KARAKTER AGRONOMI DAN MARKA SSR SEBAGAI
KARAKTER SELEKSI GENERASI AWAL UNTUK
TOLERANSI SUHU TINGGI PADA PADI
VICTOR MANOTAR PADEMAN MANALU
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul “Karakter Agronomi dan Marka SSR sebagai Karakter Seleksi Generasi Awal untuk Toleransi Suhu Tinggi pada Padi” benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Mei 2016
RINGKASAN
VICTOR MANOTAR PADEMAN MANALU. Karakter Agronomi dan Marka SSR sebagai Karakter Seleksi Generasi Awal untuk Toleransi Suhu Tinggi pada Padi. Dibimbing oleh DESTA WIRNAS dan SUDARSONO.
Penelitian ini bertujuan untuk (1) memperoleh informasi tentang aksi gen dan pewarisan sifat karakter agronomi padi terhadap cekaman suhu tinggi di rumah kaca; (2) memperoleh marka simple sequence repeat (SSR) terpaut toleransi cekaman suhu tinggi melalui metode bulk segregrant analisis dan single marker analysis.
Penelitian pertama bertujuan untuk mencari kriteria karakter seleksi dan memilih segregan terbaik di generasi awal berdasarkan beberapa karakter agronomi untuk adaptasi padi terhadap cekaman suhu tinggi. Penelitian dilaksanakan dari bulan September 2014 sampai bulan Januari 2015. Materi genetik yang digunakan adalah 213 individu segregant F2 yang berasal dari persilangan IPB 4S dan Situ Patenggang, 20 individu tetua IPB 4S, 20 individu tetua Situ Patenggang. Semua materi genetik ditanam dalam kondisi tercekam suhu tinggi di rumah kaca Institut Pertanian Bogor. Rata-rata suhu minimum dan maksimum pada penelitian ini adalah 24 °C dan 42.09 °C. Hasil penelitian menunjukkan bahwa karakter: jumlah anakan produktif dan total, jumlah gabah bernas, dan bobot gabah bernas memiliki nilai heritabilitas yang tinggi, aksi gen aditif, dan koefisien keragaman genetik yang tinggi. Karakter-karakter tersebut dapat dijadikan kriteria seleksi pada generasi awal untuk adaptasi padi terhadap cekaman suhu tinggi. Karakter seleksi dapat dilakukan baik dengan satu karakter atau beberapa kriteria karakter.
Penelitian kedua bertujuan untuk memperoleh primer SSR terpaut toleran terhadap suhu tinggi dengan menggunakan bulked segregant analysis (BSA) dan dilanjutkan dengan single marker analysis (SMA). Penelitian dilakukan di Laboratorium Biologi Molekuler Tanaman, Departemen Agronomi dan Hortikultura, Institut Pertanian Bogor (IPB). Bahan genetik yang digunakan DNA tetua IPB 4S, Situ Patenggang, DNA genotipe F2, 12 Primer SSR. Hasil penelitian menunjukkan primer SSR RM 337 mengikuti segregasi hukum Mendel, kemudian berdasarkan single marker analysis dengan menggunakan karakter bobot gabah bernas menunjukkan primer SSR RM 337 terpaut toleran terhadap suhu tinggi dengan nilai peluang yang sangat nyata. Genotipe F2 yang memiliki pola pita DNA seperti Situ Patenggang (Tetua toleran) dan daya hasil tinggi di dapatkan 54 genotipe F2. Diferensial seleksi berdasarkan genotyping dengan menggunakan primer RM 337 menghasilkan kenaikan bobot gabah bernas sebesar 37.96%. Kata kunci: aksi gen, cekaman suhu tinggi, heritabilitas, karakter seleksi, marka
SUMMARY
VICTOR MANOTAR PADEMAN MANALU. Characters Agronomy and SSR Marker as Selection Characters in Early Generation for Heat Stress Tolerance in Rice. Supervised by DESTA WIRNAS dan SUDARSONO.
This study aims to (1) obtain information about gene action and inheritance of agronomic traits of rice to stress the high temperatures in the greenhouse; (2) obtaining simple sequence repeat (SSR) markers linked to high temperature stress tolerance through bulked segregant anlysis (BSA) and single marker analysis (SMA) method.
The first research was aimed to find selection characters and to select the best segregant in early generation based on agronomic characters for adaptation of rice to heat stress. The experiment was carried out in September 2014 to Januari 2015. About 213 F2 segregants generated trough hybiridization between IPB 4S (sensitive parent) and Situ Patenggang (tolerant parent), 20 individuals of IPB 4S, and 20 indiviudals of Situ Patenggang, were used as genetic materials. All genetic materials were exposed to high temperature stress by growing in a green house of Bogor Agricultural University. The average temperature in the research period was 24 °C and 42.09 °C, respectively for minimum and maximum temperature. The results showed charachter: productive and total tiller number, total grain number, and seed weight have high heritability, gene action additive and high coeffiecient variability genetics. That charachters could be proposed as selection characters in early generation for rice breeding for adaptation to high temperature stress. The characters could be applied in either single trait or multiple traits selection.
The second study aims to obtain SSR marker linked to tolerance to high temperature using bulked segregant analysis (BSA) and continued with use single marker analysis (SMA). The experiment was conducted at the Laboratory of Plant Molecular Biology, Departement Agronomy and Horticulture, Bogor Agricultural University (IPB). The genetic materials include parental DNA IPB 4S, Situ Patenggang, and the DNA of F2 genotypes and the 12 primer SSR. The primary result showed that the primer RM 337 was folowing segragation based on Law of Mendel. After that based on single marker analysis with use the weight grain character, the primer RM 337 was linked to tolerance to high temperature with show the significant result. Based on analysis of molecular, F2 genotypes have similiar to DNA pattern of Situ Patenggang (tolerant parent) and high yielding was result 54 genotypes F2. Selection differential based on genotyping use primer RM 337 can be increase weight grain were 37.96%.
Keywords: characters selection, gene action, heat stress, heritability, molecular marker, SSR.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
KARAKTER AGRONOMI DAN MARKA SSR SEBAGAI
KARAKTER SELEKSI GENERASI AWAL UNTUK
TOLERANSI SUHU TINGGI PADA PADI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
Judul Tesis : Karakter Agronomi dan Marka SSR sebagai Karakter Seleksi Generasi Awal untuk Toleransi Suhu Tinggi pada Padi
Nama : Victor Manotar Pademan Manalu
NIM : A253130031
Disetujui oleh Komisi Pembimbing
Dr Desta Wirnas, SP, MSi Ketua
Prof Dr Ir Sudarsono, MSc Anggota
Diketahui oleh
Ketua Program Studi
Pemuliaan dan Bioteknologi Tanaman
Dr Ir Yudiwanti Wahyu E.K, MS
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji syukur penulis panjatkan ke hadirat Tuhan Yang Maha Esa karena dengan rahmat dan karunia-Nya, penulis dapat menyelesaikan penelitian dan penulisan tesis yang berjudul “Karakter agronomi dan marka SSR sebagai karakter seleksi generasi awal untuk toleransi suhu tinggi pada padi”.
Penelitian dan penulisan tesis ini berlangsung di bawah bimbingan Dr Desta Wirnas, SP, MSi selaku ketua komisi pembimbing dan Prof Dr Ir Sudarsono, MSc selaku anggota komisi pembimbing. Penulis menyampaikan terima kasih dan penghargaan yang tulus atas ilmu yang bermanfaat, motivasi, kesabaran, dan waktu yang telah diluangkan dalam mengarahkan dan membimbing penulis mulai dari perencanaan, penyusunan dan penyelesaian tesis. Ucapan terima kasih penulis sampaikan kepada Dr Ir Trikoesoemaningtyas, MSc selaku penguji luar komisi yang berkenan untuk menjadi penguji luar komisi dan atas masukannya dalam penulisan tesis ini.
Ucapan terima kasih juga penulis sampaikan kepada Direktorat Jenderal Pendidikan Tinggi (Dirjen Dikti) yang telah memberikan penulis beasiswa melalui Program Beasiswa Pendidikan Pascasarjana Dalam Negeri (BPPDN) Calon Dosen (Caldos) pada tahun 2013.
Penghargaan yang setinggi-tingginya serta rasa terima kasih yang tulus, penulis sampaikan kepada Kepala University Farm, Institut Pertanian Bogor beserta staf di rumah kaca untuk pelaksanaan penelitian, teknisi rumah kaca Bapak Mamat dan Mas Eki yang banyak membantu di lapangan, teknisi Laboratorium Plant Biology Molecular (PMB) Departemen Agronomi dan Hortikultura Yudiansyah, S.Si yang telah banyak membantu penelitian. Penghargaan yang tulus penulis sampaikan kepada seluruh staf pengajar pada Departemen Agronomi dan Hortikultura, khususnya program studi Pemuliaan dan Bioteknologi Tanaman (PBT) atas ilmu yang telah diberikan selama penulis mengikuti studi di sekolah Pascasarjana IPB. Terima kasih juga penulis sampaikan kepada teman-teman seperjuangan PBT IPB angkatan 2013, rekan-rekan di Laboratorium Pemuliaan Tanaman dan Laboratorium Plant Biology Molecular IPB, dan sahabatku Eka Jan Virgin Harquasum, SP., Merry G. Meliala, SP., Listya Pramudita, SP., untuk persahabatan dan kebersamaan dalam berbagi ilmu, berbagi suka maupun duka. Ucapan terima kasih juga disampaikan kepada Prof Dr Ir Kartina, AM, MP, Dr Ir Rusmana, MP, Dr Susiyanti, SP, MP, (Dosen di Universitas Sultan Ageng Tirtayasa-Untirta) atas surat rekomendasi untuk melanjutkan studi di Sekolah Pascasarjana IPB, dan (Alm) Ir. Sahiral Yakub, MP (Pembimbing akademik sewaktu S1 di Untirta) serta Ratna Fitry, SP, MP (Dosen S1 sewaktu di Untirta) atas saran untuk melanjutkan studi pascasarjana di bidang Pemuliaan Tanaman. Semoga segala kebaikan dibalas berlipat oleh Tuhan Yang Maha Esa.
