KARAKTERISASI ENZIM HIDROLASE BAKTERI DARI
MATA AIR SODA PARBUBU, TAPANULI UTARA
SRI ASIH
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biokimia
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
SRI ASIH. Karakterisasi Enzim Hidrolase Bakteri dari Mata Air Soda Parbubu,
Tapanuli Utara. Dibimbing oleh I MADE ARTIKA dan RASTI SARASWATI.
ABSTRACT
SRI ASIH. Characterization of Enzymes from Bacteria Hidrolase Parbubu Soda
Springs, North Tapanuli. Under the direction of I MADE A RTIKA and RASTI
SARASWATI.
Judul Skripsi
: Karakterisasi Enzim Hidrolase Bakteri dari Mata Air Soda
Parbubu, Tapanuli Utara
Nama
: Sri Asih
NIM
: G84062071
Disetujui,
Komisi Pembimbing
Dr. Ir. I Made Artika, M. App. Sc.
Dr. Rasti Saraswati
Ketua
Anggota
Diketahui
Ketua Departemen Biokimia
Dr. Ir. I Made Artika, M.App.Sc
Ketua Departemen Biokimia
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT, yang telah
melimpahkan karunia-Nya, sholawat serta salam semoga tercurahkan kepada
Rasulullah Muhammad SAW beserta keluarga dan orang-orang yang berjuang
menegakkan ajaran agama-Nya. Penelitian yang dilakukan berjudul Karakterisasi
Potensi Enzim Hidrolase Bakteri dari Mata Air Soda Parbubu, Tapanuli Utara.
Karya ilmiah ini disusun berdasarkan hasil penelitian yang dilakukan
di
Laboratorium Mikrobiolgi dan Kesehatan Tanah, Balai Penelitian Tanah, Bogor
pada bulan Maret sampai dengan Juli 2010.
Penulis mengucapkan terima kasih kepada semua pihak yang telah
membantu dalam penyusunan karya ilmah ini. Ucapan terima kasih penulis
sampaikan terutama kepada Dr. Ir. I Made Artika, M. App. Sc. Selaku
pembimbing
dan Dr. Rasti Saraswati selaku pembimbing sekaligus mendanai
penelitian ini. Ungkapan terima kasih yang tak terhingga penulis ucapkan kepada
orang tua dan keluarga atas segala doa, dukungan, dan kasih sayang yang telah
diberikan. Terima kasih penulis sampaikan pula kepada teman-teman Al Iffah, Al
Hurriyyah, ISC, dan Biokimia 43 atas motivasi dan dukungannya.
Penulis mengharapkan kritik dan saran yang membangun dari berbagai
pihak sebagai bahan masukan di kemudian hari. Semoga karya tulis ini dapat
berguna bagi penulis sendiri maupun semua pihak yang membutuhkannya demi
perkembangan dan kemajuan ilmu pengetahuan dan teknologi.
Bogor, Februari 2011
RIWAYAT HIDUP
Penulis dilahirkan di Malang pada tanggal 25 Desember 1989 dari bapak
Tukiran dan ibu Lisiati. Penulis merupakan anak kedua dari dua bersaudara.
Tahun 2006 penulis lulus dari SMA Negeri 1 Gondanglegi Kabupaten
Malang dan pada tahun yang sama lulus seleksi masuk IPB melalui jalur
Undangan Seleksi Masuk IPB (USMI) pada program mayor minor. Penulis
memilih Mayor Biokimia Fakultas Matematika dan Ilmu Pengetahuan Alam dan
Minor Teknologi Pangan.
DAFTAR ISI
Halaman
DAFTAR GAMBAR ... vii
DAFTAR LAMPIRAN ... vii
PENDAHULUAN ... 1
TINJAUAN PUSTAKA
Mata Air Soda Parbubu... 2
Enzim Hidrolase ... 2
Kurva Pertumbuhan Bakteri ... 4
Aktivitas Enzim ... 5
BAHAN DAN METODE
Alat dan Bahan ... 5
Metode ... 5
HASIL DAN PEMBAHASAN
Isolasi bakteri dari Mata Air Soda Parbubu ... 8
Aktivitas Enzim Hidrolase secara Semikuantitatif ... 8
Waktu Optimum Produksi Enzim Lipase ... 9
Aktivitas Enzim Lipase... 10
Karakterisasi Enzim Lipase ... 12
SIMPULAN DAN SARAN
Simpulan ... 13
Saran ... 13
DAFTAR PUSTAKA ... 13
DAFTAR GAMBAR
Halaman
1 Sumber mata air soda Parbubu ... 2
2 Struktur enzim selulase ... 3
3 Struktur enzim protease ... 3
4 Struktur enzim lipase ... 4
5 Pola pertumbuhan bakteri ... 5
6 Hasil uji semikuantitatif selulase pada isolat P10... 8
7 Hasil uji semikuantitatif protease pada isolat P9 ... 9
8
Hasil uji semikuantitatif lipase pada (a) isolat P3, (b) isolat P9, dan (c)
isolat P23... 9
9 Kurva pertumbuhan dan aktivitas lipase bakteri P9 ... 10
10 Reaksi hidrolisis enzimatis spesifik pada minyak zaitun... 11
11 Aktivitas lipase pada beberapa fraksi enzim... 12
12 Penentuan pH optimum ... 12
DAFTAR LAMPIRAN
Halaman
1 Bagan alir penelitian Sumber mata air soda Parbubu ... 17
2 Isolasi bakteri dari sumber mata air soda Parbubu ... 18
3 Uji potensi enzim hidrolase bakteri ... 19
4 Pembuatan kurva pertumbuhan bakteri lipolitik... 20
5 Uji aktivitas enzim lipase dengan metode titrimetrik (Paskevicius 2001)... 21
6 Karakterisasi enzim hidrolitik (Sari 2008)... 22
7 Isolat bakteri dari mata air soda Parbubu, Tapanuli Utara ... 23
8 Perhitungan Aktivitas Lipase... 26
9 Hasil uji kuantitatif aktivitas lipase pada ekstrak kasar lipase... 27
10 Aktivitas lipase pada fraksi lipase dengan pengendapan ammonium sulfat.... 28
11 Aktivitas lipase pada suhu 37°C dan berbagai pH... 29
PENDAHULUAN
Indonesia merupakan salah satu negara dengan sumber biodiversitas dunia karena memiliki 42 ekosistem daratan alami dan lima ekosistem lautan. Keragaman ekosistem tersebut membuat Indonesia menjadi tempat yang sangat potensial untuk eksplorasi berbagai jenis organisme. Salah satu bidang yang belum mendapat perhatian serius adalah keanekaragaman hayati mikrob. Pendekatan yang mungkin dilakukan adalah mengeksplorasi sebanyak-banyaknya mikroorganisme.
Pulau Sumatera merupakan pulau dengan dominasi batuan vulkanik, berupa batuan beku, piroklastik, dan breksi vulkanik. Dominasi batuan ini menjadi salah satu penyebab terdapatnya sumber panas bumi di Pulau Sumatera (Hasan et al. 2006). Morfologi Pulau Sumatera, terutama daerah Tapanuli Utara, sebagai daerah vulkanik ini mengakibatkan adanya berbagai macam sumber mata air tanah yang berasal dari daerah dataran rendah. Mata air tanah yang muncul antara lain mata air panas dan mata air soda. Mata air soda terdapat pada sumber mata air soda Parbubu. Mata air soda dengan kondisi lingkungan yang ekstrim ini menjadikannya tempat yang potensial untuk mengeksplorasi berbagai jenis mikrob. Kondisi lingkungan yang beraneka ragam pada daerah vulkanik tersebut menyebabkan terdapat beragam mikroorganisme yang mampu tumbuh pada kondisi ekstrim seperti suhu, pH, dan konsentrasi garam yang tinggi (Madigan & Mars 1997).
Sumber mata air soda yang memiliki kandungan karbonat tinggi ini menjadi pelengkap keanekaragaman biota. Keragaman mikrob yang terdapat pada sumber mata air soda sangat menarik untuk dikaji karena mikrob, terutama bakteri mampu bertahan hidup pada kondisi air yang mengandung karbonat.
