas digenea dilakukan dengan melihat ukuran tubuh, karakteristik telur dan ciri morfologisnya. Sebagian besar trematoda dewasa memiliki ukuran tubuh yang berkisar antara 1-30 mm (Lom & Dyková 2008). Ciri utama cacing daun ini adalah adanya dua buah batil isap yang terletak di sekitar ujung anterior mulut (batil isap oral) dan di bagian ventral (batil isap ventral atau asetabulum). Batil isap pada cacing trematoda digenea biasanya berotot dan tidak dilengkapi kait-kait atau pencapit seperti yang dimiliki oleh cacing monogenea. Letak batil isap ventral atau asetabulum biasanya pada pertengahan tubuh dan pada umumnya berukuran lebih besar dari batil isap oral (Moller & Andres 1986).
Dari hasil pemeriksaan insang ikan kembung 1, 5, 7 dan 10, teridentifikasi lima ekor cacing dewasa yang berasal dari kelas Trematoda, subkelas Digenea dan dari hasil pemerikasaan insang ikan kembung 2,3 dan 4, teridentifikasi enam ekor digenea yang masih berada dalam stadium larva serkaria.
Genus Pseudosteringophorus
Salah satu cacing parasitik yang ditemukan pada insang kembung 5, teridentifikasi dalam filum Platyhelminthes, kelas Digenea, ordo Prosostomata, famili Fellodistomatidae, subfamili Fellodistomatinae, genus
Pseudosteringophorus (Yamaguti 1958). Perbandingan morfometrik dan ciri morfologis genus Pseudosteringophorus yang ditemukan selama penelitian dengan pustaka acuan disajikan dalam tabel 2.
Tabel 2. Perbandingan morfometrik dan ciri morfologis genus
Pseudosteringophorus yang ditemukan dengan pustaka acuan Identifikasi morfometrik (famili
Fellostomatidae) Identifikasi ciri morfologis Kriteria (mm) Penelitian Literatur
(Dawes 1956) Panjang tubuh Lebar tubuh Diameter batil isap oral Diameter batil isap ventral Panjang telur Lebar telur 0,81 0,27 0,17 0,33 0.031 0.014 0,8-3,5 0,3-0,5 0,2 0,35 0,02-0,05 0,014-0,017
Tubuh kecil; fusiformis; diastoma, batil isap ventral (asetabulum) lebih besar daripada batil isap oral, asetabulum terletak pada pertengahan tubuh, batil isap oral pada ujung terminal; faring kecil; testes berjumlah 2 buah, simetris, bersebelahan dan terletak di belakang asetabulum; kantung sirus ovoid dan terletak di depan asetabulum; ovarium di depan testes; reseptakulum seminis tidak ada; uterus memenuhi bagian belakang tubuh penuh berisi telur (dewasa); telur berjumlah sangat banyak, bergerombol, berbentuk oval, tidak berfilamen dengan kulit yang cukup tebal; vitellaria berkembang baik, folikular, terletak lateral pada sepertiga pertengahan tubuh; kantung ekskretoris berbentuk Y-. (Yamaguti 1958); tegumen tipis (Dawes 1956)
.
Beberapa organ yang dapat terlihat pada cacing yang ditemukan pada insang kembung 5 seperti batil isap oral, faring, batil isap ventral (asetabulum), kantung sirus, vitelaria (kelenjar vitellin), uterus, telur, testes dan lubang ekskresi (Gambar 12), menjadi kunci utama dalam identifikasi cacing digenea ini.