Namboru-Amangboru (dari silsilah keluarga Manalu), Tulang-Nantulang dan Tante-Bapauda (dari silsilah keluarga Limbong dan Hutagaol) atas kasih sayang, doa dan suportnya bagi penulis untuk terus melangkah dan berjuang menyelesaikan studi. Semoga tulisan ini membawa manfaat dalam bidang pemuliaan tanaman dan menambah rasa syukur penulis kepada sang Pencipta.
Bogor, Mei 2016
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
1 PENDAHULUAN
Latar Belakang 1
Tujuan Penelitian 2
Hipotesis 2
2 TINJAUAN PUSTAKA
Tanaman Padi 5
Pemuliaan Tanaman Padi terhadap Cekaman Abiotik 6
Pengaruh Suhu Tinggi terhadap Pertanaman Padi 7
Analisis Pendugaan Nilai Parameter Genetik 8
Marka Molekuler 9
3 STUDI PEWARISAN SIFAT TOLERANSI PADI TERHADAP CEKAMAN SUHU TINGGI
Abstrak 11
Abstract 12
Pendahuluan 13
Metode Penelitian 14
Hasil dan Pembahasan 18
Simpulan 30
Daftar Pustaka 31
4 SELEKSI MARKA SSR UNTUK TOLERANSI TERHADAP CEKAMAN SUHU TINGGI PADA POPULASI F2 PADI Abstrak 33 Abstract 34 Pendahuluan 35 Metode Penelitian 35
Hasil dan Pembahasan 39
Simpulan 50
Daftar Pustaka 51
5 PEMBAHASAN UMUM 55 6 SIMPULAN DAN SARAN Simpulan 59
Saran 59
DAFTAR PUSTAKA 61
LAMPIRAN 69
DAFTAR TABEL
1 Respon tanaman padi terhadap suhu berbeda pada beberapa stadia pertumbuhan
7 2 Nilai tengah karakter agronomi tetua IPB 4S, Situ Patenggang dan F2
padi hasil persilangan IPB 4S x Situ Patenggang pada kondisi tercekam suhu tinggi
4 Nilai pendugaan komponen ragam, hertitabilitas dan Koefisien Keragaman Genetik arti luas populasi F2 padi (hasil persilangan IPB 4S dan Situ Patenggang) pada kondisi tercekam suhu tinggi
26
5 Diferensial seleksi berdasarkan karakter bobot gabah bernas pada populasi F2 padi
28 6 Diferensial seleksi multikarakter pada populasi F2 padi 29 7 Kemajuan genetik dan dugaan respon seleksi pada padi generasi F3
berdasarkan seleksi langsung dan multikarakter dengan indeks seleksi terboboti
30
8 Jenis primer SSR yang digunakan dalam penelitian ini 37 9 Nilai fenotipe bobot gabah bernas populasi F2 padi (IPB 4S x Situ
Patenggang) pada kondisi tercekam suhu tinggi
41 10 Hasil uji square pada primer RM 337 pada padi 43 11 Keragaan bobot gabah bernas 54 genotipe F2 padi hasil seleksi
berdasarkan single marker analysis
48 12 Kemajuan seleksi berdasarkan primer RM 337 pada padi 49 13 Kemajuan genetik dan dugaan respon seleksi pada padi generasi F3
berdasarkan seleksi primer RM 337
50
DAFTAR GAMBAR
1 Diagram Alir penelitian 3
2 Keragaan pertanaman padi di lapangan 19
3 Suhu maksimum dan minimum selama penelitian 20
4 Kelembapan udara selama penelitian 20
5
Pola sebaran jumlah anakan total dan jumlah anakan produktif pada padi (IPB 4S x Situ Patenggang) pada kondisi tercekam suhu tinggi
24
6
Pola sebaran kehijauan daun SPAD 105 HST dan lama pengisian biji pada padi (IPB 4S x Situ Patenggang) pada kondisi tercekam suhu tinggi
24
7
Pola sebaran tinggi tanaman saat panen dan panjang malai pada padi (IPB 4S x Situ Patenggang) pada kondisi tercekam suhu tinggi
24 8 Pola sebaran jumlah gabah bernas malai-1dan jumlah gabah
hampai malai-1 pada padi (IPB 4S x Situ Patenggang) pada kondisi tercekam suhu tinggi
9 Pola sebaran jumlah gabah total malai-1dan bobot gabah bernas
distandarisasi dengan nilai Z pada populasi F2 padi (IPB 4S x Situ Patenggang)
19 Boxplot hasil single marker analysis pada padi F2 (IPB 4S x Situ Patenggang)
2 Deskripsi padi varietas Situ Patenggang 72
3 Suhu dan kelembapan selama penelitian 73
4 Algoritma eksplorasi kecenderungan sebaran sifat-sifat kuantitatif kaitannya dengan aksi gen aditif, dominan serta epistasis komplementer dan duplikat.
74
5 Keragaan segregan padi IPB 4S x Situ Patenggang hasil seleksi berbasis 1 karakter dengan intensitas seleksi 25%
75 6 Keragaan segregan padi IPB 4S x Situ Patenggang hasil seleksi
berbasis indeks seleksi terboboti multikarakter dengan intensitas seleksi 25%
77
7 Keragaan segregan padi IPB 4S x Situ Patenggang hasil seleksi berbasis primer RM 337
1
1 PENDAHULUAN
Latar Belakang
Padi merupakan bahan makanan pokok sebagian besar penduduk Indonesia. Kebutuhan akan konsumsi beras terus semakin meningkat dari tahun ke tahun seiring dengan pertambahan penduduk.
Diduga salah satu faktor yang menyebabkan menurunnya produktivitas padi adalah pemanasan global sehingga menyebabkan peningkatan suhu permukaan bumi yang dapat mempengaruhi pertumbuhan dan perkembangan tanaman padi (Las et al. 1986; IRRI 2004). Data peningkatan suhu muka bumi di beberapa kota besar Indonesia antara lain: Jakarta antara 1.04 – 1.4 °C 100 tahun-1,
Cilacap antara 3.38 – 3.41°C 100 tahun-1, Medan antara 1.55 –1.98 °C 100 tahun-1, Surabaya antara 1.46 – 3.29 °C 100 tahun-1, Bau Bau hingga 3.63 °C 100 tahun-1 dan Makassar antara 1.84 – 2.83 °C 100 tahun-1 (Aldrian et al. 2014). Terdapat kemungkinan bahwa wilayah disekitarnya juga mengalami peningkatan suhu.
Menurut Taiz dan Zeiger (2002) cekaman suhu tinggi didefinisikan sebagai kenaikan suhu yang melebihi ambang kerusakan untuk periode waktu cukup menyebabkan kerusakan yang tidak dapat balik (irreversible) pada pertumbuhan dan perkembangan tanaman. Berbagai hasil penelitian menunjukkan bahwa peningkatan suhu secara langsung akan mempengaruhi pertumbuhan dan perkembangan tanaman padi (Yoshida 1978; Wassmann et al. 2009), mulai dari perkecambahan, pertumbuhan vegatatif sampai dengan pertumbuhan generatif (Jagadish et al. 2008). Cekaman suhu tinggi selama fase berbunga padi menyebabkan penurunan vigor serbuk sari, dan fertilitas spikelet (Prasad et al. 2006; Jagadish et al. 2011; Poli et al. 2013). Penurunan vigor serbuk sari dapat dianggap sebagai faktor fisiologi yang bertanggung jawab terhadap penurunan produksi gabah pada kondisi suhu tinggi (Tang et al. 2008; Zhang et al. 2008; Xiao et al. 2011).
Sifat toleransi padi terhadap cekaman suhu tinggi dapat diperbaiki melalui seleksi dalam program pemuliaan tanaman. Keberhasilan seleksi atau efisiensi seleksi untuk meningkatkan daya hasil ditunjukkan oleh perolehan kemajuan genetik dari genotipe terpilih. Kemajuan genetik yang diperoleh sangat tergantung kepada variabilitas genetik, heritabilitas, dan korelasi genetik (Roy 2000). Kemajuan genetik dapat dimaksimalkan dengan memilih karakter seleksi yang tepat, yaitu yang memiliki nilai heritabilitas tinggi dan berkorelasi dengan hasil. Oleh karena itu informasi tentang pola pewarisan diperlukan.
Seleksi pada lingkungan bercekaman harus dilakukan di lingkungan target dengan tujuan untuk dapat memaksimalkan ekspresi gen-gen yang mengendalikan daya hasil maupun daya adaptasi (Cooper dan Byth 1996). Seleksi pada kondisi bercekaman dapat dilakukan berdasarkan fenotipe, marka molekuler, dan gabungan antara fenotipe dan marka molekuler (Bernando 2002).
2
Kemajuan dalam bidang biologi molekuler memungkinkan keragaman genetik suatu populasi dapat diamati pada tingkat deoxyribo nucleic acid (DNA). Marka molekuler ini tidak dipengaruhi oleh faktor lingkungan. Marka molekuler yang digunakan dalam penelitian ini adalah marka simple sequence repeat (SSR) yang berdasarkan pada sejumlah sekuen DNA berulang (2 – 5 nukleotida) yang terdapat dalam mikrosatelit. Jumlah pengulangan SSR ini bervariasi antar individu dan merupakan sumber polimorfisme di tanaman. Marka SSR merupakan marka berlokus tunggal, multialelik dan kodominan (Acquaah 2007). Marka SSR dipilih karena jumlahnya yang melimpah dan terdistribusi merata, dihasilkan cepat melalui mesin polymerase chain reaction (PCR), mudah diskoring dan informasi sekuen primernya mudah diakses melalui publikasi (Saghai-Maroof et al. 1994). Penggunaan marka molekuler SSR sebagai alat bantu seleksi telah dilakukan pada padi untuk sifat ketahanan terhadap suhu tinggi (Zhu et al. 2005; Xiao et al. 2011; Buu et al. 2014).