Potensi yang dimiliki oleh mata air soda Parbubu ini belum dikaji, bahkan pemanfaatannya hanya sebagai pemandian saja. Oleh karena itu, diperlukan kajian yang lebih mendalam terhadap potensi yang dimiliki oleh mata air soda ini agar dapat dimanfaatkan secara tepat. Untuk dapat memanfaatkan potensi yang tersimpan tersebut diperlukan upaya-upaya untuk mengeksplorasi kekayaan mikrob yang terdapat di dalamnya. Upaya untuk mendapatkan mikrob, terutama bakteri yang terdapat di mata air soda tersebut dilakukan
dengan cara mengisolasi, memurnikan, dan mengidentifikasi potensi enzimatik yang dimiliki isolat, serta optimasi potensi bakteri. Analisis ini menjadi sangat penting untuk mengetahui besarnya potensi yang dimiliki oleh bakteri dalam air soda.
Keistimewaan isolat bakteri ini adalah dapat hidup pada kondisi lingkungan dengan kadar karbonat yang tinggi, yang mengakibatkan air menjadi sadah dan turunnya pH air menjadi asam (DITA 2003). Kesadahan air yang tinggi dan pH yang relatif rendah dapat menghambat organisme untuk hidup. Kondisi ekstrim air soda ini memungkinkan hidupnya bakteri yang memiliki ketahanan yang tinggi pula pada kondisi ekstrim yang dibutuhkan saat proses industri.
Pengkajian terhadap potensi enzimatik bakteri ini juga diperlukan untuk memanfaatkan bakteri sebagai pabrik biologis. Bakteri yang dapat tumbuh pada lingkungan ekstrim dapat menghasilkan metabolit, salah satunya enzim, yang memiliki ketahanan terhadap kondisi ekstrim pula. Enzim-enzim yang mampu bertahan dan aktif pada kondisi ekstrim sangat diperlukan dalam dunia industri berbasis enzim. Penggunaan enzim yang mampu bertahan pada suhu tinggi dalam bidang bioteknologi dapat menurunkan biaya operasi dan meningkatkan kecepatan reaksi
Enzim yang diujikan aktivitasnya antara lain protease, lipase, dan selulase. Pemilihan ketiga jenis enzim dimaksudkan untuk mendegradasi makromolekul utama yang terdapat banyak di alam, sehingga cakupan aplikasi enzim yang diproduksi bakteri ini dapat lebih luas. Aplikasi penggunaan enzim dalam industri selanjutnya disesuaikan dengan potensi mikrob yang diperoleh saat analisis.
Tujuan penelitian ini adalah mengisolasi bakteri dari mata air soda Parbubu dan menguji aktivitas enzim hidrolasenya, yaitu lipase, protease, dan selulase serta mengkarakterisasi enzim hidrolase terbaik pada kondisi suhu dan pH tertentu.
Menurut Komisi Tata International Union of Bioch Molecular Biologist, protease dalam kelompok III subkelompo (E.C.3.4), yaitu golongan hidrola substrat protein. Terdapat beb klasifikasi protease yang digunaka Hartley (1960) (dalam Widya mengklasifikasikan protease ber sifat kimia sisi aktif enzim, yait serin (memiliki asam amino serin aktifnya), protease sulfhidril (mem sulfhidril pada sisi aktifnya), pro (memiliki ion logam pada sisi ak protease asam (memiliki dua gugu pada sisi aktifnya).
Enzim Lipase (EC 3.1)
Enzim yang bekerja dalam lemak dan minyak dapat dike menjadi dua kelompok besar, yait esterase (Winarno 1985). Lipas golongan enzim dengan fungs karena dapat menghidrolisis ikata lemak sehingga terbentuk asam le dan gliserol. Lipase juga termasu enzim esterase, sebab dapat me ikatan ester. Selain itu, lipase enzim yang dapat memutuskan i panjang asam lemak ester dan me ilmiah triasilgliserol asilhidrolase 2007).
Enzim lipase atau asilgliser (E.C 3.1.1.3) (Gambar 4) merup yang dapat menghidrolisis ran trigliserida. Enzim ini memiliki po digunakan memproduksi asam le merupakan prekursor berbagai ind Produksi asam lemak secar menggunakan katalis kimia m efek samping bagi lingkungan. enzim lipase telah banyak diken cakupan aplikasi yang amat bidang bioteknologi, seperti pestisida, pengolahan limbah makanan, biosensor, deterjen, untu
Gambar 4 Struktur enzim lip
Tata Nama iochemist and ase termasuk pok IV enzim olase pemecah eberapa cara akan para ahli. yastuti 2000) berdasar sifat-yaitu protease erin pada sisi emiliki gugus protease metal i aktifnya), dan ugus karboksil
lam hidrolisis ikelompokkan aitu lipase dan ipase termasuk gsi hidrolase atan ester pada lemak bebas suk kelompok menghidrolisis se merupakan n ikatan rantai memiliki nama lase (Tikaet al
serol hidrolase upakan enzim antai panjang i potensi untuk lemak, yang industri kimia. cara industri menghasilkan n. Selain itu, enal memiliki t luas dalam ti biomedikal, ah, industri ntuk industri
lipase.
kulit dan industri oleokimia (mempro asam lemak dan turunannya) (Macra 1983).
Kurva Pertumbuhan Bakteri
Pertumbuhan merupakan penam secara teratur semua komponen sel jasad. Pertumbuhan dapat diartikan se penambahan jumlah sel. Penambahan ju sel pada jasad bersel banyak (multise tidak menghasilkan penambahan ju individunya, melainkan terbent jaringan baru atau peningkatan ukuran jasad. Penambahan sel pada jasad berse (uniseluler) berarti penambahan ju individu, seperti pertumbuhan yang pada suatu kultur mikrob.
Mikrob jika dimasukkan pada baru yang sesuai akan tumbuh memperbanyak diri. Perbanyakan dilakukan melalui pembelahan biner, a satu sel akan membelah diri menjadi d yang identik. Sel tersebut kemudian m masing membelah diri lagi menjadi du seterusnya. Pertumbuhan seperti ini ter dalam tipe pertumbuhan ekspon (Sokatch 1969). Pada kondisi yang s beberapa jenis bakteri dapat mem menjadi dua sel baru atau memb generasi baru dalam 15 menit, seda pada kondisi yang tidak sesuai, dipe waktu hingga 24 jam atau lebih. Waktu diperlukan untuk membelah diri dari sa menjadi dua sel sempurna disebut regenerasi (Fosteret al. 1961).
Kurva pertumbuhan diperoleh m perbandingan jumlah bakteri yang di pada waktu-waktu tertentu seh menunjukkan pola pertumbuhan b tersebut. Pola pertumbuhan bakteri umumnya memiliki empat fase pertumb yaitu fase permulaan atau adaptasi (fas fase pertumbuhan eksponensial logaritma (fase log), fase stasioner, da kematian.
Fase pertumbuhan awal atau fa merupakan awal pertumbuhan mikrob bakteri baru menyesuaikan diri d lingkungannya. Fase ini ditandai d perubahan komposisi kimiawi penambahan ukuran, substansi seluler waktu generasinya relatif panjang. selanjutnya adalah fase pertum eksponensial atau fase log yang meru fase pertumbuhan sel paling cepat dan aktif. Pada fase ini, metabolisme sel aktif, sintesis bahan sel sangat cepat d
produksi crae AR
ri
ambahan el suatu sebagai n jumlah tiseluler) jumlah entuknya ran suatu rsel satu jumlah g terjadi
a media uh dan an diri r, artinya i dua sel masing-dua, dan termasuk onensial g sesuai, embelah mbentuk dangkan iperlukan ktu yang i satu sel t waktu
melalui dihitung sehingga bakteri teri pada mbuhan, fase lag), l atau dan fase
pertama/ 10-1). Pengenceran terus dilakukan sampai pengenceran keenam atau diperoleh konsentrasi 10-6). Hasil pengenceran diambil sebanyak 0.1 mL dan dimasukkan ke dalam cawan petri, lalu ditambah dengan media hangat yang belum memadat sebanyak 20 mL. Media yang ditambahkan ada dua macam, yaitu media peptone glucose agar (PGA) dan potato dextrose agar (PDA), sehingga diperoleh koloni yang berbeda. Selanjutnya cawan digoyang perlahan untuk homogenisasi, lalu dibiarkan memadat. Cawan selanjutnya dimasukkan dalam plastik dalam posisi terbalik dan disimpan dalam inkubator suhu 37ºC selama dua sampai lima hari (Nurkanto 2007; Ilyas 2007).