a b
Gambar 12. Genus Pseudosteringophorus (a) Yamaguti (1958), (b) hasil penelitian (perbesaran 4X)
Gambar 13. Struktur batil isap oral dan ventral (asetabulum) genus
Pseudosteringophorus yang ditemukan pada insang kembung 5 (perbesaran 40X)
Cacing ini dimasukkan dalam subordo Prosostomata karena letak mulut dengan batil isap oralnya yang dekat dengan ujung anterior tubuh (Dawes 1956). Kunci identifikasi famili Fellodistomatidae berdasarkan bentuk tubuhnya yang pipih, rata dan oval memanjang, tegumen tipis, tidak adanya prefaring namun adanya faring yang kecil. Famili Fellodistomatidae memiliki esofagus yang pendek atau panjang tergantung spesies namun pada preparat esofagus tidak dapat terlihat kemungkinan disebabkan oleh pewarnaan yang kurang sempurna. Cacing ini juga dicirikan dengan adanya batil isap oral dan ventral (asetabulum) (Gambar 13), kantung sirus yang pendek dan tebal, dua buah testes yang membulat dan simetris dimana testes yang satu berada di sebelah testes yang lain, ovarium yang terletak di depan testes namun tidak tampak pada preparat, adanya uterus yang meluas memenuhi bagian posterior tubuh dan berisi telur dalam jumlah besar, vitelaria yang berkembang baik dan saluran ekskretoris yang berbentuk V- atau Y- (Dawes 1956). Lubang genital dapat berada median, submedian atau lateral tubuh di depan asetabulum serta vitelaria yang dibatasi perluasannya dan terbagi ke dalam bentuk folikular, tubulus atau seperti bentuk cabang yang mengelompok (Yamaguti 1958). Kunci identifikasi genus Pseudosteringophorus dapat dilihat pada tabel 2.
Cacing digenea dewasa umum ditemukan pada saluran pencernaan ikan yang diinfeksinya, namun cacing ini juga dapat ditemukan di rongga mulut, paru-paru atau beberapa organ dalam lainnya karena sifatnya yang endoparasit pada tubuh induk semangnya (Williams & Williams 1996). Pseudosteringophorus
merupakan endoparasit pada ikan air laut dan habitat alaminya adalah saluran pencernaan khususnya lambung dan usus ikan yang diinfeksinya (Yamaguti 1958), namun dalam penelitian ini Pseudosteringophorus berhasil ditemukan pada insang yang bukan merupakan habitat alaminya. Hal ini dapat disebabkan oleh beberapa kemungkinan seperti kontaminasi di laboratorium dan kontaminasi di alam oleh cacing ini yang bermigrasi dari saluran pencernaan ke lingkungan perairan lalu menempel ke insang ikan atau karena migrasi cacing ini dari saluran cerna (terutama lambung) langsung ke insang. Migrasi cacing dapat terjadi premortem akibat populasi cacing sudah terlalu padat dalam saluran cerna ikan atau postmortem akibat ikan sebagai induk semangnya mati sehingga cacing merasa tidak nyaman lagi dan kemudian bermigrasi ke lingkungan.
Satyu Yamaguti pernah menemukan cacing famili Fellostomatidae spesies
Symmetrovesicula chaetodontis dalam bentuk spesimen gravid pada usus ikan
Chaetodon sp. di Makassar, Indonesia pada tahun 1953 (Yamaguti 1958). Yamaguti (1958) juga menyebutkan, spesies Pseudosteringophorus hoplognathi
pernah ditemukan di Jepang dalam usus ikan Hoplognatus punctata, tahun 1940.
Genus Lecithocladium
Digenea lain ditemukan pada insang kembung 10, berjumlah dua buah dan teridentifikasi dalam filum Platyhelminthes, kelas Digenea, ordo Prosostomata, famili Hemiuridae, subfamili Dinurinae, genus Lecithocladium (Yamaguti 1958). Cacing ini termasuk dalam ordo protostomata karena letak mulut beserta batil isap oralnya yang dekat dengan ujung anterior tubuh (Dawes 1956).
