METODE
POLYMERASE CHAIN REACTION
(PCR)
FIDYATUN KHOIRIYAH
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa tesis Identifikasi Molekular Isolat Lokal
Staphylococcus aureus Dengan Metode Polymerase Chain Reaction (PCR) adalah karya saya sendiri dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumebr informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, September 2011
FIDYATUN KHOIRIYAH. Molecular Identification Local Isolates of Staphylococcus aureus by Polymerase Chain Reaction (PCR) Method. Under direction of RATIH DEWANTI-HARIYADI and SRI HENDRASTUTI HIDAYAT.
Foodborne disease is a major concern worldwide, and S. aureus is continuously involved in foodborne disease outbreaks. Staphylococcal enterotoxins (SE) are important virulence factor for the bacterial intoxications. This research aimed to identify local isolates of S. aureus based on universal 16S rRNA gene and to identify whether they possess SE A and C1 genes. Bacterial DNA isolation was conducted using Doyle and Doyle extraction method with some modification. Amplification of genes encoding for 16S rRNA, SEA
and SEC1 were investigated using primers 63f/1387r, SEA-1/SEA-2 and SEC1-1/SEC1-2, consecutively. The amplification products of 16S rRNA were
sequenced and then analyzed for their relatedness to S. aureus. The resulted sequence was analyzed using BLAST (Basic Local Alignment Search Tool) program and it was found that 5 out of 14 isolates were confirmed as S. aureus. The local isolates of NU4, NU5 and NU9 have 87 – 92% similarity, but all of the local isolates do not have similarity with S. aureus ATCC 25923. Based on amplification of the SE genes, 1 local isolates of S. aureus (NU5) was capable of producing both SEA and SEC1 while 1 local isolate (NU1) was only capable of producing SEA.
aureus Dengan Metode Polymerase Chain Reaction (PCR). Dibimbing oleh RATIH DEWANTI-HARIYADI and SRI HENDRASTUTI HIDAYAT.
Bakteri S. aureus telah banyak dilaporkan sebagai patogen asal pangan yang dapat menyebabkan keracunan pada manusia. Karakteristik S. aureus antara lain termasuk bakteri bulat Gram positif yang bergerombol seperti anggur, membentuk pigmen kuning keemasan, tidak membentuk spora, non-motil, bersifat aerob dan anaerob fakultatif serta membentuk koagulase dan katalase positif. Umumnya S. aureus tidak memiliki kemampuan untuk berkompetisi baik dengan mikroflora normal pada makanan. Keberadaan S. aureus pada pangan umumnya disebabkan oleh kontaminasi silang dari pekerja maupun peralatan pengolahan yang digunakan serta perlakuan pangan setelah diolah. Bakteri ini sendiri ditemukan di dalam saluran pernapasan, permukaan kulit dan rambut hewan berdarah panas termasuk manusia. Lebih dari 30 – 50% populasi manusia adalah “carrier” S. aureus (Le Loir et al., 2003).
Faktor virulensi yang dihasilkan oleh S. aureus sehingga menyebabkan keracunan pangan salah satunya adalah enterotoksin stafilokoki (SE). SE merupakan kelompok protein globular rantai tunggal yang bersifat antigentik dengan berat molekul rendah. SE juga bersifat larut air, termostabil dan kaya akan beberapa residu asam amino. Terdapat 14 jenis enterotoksin stafilokoki yang telah diidentifikasi. SE yang umumnya mengontaminasi makanan diantaranya SEA, SEB, SEC1 dan SED. Hanya sedikit galur yang menghasilkan SEB, proporsinya 1:10 dengan galur penghasil SEA (Bennett dan Amos, 1982). Selain SEA, enterotoksin stafilokoki yang paling sering mengontaminasi produk pangan antara lain SEC1 dan SED. Menurut Pelisser et al., (2009), pada produk keju dan daging ditemukan 10 galur S. aureus yang mengandung SEA dan 12 galur mengandung SED. Sedangkan pada produk susu mentah dan pasteurisasi terdapat 16 galur S. aureus yang mengandung SEA dan 8 galur yang mengandung SEC1 (Rall et al., 2008). Menurut Maslankova et al., (2009), pada produk olahan susu berturut-turut ditemukan S. aureus yang mengandung SEC 24.1%, SEB 13.9% dan SED 10.1%. Berdasarkan perbandingan sekuen asam amino, SEA berada satu kelompok dengan SED, sedangkan SEB berada satu kelompok dengan SEC1 (Argudin et al., 2010).