Setelah masa inkubasi, pertumbuhan bakteri dalam cawan diamati. Bakteri yang tumbuh dalam cawan tersebut bersifat heterogen, sehingga dilakukan pemurnian dengan cara memindahkan masing-masing bakteri yang tumbuh dengan ciri morfologi berbeda ke dalam media dalam cawan baru dan selanjutnya dipindahkan pada agar miring. Pemindahan dilakukan dengan cara mengambil sebanyak satu ose biakan dari cawan lalu dipindahkan ke agar miring dengan goresan zig-zag dari arah pangkal agar miring hingga ujung. Pemindahan ini dilakukan secara aseptik. Isolat yang telah murni selanjutnya disimpan pada media penyimpanan dengan suhu 4ºC (Akhdiya 2003).
Uji Potensi Enzim Hidrolase Bakteri
Pengujian Potensi Selulolitik.
Pengujian potensi bakteri selulolitik secara semikuantitatif dilakukan dengan menguji aktivitas bakteri dalam mendegradasi selulosa. Metode yang digunakan adalah modifikasi metode Miller (1959) pada substrat selulosa murni, yaitucarboxy methyl cellulose(CMC). Media CMC mengandung 1 g CMC, 0.02 g MgSO4, 7H2O, 0.075 g KNO3, 0.05 g, K2HPO4, 0.002 g FeSO4,7H2O, 0.004 g CaCl2, 2H2O, 0.2 g ekstrak ragi, 1.5 g agar-agar bakto, dan 0.1 g glukosa (Meryandiniet al.2009). Isolat yang telah murni tersebut selanjutnya diambil menggunakan lup dan ditotolkan pada media CMC lalu diinkubasi selama 48 jam pada suhu 37ºC (Syam 2008). Koloni yang sudah tumbuh dicuci dengan larutan merah kongo 0.1% selama sepuluh menit dan dibilas menggunakan NaCl 1%. Selanjutnya diukur diameter koloni dan zona bening yang dihasilkan oleh isolat. Aktivitas selulolitik
ditentukan berdasar indeks selulolitik (IS) yang diperoleh dari perbandingan diameter zona bening dan diameter koloni (Nurkanto 2007).
Pengujian Potensi Proteolitik.
Pengujian potensi bakteri proteolitik secara semikuantitatif dilakukan dengan menguji aktivitas bakteri dalam mendegradasi protein. Metode pengujian yang dilakukan berdasarkan metode Durham et al. (1987). Isolat yang telah murni ditotol pada medium agar susu skim. Agar susu skim dibuat dengan mencampurkan 100 mL susu skim pasteurisasi dicampur dalam kondisi hangat dengan larutan lain yang berisi 7.5 g bakto agar yang dilarutkan dalam 200 mL akuades dan telah diautoklaf selama 15 menit pada suhu 121ºC. Campuran tersebut lalu dimasukkan dalam cawan dan dibiarkan memadat (Amri 2004). Isolat yang telah ditotolkan diinkubasi pada suhu 37ºC selama 48 jam. Setelah masa inkubasi, diukur diameter koloni dan zona beningnya. Aktivitas proteolitik ditentukan berdasar indeks proteolitik (IP) yang diperoleh dari perbandingan diameter zona bening dan diameter koloni (Akhdiya 2003).
Pengujian Potensi Lipolitik. Pengujian potensi bakteri lipolitik secara kualitatif dilakukan dengan menguji aktivitas bakteri dalam mendegradasi lipid. Pengujian dilakukan menurut Sensitive Plate Assay (Kouker & Jaeger 1987 dalam Tika dkk 2007). Koloni yang tumbuh pada media diinokulasikan sebagai spot kecil pada media Tween 80 agar (Gilbertet al.1991). Tween 80 atau disebut juga polisorbat 80 merupakan surfaktan nonionik dan emulsifier yang berasal dari sorbitan polietoksilat dan asam oleat, dan sering digunakan dalam makanan. Polisorbat 80 adalah cairan, kuning kental larut dalam air. Tween 80 mengandung Tween 80, NaCl 5 g, CaCl2.H2O 0.1 g, agar 20 g, dan Bromkresol ungu 25 mg. Selanjutnya medium uji diinkubasi pada 37ºC selama 48 jam. Aktivitas lipolitik ditentukan berdasar zona yang terbentuk pada perubahan warna media dari kuning menjadi ungu.
Penentuan Waktu Optimum Produksi Enzim Lipase
tertinggi. Penentuan waktu optimum inkubasi dilakukan dengan mengkaji hubungan antara kurva pertumbuhan bakteri dan kurva aktivitas enzim lipasenya.
Pembuatan Kurva Pertumbuhan. Sebanyak satu lup isolat bakteri terpilih (data nilai indeks lipolitik terbesar), diinokulasikan ke dalam 50 mL media PGA sebagai starter dan diinkubasi selama 20 jam. Selanjutnya dimasukkan 1% starter, yaitu sebanyak 3 mL ke dalam 300 mL media cair minyak zaitun 1% dan diinkubasi pada inkubator bergoyang. Setiap jam diambil sebanyak 1 mL larutan sampel, kemudian dilakukan pengenceran. Selanjutnya, dilakukan pengukuran jumlah sel dengan metode turbidimetri melalui perhitungan optical density (OD) menggunakan spektrofotometer pada panjang gelombang 600 nm sehingga diperoleh kurva pertumbuhan (Martharina 2009).
Aktivitas Enzim Lipase. Sebanyak satu lup isolat bakteri terpilih (data nilai indeks lipolitik terbesar), diinokulasikan ke dalam 50 mL media PGA sebagai starter dan diinkubasi selama 20 jam. Selanjutnya dimasukkan 1% starter, yaitu sebanyak 3 mL ke dalam 300 mL media cair minyak zaitun 1% dan diinkubasi pada inkubator bergoyang. Pada saat bakteri mulai memasuki fase pertumbuhan eksponensial, yaitu jam keempat, dilakukan pengambilan sampel setiap tiga jam sampai akhir fase pertumbuhan untuk mendapatkan ekstrak kasar enzim per tiga jam (Maranatha 2008). Ekstrak kasar enzim diperoleh dengan sentrifugasi kultur sel pada 3500 rpm selama 15 menit pada suhu 4°C. Pengukuran aktivitas enzim ekstrak kasar dilakukan dengan metode titrimetri. Dicampurkan sebanyak 2 mL minyak zaitun, 4 mL buffer sitrat pH 6, dan 1 mL ekstrak kasar enzim lipase. Campuran dikultivasi pada shaker dengan suhu 37ºC selama 1 jam. Setelah dikultivasi, campuran substrat enzim diinaktifkan dengan penambahan larutan aseton:alkohol (1:1) sebanyak 10 mL kemudian dititrasi dengan NaOH 0.05 N dengan menambah 2-3 tetes fenolftalein 1% sebagai indikator. Aktivitas enzim lipase ditunjukkan dengan perubahan warna saat titrasi dari tidak berwarna menjadi berwarna merah muda. Perlakuan untuk kontrol dilakukan setelah kultivasi selama 1 jam. Aktivitas lipase ditentukan berdasar perhitungan volume NaOH terkoreksi, yaitu volume terpakai sampel dikurangi volume NaOH terpakai blanko. Volume NaOH terpakai selanjutnya dikalikan dengan
konsentrasi NaOH dan dibagi dengan bobot minyak dan masa inkubasi (Paskevicius 2001).
Pengukuran Aktivitas Enzim Lipase Produksi Enzim Lipase. Produksi enzim lipase dilakukan dengan cara mengkultivasi satu isolat bakteri ke dalam 50 mL media PGA sebagai starter dan diinkubasi selama 20 jam. Selanjutnya dimasukkan 1% starter, yaitu sebanyak 3 mL ke dalam 300 mL media cair minyak zaitun 1% dan diinkubasi pada inkubator bergoyang selama waktu optimum panen berdasar data pengamatan sebelumnya. Setelah itu dilakukan sentrifugasi dengan kecepatan 3500 rpm selama 15 menit. Filtrat hasil sentrifugasi disebut ekstrak kasar enzim.
Semipurifikasi Enzim Lipase Melalui Pengendapan dengan Ammonium Sulfat.
Proses pemurnian sampel ekstrak kasar enzim lipase dilakukan dengan fraksinasi bertingkat menggunakan garam ammonium sulfat dengan tingkat kejenuhan (0-20)%, (20-40)%, (40-60)%, dan (60-80)%. Fraksinasi dengan amonium sulfat dilakukan dengan cara menambahkan amonium sulfat sedikit demi sedikit pada larutan ekstrak kasar enzim sambil diaduk. Pengadukan diusahakan sedemikian rupa sehingga tidak menimbulkan busa selama kurang lebih 20 menit. Setiap endapan protein enzim yang didapat dipisahkan dari filtratnya dengan menggunakan sentrifugasi pada kecepatan 10.000 rpm selama 15 menit. Pellet yang dihasilkan selanjutnya dilarutkan dalam larutan buffer sitrat pH 6 untuk dilakukan pengujian selanjutnya (Nurhasanah & Herasari 2008).