Menurut Yamaguti (1958), identifikasi famili Hemiuridae terutama dilakukan berdasarkan bentuk tubuhnya yang silindris memanjang dan adanya dua buah batil isap yang berukuran kecil hingga sedang pada bagian oral dan ventral tubuhnya (Gambar 15). Letak batil isap oral dan ventral (asetabulum) yang berdekatan merupakan karakteristik yang khas dari famili Hemiuridae. Beberapa
jenis cacing famili ini memiliki bagian tubuh yang menyerupai ekor. Tegumennya tipis atau seringkali tampak sedikit beranulus, bertingkat atau dengan tepi bergerigi. Batil isap oral, faring dan asetabulum berkembang baik dengan asetabulum biasanya terletak di dekat batil isap oral (dekat ujung anterior tubuh). Esofagus pendek, testis berjumlah dua buah dan terletak diagonal atau simetris. Cacing ini memiliki seminal vesikel dan duktus hermafrodit, vitelaria kompak, berlobus atau berbentuk tubulus, uterus berisi telur dalam jumlah besar, dan saluran ekskretoris berbentuk Y-. Telur cacing ini memiliki karakteristik telur digenea, yaitu berbentuk bulat oval, tanpa dilengkapi filamen atau alat gerak lain seperti telur cacing monogenea dan bergerombol dalam jumlah banyak. Yamaguti (1958) mengatakan bahwa cacing famili Hemiuridae merupakan parasit yang memiliki habitat utama esofagus dan lambung, kadang-kadang ditemukan di usus atau gelembung renang dan dapat pula di luar saluran pencernaan ikan.
Berdasarkan Yamaguti (1958), subfamili Dinurinae teridentifkasi oleh bentuk tubuh yang memanjang, testes terletak di belakang asetabulum, ovarium tidak berlobus dan terletak di belakang testes, vitelaria berlobus panjang, sempit dan teringkali tampak berbelit-belit. Dawes (1956) mengatakan bahwa vitelaria pada cacing subfamili Dinurinae tumbuh memanjang dan berbentuk tubulus. Cacing ini juga memiliki uterus yang meluas di bagian posterior tubuhnya ke arah vitelaria. Cacing ini dimasukkan dalam genus Lecithocladium karena tubuhnya yang langsing memanjang dan berukuran kecil hingga sedang. Genus
Lecithocladium memiliki batil isap oral yang bentuknya menyerupai mangkok dengan asetabulum yang mencolok dan terletak di ujung anterior tubuh. Testis dua buah berdampingan dan terletak saling diagonal, biasanya pada bagian anterior hingga petengahan tubuh. Vesikula seminalis sederhana dan pada umumnya dilengkapi dengan dinding otot yang tebal serta terletak anterodorsal dari testes. Ovarium terletak di dekat pertengahan tubuh. Vitelaria terdiri atas tujuh buah tubulus panjang dan uterus yang meluas hingga bagian ekor tubuh (Yamaguti 1958). Dawes (1956) menyebutkan bahwa cacing yang termasuk dalam genus
Lecithocladium memiliki tegumen dengan karakteristik berkerut-kerut di daerah anterior tubuhnya kecuali untuk spesimen yang memiliki panjang tubuh 1-1,5 mm. Menurut Yamaguti (2007), cacing dari genus Lechitocladium memiliki telur
yang berbentuk oval, kadang seperti ginjal (reniform), berembrio dan berukuran 0,016-0,021 x 0,009-0,012 mm. Dua buah spesimen digenea yang ditemukan pada insang kembung 10 memiliki karakteristik yang menyerupai karakteristik yang dimiliki oleh genus Lecithocladium dengan ukuran tubuh cacing pertama 2,09 x 0,32 mm serta cacing kedua 1,52 x 0,29 mm. Selain itu, kedua cacing terlihat memiliki telur yang berbentuk oval, agak memanjang seperti ginjal atau kacang merah, berukuran panjang antara 0,017 hingga 0,019 mm dan lebar antara 0,008 hingga 0,009 mm, sehingga dapat disimpulkan bahwa kedua cacing yang ditemukan pada insang kembung 10 termasuk genus Lecithocladium.