Informasi marka SSR yang dapat digunakan sebagai alat bantu seleksi padi untuk adaptasi pada cekaman suhu tinggi di Indonesia masih perlu diperoleh. Salah satu metode untuk mengembangkan marka seleksi berbasis marka molekuler adalah bulked segregant analysis (BSA) atau single marker analyisis (SMA).
Tujuan Penelitian Tujuan penelitian ini adalah:
1. Memperoleh informasi tentang pola pewarisan karakter agronomi pada padi toleran suhu tinggi pada populasi F2.
2. Memperoleh karakter kuantitatif sebagai karakter seleksi untuk toleransi terhadap suhu tinggi.
3. Memperoleh marka molekuler yang spesifik terpaut toleransi suhu tinggi. Hipotesis
Adapun hipotesis dalam penelitian ini adalah:
3
Gambar 1 Alur penelitian Persilangan
F1 IPB 4S x Situ Patenggang
Populasi Padi F2 Penelitian
Sebelumnya
Percobaan 1 Studi Pewarisan Percobaan 2 Seleksi Marka
Seleksi Marka SSR pada tetua
1. Informasi pola pewarisan. 2. Karakter seleksi
3. Marka seleksi SSR terpaut toleran suhu tinggi tinggi.
Genotyping
Verifikasi dengan Single Marker Analysis pada Populasi F2 untuk karakter bobot gabah bernas Screening 64 genotipe padi, terpilih yang ekstrim peka dan toleran:
Peka: IPB 6R, IR64, IPB 4S,
Toleran: Mekongga, Situ Patenggang dan Marinah.
1 kelompok Bulk Toleran
1 kelompok Bulk Peka
Metode Bulked Segregant Analysis (BSA)
5
2 TINJAUAN PUSTAKA
Tanaman Padi
Padi (Oryza sativa L.) termasuk ke dalam genus oryza. Genus oryza terdiri dari 22 spesies dengan 2n = 2x = 24 dan 2n = 4x = 48 yang mewakili genom-genom AA, BB, CC, BBCC, CCDD, EE dan FF (Vaughan 1994). Dengan menggunakan pendekatan molekuler, yaitu hibridisasi total genom DNA, telah berhasil ditemukan genom baru untuk dua komplek spesies liar, O. meyeriana dan O. ridleyi. Genom GG untuk O. meyeriana diploid dan HHJJ untuk komplek O. rdleyi alotetraploid (IRRI 1997).
Padi merupakan golongan tumbuhan Graminae dengan batang yang tersusun dari beberapa ruas. Ruas-ruas ini merupakan bumbung kosong yang ditutup oleh buku dan panjang ruasnya tidak sama. Ruas yang terpendek berada di pangkal batang, ruas yang kedua dan seterusnya lebih panjang dari ruas-ruas yang lebih bawah. Pada buku bagian bawah dari ruas, tumbuh daun pelepah yang membalut ruas sampai buku bagian atas (Siregar 1987). Tepat pada buku bagian atas ujung daun pelepah meperlihatkan percabangan dengan cabang yang terpendek menjadi ligule (lidah) daun serta bagian yang terpanjang dan terbesar menjadi helaian daun. Daun pelepah itu menjadi ligule dan pada helaian daun terdapat dua embel sebelah kiri dan kanan yang disebut auricular. Auricular dan ligule yang kadang-kadang berwarna hijau dan ungu dapat digunakan sebagai alat untuk mendeterminasi dan identifikasi suatu varietas (Siregar 1987). Daun pelepah yang membalut ruas yang paling atas batang umumnya disebut daun bendera. Tepat dimana daun pelepah teratas menjadi ligule dan daun bendera, disitulah timbul ruas yang menjadi bulir padi (de Datta 1981).
Bunga padi adalah bunga terminal yang berbentuk malai terdiri dari bunga-bunga tunggal (spikelet). Tiap bunga tunggal terdiri dari 2 lemma steril, lemma (sekam besar), palea (sekam kecil), 6 buah benang sari yang masing-masing memiliki 2 kotak sari dan sebuah putik (Syukur et al. 2012). Pada dasar bunga terdapat ladicula (berperan penting terhadap mekarnya bunga). Ladicula berfungsi mengatur dalam pembuahan palea, pada waktu berbunga ia menghisap air dari bakal buah, sehingga mengembang. Pengembangan ini mendorong lemma dan palea terpisah dan terbuka (Grist 1975).
Buah padi yang disebut biji padi atau bulir/gabah merupakan buah padi yang tertutup oleh lemma dan palea. Lemma dan 4 palea serta bagian lain akan membentuk sekam atau kulit gabah, lemma selalu lebih besar dari palea dan menutupi hampir 2/3 permukaan beras, sedangkan sisi palea tepat bertemu pada bagian sisi lemma. Gabah terdiri atas biji yang terbungkus sekam. Sekam terdiri atas gluma rudimenter dan sebagian dari tangkai gabah (pedicel) (Badan Litbang 2009).
6
Tanaman padi membentuk rumpun dengan anakannya. Anakan akan tumbuh pada dasar batang. Pembentukan anakan akan terjadi secara bersusun, yaitu: 1) anakan pertama (primer), anakan primer ini tumbuh di antara dasar batang dan daun sekunder, sedangkan pada pangkal batang anakan primer terbentuk perakaran. Anakan primer ini tetap melekat pada batang utama hingga masa pertumbuhan berikutnya, namun dalam mendapatkan zat makanan, anakan primer tidak tergantung pada batang utama sebab memiliki perakaran sendiri, 2) anakan kedua (sekunder), anakan ini tumbuh pada batang bawah anakan primer, yaitu pada buku pertama dan juga membentuk perakaran sendiri. 3) anakan ketiga (tersier), anakan tersier yang tumbuh pada buku pertama pada batang anakan sekunder dengan bentuk yang serupa dengan anakan primer dan sekunder (Yoshida 1981; Siregar 1987).
Pemuliaan Tanaman Padi terhadap Cekaman Abiotik
Salah satu tahapan dalam pembentukan varietas baru adalah melakukan perbaikan daya adaptasi tanaman terhadap lingkungan tumbuh yang menjadi target pengembangan. Secara genetik adaptasi memiliki makna sebagai suatu kondisi yang berkaitan dengan konstitusi genetik dari suatu genotipe sehingga menjadikan genotipe tertentu cocok pada kondisi lingkungan tumbuh dimana genotipe itu berada. Adaptasi adalah fungsi dari gen yang dimiliki tanaman dalam merespon lingkungan tumbuh. Gen-gen tersebut mengatur dan mengendalikan proses biokimia dan fisiologi tanaman selama proses pertumbuhan dan perkembangan tanaman sehingga beradaptasi dan sesuai atau sebaliknya, dengan sumber daya lingkungan yang tersedia, dan dengan berbagai kemungkinan cekaman yang ada (Bidinger et al. 1996).
Cekaman abiotik merupakan salah satu faktor pembatas baik dalam kegiatan ekstensifikasi maupun intensifikasi pertanian. Lingkungan bercekaman didefinisikan sebagai lingkungan suboptimum untuk pertumbuhan dan produksi tanaman (Wirnas 2007). Upaya perbaikan daya hasil dan adaptasi tanaman terhadap cekaman abiotik dapat dilakukan melalui serangkaian program pemuliaan tanaman.
Berbagai upaya pemuliaan tanaman dengan tujuan untuk memperbaiki karakter terhadap toleransi cekaman abiotik pada tanaman padi telah dilakukan oleh para pemulia, antara lain untuk toleransi terhadap kekeringan (Subashri et al. 2009), toleransi terhadap genangan (Neeraja et al. 2007), toleransi terhadap salinitas (Islam et al. 2011), toleransi terhadap defisiensi unsur P (Mu et al. 2008), toleransi terhadap cekaman Al (Nguyen et al. 2002), toleransi terhadap cekaman suhu tinggi (Zhang et al. 2009), toleransi sifat ketahanan terhadap suhu tinggi (Zhu et al. 2005; Zhang 2009; Buu et al. 2014).
7 Pengaruh Suhu Tinggi terhadap Pertanaman Padi
Budi daya padi sawah sering dianggap sebagai penyumbang terhadap perubahan iklim melalui emisi gas CO2, CH4 dan N2O, tetapi budi daya padi juga
terkena dampak negatif dari perubahan iklim. Menurut Koesmaryono (2014) dampak perubahan iklim di Indonesia antara lain: pergeseran musim hujan dan kemarau, pergeseran waktu tanam padi, ledakan serangan hama dan penyakit (seperti ledakan wereng batang coklat dan blast akibat keberadaan tanaman yang tidak serempak di lapangan dan suhu yang rendah di malam hari pada daerah yang relatif kering), kerusakan lahan sawah akibat kekeringan dan kebanjiran.
Cekaman panas akibat suhu udara yang tinggi merupakan ancaman yang serius bagi produksi tanaman di seluruh dunia. Emisi gas yang diakibatkan aktifitas manusia secara substansial menambah konsentrasi gas-gas rumah kaca, terutama CO2, metana, klorofluorokarbon, dan oksida-oksida nitrus. Sirkulasi
global yang berbeda memprediksi bahwa gas-gas rumah kaca secara bertahap akan meningkatkan rata-rata suhu udara dunia.
Menurut laporan International Panel on Climatic Change (IPCC), rata – rata suhu global akan meningkat 0.3 °C setiap dekadenya (Jones et al. 1999), secara berturut-turut akan mencapai kurang lebih 1 dan 3 °C di atas nilai suhu sekarang pada tahun 2025 dan 2100, dan mengarah pada pemanasan global. Peningkatan suhu dapat mengakibatkan perubahan persebaran geografis dan musim tanam komoditas pertanian dengan cara menciptakan ambang batas suhu untuk awal musim dan menyebabkan kemasakan tanaman yang lebih awal (Porter 2005).
Menurut Yoshida (1978) suhu kritis untuk stadium perkecambahan, proses anakan, inisiasi dan perkembangan inflorens bunga, serta proses pemasakan bulir padi telah diidentifikasi (Tabel 1).