Uji Aktivitas Enzim Lipase pada Berbagai Fraksi. Pengujian aktivitas enzim lipase dari fraksi-fraksi enzim yang diproduksi menggunakan metode titrimetri oleh Paskevicius (2001) seperti yang telah dilakukan pada pengujian sebelumnya). Pengujian dilakukan pada kondisi yang sama, yaitu nilai keasaman dan suhu percobaan.
Karakterisasi Enzim Lipase dengan Aktivitas Terbaik
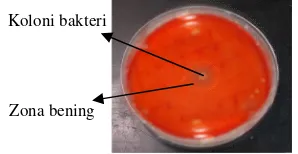
Koloni bakteri
Zona bening 5, 7, dan 9 menggunakan buffer sitrat, buffer
fosfat, dan buffer bórax-borat. Pengujian pH optimum dilakukan pada suhu 37°C. Penentuan suhu optimum dilakukan dengan cara menguji aktivitas enzim hidrolase pada berbagai suhu dalam buffer pH optimum dan diinkubasi 30 menit. Variasi suhu yang digunakan adalah 30ºC, 50ºC, 70ºC, dan 90ºC (Sari 2008).
HASIL DAN PEMBAHASAN
Isolasi Bakteri dari Mata Air Soda Parbubu
Bakteri yang digunakan dalam penelitian ini merupakan bakteri yang diisolasi dari mata air soda Parbubu, Tapanuli Utara. Berdasarkan hasil isolasi yang dilakukan, diperoleh 23 isolat bakteri dari mata air soda Parbubu. Isolat-isolat yang diperoleh dibedakan berdasarkan ciri morfologisnya, yaitu warna, bentuk tepian, ukuran, dan kecembungan. Isolasi bakteri dilakukan dengan menggunakan sampel air dari mata air soda Parbubu, Tarutung, Tapanuli Utara diisolasi dengan metode cawan sebar. Media yang digunakan adalah media peptone glucose agar (PGA) dan potato dextrose agar (PDA), sehingga diperoleh koloni yang berbeda (Nurkanto 2007; Ilyas 2007).
Aktivitas Enzim Hidrolase secara Semikuantitatif
Pengujian potensi bakteri dalam menghasilkan enzim hidrolase dilakukan terhadap tiga enzim, yaitu enzim selulase, protease, dan lipase. Pemilihan tiga jenis enzim ini dimaksudkan karena enzim ini merupakan enzim yang dapat mendegradasi makromolekul utama yang ada di alam, yaitu selulosa, protein, dan lipid. Pengujian potensi enzim dilakukan secara semikuantitatif, yaitu dengan mengukur indeks hidrolitik dari bakteri. Pengujian ini digunakan sebagai informasi awal dalam menentukan enzim yang memiliki aktivitas terbesar dalam mendegradasi substrat.
Aktivitas Enzim Selulase secara Semikuantitatif
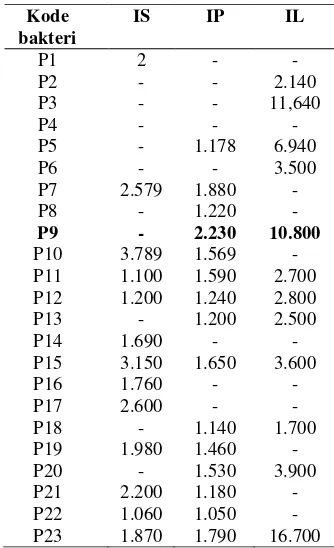
Berdasarkan pengujiian secara semikuantitatif terhadap 23 isolat bakteri Parbubu, sebanyak tiga belas isolat memiliki aktivitas selulase. Isolat P10 merupakan isolat bakteri yang memiliki aktivitas selulase
terbaik dengan nilai indeks selulolitik (IS) sebesar 3,789.
Pengujian enzim selulase secara semikuantitatif dilakukan menggunakan modifikasi metode Miller (1959) pada substrat selulosa murni, yaitucarboxy methyl cellulose (CMC). Menurut Nurkanto (2007) pengujian dilakukan dengan mengukur indeks selulolitik berdasarkan zona bening yang terlihat disekitar koloni bakteri yang ditotolkan pada media CMC. Senyawa CMC merupakan turunan selulosa yang mudah larut dalam air. Oleh karena itu CMC mudah dihidrolisis menjadi gula-gula sederhana oleh enzim selulase dan selanjutnya difermentasi menjadi etanol oleh bakteri. Hal itu yang mendasari pemilihan CMC sebagai substrat untuk pengujian enzim lipase. Pengujian ini berdasarkan proses enzimatik yang dilakukan oleh koloni bakteri yang dapat menghasilkan enzim selulase dan mendegradasi selulosa menjadi glukosa. Hasil degradasi media oleh enzim selulase terlihat sebagai zona bening, sedangkan media agar yang masih mengandung selulosa terlihat berwarna merah karna adanya ikatan dengan pewarna merah kongo.
Gambar 6 Hasil uji semikuantitatif selulase rpada isolat P10.
Aktivitas Enzim Protease secara Semikuantitatif
Zona bening Koloni bakteri
a b c
yang dilaporkan Akhdiya (2003) disebut sebagai indeks proteolitik (IP) yang diperoleh dari perbandingan diameter zona bening dan diameter koloni.
Gambar 7 Hasil uji semikuantitatif protease pada isolat P9.
Aktivitas Enzim Lipase secara Semikuantitatif
Pengujian enzim lipase secara semikuantitatif dilakukan dengan menguji aktivitas bakteri dalam mendegradasi lipid menggunakan metode Sensitive Plate Assay (Kouker & Jaeger 1987 dalam Tika et al. 2007). Aktivitas lipase ditentukan berdasar zona yang terbentuk pada perubahan warna media yang mengandung bromkresol ungu dari kuning menjadi ungu. Bromkresol ungu merupakan indikator pH dengan rentang pH 5.2 berwarna ungu. Pada pengujian ini, terdapat perubahan pH dari asam menjadi basa dengan dihasilkannya asam lemak dan gliserol dari polimer lipid sehingga dapat diamati secara semikuantitatif banyaknya enzim yang dihasilkan oleh sel untuk mendegradasi polimer lipid. Berdasarkan pengujiian secara semikuantitatif terhadap 23 isolat bakteri Parbubu, sebanyak dua belas isolat memiliki aktivitas lipase. Isolat bakteri yang memiliki indeks lipolitik (IL) besar adalah isolat P3, P9, dan P23 dengan nilai indeks lipolitik berturut-turut sebesar 11,640; 10,80; 16,700.
Uji kualitatif tidak selalu menjadi dasar yang baik untuk melihat aktivitas enzim, sehingga perlu dilakukan uji lanjutan terhadap aktivitas proteasenya. Menurut Ward (1985) tidak selalu terdapat korelasi yang baik antara daerah jernih di sekitar koloni pada media padat dengan kemampuan organisme tersebut. Beberapa faktor yang diduga menjadi penyebab tidak terkorelasinya nilai aktivitas hidrolisis secara kualitatif dengan nilai aktivitas enzim secara kuantitatif adalah kecepatan pertumbuhan setiap isolat pada medium padat atau cair dan jumlah inokulum yang diberikan pada kedua medium.
Berdasarkan seluruh pengujian terhadap ketiga enzim hidrolase, aktivitas enzim
tertinggi merupakan enzim lipase. Ketiga koloni bakteri yang memiliki nilai IL tertinggi, yaitu P3, P9, dan P23 dipilih koloni P9 untuk diuji lanjut secara kuantitatif. Dipilih koloni P9 karena koloni bakteri ini juga memiliki aktivitas yang tertinggi pada uji semikuantitatif enzim protease.
Gambar 8 Hasil uji semikuantitatif lipase wpada (a) isolat P3, (b) isolat P9, wdan (c) isolat P23.
Tabel 1 Hasil uji aktivitas enzim hidrolase secara semikuantitatif
Waktu Optimum Produksi Enzim Lipase
Hasil pembuatan kurva pertumbuhan dan kurva produksi dapat dilihat pada Gambar 9. Kurva tersebut menunjukkan bahwa aktivitas enzim optimum terjadi pada jam ketujuh yang merupakan fase bagian awal dari fase pertumbuhan eksponensial atau fase log. Hal ini berarti isolat bakteri P9 dapat dilakukan pemanenan enzim lipase pada jam ketujuh setelah masa kultivasi.