Lecithocladium scombri (Gambar 14) pernah ditemukan di Makassar, Indonesia pada lambung ikan Scomber kanagunta dan Scomber microlepidotus
oleh Satyu Yamaguti pada tahun 1953, memiliki ukuran panjang 1,9-3,2 mm dan lebar 0,3-0,48 mm (Yamaguti 2007). Selain itu, Manter (1954) menyebutkan pernah ditemukannya spesies lain dari genus Lecithocladium yaitu Lecithocladium magnacetabulum yang memiliki ukuran tubuh 1,4-1,960 x 0,49-1,960 mm,
Lecithocladium excisum yang memiliki panjang 1,655 mm dan Lecithocladium seriolellae yang berukuran 1,344-1,736 x 0,339-0,547 mm di wilayah perairan New Zealand. Cacing dari genus Lecitholadium sangat umum menyerang berbagai jenis ikan laut seperti spesies Lecitholadium excisum yang pertama kali ditemukan oleh Rudolphi pada tahun 1819, memiliki sinonim Distoma excisum. Cacing ini memiliki inang antara utama ikan dari genus Decapterus seperti makarel (Decapterus macarelus) dengan habitat alaminya saluran pencernaan terutama lambung ikan yang diserangnya (Dawes 1956). Meskipun induk semang utama cacing ini adalah ikan makarel tetapi tidak menutup kemungkinan baginya untuk menginfeksi jenis ikan laut lainnya karena sifatnya yang tidak host spesific. Cacing dewasa dari genus Lecithocladium bersifat endoparasit dan umum ditemukan pada saluran pencernaan ikan terutama usus dan lambung, namun cacing ini dapat juga ditemukan pada lokasi lain pada tubuh ikan seperti insang akibat migrasi cacing ke lingkungan dan kemudian masuk serta menempel pada lamela insang saat ikan bernafas dan membuka operkulumnya. Selain itu, migrasi juga dapat terjadi postmortem sama seperti cacing digenea dewasa lainnya atau karena adanya kontaminasi di laboratorium
Gambar 14. (a) Lecithocladium scombri, (b) Lecithocladium megalapsis (Yamaguti 2007)
Gambar 15. Genus Lechitocladium dari insang kembung 10 (perbesaran 4X)
Gambar 16. Struktur vitelaria dan ovarium Lecithocladium 1 dari insang kembung 10 (perbesaran 40 X)
Digenea A
Cacing lain yang ditemukan pada insang kembung 1 juga teridentifikasi dalam subkelas digenea. Hal ini dikarenanya oleh adanya batil isap oral pada bagian anterior tubuhnya dan telur yang memenuhi uterusnya. Telur yang terdapat pada tubuh cacing yang ditemukan pada insang kembung 1 ini memiliki karakteristik telur digenea. Telur ini berbentuk bulat sedikit oval, berembrio, memiliki lapisan telur yang tipis pada bagian luarnya, tidak memiliki filamen seperti yang dimiliki telur monogenea dan bergerombol dalam jumlah banyak memenuhi ruang uterus (Gambar 17B).
Cacing ini diduga berasal dari famili yang sama dengan digenea sebelumnya yaitu Hemiuridae dikarenakan bentuk tubuhnya yang memanjang dan silindris dengan tegumen yang berigi dan tampak seperti memiliki ekor pada bagian posterior tubuhnya (Gambar 17A). Cacing ini juga tampak memiliki vitelaria yang padat, berbentuk tubulus dan berbelit-belit (Gambar 17B). Ukuran digenea ini lebih besar dibandingkan digenea sebelumnya dengan panjang 3,2 mm dan lebar 0,42 mm. Identifikasi cacing ini tidak dapat dilakukan hingga tingkat famili bahkan genus karena keadaan preparat yang kurang baik sehingga tidak ada organ dalam lain yang merupakan kunci identifikasi cacing ini yang dapat terlihat.