Tabel 1 Respon tanaman padi terhadap suhu berbeda pada beberapa stadia pertumbuhan (Yoshida 1978)
Stadia pertumbuhan Kisaran suhu kritis (°C)
Rendah Tinggi Optimum
8
daerah tropis dapat mengalami gangguan pertumbuhannya akibat cekaman suhu tinggi. Gejala gangguan cekaman suhu tinggi beragam pada berbagai fase pertumbuhan padi. Beberapa gangguan yang dapat terjadi pada pertumbuhan padi antara lain turunnya daya berkecambah pada benih padi, klorosis daun, penurunan tinggi tanaman, penurunan jumlah anakan pada fase vegetatif, pemutihan spikelet, penurunan jumlah bulir padi yang terbentuk, perlambatan fase heading, peningkatan jumlah bulir hampa, peningkatan spikelet steril, proses pemasakan bulir padi tidak sempurna (Nishiyama dan Satake 1981; Prasaad et al. 2006; Jagadish et al. 2008; Wasmann et al. 2009).
Suhu daun yang tinggi dan kahat air akan mengarah pada cekaman panas. Suhu tinggi mengurangi stabilitas membran. Salah satu bentuk adaptasi cekaman suhu tinggi melindungi daun dari panas berlebihan (Levitt 1980). Menurut Zhang et al. (2005), respon tanaman terhadap suhu tinggi di antaranya adalah penurunan sintesa protein normal dan percepatan transkripsi dan translasi heat shock protein (HSP). Faktor transkripsi memediasi akumulasi HSP dalam respon terhadap kejutan panas. Secara umum perubahan yang terjadi pada tanaman yang tercekam suhu tinggi dikelompokkan menjadi beberapa tipe yaitu perubahan morfologis, anatomis, fenologis, dan fisiologis (Wahid et al. 2007).
Suhu bersama dengan penyinaran adalah pendorong utama untuk perkembangan tanaman (Kroff et al. 1995). Suhu optimum untuk perkembangan normal padi berkisar 27 – 32 °C (Yin et al. 1996). Suhu tinggi mempengaruhi hampir semua tahap pertumbuhan padi, yaitu dari munculnya bunga, pemasakan, dan panen. Penyerbukan memainkan peran yang dominan dalam produktivitas tanaman. Umumnya, reproduksi jantan pada perkembangan padi dikenal lebih sensitif terhadap stress panas (Wassmann dan Dobermann 2007). Prasad et al. (2006) melaporkan bahwa stress suhu tinggi selama berbunga pada padi menyebabkan penurunan produksi serbuk sari. Penurunan gabah disebut karakter fenotipe tanaman padi pada suhu tinggi, sedangkan penurunan perkecambahan serbuk sari dan aktivitasnya dapat dianggap sebagai faktor fisiologi yang bertanggung jawab terhadap penurunan gabah (Tang et al. 2008).
Pendugaan Nilai Parameter Genetik
9 al. 2000). Heritabilitas dari suatu karakter dapat diketahui dengan menduga komponen ragam menggunakan studi generasi dasar yaitu populasi P1, P2, F1, F2, dan backcross (Syukur 2012).
Pendekatan untuk pendugaan aksi gen berupa pendugaan menggunakan parameter kemenjuluran kurva (skewness) dan keruncingan kurva (kurtosis) serta sebaran populasi bersegregasinya pada F2. Nilai kemenjuluran kurva dapat digunakan untuk menunjukkan aksi gen yang mengendalikan suatu karakter dan nilai keruncingan kurva digunakan untuk menduga jumlah gen pengendali (Roy 2000).
Koefisien keragaman genetik digunakan untuk menduga luas atau tidaknya keragaman genetik yang dimiliki masing-masing karakter. Bila tingkat keragaman genetik sempit maka keragaman antar individu dalam populasi relatif seragam, sehingga seleksi untuk perbaikan sifat menjadi kurang efektif. Kemudian apabila keragaman genetik semakin luas, maka peluang keberhasilan seleksi dalam meningkatkan frekuensi gen yang diinginkan semakin besar pula (Chahal dan Gosal 2003).
Seleksi untuk karakter kuantitatif dapat dijelaskan dalam perubahan atau perbaikan nilai tengah dari generasi yang diturunkan dari individu-individu hasil seleksi. Seleksi dilakukan dengan memilih individu-individu yang mempunyai nilai fenotipe yang melebihi dari batas yang ditentukan. Selisih nilai tengah antara populasi hasil seleksi dengan populasi dasarnya disebut diferensial seleksi. Diferensial seleksi menunjukkan superioritas dari individu-individu yang terpilih dibandingkan dengan populasi dasarnya (Falconer dan Mackay 1996).
Marka Molekuler
Penanda molekuler merupakan suatu penanda yang mampu membedakan setiap spesies tanaman atau genotipe tanaman tanpa dipengaruhi oleh lingkungan. Potensi penggunaan marka sebagai alat untuk melakukan karakterisasi genetik tanaman telah dikenal sejak lama. Marka bisa dikategorikan sebagai marka morfologi, sitologi dan yang terbaru adalah marka molekuler (Moritz dan Hilis 1996).
Marka molekuler seringkali dikenal sebagai sidik jari DNA karena mengacu pada pita polimorfisme berupa fragmen DNA. Keunggulan utama penanda molekuler adalah (a) keakuratan tinggi dan tidak dipengaruhi lingkungan yang mempengaruhi ekspresi gen, (b) dapat diuji pada semua tingkat perkembangan tanaman, (c) pada pengujian hama dan penyakit tidak tergantung pada organisme pengganggu (d) penggunaannya pada kegiatan seleksi pemuliaan tanaman dapat mempercepat proses seleksi dan lebih hemat pada pengujian selanjutnya di lapangan (Kasim dan Azrai 2004).
10
merupakan alat bantu yang sangat akurat untuk membedakan genotipe, evaluasi kemurnian benih, pemetaan dan seleksi genotipe untuk karakter yang diinginkan; (c) studi genetik populasi dan analisis diversitas genetik (Powell et al. 1996).
Variasi jumlah pengulangan untuk suatu batasan lokus diantara genotip yang berbeda dengan mudah dapat dideteksi dengan teknik PCR (Hamada et al. 1982). Teknik PCR pada mikrosatelit hanya menggunakan DNA dalam jumlah sedikit dengan daerah amplifikasi yang kecil 100-300 bp dari genom. Selain itu marka mikrosatelit dapat diaplikasikan tanpa merusak bahan tanaman karena sample yang dibutuhkan untuk ekstraksi DNA sangat sedikit, selain itu dapat menggunakan bagian tanaman lain seperti biji atau serbuk sari (Senior et al. 1996). Produk amplifikasi hasil PCR tersebut dapat dideteksi menggunakan elektroforesis pada suatu gel dengan standar sekuen. Bila nol alel teramati maka kondisi alel tidak teramplifikasi selama PCR(Rohrer et al. 1994).
Peta genetik dan peta fisik dari sekuensing genom memudahkan pencarian marka molekuler yang secara kuat terpaut dengan suatu sifat yang menjadi target dalam pemuliaan tanaman. Pemanfaatan marka molekuler tersebut dalam seleksi materi pemuliaan tanaman disebut molecular marker assited selection (MAS). Teknik MAS memiliki kelebihan, antara lain sifatnya yang stabil dan tidak terpengaruh lingkungan. MAS dapat diujikan pada tanaman, bahkan pada saat tanaman masih muda, dan ditanam di rumah kaca maupun di lapang, tanpa terpengaruh musim. Beberapa kelebihan tersebut menyebabkan seleksi berdasarkan marka molekuler berpotensi memberikan hasil yang lebih akurat dibandngkan dengan seleksi berdasarkan fenotipe tanaman yang terpengaruh oleh musim, iklim mikro, spesifik organ, dan fase pertumbuhan tanaman (Susanto et al. 2008).
Identifikasi marka molekuler untuk MAS dapat diupayakan salah satunya melalui single marker analysis (SMA). Doerge (2002) menyatakan bahwa single marker analysis merupakan analisis segregasi fenotipe dengan marker genotipe dimana marker yang akan kita analisis apakah ada hubungan atau asosiasi pada karakter kuantitatif yang kita inginkan. Collard et al. (2005) menyatakan bahwa single marker analysis merupakan salah satu metode analisis quantitative trait loci (QTL) dengan memanfaatkan analysis of varian (ANOVA). Single marker analysis pada padi untuk toleransi cekaman suhu tinggi telah digunakan oleh Poli et al. (2013).
11
3 STUDI PEWARISAN SIFAT TOLERANSI PADI
TERHADAP CEKAMAN SUHU TINGGI
Abstrak
Perakitan varietas baru yang toleran terhadap cekaman suhu tinggi diperlukan untuk keberlanjutan produksi padi dalam antisipasi pemanasan global di masa yang akan datang. Penelitian ini bertujuan untuk mencari kriteria karakter seleksi dan memilih segregan terbaik di generasi awal berdasarkan beberapa karakter agronomi untuk adaptasi padi terhadap cekaman suhu tinggi. Penelitian dilaksanakan dari bulan September 2014 sampai bulan Januari 2015. Materi genetik yang digunakan adalah 213 individu segregant F2 yang berasal dari persilangan IPB 4S dan Situ Patenggang, 20 individu tetua IPB 4S, 20 individu tetua Situ Patenggang. Semua materi genetik ditanam dalam kondisi tercekam suhu tinggi di rumah kaca Institut Pertanian Bogor. Rata-rata suhu minimum dan maksimum pada penelitian ini adalah 24 °C dan 42.09 °C. Hasil penelitian menunjukkan bahwa karakter: jumlah anakan produktif dan total, jumlah gabah bernas, dan bobot gabah bernas memiliki nilai heritabilitas yang tinggi, aksi gen aditif, dan koefisien keragaman genetik yang tinggi. Karakter-karakter tersebut dapat dijadikan kriteria seleksi pada generasi awal untuk adaptasi padi terhadap cekaman suhu tinggi. Seleksi dapat dilakukan baik dengan satu karakter atau beberapa kriteria karakter.