Kode bakteri
IS IP IL
P1 2 -
-P2 - - 2.140
P3 - - 11,640
P4 - -
-P5 - 1.178 6.940
P6 - - 3.500
P7 2.579 1.880
-P8 - 1.220
-P9 - 2.230 10.800
P10 3.789 1.569
-P11 1.100 1.590 2.700 P12 1.200 1.240 2.800
P13 - 1.200 2.500
P14 1.690 -
-P15 3.150 1.650 3.600
P16 1.760 -
-P17 2.600 -
-P18 - 1.140 1.700
P19 1.980 1.460
-P20 - 1.530 3.900
P21 2.200 1.180
-P22 1.060 1.050
Pengujian aktivitas enzim untuk pembuatan kurva pertumbuhan dilakukan pada titik-titik kurva pertumbuhan. Pada pengujian ini dilakukan pengukuran aktivitas enzim setiap tiga titik (jam) diawali pada awal fase pertumbuhan eksponensial. Menurut Martharina (2010), penentuan waktu inkubasi optimum dimaksudkan untuk memperoleh waktu yang tepat disaat aktivitas enzim berada paling tinggi sehingga dapat dilakukan pemanenan enzim dengan hasil terbaik. Waktu inkubasi optimum ditentukan berdasarkan kurva pertumbuhan bakteri dan kurva produksi dari bakteri. Selain itu, kurva ini juga dapat digunakan untuk memperkirakan penggolongan jenis metabolit dari enzim yang dihasilkan. Berdasarkan hasil pengujan, isolat bakteri P9 menghasilkan enzim dengan aktivitas optimum pada jam ketujuh yang merupakan awal fase pertumbuhan eksponensial. Hal ini menunjukkan bahwa isolat bakteri P9 menghasilkan enzim yang merupakan metabolit primer. Menurut Poedjiadi (1994), metabolit primer merupakan suatu zat yang dihasilkan pada fase pertumbuhan karena digunakan sebagai zat yang menopang pertumbuhan suatu makhluk hidup.
Aktivitas Enzim Lipase
Pengukuran aktivitas enzim lipase dilakukan pada beberapa fraksi enzim lipase, yaitu fraksi ekstrak kasar enzim (crude enzyme), fraksi enzim dengan pengendapan (0-20)%, fraksi (20-40)%, fraksi (40-60)%, dan fraksi (60-80)% ammonium sulfat. Pengukuran aktivitas enzim lipase pada fraksi ekstrak kasar enzim ini menunjukkan bahwa aktivitas rata-rata enzim hasil pengukuran duplo adalah 0.5882 U/mL. Hasil pengukuran terhadap fraksi enzim dengan pengendapan ammonium sulfat menunjukkan nilai aktivitas
rata-rata enzim hasil pengukuran duplo pada fraksi (0-20)% adalah 1.3627 U/mL, fraksi (20-40)% adalah 2.3235 U/mL, fraksi (40-60)% adalah 2.4410 U/mL, dan fraksi (60-80)% adalah 3.5245 U/mL.
Produksi ekstrak kasar enzim diperoleh dengan mengkultivasi satu isolat bakteri P9 sebagai isolat terpilih ke dalam media yang yang mengandung minyak zaitun sebagai penginduksi. Menurut Rajendran dan Tangavelu (2007), minyak zaitun merupakan salah satu minyak nabati terbaik setelah minyak kacang tanah untuk digunakan sebagai penginduksi enzim lipase. Selain itu, minyak zaitun juga telah banyak digunakan dalam berbagai pengujian enzim lipase. Media tersebut selanjutnya diinkubasi pada inkubator bergoyang dengan suhu ruang selama tujuh jam sesuai dengan hasil pengamatan waktu produksi optimum enzim. Inkubasi pada kondisi optimum pertumbuhan dimaksudkan untuk memperoleh enzim lipase dalam jumlah cukup besar. Setelah itu dilakukan pemisahan antara media dengan sel menggunakan sentrifugasi dengan kecepatan 3500 rpm selama 15 menit suhu 4°C. Filtrat hasil sentrifugasi merupakan campuran antara media dan enzim yang selanjutnya disebut ekstrak kasar enzim (Paskevicius 2001). Sedangkan sel yang memiliki massa yang yang lebih besar tertarik ke pusat sentrifugal sehingga menjadi pellet yang menempel pada dinding tabung sentrifus.
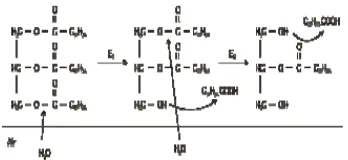
Ekstrak kasar enzim selanjutnya diuji aktivitas lipasenya menggunakan metode titrimetri. Pengujian menggunakan minyak zaitun sebagai substrat, buffer sitrat pH 6, dan fraksi enzim. Campuran dikultivasi pada inkubator bergoyang dengan suhu 37ºC selama 1 jam. Minyak zaitun terdiri atas trigliserida (trioleogliserol). Setiap 2 mL minyak zaitun memiliki massa 1.7 gram. Trioleogliserol dengan rumus molekul Gambar 9 Kurva pertumbuhan dan aktivitas lipase isolat bakteri P9.
Kurva pertumbuhan bakteri P9
Gambar 10 Reaksi hidrolisis enzimatis spesifik pada minyak zaitun.
(C17H35COO)3C3H5 memiliki berat molekul 884 g/mol, sehingga dalam 2 mL sampel minyak zaitun terdapat 1.923 mmol trigliserida. Pada masa inkubasi, minyak zaitun, (C17H35COO)3C3H5,akan dihidrolisis oleh lipase menjadi asam lemak
C17H34COOH dan gliserol
berdasarkan reaksi pada Gambar 10. Setelah diinkubasi, campuran substrat enzim diinaktifkan dengan penambahan larutan aseton:alkohol (1:1). Campuran aseton dan alkohol mendenaturasi struktur enzim yang merupakan protein dengan mengganggu rantai ikatan hidrogen intramolekul samping. Hal ini mengakibatkan rusaknya struktur sekunder dan tersier dari enzim sehingga enzim berhenti bekerja. Kemudian campuran tersebut dititrasi dengan NaOH 0.05 N dengan sebelumnya menambahkan 2-3 tetes fenolftalein 1% sebagai indikator. Asam lemak hasil hidrolisis bereaksi dengan NaOH menjadi C17H34COONa dan H2O (Wulan et al.2007).
Aktivitas enzim lipase ditunjukkan dengan perubahan warna saat titrasi dari tidak berwarna menjadi berwarna merah muda akibat terjadinya perubahan pH pada larutan. Awal titrasi netralisasi, larutan tidak menghasilkan warna karena NaOH yang diteteskan berikatan dengan asam lemak, sehingga tidak dapat mengubah pH larutan, akibatnya fenolftalein sebagai indikator pH tidak memberikan perubahan warna. Warna merah muda muncul saat NaOH tidak lagi dapat berikatan dengan asam lemak, sehingga memberikan sifat basa pada larutan dan menimbulkan warna merah muda sebagai indikasi perubahan pH larutan. Banyaknya NaOH yang dapat berikatan dengan asam lemak menunjukkan banyaknya asam lemak yang terdapat dalam larutan sebagai hasil hidrolisis minyak zaitun. Perlakuan untuk kontrol dilakukan setelah kultivasi selama 1 jam, yaitu berarti sebelum terjadi reaksi degradasi lipid oleh lipase. Aktivitas lipase ditentukan berdasar perhitungan volume NaOH terkoreksi, yaitu volume terpakai sampel dikurangi volume NaOH terpakai blanko. Volume NaOH terpakai selanjutnya dikalikan dengan konsentrasi NaOH dan dibagi dengan bobot minyak dan masa inkubasi (Paskevicius 2001).