beras ketan dan setelah diuji laboratorium ternyata positif mengandung S. aureus (Gentina et al., 2008). Menurut US FDA (2001), kasus keracunan makanan biasanya terjadi apabila jumlah S. aureus mencapai 105CFU/g atau lebih. SE dapat menyebabkan keracunan pada dosis yang sangat rendah yaitu 0.1 – 1 µg/kg (ICSMF, 1996). Data mengenai keberadaan bakteri S. aureus di Indonesia berdasarkan gen penyandi SE dengan menggunakan teknik Polymerase Chain Reaction (PCR)yang berasal dari susu segar hewan dan produk pangan olahan asal hewan sudah pernah dilaporkan (Salasi et al., 2009), sedangkan data mengenai keberadaan bakteri S. aureus di Indonesia berdasarkan gen penyandi SE yang berasal dari produk-produk pangan tradisional siap santap ataupun produk-produk olahan pangan lainnya belum pernah dilaporkan.
Penelitian ini dilakukan untuk mengidentifikasi isolat-isolat lokal S. aureus berdasarkan gen 16S rRNA dan mendeteksi keberadaan gen penyandi
SEA serta SEC1 pada isolat-isolat tersebut. Isolasi DNA genom bakteri S. aureus menggunakan metode Doyle dan Doyle (1989) dengan beberapa modifikasi. Amplifikasi gen target menggunakan 3 pasang primer berturut-turut, yaitu 63f/1387r, SEA-1/SEA-1 dan SEC1-1/SEC1-2. Hasil amplifikasi untuk gen 16S rRNA dilakukan sekuensing dan dianalisis menggunakan program BLAST pada situs NCBI (www.ncbi.nlm.nih.gov).
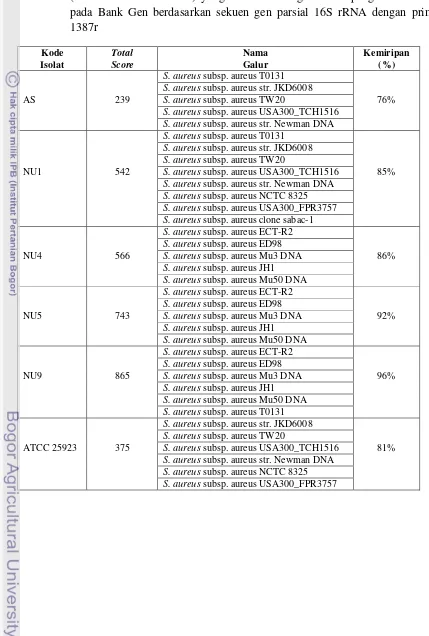
Berdasarkan hasil amplifikasi parsial gen 16S rRNA, 5 dari 14 isolat lokal yang diteliti adalah S. aureus, meskipun secara uji biokimiawi 14 isolat lokal tersebut telah diidentifikasi sebagai S. aureus (Apriyadi, 2010). Dalam penelitian ini, 9 isolat lokal lainnya tidak teridentifikasi sebagai spesies apapun. Tingkat kemiripan tinggi ditemukan pada isolat lokal NU4, NU5 dan NU9, yaitu berkisar antara 87 – 92%. Dapat disimpulkan bahwa 3 isolat lokal tersebut merupakan spesies yang sama. Berdasarkan sumber isolat-isolat lokal S. aureus yang diperoleh, isolat lokal AS yang berasal dari ayam suwir memiliki kemiripan yang relatif rendah (76%) dengan isolat lokal yang berasal dari nasi uduk (NU1). Tidak terdapat kemiripan antara kelima isolat lokal S. aureus dengan S. aureus ATCC 25923 sebagai isolat pembanding.
Hasil amplifikasi gen penyandi SE menunjukkan bahwa dari 2 isolat lokal S. aureus, 1 isolat mengandung kedua gen penyandi SEA dan SEC1 (NU5) dan 1 isolat hanya mengandung SEA (NU1).