12
Inheritance Trait Study for Heat Stress Tolerance in Rice
Abstract
Growing heat tolerance varieties is an essential way to sustain rice production in future anticipation of global warming. This research was aimed to find selection characters and to select the best segregant in early generation based on agronomic characters for adaptation of rice to heat stress. The experiment was carried out in September 2014 to Januari 2015. About 213 F2 segregants generated trough hybiridization between IPB 4S (sensitive parent) and Situ Patenggang (tolerant parent), 20 individuals of IPB 4S, and 20 indiviudals of Situ Patenggang, were used as genetic materials. All genetic materials were exposed to high temperature stress by growing in a green house of Bogor Agricultural University. The average temperature in the research period was 24 °C and 42.09 °C, respectively for minimum and maximum temperature. The results showed charachter: productive and total tiller number, total grain number, and seed weight have high heritability, gene action additive and high coeffiecient variability genetics. That charachters could be proposed as selection characters in early generation for rice breeding for adaptation to high temperature stress. The characters could be applied in either single trait or multiple traits selection.
13 Pendahuluan
Padi merupakan salah satu tanaman budidaya terpenting dalam peradaban manusia karena padi merupakan sumber karbohidrat utama bagi mayoritas penduduk dunia. Produksi padi dunia menempati urutan ketiga dari semua serealia setelah jagung dan gandum (Idham dan Tjahyono 1995).
Cekaman suhu tinggi menjadi salah satu penyebab pembatas dalam upaya peningkatan produktifitas padi. Menurut Taiz dan Zeiger (2002) cekaman suhu tinggi sering didefinisikan sebagai kenaikan suhu yang melebihi ambang batas kerusakan dalam suatu periode waktu yang cukup menyebabkan kerusakan yang tidak dapat balik (irreversible) pada pertumbuhan dan perkembangan tanaman. Suhu optimum untuk pertumbuhan dan perkembangan padi berkisar 27-32 °C (Yoshida 1978). Wassmann et al. (2009) menyatakan bahwa suhu optimum padi berada pada suhu sekitar 33 °C. Batasan suhu tinggi untuk tiap tanaman akan tergantung habitat asal tanaman.
Menurut laporan International Panel on Climatic Change (IPCC), rata – rata suhu global akan meningkat 0.3 °C setiap dekadenya (Jones et al. 1999), secara berturut – turut akan mencapai kurang lebih 1 dan 3 °C di atas nilai suhu sekarang pada tahun 2025 dan 2100, dan mengarah pada pemanasan global. Peningkatan suhu dapat mengakibatkan perubahan persebaran geografis dan musim tanam komoditas pertanian dengan cara menciptakan ambang batas suhu untuk awal musim dan menyebabkan kemasakan tanaman yang lebih awal (Porter 2005).
Sifat toleransi padi terhadap cekaman suhu tinggi dapat diperbaiki melalui program pemuliaan tanaman, yaitu seleksi. Keberhasilan seleksi atau efisiensi seleksi untuk meningkatkan daya hasil ditunjukkan oleh perolehan kemajuan genetik dari genotipe terpilih. Kemajuan genetik yang diperoleh sangat tergantung kepada variabilitas genetik, heritabilitas, dan korelasi genetik (Roy 2000). Kemajuan genetik dapat dimaksimalkan dengan memilih karakter seleksi yang tepat, yaitu yang memiliki nilai heritabilitas tinggi, aksi gen aditif dan koefisien keragaman genetik yang luas. Pemilihan karakter seleksi dapat dilakukan berdasarkan analisa genetik untuk memperoleh pola pewarisan dari karakter yang ingin diperbaiki.
14
Metode Penelitian Tempat dan Waktu Penelitian
Penelitian telah dilakukan di rumah kaca Kebun Percobaan University Farm IPB, Cikabayan, Dramaga, Bogor. Penelitian akan dilaksanakan bulan September 2014 sampai Januari 2015. Pengamatan pasca panen akan dilakukan di Laboratorium Penelitian Pemuliaan Tanaman Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor.
Bahan dan Alat
Bahan tanam yang digunakan adalah benih P1 (IPB 4S) ditanam sebanyak 20 individu, P2 (Situ Patenggang) ditanam sebanyak 20 individu dan F2 hasil kombinasi persilangan IPB 4S x Situ Patenggang ditanam sebanyak 213 individu, Media tanam yang digunakan adalah campuran tanah dan pupuk kandang dengan perbandingan 2:1. Pupuk dasar yang digunakan adalah Urea 250 kg/ha, SP-36 100 kg/ha, KCl 75 kg/ha. Bahan lain yang diperlukan adalah insektisida dan fungisida. Alat-alat yang digunakan adalah alat-alat pengolahan tanah, ember, bak plastik penyemaian, meteran, alat tulis, SPAD 502 digital, termometer maksimum-minimum, oven, timbangan digital, dan timbangan analitik.
Prosedur Penelitian
Penelitian telah dilaksanakan dalam beberapa tahapan kegiatan yang meliputi persiapan media tanam yaitu ember yang diisi campuran tanah dan pupuk kandang dengan perbandingan 2:1. Penyemaian yaitu benih disemai pada bak plastik di tempat persemaian dengan media tanam tanah dan pupuk kandang dengan perbandingan 1:1. Bibit hasil persemaian dipindah tanam setelah berumur 14 hari ke media tanam dalam ember. Bibit ditanam satu bibit per ember. Pupuk Urea diberikan sebanyak dua kali yaitu ½ bagian diberikan seminggu setelah tanam (MST) dan sisanya pada 3 MST. Perlakuan cekaman suhu tinggi diberikan dengan membiarkan tanaman berada di lapang pada bulan dengan suhu rata-rata diatas 35 °C.
Penyiraman dilakukan berselang yaitu mengisi air dalam ember sampai penuh dan setelah airnya kering lalu disiram kembali yang dilakukan sampai 10 hari sebelum panen. Pengendalian gulma dilakukan secara manual. Pengendalian hama dan penyakit dilakukan sesuai dengan tingkat serangan.
15 dilakukan secara manual dan penimbangan bobot 100 butir dan bobot gabah bernas dilakukan menggunakan timbangan analitik.
Pengamatan telahdilakukan pada masing-masing individu dari populasi P1, P2
dan F2. Pengamatan dilakukan pada semua tanaman untuk karakter sebagai berikut: 1) kehijauan daun 105 HST dengan SPAD Merk Minolta, 2) Tinggi tanaman saat panen, 3) Jumlah anakan total, 4) jumlah anakan produktif, 5) Panjang malai, 6) Jumlah gabah bernas dari setiap malai contoh, 7) Jumlah gabah hampa dari setiap malai contoh, 8) Jumlah gabah total dari setiap malai contoh, 9) lama pengisian biji, 10) bobot 100 butir per tanaman, 11) bobot gabah bernas per tanaman yaitu pada saat gabah kering panen.
Analisis Data
1. Analisis Nilai Tengah Karakter pada Tetua dan Populasi F2
Analisis nilai tengah karakter pada tetua dilakukan dengan membandingkan antara tetua IPB 4S dengan Situ Patenggang menggunakan uji-T. Nilai tengah populasi F2 dihitung nilai rata-ratanya dan kisaran nilai genotipe. 2. Pendugaan Aksi Gen dan Jumlah Gen
Sebaran data setiap karakter pada populasi F2 dianalisis dengan melakukan uji kenormalan dengan memanfaatkan program Minitab 14. Aksi gen diduga dengan menggunakan metode skewness (kemenjuluran) dan jumlah gen diduga dengan kurtosis (tinggi kurva).
Nilai skewness dihitung menggunakan persamaan Roy (2000): Skewness (S) = ∑��= ��−�
�− �
Uji statistik yang digunakan adalah sebagai berikut: Zs = S/SES dengan nilai
kritik Z0.05/2 = 1.96 dan Z0.01/2 = 2.58, dimana galat baku skewness sesuai dengan
persamaan Sing dan Chaudary (1979):
SES =√ � �−
�− �+ �+
Nilai kurtosis dihitung menggunakan persamaan Roy (2000): Kurtosis (K) = ∑��= ��−�
�− �
Uji statistik yang digunakan adalah sebagai berikut: Zs = S/SEK dengan
nilai kritik Z0.05/2 = 1.96 dan Z0.01/2 = 2.58, dimana galat baku kurtosis sesuai
dengan persamaan Sing dan Chaudary (1979):
SEK = 2(K) x√ n −
16
Interpretasi kemungkinan aksi gen dan jumlah gen berdasarkan Jambormias (2014) terdapat pada Lampiran 4. Jika skewness bernilai mendekati nol atau sama dengan nol maka karakter dikendalikan oleh aksi gen aditif, skewness < 0 karakter dikendalikan aksi gen aditif dengan epistasis duplikat, dan jika skewness > 0 karakter dikendalikan aksi gen aditif dengan epistasis komplementer. Jika kurtosis = 3, bentuk grafik sebaran mesokurtik. Jika kurtosis bernilai > 3, bentuk grafik sebaran data leptokurtic berarti karakter dikendalikan oleh sedikit gen. Jika sebaran populasi F2 bersifat kontinu maka karakter merupakan karakter kuantitatif atau poligenik, sedangkan jika sebaran populasi F2 bersifat tidak kontinu maka karakter tersebut bersifat kualitatif atau sedikit gen.
3. Pendugaan Komponen Ragam dan Heritabilitas
Analisa data dilanjutkan untuk menghitung ragam fenotip dan lingkungan dan serta pendugaan ragam genotip, heritabilitas, nilai koefisien keragaman genetik dan korelasi dari genotipe F2. Formula yang digunakan untuk menghitung nilai parameter genetik mengikuti formula yang dikemukakan oleh Singh dan Chaudhary (1979), sebagai berikut:
Ragam fenotipe (�� ) = � F2 = �� = �� + ��
Ragam lingkungan (��) = {�� + �� }/2 Ragam genetik (��) = �� –��
Pendugaan heritabilitas merupakan proporsi antara ragam genetik dengan ragam fenotipe yang dihitung berdasarkan rumus Allard (1960):
ℎ�� = ����
Keterangan: ℎ�� = heritabilitas arti luas, �� = ragam genetik, �� = ragam fenotipe heritabilitas dikategorikan mengikuti Stansfield (1983) nilai heritabilitas digolongkan menjadi tiga kriteria yaitu nilai heritabilitas tinggi (h2bs > 50), heritabilitas sedang (20 < h2bs < 50), dan
heritabilitas rendah (h2bs < 20).