Setelah diperoleh ekstrak kasar enzim, dilanjutkan dengan purifikasi enzim secara fraksinasi bertingkat menggunakan garam ammonium sulfat. Pengendapan menggunakan garam ammonium sulfat merupakan salah satu cara yang mudah untuk
mengendapkan protein enzim, selain denaturasi oleh panas atau pH, sentrifugasi diferensial, filtrasi gel, dan elektroforesis (Martinet al. 1995). Penggunaan ammonium sulfat disebabkan karena ammonium sulfat mudah didapatkan, harganya relatif murah, bersifat menstabilkan enzim (Yurnaliza 2002). Penambahan garam ammonium sulfat dilakukan secara perlahan dan dilakukan pengadukan secara terus menerus. Saat pengadukan, ammonium sulfat akan berikatan dengan gugus ionik yang berada pada ujung-ujung protein enzim. Pada kondisi tertentu, garam ammonium sulfat berada pada kondisi jenuh dan tidak dapat terlarut lagi. Pada kondisi ini, molekul-molekul enzim yang telah berikatan dengan ammonium sulfat meningkat bobot molekulnya sehingga terendapkan menjadi pellet ketika disentrifugasi. Ekstrak kasar enzim dengan penambahan ammonium sulfat secara bertingkat selanjutnya disebut sebagai fraksi (0-20)%, fraksi (20-40)%, fraksi (40-60)%, dan fraksi (60-80)% ammonium sulfat. Setelah penambahan garam, dilakukan pemisahan enzim dengan sentrifugasi 10.000 rpm selama 15 menit pada suhu 4°C. sentrifuse dilakukan pada suhu 4°C agar enzim yang merupakan protein tidak rusak akibat panas yang ditimbulkan oleh sentrifuse. Enzim yang sudah terendapkan setelah mengalami sentrifus berda pada pusat sentrifugal,yaitu menjadi pellet yang menempel pada dinding tabung sentrifuse, sedangkan media terpisahkan berupa filtrat. Pellet yang dihasilkan selanjutnya dilarutkan dalam larutan buffer sitrat pH 6 untuk dilakukan pengukuran aktivitas enzim lipase (Nurhasanah & Herasari 2008).
sebesar 5.992 kali dibandingkan e enzim. Aktivitas spesifik enzim oleh kadar protein, semakin ting spesifik suatu enzim maka sem kemurnian enzim tersebut. menunjukkan terjadinya pemisah lain yang bukan enzim. M aktivitas spesifik pada tiap tahap menunjukkan bahwa purifikasi e dilakukan cukup baik (Nurh Herasari 2008). Enzim deng kemurnian dan aktivitas tertinggi dilakukan tahap karakterisasi e perlakuan pH dan suhu.
Karakterisasi Enzim Lip
Berdasarkan pengujian akti lipase, fraksi enzim dengan pem 80)% ammonium sulfat memili terbaik. Fraksi enzim ini dilakukan karakterisasi enzim pad kondisi pH dan suhu untuk kondisi pH dan suhu optim enzimatis.
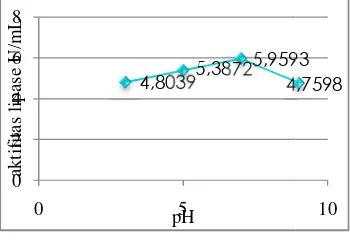
Hasil pengujian aktivitas e beberapa kondisi pH (Ga menunjukkan bahwa enzim lipas aktivitas optimum pada pH 7, y 5.9593 U/mL. Pengujian aktiv lipase pada pH optimum pada be menunjukkan bahwa aktivitas lipa pada suhu 90ºC dengan aktivita 4.9019 U/mL (Gambar 13).
Penentuan pH optimum dengan mengukur aktivitas enzim substrat minyak zaitun dalam be Variasi pH media yang digunakan 3, 5, 7, dan 9 menggunakan buffe dan 5, buffer fosfat pH 7, serta b borat pH 9. Pengujian pH optimum pada suhu 37°C dengan m metode titrimetri.
Gambar 11 Aktivitas lipase pad wfraksi enzim.
n ekstrak kasar im dipengaruhi tinggi aktivitas emakin tinggi t. Hal ini isahan protein Meningkatnya ap pemurnian i enzim yang urhasanah & ngan tingkat gi selanjutnya i enzim pada
Lipase
ktivitas enzim emurnian (60-iliki aktivitas i selanjutnya pada beberapa menentukan timum reaksi
s enzim pada Gambar 12) ipase memiliki , yaitu sebesar tivitas enzim beberapa suhu lipase optimum tivitas sebesar
m dilakukan im lipase pada berbagai pH. kan adalah pH ffer sitrat pH 3 buffer bórax-um dilakukan menggunakan
Gambar 12 menunjukkan terdapat variasi aktivitas enzim pada k keasaman yang berbeda-beda. Ha umumnya terjadi karena adanya peru struktur sekunder dan tersier dari enzim pH yang optimum muatan gugus sa asam amino berada pada keadaan yang sehingga enzim sangat efisien mempercepat reaksi biokimia yang spesifik. Aktivitas optimum lipase d pada pH 7. Hal ini disebabkan karena kondisi pH 7 gugus pemberi dan pen proton yang penting pada sisi katalitik berada pada keadaan yang diing sehingga aktivitas katalitiknya (Nurhasanah & Herasari 2008).
Penentuan suhu optimum dila dengan cara menguji aktivitas enzim pada beberapa suhu dalam buffer sebagai kondisi pH optimum lipase diinkubasi 30 menit. Variasi suhu digunakan adalah 30ºC, 50ºC, 70ºC, dan (Sari 2008).
Hasil pengujian aktivitas lipase beberapa pH tertera pada Gambar 13. temperatur kurang dari 50°C enzim stabil, tetapi hidrolisis substrat minyak oleh enzim tidak berjalan secara mak Dengan meningkatnya temperatur, kinetik molekul-molekul yang be bertambah sehingga molekul yang be semakin banyak dan produk yang diha semakin besar. Enzim terus meng peningkatan aktivitas sampai pada 90°C. Di atas temperatur optimum, ak enzim menurun tajam hal ini terjadi k enzim mengalami denaturasi protein dapat merubah konformasi struktur m sehingga enzim kehilangan sifat alamia Pada temperatur tinggi, substrat juga mengalami perubahan konformasi seh gugus reaktifnya mengalami hambatan memasuki sisi aktif enzim (Suhartono 1
Gambar 12 Penentuan pH optimum en sssssssssssslipase. ada beberapa 4,80395,3872 5,9593 4,7598 0 2 4 6 8 0 5 ak tif ita s lip ase U/ m L pH bahwa a kondisi Hal ini erubahan im. Pada samping ng sesuai dalam g sangat dicapai ena pada penerima litik enzim iinginkan tinggi ilakukan m lipase r pH 7 ase dan hu yang dan 90ºC
Sedangkan pada suhu yang lebih suhu optimum, aktivitas enzim ju Hal ini disebabkan karena renda aktivasi yang tersedia. Energ dibutuhkan untuk menciptaka tingkat kompleks aktif, baik da enzim atau molekul substrat (Nu Herasari 2008).
SIMPULAN DAN SARAN
Simpulan
Mata air soda Parbubu, Tap memiliki potensi mikrobial. Se isolat bakteri diperoleh dari ma tersebut. Isolat tersebut memili enzim lipase, protease, dan selula enzim lipase sebagai enzim deng terbaik. Enzim lipase dengan p (60-80)% ammonium sulfat aktivitas tertinggi dibandingkan yaitu sebesar 3.5245 U/mL. En memiliki aktivitas optimum pada yaitu sebesar 5.9593 U/mL dan 90ºC dengan aktivitas sebesar 4.90
Saran
Perlu dilakukan penelitian untuk identifikasi bakteri P9, k enzim lipase pada substrat ya logam-logam inhibitor, nilai Km Selain itu juga perlu dilakukan lebih lanjut tentang aktivitas dan k enzim protease dan selulase dari yang diperoleh dari mata air so lainnya.
DAFTAR PUSTAK
Akhdiya A. 2003. Isolasi bakterenzim protease alkalin Buletin Plasma Nutfah9: 98-Gambar 13 Penentuan suhu optim ssssssssssssenzim lipase.
3,9657 3,8294 0 1 2 3 4 5 6 0 50 A k ti fi ta s L ip as e (U /m L ) Suhu
ih rendah dari juga rendah. dahnya energi ergi tersebut takan kondisi dari molekul Nurhasanah &
ARAN
apanuli Utara Sebanyak 23 mata air soda iliki aktivitas lulase dengan ngan aktivitas pengendapan lfat memiliki an fraksi lain, Enzim lipase ada pH netral, an pada suhu .9019 U/mL.
n lebih lanjut , karakterisasi yang sesuai, Km dan Vm. an pengkajian n karakterisasi ri isolat-isolat soda Parbubu
AKA
teri penghasil termostabil. 98-102.
Amri E. Aktivitas amilase dan protease dihasilkan oleh isolat aktinom [skripsi]. Bogor: Fakultas Matem dan Ilmu Pengetahuan Alam, I Pertanian Bogor.