a b
Gambar 17. (a). Preparat cacing digenea A dari insang kembung 1 (perbesaran 4X), (b). gambaran telur dan vitelaria digenea A (perbesaran 40X)
Digenea B
Cacing yang ditemukan pada insang kembung 7 juga teridentifikasi dalam subkelas digenea. Identifikasi ini dilakukan berdasarkan bentuk tubuhnya yang pipih dan adanya telur dalam jumlah banyak dan bergerombol yang memenuhi
uterusnya (Gambar 18). Telur tampak berbentuk bulat oval, sedikit memanjang seperti ginjal (Gambar 19). Telur juga tidak memiliki filamen atau benang seperti yang dimilki oleh telur cacing monogenea. Cacing ini memiliki panjang 1,59 mm dan lebar 0,18 mm. Jika dilihat dari karakteristik telur, ukuran tubuh, struktur vitelaria, serta adanya tegumen yang sedikit berigi, kemungkinan cacing ini juga berasal dari famili Hemiuridae, namun identifikasi hingga tingkat famili sangat sulit dilakukan karena kondisi preparat yang kurang baik sehingga tidak terlihat organ lain selain uterus dan vitelaria yang dibutuhkan untuk proses identifikasi.
Gambar 18. Preparat cacing digenea B dari insang kembung 7 (perbesaran 4X)
Gambar 19. Struktur telur, uterus dan vitelaria digenea B dari insang kembung 7 (perbesaran 40X)
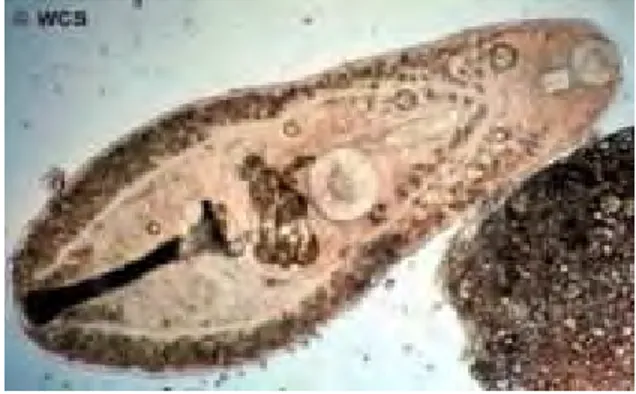
Serkaria Digenea
Serkaria merupakan salah satu stadium larva trematoda digenea, mempunyai batil isap oral maupun ventral dan ciri utama dari stadium ini adalah sebuah ekor pada bagian posterior tubuhnya (Noble & Noble 1989). Jika melihat siklus hidup trematoda digenea, dapat diketahui bahwa serkaria yang berhasil
ditemukan memiliki target jaringan insang dan di insang inilah ia akan menetap dan berdiferensiasi menjadi metaserkaria yang berbentuk kista sebelum dimakan oleh induk semang definitifnya. Paperna (1995) menyatakan bahwa ada beberapa jenis serkaria yang dapat langsung berkembang menjadi digenea dewasa pada inang definitif tanpa melalui stadium metaserkaria.
Gambar 20. Gambaran umum serkaria digenea (Winterbourn 1973)
Berdasarkan struktur anatomis dan ciri morfologisnya, Noble (1989) membagi serkaria menjadi beberapa tipe: serkaria longifurkata, serkaria plerofoserkosa, serkaria monostoma, serkaria gimnosefalosa, serkaria ekinostoma, xifidoserkaria, serkaria amfistoma, dan serkaria brevifurkata. Dari beberapa tipe serkaria tersebut, maka serkaria yang ditemukan kemungkinan termasuk dalam tipe serkaria monostoma karena hanya terdapat satu batil isap pada tubuhnya. (Pardede 2000). Selain dari ada atau tidaknya batil isap, identifikasi serkaria juga dilakukan dengan melihat ukuran tubuhnya. Winterbourn (1973) menyatakan bahwa serkaria monostomata memiliki panjang rata-rata 0,37-0,44 mm dengan lebar 0,11-0,14 mm sedangkan serkaria yang diperoleh dari hasil penelitian memiliki panjang 0,38-0,46 mm dan lebar 0,09-0,11 mm.
Gambar 22. Preparat serkaria digenea dari insang kembung 3 (Perbesaran 10X)
a b c
Gambar 23. Preparat serkaria digenea dari insang kembung 4 (Perbesaran 10X)
Khusus gambar 23 a dan c, masih dalam pertimbangan apakah cacing ini termasuk dalam stadium serkaria atau sudah merupakan bentuk digenea dewasa karena tidak jelasnya pemisahan antara badan dan ekor serkaria.