Koefisien keragaman genetik (KKG) digunakan untuk menduga luas atau tidaknya keragaman genetik yang dimiliki masing-masing karakter, dengan rumus sebagai berikut (Singh dan Chaudhary 1979).
��� =√��
x × %
17 4. Diferensial Seleksi
Seleksi segregan terbaik berdaya hasil tinggi menggunakan intensitas seleksi 25% dianalisis menggunakan seleksi langsung dan seleksi multikarakter. Diferensial seleksi diestimasi menggunakan formula Falconer dan Mackay (1996):
S = x�− x
x x %
Keterangan: S = diferensial seleksi, x� = nilai tengah populasi terseleksi, x = nilai tengah populasi sebelum seleksi.
Seleksi multikarakter dilakukan dengan menggunakan indeks seleksi terboboti. Pemilihan karakter seleksi didasarkan pada aksi gen aditif, nilai heritabilitas yang tinggi dan koefisien keragaman genetik yang luas. Persamaan indeks seleksi sebagai berikut:
I = w1Z1 + w2Z2 + w3Z3 + ... + wnZn
Keterangan: I = nilai indeks seleksi, wn = pembobot untuk indeks seleksi, Zn = nilai amatan yang dibakukan.
Berdasarkan penelitian yang dilakukan didapatkan bahwa karakter jumlah anakan total, jumlah anakan produktif, jumlah gabah total dan bobot gabah bernas yang digunakan untuk indeks seleksi. Nilai pembobot yang digunakan dalam penelitian ini antara lain: 3 untuk karakter bobot gabah bernas, 2 untuk karakter jumlah anakan produktif dan -1 untuk karakter jumlah anakan total dan jumlah gabah total, digunakan -1 karena ada residu jumlah anakan yang tidak produktif dan jumlah gabah hampa pada kedua karakter ini.
Nilai duga kemajuan genetik diestimasi menggunakan formula Falconer dan Mackay (1996):
R = i �� h2
Keterangan: R = dugaan kemajuan genetik, i = intensitas seleksi (pada penelitian ini intensitas seleksi 25% adalah 1.265), �� = standar deviasi dari ragam fenotipe, h2 = heritabilitas.
18
Hasil dan Pembahasan Keragaan Tetua dan Populasi F2
Berdasarkan hasil penelitian yang telah dilakukan di Green House Cikabayan, University Farm IPB maka telah diperoleh informasi mengenai pertumbuhan dan produksi tetua IPB 4S dan Situ Patenggang serta diperoleh informasi pola pewarisan sifat karakter agronomi populasi F2 hasil persilangan IPB 4S dan Situ Patenggang pada kondisi tercekam suhu tinggi.
Suhu lingkungan selama penelitian mencapai 24.69 °C (suhu minimum) dan 42.33 °C (suhu maksimum) pada fase vegetatif (0-54 HST), sedangkan saat fase vegetatif akhir (55-70 HST), suhu rata-rata mencapai 24.64 °C (suhu minimum) dan 34.42 °C (suhu maksimum). Fase generatif (71 HST-panen), suhu harian minimum mencapai 27.49 °C dan suhu maksimum 38.59 °C. Awal panen suhu harian maksimum mencapai 39.57 °C. Hal tersebut mendeskripsikan kondisi suhu penelitian di wilayah Kebun Percobaan University Farm Cikabayan (Gambar 3 dan Lampiran 3). Kondisi penelitian menunjukkan tanaman tercekam suhu tinggi.
Menurut Yoshida (1978) suhu optimal untuk stadia penganakan yang optimal berada pada kisaran suhu 25-31°C, stadia antesis suhu optimum berada pada kisaran 30-33 °C dan stadia pematangan pada suhu 20-29 °C. Koesmaryono (2009) menyatakan bahwa suhu udara merupakan unsur iklim yang sangat berperan dalam memberikan lingkungan yang kondusif bagi pertumbuhan dan perkembangan tanaman. Kelembaban relatif (RH) pada fase vegetatif (0-54 HST) sebesar 64.20%, fase vegetatif akhir (55-70 HST) sebesar 71.96% dan pada fase generatif (71 HST-panen) sebesar 64.92% (Gambar 4). Kelembapan merupakan ukuran jumlah uap air yang dikandung oleh massa udara melalui proses evaporasi. Tenorio (2013) dalam penelitiannya untuk penapisan plasma nutfah toleran suhu tinggi pada padi mendapatkan kelembapan berkisar 70-75%.
Penampilan pertumbuhan dan perkembangan tanaman terlihat terganggu selama berada di rumah kaca dari awal tanam hingga menjelang fase vegetatif akhir tanaman (0-52 HST), yang ditandai dengan: pertambahan tinggi tanaman yang relatif lambat, pengeringan daun pada beberapa nomor F2, tanaman menjadi kerdil, penggulungan daun (leaf rolling) dan jumlah anakan yang cenderung sedikit. Oleh karena itu, untuk memberikan kesempatan pertumbuhan dan perkembangan yang optimal maka menjelang fase vegetatif akhir (52 HST) tanaman dikeluarkan dari rumah kaca kemudian memasuki fase generatif (72 HST) semua tanaman dimasukkan kembali ke dalam rumah kaca. Hal yang sama juga pernah dilakukan oleh Ginting (2014) dan Sasti (2014). Keragaan tanaman selama penelitian disajikan pada Gambar 2.
19 pertumbuhan. Hal ini dilakukan dalam upaya antisipasi terhadap pengaruh pemanasan global di masa yang akan datang, terlebih lagi Indonesia sebagai negara tropis, dimana cekaman suhu tinggi pada negara tropis dialami tanaman dari semenjak tanaman berada dilapangan. Koesmaryono (2009) menyatakan bahwa pada musim kering di Indonesia suhu rata-rata harian bisa mencapai di atas 33 °C, terutama daerah seperti Nusa Tenggara Timur akan mencapai suhu sekitar 35-40 °C.
A B
C D
E F
20
Gambar 3 Suhu minimum dan maksimum selama penelitian
Gambar 4 Kelembapan udara selama penelitian
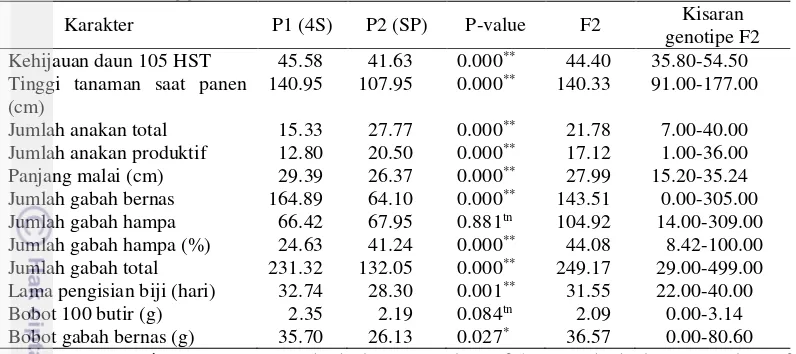
Berdasarkan hasil analisa data diketahui bahwa nilai tengah karakter jumlah anakan produktif, jumlah anakan total, kehijauan daun 105 HST, lama pengisian biji, tinggi tanaman saat panen, panjang malai, jumlah gabah bernas, jumlah gabah hampa, jumlah gabah hampa (%), jumlah gabah total dan bobot gabah bernas antara kedua tetua menunjukkan berbeda nyata kecuali pada karakter bobot 100 butir dan jumlah gabah hampa malai-1. Jumlah gabah hampa malai-1 menunjukkan tidak berbeda nyata karena yang dibandingkan adalah jumlah gabah hampanya, akan tetapi jika berdasarkan persentase gabah hampa maka kedua tetua ini berbeda nyata.
Tabel 2 memperlihatkan bahwa nilai tengah populasi F2 untuk karakter agronomi hasil yang meliputi jumlah gabah bernas malai-1 dan bobot gabah bernas tanaman-1, melebihi nilai tengah dari kedua tetua. Hal ini berarti terdapat
21 4S : IPB 4S; SP : Situ Patenggang; * = berbeda nyata pada taraf 5%; ** = berbeda nyata pada taraf 1% dan tn = tidak nyata pada taraf 5% berdasarkan uji-t antara tetua IPB 4S dan Situ Patenggang; HST= Hari Setelah Tanam; F2= nilai rata-rata populasi F2
Nilai tengah karakter agronomi tetua IPB 4S pada penelitian ini terlihat lebih unggul pada hampir semua karakter dibandingkan tetua Situ Patenggang. Hal ini di duga karena varietas IPB 4S merupakan varietas unggul baru yang berdaya hasil hasil tinggi namun sangat peka terhadap cekaman suhu tinggi berdasarkan hasil penapisan yang dilakukan oleh Mubarrozah (2013) dan Wirnas et al. (2015a), sementara varietas Situ Patenggang merupakan varietas yang toleran terhadap cekaman suhu tinggi namun berdaya hasil rendah.
Berdasarkan hasil penapisan yang di lakukan oleh Mubarrozah (2013) menunjukkan bahwa pada kondisi normal varietas IPB 4S menghasilkan bobot gabah bernas sebesar 59.1 g dan kondisi tercekam suhu tinggi 33.4 g, sementara varietas Situ Patenggang pada kondisi normal menghasilkan bobot gabah bernas sebesar 34.5 g dan pada kondisi tercekam suhu tinggi 33.6 g. Tenorio (2013) melakukan penapisan terhadap sekitar 511 varietas dan galur serta 455 aksesi dari IRRI dalam empat musim tanam untuk mencari sumber daya genetik yang toleran terhadap suhu tinggi. Hasil penelitian tersebut didapatkan varietas Milyang23 dan IR2006-P12-12-2-2 toleran pada saat fase pembungan dan Giza178 toleran pada fase pengisian gabah.