Choughlan MP. 1985. The propertie fungal and bacterial cellulose comment on their production application. Biotechnology and G Engineering3:39-109.
Crueger W, Crueger A, Brock TD, 1984. Biotechnology: A text bo industrial Microbiology. Sunde Minuaer Associates.
[DITA] Direktori Ikan dan Tanaman 2003. Kesadahan air. Ornamenta Information Service Highligh [terhubung berkala]. htt fish.com/direktori.php [22 fe 2010].
Durham DR, Stewart DB, Stellwag EJ. Novel alkaline and heat stable proteases from alkalaphilicBacill strain GX6638. J. Bacterial 169: 2768.
Foster EM, Nelson FE, ML Speck Doetsch, Olson JCJr. 1961. Microbiology. New Jersey: Pr Hall.
Gilbert EJ, Cornish A, Jones C. Purification and properties extracellulear lipase fromPseudo aeruginosa EF2. Journal of G Microbiology137:2223-2229.
Hardjoko S, Indrati NS, T Bantacut. Biokonversi: Pemanfaatan Li Industri Pertanian. Bogor:PAU IP
Hasan R, Setiadama, Risdianto D, Supa 2007. Geologi daerah panas Sipoholon-Tarutung, kabu Tapanuli Utara, Sumatera Utara. Sumber Daya Geologi. [terh berkala]. http://www.bgl.esdm [20 Februari 2010].
Ilyas M. 2007. Isolasi dan ident mikoflora kapang pada sampel s daun tumbuhan di Kawasan G Lawu, Surakarta, Jawa Te Biodiversitas8: 105-110.
optimum
4,45094,9019
100
ase yang omisetes tematika , Institut
erties of se with tion and Genetic
, editor. book of nderland:
an Air. ntal Fish lights. .
http://o-februari
J. 1987. le serine cillussp.
69:2762-eck, RN . Dairy Prentice
. 1991. ties of domonas General
ut. 1989. Limbah IPB.
pardi K. s bumi abupaten ra. Pusat terhubung m.go.id/
Koolman J, Rohm K. 2001.Atlas Berwarna dan Teks Biokimia. Septelia, penerjemah. Jakarta: Hipokrates.
Kouker G and Jaeger KE. 1987. Specific and sensitive plate assay for bacterial lipases. Appl Environ Microbiol 53: 211-213.
Kullp K. 1975. Carbohydrate. Di dalam: Reed G, editor. Enzyme and Food Processing. New York: Academic Pr.
Kusumaningtyas E, Tan WS, Zamrod Z, Eshaghi M, Yusoff K. 2004. Existence of two forms of L protein ofNewcastle disease virus isolates due to a compensatory mutation in Domain V. Arch Virol 149: 1859-1865.
Lehninger AL. 1982.Dasar-dasar Biokimia. Thenawidjaja M, penerjemah; Jakarta: Erlangga. Terjemahan dari: Principles of Biochemistry.
Macrae AR. 1983. Extracelullar microbial lipases. Di dalam Fogarty WM, editor. Microbial Enzymes and biotecknology. England: Applied Science Publiser.
Madigan MT, Marrs BL. 1997. Extremophiles.Sci. Am82-87.
Maranatha. 2008. Aktivitas enzim selulase isolat asal Indonesia pada berbagai substat limbah pertanian. [skripsi]. Bogor: Fakultas Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Martharina D. 2010. Karakterisasi bakteri asam laktat termofilik dari sumber air panas Gunung Pancar, Bogor [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Martin DW, Mayes PA, Rodwell VW. 1981. Harper’s Review of Biochemistry. California: Lange Medical Publication.
Meryandini A et al. 2009. Isolasi bakteri selulolitik dan karakterisasi enzimnya. Makaira Sains13:33-38.
Miller GL. 1959. Use of Dinitrosalicylic Acid Reagent of Determination of Reducing sugar.Anal Chem 31: 246-248.
Nurkanto A. 2007. Identifikasi aktinomisetes tanah hutan pasca kebakaran bukit Bangkirai Kalimantan Timur dan potensinya sebagai pendegradasi selulosa dan pelarut fosfat.Biodiversitas 8: 314-319.
Nurhasanah, Herasari D. 2008. Pemurnian Enzim Lipase dari Bakteri Lokal dan Aplikasinya dalam Reaksi Esterifikasi. Prosiding Seminar Nasional Sains dan Teknologi-II 2008 Universitas Lampung. hlm 267-277.
Paskevicius A. 2001. Lipase activity of yeast and yeasts-like fungi functioning under natural conditions. Institute Of Botany.
[terhubung berkala].
http://images.katalogas.lt/maleidykla/bi o4/B-16.pdf [23April 2010].
Pelczar MJJr, Chan ECS. 1986.Dasar-Dasar Mikrobiologi. Volume ke-1. Hadioetomo RS, Imas T, Tjitrosomo SS, Angka SL, penerjemah; Jakarta: UI Pr. Terjemahan dari: Elements of Microbiology.
Poedjiadi A. 1994. Dasar-Dasar Biokimia. Jakarta: UI Press.
Rahman A, Fardiaz S, Rahaju WP, Suliantari, Nurwitri CC. 1992. Teknologi Fermentasi Susu. Bogor: Departemen Pendidikan dan Kebudayaan Direktorat Jenderal Pendidikan Tinggi, Institut PertaniSan Bogor.
Rajendran A, Thangavelu V. 2007. Optimization of medium compotition for lipase production by candida rugosa NCIM 3462 using response surface methodology. Canada Journal Microbiol53:643-655.
Ruhiman B. 2009. Ekspedisi Geografi Indonesia Sumatera Utara 2009. Bandung: Badan Koordinasi Survei dan Pemetaan Nasional.
Sari WW. 2008. Karakterisasi selulase bakteri asal tanah pertanian Jawa Tengah dan Jawa Barat [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Tarutung, Tapanuli Utara, Sumatera Utara. Pusat Sumber Daya Geologi.
[terhubung berkala].
http://www.bgl.esdm.go.id/ [20 Februari 2010].
Subba R. 1995. Soil Microorganisms and Plant Growth,3rd ed. New Hampshire: Science Publisher.
Suhartono, M.T. 1989. Enzim dan Bioteknologi. PAU Bioteknologi IPB. Bogor.
Suhartono MT. 2000. Eksplorasi protease bakteri asal Indonesia untuk aplikasi industri dan riset bioteknologi. Di dalam: Suwahyono U, Rachman MA, Tambunan J, Setyahadi S, Angkoso G, Wahyudi P, Laily N, Retno WK, Trismilah, editor. Mikrobiologi, Enzim dan Bioteknologi. Prosiding Seminar Nasional Industri Enzim dan Bioteknologi II. hlm 125 -130.
Syam KA. 2008. Optimasi produksi dan aktivitas enzim selulse dari mikrob selulolitik asal rayap [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Tika IN, Redhana IW, Ristiati NP. 2007. Isolasi enzim lipase termostabil dari bakteri termofilik isolat air panas Banyuwedang Kecamatan Gerogak, Buleleng Bali.Akta Kimia Indonesia 2: 109–112.
Ward OP. 1985. Proteolytic enzymes. Di dalam: Young MM, editor. Comprehensive Biotechnology: The principles,Applications, and Regulations of Biotechnology in Industry, Agriculture and Medicine. Vol. 3.Oxford: Pergamon Press.
Widyastuti N, D Astuty. 2000. Optimasi produksi dan semikarakterisasi protease dariBacillussp. Laporan teknik.
Winarno FG. 1987. Biokimia Pangan. Jakarta: PT. Gramedia Utama.
Wulan PPDK, Rejoso MT, Hermansyah H. 2007. Reaksi Hidrolisis Minyak Zaitun Menggunakan Lipase Rhizopus oryzae yang Diimobilisas Melalui Metode
Adsorpsi. Depok: Departemen Pendidikan dan Kebudayaan Direktorat Jenderal Pendidikan Tinggi, Universitas Indonesia.