Genus Anisakis
Cacing parasitik lainnya yang ditemukan teridentifikasi dalam filum Nemathelminthes, kelas Nematoda, subkelas Secernentea, ordo Ascaridida, subordo Ascaridina, famili Anisakidae, genus Anisakis (Grabda 1991). Menurut Smith & Wootten (1978), Anderson (1992), Williams & Jones (1994), Anisakis
baik dalam bentuk larva maupun dewasa, merupakan parasit yang sangat umum dijumpai pada spesies ikan air laut. Grabda (1991) menyebutkan bahwa Anisakis
merupakan golongan cacing nematoda yang berukuran besar dengan tiga buah bibir yang mengelilingi mulutnya. Berdasarkan Koyama et al. (1969), Berland (1989), Ishikura & Namiki (1989), Anderson (2000), Shih & Jeng (2002) dan Shih (2004), identifikasi cacing nematoda famili Ansakidae dilakukan melalui perbandingan karakteristik morfologis dari masing-masing tipe larva. Identifikasi nematoda famili Anisakidae hingga tingkat spesies dan penentuan stadium perkembangan larva dilakukan berdasarkan morfologi sistem pencernaan; bentuk dan keberadaan gigi pembor atau tiga buah bibir yang terletak pada ujung anterior tubuhnya. Selain itu posisi lubang ekskretoris; panjang dan bentuk ventrikulus; keberadaan, panjang dan posisi dari intestinal sekum anterior dan ventrikular appendiks posterior serta bentuk dari ekor post-anal dengan suatu tipe terminal mukron turut menjadi kunci identifikasi famili Anisakidae.
Cacing Anisakis memiliki tiga buah bibir yang mengelilingi mulutnya: satu terletak di dorsal dan dua lainnya di sisi ventro-lateral. Beberapa spesies memiliki bibir yang dipisahkan oleh interlabia yang berukuran lebih kecil (Grabda 1991). Adanya bibir yang berkembang baik pada famili Anisakidae dewasa merupakan karakteristik khas yang membedakannya dari famili lain dalam ordo Ascaridida (Meyers 1975).
Gambar 24. Struktur mulut Anisakis simplex (Anonim 2008)
Kutikula jelas terlihat beralur transversal di sepanjang tubuhnya dan tembus cahaya (Nuchjangreed et al. 2006). Anisakis memiliki esofagus yang lurus, berbentuk silindris atau sedikit mengalami pelebaran di bagian posteriornya, terdiri atas dua bagian, yaitu bagian anterior yang berupa otot dan bagian posterior yang berbentuk kelenjar, dikenal sebagai ventrikulus. Bagian ventrikulus
berhubungan dengan usus halus dan bagian terminal dari sistem pencernaannya adalah rektum yang membuka keluar melalui anus dengan tiga kelenjar anal besar yang berasosiasi dengan rektum.
Anisakis tidak memiliki ujung lobus yang tumpul (sekum dan appendiks) pada pertemuan ventrikulus-sekum maupun berbagai variasi konfigurasi esofageal-intestinal seperti pada beberapa genus lainnya dalam famili Anisakidae. Bagian anterior berhubungan langsung dengan appendiks dan bagian posterior dengan sekum (Meyers 1975). Berdasarkan Grabda (1991), genus Raphidascaris
memiliki satu lobus (appendiks) yang keluar dari ventrikulus dan menjulur ke belakang; pada Pseudoterranova dan Phocanema lobus (sekum) menjulur ke depan dari usus halus, sedangkan pada Contracaecum dan Thynnascaris satu lobus keluar dari ventrikulus, menjulur ke belakang dan satu lobus lagi dari usus halus, menjulur ke depan. Tidak ada lobus yang ditemukan pada genus Anisakis
dan ventrikulus terhubung secara obliq dengan intestin.