22
Aksi Gen yang Mengendalikan Karakter Pertumbuhan dan Hasil
Karakter kuantitatif tanaman dalam suatu populasi sangat berhubungan erat dengan pertumbuhan dan hasil tanaman. Karakter kuantitatif merupakan karakter yang sangat dipengaruhi oleh lingkungan. Karakter kuantitatif dikendalikan oleh banyak gen dan pengaruh masing-masing gen terhadap penampilan karakter (fenotipe) lebih kecil dan bersifat aditif, walaupun gen-gen tersebut secara bersama-sama mempunyai pengaruh yang lebih besar dari pengaruh lingkungan (Fehr 1987).
Menurut Roy (2000) karakteristik pola sebaran karakter kuantitatif dapat dijelaskan oleh nilai tengah, median, range, ragam, standar deviasi, standar eror, skewness, dan kurtosis. Statistik deskriptif tersebut dapat digunakan untuk menduga jumlah gen dan aksi gen yang mengendalikan karakter tersebut di populasi bersegregasi.
Analisis sebaran data pada populasi F2 bertujuan untuk menduga sifat suatu karakter apakah kualitatif (dikendalikan oleh gen-gen mayor) atau kuantitatif (dikendalikan oleh gen-gen minor atau poligenik), aditif, atau ada tidaknya epistasis. Jika sebaran data populasi F2 menghasilkan grafik yang membentuk sebaran kontinyu, menyebar normal, dan satu puncak maka karakter tersebut dikendalikan oleh gen-gen minor. Jika sebaran data populasi F2 menghasilkan grafik yang membentuk sebaran kontinyu, tetapi tidak menyebar normal, dengan lebih dari satu puncak maka karakter tersebut dikendalikan oleh gen-gen minor dan satu atau dua gen mayor. Jika grafik sebaran data populasi F2 tidak kontinyu dan data tidak menyebar normal, diduga bahwa karakter tersebut dikendalikan oleh gen-gen mayor.
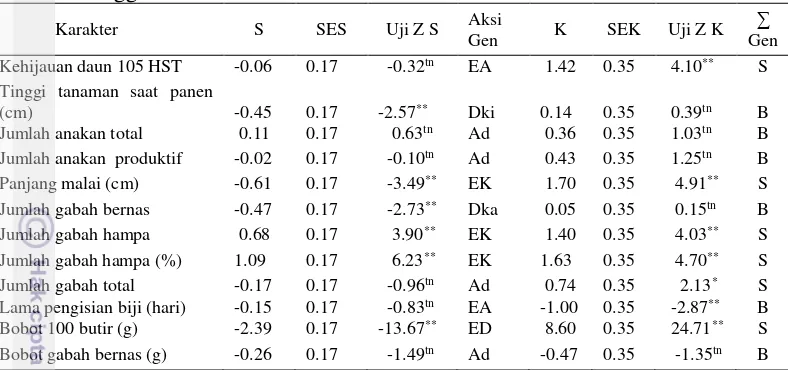
Analisis sebaran pada karakter agronomi yang diamati terdapat pada Gambar 5 sampai 10 serta nilai skewness dan kurtosis untuk karakter agronomi yang diamati ditampilkan pada Tabel 3. Berdasarkan Gambar 5 hingga Gambar 10 memperlihatkan bahwa terdapat segregan transgesif pada populasi F2. Hal ini diduga terjadi karena pada populasi F2 keberadaan segregan transgesif akibat dikendalikan oleh gen-gen aditif dan terdapat pengaruh epistasis komplementer, aditif, duplikat dan pengaruh aksi gen dominansi. Keberadaan segregan transgesif untuk menjadi bahan seleksi sangat menguntungkan karena nilai segregan transgesif yang diperoleh melebihi nilai tengah kedua tetua.
Hasil uji Z skewness dan kurtosis untuk karakter jumlah anakan produktif, jumlah anakan total, bobot gabah bernas berbeda tidak nyata. Hal ini menunjukkan bahwa, jumlah anakan produktif, jumlah anakan total, jumlah gabah total dan bobot gabah bernas memiliki pola sebaran mendekati normal dan dikendalikan banyak gen dengan aksi gen aditif dan tidak terdapat pengaruh gen-gen mayor (Tabel 3). Situmorang (2015) dan Jaisyurahman (2015) melaporkan aksi gen aditif untuk karakter jumlah gabah total dan jumlah anakan 45 HST pada tanaman padi.
23 Tabel 3 Pendugaan aksi gen dan jumlah gen karakter agronomi populasi F2 hasil persilangan IPB 4S dan Situ Patenggang pada kondisi tercekam suhu taraf 5%; **= sangat nyata pada taraf 1%; tn= tidak nyata; HST=Hari Setelah Tanam; F2=nilai rata-rata populasi F2
Skewness merupakan ukuran kemenjuluran kurva, sedangkan kurtosis merupakan ukuran kegemukan kurva. Jika nilai skewness mendekati nol atau sama dengan nol, dapat diduga bahwa aksi gennya bersifat aditif. Jika skewness bernilai negative, maka terdapat aksi gen aditif dengan pengaruh epistasis duplikat, sedangkan jika skewness bernilai positif maka karakter tersebut dikendalikan oleh aksi gen aditif dengan pengaruh epistasis komplementer. Pendugaan nilai kurtosis digunakan untuk menduga jumlah gen pengendalinya, jika kurtosis bernilai negatif sebarannya disebut leptokurtik karakter tersebut dikendalikan oleh sedikit gen, sedangkan jika kurtosis bernilai positif sifat yang diteliti dikendalikan oleh banyak gen (polygenic) (Roy 2000).
Karakter panjang malai, jumlah gabah hampa, persentase jumlah gabah hampa, dan bobot 100 butir memiliki pola sebaran data tidak normal karena hasil uji Z skewness dan kurtosis menunjukkan perbedaan yang sangat nyata. Hal ini berarti karakter-karakter tersebut selain dikendalikan oleh aksi gen interaksi alel dari lokus yang berbeda (epistasis).
24
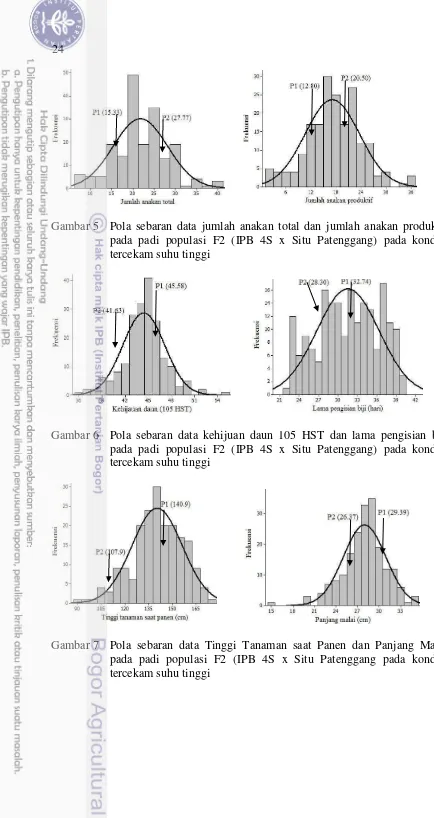
Gambar 5 Pola sebaran data jumlah anakan total dan jumlah anakan produktif pada padi populasi F2 (IPB 4S x Situ Patenggang) pada kondisi tercekam suhu tinggi
Gambar 6 Pola sebaran data kehijuan daun 105 HST dan lama pengisian biji pada padi populasi F2 (IPB 4S x Situ Patenggang) pada kondisi tercekam suhu tinggi
25
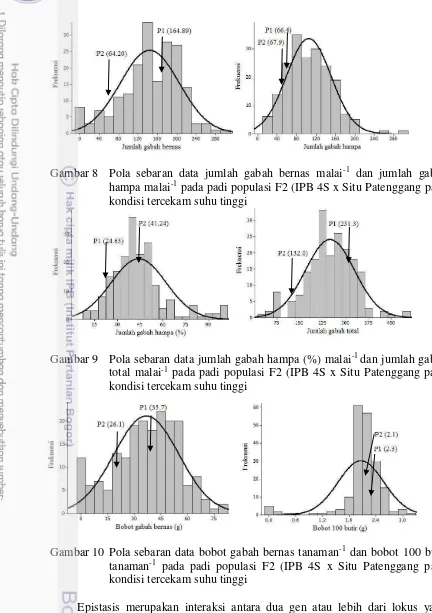
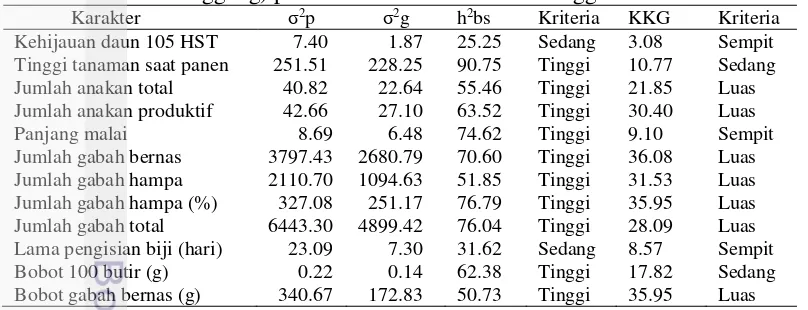
Gambar 8 Pola sebaran data jumlah gabah bernas malai-1 dan jumlah gabah hampa malai-1 pada padi populasi F2 (IPB 4S x Situ Patenggang pada kondisi tercekam suhu tinggi
Gambar 9 Pola sebaran data jumlah gabah hampa (%) malai-1 dan jumlah gabah total malai-1 pada padi populasi F2 (IPB 4S x Situ Patenggang pada kondisi tercekam suhu tinggi
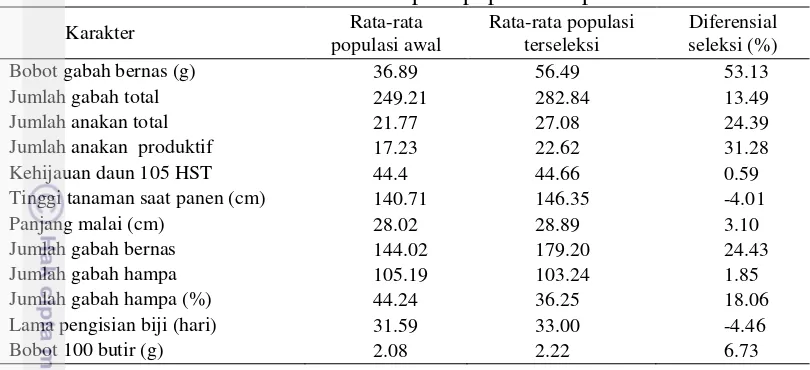
Gambar 10 Pola sebaran data bobot gabah bernas tanaman-1 dan bobot 100 butir
tanaman-1 pada padi populasi F2 (IPB 4S x Situ Patenggang pada kondisi tercekam suhu tinggi
26
hanya jika dua gen menghasilkan bahan yang sama untuk membentuk fenotipe yang sama (Griffiths et al. 2005).