Lampiran 1 Bagan alir penelitian Sumber mata air soda Parbubu
Air soda dari sumber mata air soda Parbubu
Isolat Bakteri
Uji potensi enzim hidrolase dari isolat bakteri
Uji potensi selulolitik
Penentuan waktu inkubasi optimum melalui kurva
pertumbuhan
Karakterisasi enzim dengan variasi suhu dan pH
Uji potensi proteolitik
Uji potensi lipolitik
Lampiran 2 Isolasi bakteri dari sumber mata air soda Parbubu (Nurkanto 2007;
Ilyas 2007)
Air soda Parbubu
Pengenceran 9 kali menggunakan larutan NaCl
0.85%
Media PGA hangat 1 mL sampel
Inkubasi pada suhu ruang selama 48 jam
Pemurnian bakteri dengan morfologi yang berbeda
Lampiran 3 Uji potensi enzim hidrolase bakteri
1. Pengujian potensi selulolitik (Meryandini 2007)
2. Pengujian potensi proteolitik (Akhdiya 2003)
3. Pengujian potensi lipolitik (Kouker & Jaeger 1987)
Isolat bakteri Ditotol pada media
susu skim padat
Inkubasi pada suhu ruang selama 48 jam
Ukur koloni dan zona bening Hitung indeks proteolitik
Isolat bakteri Ditotol pada media
Tween 80
Inkubasi pada suhu ruang selama 48 jam
Ukur koloni dan zona ungu Hitung indeks lipolitik
Isolat bakteri Ditotol pada media
CMC padat
Inkubasi pada suhu ruang selama 48 jam
Bilas dengan larutancongo
red1% selama 10 menit
Bilas dengan NaCl 1%
Ukur koloni dan zona bening
Lampiran 4 Pembuatan kurva pertumbuhan bakteri lipolitik
Isolat bakteri Inokulasi pada media
starter cair (PGB)
Inkubasi pada inkubator bergoyang
selama 6 jam
Inokulasi 1% starter pada media cair minyak zaitun 1% Inkubasi pada
inkubator bergoyang
Pengukuran OD pada 600 nm setiap jam
Lampiran 5 Uji aktivitas enzim lipase dengan metode titrimetrik (Paskevicius
2001)
Isolat bakteri
Peremajaan pada media minyak zaitun
Inkubasi 1-3 hari pada suhu ruang
Ditambah 25 mL buffer sitrat pH 6.0, 50 mM
Sentrifuse selama 15 menit pada 3500 rpm, suhu 4-5ºC
Dicampur dengan minyak zaitun & buffer sitrat
Diinkubasi bergoyang dengan suhu 37ºC selama 1 jam
Inaktifasi enzim dengan larutan aseton:alkohol (1:1)
Titrasi dengan NaOH 0.05 N dengan
indikatorphenolphtalein1%
Lampiran 6 Karakterisasi enzim lipase (Sari 2008)
Ekstrak enzim kasar (crude enzyme)
Dicampur dengan substrat minya zaitun
Perlakuan pH (3,5,7,9) dengan penambahan buffer sitrat, fosfat, dan tris-HCl
Perlakuan suhu dengan inkubasi bergoyang pada suhu 37ºC, 50ºC, 70ºC, dan 90ºC selama
1 jam
Inaktivasi enzim
Kode Bakteri
Warna deskripsi Foto
P1 jingga Mengkilat, sekitar koloni lebih terang, ukuran cukup besar (2-10 mm), bentuk bergelombang mirip bunga
P2 Jingga Mengkilat, sekitar koloni lebih terang, ukuran cukup besar (2-10 mm), bentuk bergelombang mirip bunga, terlihat ada serabut
P3 Putih Bulat, tidak mengkilat, pinggir tipis agak mengkilat, ukuran kecil (0.5-2mm)
P4 jingga Mengkilat, sekitar koloni lebih terang, ukuran lebih besar (0.8-1.8 cm), bentuk memanjang
P5 bening Mengkilat, bundar, kecil sekali (tidak dapat diukur)
P6 kuning Menonjol ke permukaan media, bulat, tidak mengkilat, pinggir halus, diameter 7 mm
P7 Putih
gading
Center menonjol, mengkilat, pinggir halus agak bergaris, D=1.2 cm
P8 Putih
kekuning an
Bulat, kecil (D=0.5 cm)
P9 Putih
gading
Bulat, mengkilat, ukuran sangat kecil (tidak bisa diukur)
Lampiran 7 Isolat bakteri dari mata air soda Parbubu, Tapanuli Utara
P11 kuning Bulat, mengkilat, kecil (D= 1mm)P12 Putih
gading
Bulat, mengkilat, ukuran sangat kecil (tidak bisa diukur)
P13 Putih
kekuning an
Bulat, kecil (D=0.5 cm)
P14 bening Mengkilat, bundar, kecil sekali (tidak dapat diukur)
P15 Putih
gading
Bulat, mengkilat, ukuran sangat kecil (tidak bisa diukur)
P16 putih Mengkilat, bundar, tepian halus, zona sekitar koloni memudar, D= 1-4 mm
P17 putih Bulat, pinggir tidak halus, tidak mengkilat, D= 1 mm
P18 Putih
gading
Bulat (seperti mutiara), mengkilat, tepian halus tanpa zona yang memudar, D= 1mm
P19 Putih
kekuning an
Lampiran 7 Isolat bakteri dari mata air soda Parbubu, Tapanuli Utara
P20 putih Mengkilat, tepian menonjol, ada zona memudar disekitar koloni, D= 2-4 mm
P21 Putih Bulat, tidak mengkilat, pinggir tipis agak mengkilat, ukuran kecil (0.5-2mm)
P22 jingga Mengkilat, sekitar koloni lebih terang, ukuran cukup besar (2-10 mm), bentuk bergelombang mirip bunga
Lampiran 8 Perhitungan aktivitas lipase
Aktivitas hidrolisis lipase =
(
)
Keterangan:
Satu unit aktivitas lipase adalah banyaknya enzim yang dibutuhkan untuk
menghidrolisis minyak menghasilkan satu µmol produk selama satu jam.
A
: volume NaOH (titran) untuk titrasi sampel (mL)
B
: volume NaOH (titran) untuk titrasi blanko (mL)
N NaOH
: Normalitas NaOH yang digunakan
1000
: faktor konversi dari mmol keµmol
W
: berat minyak (mg)
Lampiran 9 Hasil uji kuantitatif aktivitas lipase pada ekstrak kasar lipase
Jam ke-
Ulangan
Volume titran
terpakai (mL)
Rata-rata volume
titran terpakai (mL)
Aktivitas lipase
(U/mL)
Blanko
1
2.00
1.975
0
2
1.95
4
1
2.35
2.300
0.1354
2
2.25
7
1
2.75
2.575
0.3229
2
2.40
10
1
-
2.300
0.1354
2
2.30
13
1
2.70
2.500
0.2187
2
2.30
16
1
2.25
2.250
0.1146
2
2.25
19
1
2.20
2.300
0.1354
2
2.40
22
1
2.40
2.325
0.1458
2
2.25
25
1
2.30
2.300
0.1354
2
-28
1
2.25
2.350
0.1562
Lampiran 10 Aktivitas lipase pada fraksi lipase dengan pengendapan ammonium
sulfat
Konsentrasi
(NH
4)
2SO
4Ulangan
Volume titran
terpakai (mL)
Rata-rata volume
titran terpakai (mL)
Aktivitas
lipase (U/mL)
Blanko
1
2.65
2.55
0
2
2.45
0%
1
2.92
3.15
0.5882
2
3.38
(0-20)%
1
3.65
3.94
1.3627
2
4.23
(20-40)%
1
4.57
4.92
2.3235
2
5.27
(40-60)%
1
4.93
5.04
2.4410
2
5.15
(60-80)%
1
5.52
6.145
3.5245
Lampiran 11 Aktivitas lipase pada suhu 37°C dan berbagai pH
pH
Bufer
Ulangan
Volume titran
blanko (mL)
Volume titran
sampel (mL)
Aktivitas
lipase (U/mL)
Terpakai
Rata-rata
Terpakai
Rata-rata
3
1
10.93
10.925
18.61
17.755
4.8039
2
10.92
16.90
5
1
4.98
4.900
10.42
10.395
5.3872
2
4.82
10.37
7
1
3.68
3.571
10.65
9.65
5.9593
2
3.46
8.65
9
1
3.07
3.36
8.55
8.215
4.7598
Lampiran 12 Aktivitas lipase pada pH 7 dan berbagai suhu
Suhu
Ulangan
Volume titran
blanko (mL)
Volume titran
sampel (mL)
Aktivitas
lipase (U/mL)
Terpakai
Rata-rata
Terpakai
Rata-rata
30°C
1
3.30
3.375
7.07
7.42
3.9657
2
3.45
7.77
50°C
1
3.40
3.42
6.60
7.326
3.8294
2
3.44
8.05
70°C
1
3.45
3.41
7.40
7.95
4.4509
2
3.37
8.60
90°C
1
3.60
3.60
9.70
8.60
4.9019