Gambar 25. Ujung anterior famili Anisakidae stadium ketiga (Meyers 1975)
Nuchjangreed et al. (2006) menyatakan bahwa ekor Anisakis jantan dewasa dapat teridentifikasi dengan jelas dengan adanya spikula dan bursa kopulatoris. Lubang ekskresi terletak di sebelah ventral yang pada beberapa spesies dapat berada di
bagian puncak kepala pada basis ventro-lateral bibir atau di dekat cincin saraf (Grabda 1991).
Siklus hidup cacing genus Anisakis sangat kompleks (Gambar 26). Dixon (2006) menjelaskan siklus hidup Anisakis sebagai berikut: telur dikeluarkan oleh cacing dewasa melalui feses mamalia laut yang berperan sebagai induk semang definitif. Telur tersebut tenggelam ke dasar laut dan kemudian menetas menjadi larva stadium kedua. Larva stadium kedua hidup bebas di dalam air dan dapat bertahan selama beberapa hari hingga minggu tergantung temperatur air. Larva ini kemudian dimakan oleh krustasea laut yang berperan sebagai induk semang antara pertama dan akan memfasilitasinya untuk melanjutkan perkembangan hidupnya menjadi larva stadium ketiga yang infektif. Ketika krustasea dimakan oleh ikan, larva stadium ketiga tersebut akan bermigrasi ke berbagai jaringan induk semang antara kedua ini dan berkembang menjadi larva stadium ketiga yang lebih maju serta tinggal menetap di organ dalam atau otot. Saat ikan yang terinfeksi Anisakis
ini dimakan oleh induk semang definitifnya, seperti mamalia laut, larva akan dilepaskan ke dalam saluran cerna. Di dalam saluran cerna induk semang definitifnya, larva akan mengalami pergantian kulit (moulting), berkembang menjadi larva stadium keempat dan kemudian menjadi dewasa. Manusia hanya bertindak sebagai induk semang asidental yang tidak memiliki pengaruh terhadap proses transmisi parasit ini.
Distribusi dan lokalisasi infeksi cacing ini pada ikan terbesar ditemukan pada usus kemudian hati dan lambung dan tidak tertutup kemungkinan terjadinya infeksi pada bagian lain dari tubuh ikan seperti sirip, paru-paru, telur di uterus dan insang. Berdasarkan hasil observasi yang sudah pernah dilaporkan, tidak pernah ditemukan adanya migrasi postmortem dari cacing dewasa karena cacing ini tidak dapat bermigrasi ke daging ataupun bagian tubuh dengan tingkat vaskularisasi yang tinggi. Dalamnya distribusi cacing ini mengindikasikan kemampuannya bermigrasi pada lokasi yang berbeda dari organ-organ tubuh ikan (Nuchjangreed
et al. 2006).
Identifikasi terhadap Anisakis selama penelitian dilakukan berdasarkan ciri morfologi dan ukuran tubuhnya. Cacing yang ditemukan tampak jelas memiliki bagian kepala pada ujung anterior tubuhnya dan kutikula yang beralur transversal pada seluruh permukaan tubuhnya (gambar 27).
(a) (b)
Gambar 27. Ujung anterior yang memperlihatkan struktur kepala dan kutikula. (a) Anisakis (Nuchjangreed et al. 2006), (b) Anisakis (penelitian, perbesaran 10X)
Selain dengan melihat ujung anterior, ujung posterior cacing ini juga merupakan kunci identifikasi penting dalam penentuan genus Anisakis. Cacing
yang terlihat jelas. Mukron merupakan suatu penjuluran kontraktil dengan kutikula yang tipis (Grabda 1991). Mukron terdapat pada larva Anisakis simplex, baik jantan maupun betina namun tidak terdapat pada cacing dewasa (Nuchjangreed et al. 2006). Meskipun bursa kopulatoris merupakan struktur yang umum ditemukan pada ekor cacing famili anisakidae jantan, namun struktur tersebut tidak terdapat pada spesies Anisakis simplex dan Pseudoterranova ceticola (Abollo & Pascual 2002).