Pendugaan Nilai Ragam Genetik dan Heritabilitas
Keragaman genetik terdiri atas ragam genetik aditif, dominan, dan epistasis. Ragam genetik aditif adalah ragam genetik yang menyebabkan terjadinya kesamaan sifat diantara tetua dan turunannya. Heritabilitas menunjukkan kemampuan suatu sifat diwariskan dari tetua ke keturunannya. Peningkatan keragaman genetik terjadi dengan membentuk famili-famili homozigot. Pendugaan komponen ragam dan heritabilitas untuk mengetahui proporsi keragaman yang disebabkan oleh faktor genetik dan lingkungan. Nilai heritabilitas sangat dipengaruhi oleh lingkungan (Roy 2000).
Hasil analisa data pada Tabel 4 menunjukkan bahwa pada populasi F2 karakter jumlah anakan produktif, jumlah anakan total, tinggi tanaman saat panen, panjang malai, jumlah gabah bernas, jumlah gabah hampa, jumlah gabah total, persentase jumlah gabah hampa, bobot gabah bernas, bobot 100 butir, memiliki nilai heritabilitas arti luas tergolong tinggi, sedangkan untuk karakter kehijauan daun 105 HST, lama pengisian biji, memiliki nilai heritabilitas arti luas tergolong sedang.
Heritabilitas arti luas (broad sense heritability) (h2
bs) didefinisikan nisbah
ragam genotipe terhadap ragam fenotipe. Heritabilitas merupakan salah satu alat ukur yang banyak digunakan dalam pemuliaan tanaman. Secara sederhana, istilah heritabilitas adalah suatu perbandingan antara besaran ragam genotipe terhadap besaran total ragam fenotipe dari suatu karakter (Bari et al. 1982). Fenotipe merupakan interaksi antara genetik dan lingkungan, ini berarti besaran fenotipe sebagian ditentukan oleh pengaruh genetik dan sebagian oleh pengaruh lingkungan (Poespodarsono 1988).
Tabel 4 Nilai pendugaan komponen ragam, heritabilitas arti luas dan Koefisien Keragaman Genetik (KKG) populasi F2 (hasil persilangan IPB 4S dan Situ Patenggang) pada kondisi tercekam suhu tinggi
Karakter σ2p σ2g h2bs Kriteria KKG Kriteria
27 nilai 0, berarti pekerjaan seleksi tidak akan banyak berarti sehingga seleksi harus dilakukan pada generasi lanjut (Poespodarsono 1988).
Apabila suatu karakter memiliki nilai heritabilitas arti luas tinggi, maka karakter tersebut potensial untuk diwariskan pada generasi awal dan dapat dijadikan sebagai kriteria seleksi untuk adaptasi padi terhadap cekaman suhu tinggi karena karakter tersebut lebih respon terhadap seleksi. Hal ini sesuai dengan pendapat Syukur dan Sobir (2015) yang mengatakan bahwa karakter yang memiliki nilai heritabilitas sedang sampai tinggi maka karakter tersebut lebih banyak dikendalikan oleh faktor genetik dibandingkan dengan faktor lingkungan.
Tabel 4 menunjukkan nilai KKG pada masing-masing karakter. Kisaran nilai KKG pada karakter yang di uji antara 3.08 – 36.08. Nilai KKG terendah terdapat pada karakter kehijauan daun 105 HST sebesar 3.08%, sedangkan nilai KKG tertinggi terdapat pada jumlah gabah bernas sebesar 36.08%. Stanfield (1983) menerangkan bahwa keragaman genetik yang tinggi menunjukkan adanya pengaruh genetik yang lebih dominan daripada pengaruh lingkungan, begitu pula sebaliknya bahwa keragaman genetik yang rendah menunjukkan adanya pengaruh yang dominan dari lingkungan. Lebih lanjut Febrianto et al. (2015) menyatakan bahwa nilai KKG yang tinggi menunjukkan suatu peluang terhadap usaha-usaha perbaikan yang efektif melalui seleksi sehingga seleksi terhadap karakter kuantitatif dapat dilakukan tanpa mengabaikan nilai tengah populasi yang bersangkutan.
Keragaman genetik adalah perbedaan penampakan yang muncul dari berbagai tanaman yang diuji. Adapun hal yang mempengaruhi keragaman genetik pada generasi selanjutnya dipengaruhi oleh faktor genetik, faktor lingkungan dan interaksi genetik x lingkungan (G x E). Keragaman genetik yang diturunkan merupakan salah satu indikator dalam mementukan keberhasilan suatu program pemuliaan tanaman. Hal ini digunakan untuk mengetahui sifat-sifat kuantitatif yang diharapkan muncul pada keturunannya (Allard 1960). Komponen ragam terdiri dari ragam lingkungan, ragam genetik, dan ragam fenotip. Hasil penelitian menunjukkan bahwa heritabilitas karakter yang diuji memiliki kisaran nilai antara 28.67% - 90.75% (Tabel 4). Stansfield (1983) menyatakan bahwa heritabilitas adalah proporsi varians fenotipik total yang disebabkan oleh semua tipe efek gen. nilai heritabilitas digolongkan menjadi tiga kriteria yaitu nilai heritabilitas tinggi (h2 > 50), heritabilitas sedang (20 < h2 < 50), dan heritabilitas rendah (h2 < 20).
Diferensial Seleksi Langsung
28
pada tanaman padi. Penggunaan diferensial seleksi langsung dengan menggunakan bobot gabah bernas pada tanaman padi dilaporkan antara lain oleh Jaisyurahman (2015) dan Situmorang (2015).
Tabel 5 Diferensial seleksi berdasarkan karakter bobot gabah bernas pada populasi F2 padi
Tinggi tanaman saat panen 140.71 147.92 -5.12
Jumlah anakan total 21.77 24.04 10.43
Jumlah anakan produktif 17.12 24.96 31.41
Panjang malai 28.02 29.07 3.75
Jumlah gabah bernas 144.02 193.42 34.30
Jumlah gabah hampa 105.19 116.56 10.81
Jumlah gabah hampa (%) 44.24 37.01 16.34
Jumlah gabah total 249.21 309.98 24.39
Lama pengisian biji (hari) 31.59 33.86 -7.19
Bobot 100 butir (g) 2.08 2.26 8.65
Tabel 5 menunjukkan nilai diferensial seleksi langsung berdasarkan karakter bobot gabah bernas. Seleksi langsung dengan intensitas 25% akan memberikan perbaikan nilai tengah bobot gabah bernas sebesar 58.39%. Seleksi langsung pada penelitian ini mempunyai keunggulan yaitu meningkatnya nilai tengah jumlah anakan produktif, jumlah anakan total, kehijauan daun 105 HST, panjang malai, jumlah gabah bernas, jumlah gabah total dan bobot 100 butir, sedangkan kelemahannya adalah meningkatnya tinggi tanaman sebesar 5.12%, lama pengisian biji sebesar 7.19% dan jumlah gabah hampa sebesar 10.81% yang hal ini tidak diinginkan pemulia. Keragaan 50 segregan terbaik dengan menggunakan karakter bobot gabah bernas disajikan pada Lampiran 5.
Diferensial Seleksi Multikarakter Indeks Terboboti
29 jumlah gabah hampa pada kedua karakter ini. Hasil nilai indeks seleksi pada 50 genotipe terbaik disajikan pada Lampiran 6.
Tabel 6 Diferensial seleksi multikarakter pada populasi F2 padi
Karakter Rata-rata
Tinggi tanaman saat panen (cm) 140.71 146.35 -4.01
Panjang malai (cm) 28.02 28.89 3.10
Tabel 6 menunjukkan nilai diferensial seleksi multikarakter berdasarkan karakter bobot gabah bernas, jumlah gabah total, jumlah anakan total dan jumlah anakan produktif. Seleksi dengan menggunakan 4 karakter seleksi dan intensitas seleksi 25% akan memberikan perbaikan nilai tengah bobot gabah bernas sebesar 53.13%, menurunkan jumlah gabah hampa sebesar 1.85%, dan jumlah gabah hampa (%) sebesar 18.06% dimana karakter jumlah gabah hampa dan jumlah gabah hampa (%) yang sedikit berarti bagi pemulia tanaman dan memberikan perbaikan terhadap bobot 100 butir sebesar 6.73%. Seleksi multikarakter pada penelitian ini mempunyai keunggulan yaitu meningkatnya nilai tengah jumlah anakan produktif, jumlah anakan total, panjang malai, jumlah gabah bernas, dan jumlah gabah total sedangkan kelemahannya adalah meningkatnya tinggi tanaman sebesar 4.01%, dan lama pengisian biji sebesar 4.46% yang hal ini tidak diinginkan pemulia. Seleksi berdasarkan bobot gabah bernas per tanaman dan seleksi multikarakter (jumlah anakan produktif, jumlah anakan total, jumlah gabah total, dan bobot gabah bernas) terseleksi 50 segregan F2 terbaik berdaya hasil tinggi pada kondisi cekaman suhu tinggi. Keragaan 50 segregan terbaik dengan menggunakan multikarakter disajikan pada Lampiran 6.
Kemajuan Genetik dan Dugaan Respon Seleksi