Gambar 28. Ujung posterior yang memperlihatkan struktur mukron. (a) Anisakis
(Nuchjangreed et al. 2006), (b) Anisakis (penelitian, perbesaran 10X)
Larva Anisakis simplex stadium ketiga berwarna putih agak pink, translusen dan berukuran kecil (Grabda 1991), dengan panjang yang bervariasi tergantung spesies. Nuchjangreed et al. (2006) menyatakan bahwa larva Anisakis
memiliki panjang yang berkisar antara 9-36 mm dan lebar 0,5-1 mm, sedangkan menurut Anonim(2008) larva Anisakis memiliki panjang antara 0,5 hingga 3 mm. Cacing yang ditemukan selama penelitian memiliki panjang 1,1 mm.
Berdasarkan siklus hidupnya dan dengan mempertimbangkan ukuran serta ciri morfologis yang terlihat selama proses pengamatan, termasuk tidak adanya lobus sekum maupun appendiks pada pertemuan ventrikulus-sekum, seperti yang dimiliki beberapa genus lainnya dalam famili Anisakidae, dapat disimpulkan bahwa cacing famili Anisakidae yang berhasil diisolasi dari insang ikan kembung
ini berada pada stadium larva infektif atau stadium larva ketiga dan termasuk dalam genus Anisakis.
Gambar 29. Preparat utuh Anisakis, larva stadium ketiga (Perbesaran 4X)
Anisakis pernah dilaporkan terdapat pada beberapa jenis ikan kembung yang berada pada perairan Pulau Panggang, Kepulauan Seribu, Jakarta Utara dalam observasi selama tahun 1974 hingga tahun 1975 (Hadidjaja 1977). Larva
Anisakis dapat menginfeksi manusia melalui ikan yang dimakannya (fish borne zoonosis). Infeksi yang terjadi umumnya disebabkan oleh larva stadium ketiga (Buchmann & Bresciani 2001) dan dikenal sebagai agen penyebab penyakit pada manusia yang disebut anisakiosis (Van Thiel 1962). Anisakiosis terjadi karena adanya migrasi larva Anisakis ke dalam saluran pencernaan manusia akibat memakan larva hidup cacing Anisakis namun larva ini tidak akan pernah menjadi dewasa di dalam saluran pencernaan manusia, induk semang asidentalnya (Huss & Embarek 2003).
Cacing Anisakis beresiko terhadap kesehatan manusia melalui dua cara, yaitu melalui infeksi oleh larva cacing yang berasal dari ikan yang dimasak kurang sempurna dan kemudian bermigrasi ke dalam usus atau melalui reaksi alergi oleh bahan kimia yang ditinggalkan cacing pada daging sehingga ikan yang terinfeksi cacing ini dapat menghasilkan reaksi anafilaktik (alergi) pada manusia yang memakannya yang sensitif terhadap immunoglobulin E (IgE) (Anonim 2008). Reaksi anafilaktik inilah yang sering dikenal sebagai ”alergi makanan laut” atau sea food allergy. Anisakiosis menimbulkan gejala yang tidak spesifik bahkan sering tidak terdiagnosa namun pada saluran pencernaan manusia telah terbentuk ulkus akibat memakan larva hidup cacing Anisakis ini. Spesies Anisakidae yang paling sering menyebabkan penyakit pada manusia (bersifat zoonosis) adalah
Anisakis simplex dan Pseudoterranova decipiens. Stadium larva infektif cacing parasit ini dapat ditemukan pada seluruh bagian tubuh ikan terutama organ dalam dan otot sejumlah ikan konsumsi dan yang memiliki nilai ekonomi tinggi seperti kembung, salmon, cod, makarel dan termasuk cumi-cumi (Dixon 2006).
Tindakan pencegahan kejadian anisakiosis yang paling efektif adalah mencegah penularan dari ikan terinfeksi larva Anisakis yang akan dikonsumsi ke manusia, antara lain tidak memakan ikan yang mentah, pemanasan ikan yang akan dimakan minimal hingga suhu 600C, pembekuan hingga suhu -200C selama 24 jam, penggaraman pada larutan garam pekat selama 10 hari (Wootten 2008).