METODE
POLYMERASE CHAIN REACTION
(PCR)
FIDYATUN KHOIRIYAH
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Ilmu Pangan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
© Hak cipta milik Institut Pertanian Bogor, tahun 2011
Hak cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber. Pengutipan hanya dilakukan untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah. Pengutipan tidak mengurangi kepentingan yang wajar Institut Pertanian Bogor.
Nama : Fidyatun Khoiriyah NIM : F251070191
Disetujui Komisi pembimbing
Dr. Ir. Ratih Dewanti-Hariyadi, M.Sc Dr. Ir. Sri Hendrastuti Hidayat, M.Sc
Ketua Anggota
Diketahui
Ketua Program Studi Ilmu Pangan Dekan Sekolah Pascasarjana IPB
Dr. Ir. Ratih Dewanti-Haryadi, M.Sc Dr. Ir. Dahrul Syah, M.Sc.Agr
Alhamdulillahi robbil „alamin, puji syukur penulis ucapkan kepada Alloh SWT atas segala nikmat, karunia dan hidayah-Nya sehingga pada akhirnya penulis dapat menyelesaikan karya ilmiah yang berjudul Identifikasi Molekular Isolat Lokal Staphylococcus aureus Dengan Metode Polymerase Chain Reaction (PCR).
Selama perjalanannya menempuh pendidikan S2 ini, penulis merasa dibantu oleh banyak pihak. Oleh karena itu pada kesempatan ini, penulis ingin mengucapkan terima kasih kepada Dr. Ir. Ratih Dewanti-Hariyadi, MSc, selaku ketua komisi pembimbing dan Dr. Ir. Sri Hendrastuti Hidayat, MSc, selaku anggota komisi pembimbing atas segala arahan, bimbingan, semangat, perhatian dan kesabaran penuh seperti layaknya ibu sendiri. Semoga Alloh SWT memberikan balasan yang terbaik atas segala pengorbanan waktu, tenaga, pikiran dan ilmu yang bermanfaat bagi penulis.
Kepada Antung Sima Firlieyanti, STP, MSi, selaku dosen penguji luar komisi yang telah bersedia meluangkan waktu untuk memberi masukan guna perbaikan karya ilmiah ini.
Ucapan terima kasih juga penulis sampaikan kepada SEAFAST Center – LPPM IPB yang telah mendanai penelitian ini dan memberi kemudahan bagi penulis untuk dapat melaksanakan penelitian hingga selesai.
Terima kasih penulis ucapkan kepada: (1) Tim mahasiswa satu bimbingan, yaitu Bombay, Veni, Mbak Desty, Winnie, Ipit, Ivani, Juli dan Ipan, yang telah mengisi hari-hari penulis selama penelitian dan membantu dalam banyak hal, (2) Mbak Ari, Sofah dan Yeris serta para teknisi lainnya di laboratorium SEAFAST Center, (3) Mbak Tuti di Laboratorium Virologi Tumbuhan, HPT IPB, yang telah banyak mengajarkan ilmu dan hal-hal teknis maupun non-teknis tentang bidang molekular, (4) Teman-teman IPN, terutama Zaim dan Uni Rita, yang telah memberi semangat dan banyak membantu di detik-detik terakhir penyelesaian masa studi, (5) Teman-teman di Laboratorium Bioteknologi Pangan SEAFAST Center, Midun, Mbak Dwi, Mbak Elfi, Mbak Maya, Dilla, Yogi, Dita, Victor, Ahmad dan Goy yang selalu memberi semangat dan bantuan, (6) Teman-teman Liqo, serta (7) Semua pihak yang tidak dapat penulis sebutkan satu per satu, terima atas dukungannya.
Tak lupa dengan sangat penulis mengucapkan terima kasih kepada papa, mama, adik-adik dan seluruh keluarga besar yang tak henti-hentinya memberikan doa, kasih sayang, semangat dan bantuan. Terima kasih yang luar biasa teruntuk suami dan mujahid kecilku, Ukasyah, penulis merasa amat sangat bersyukur memiliki kalian berdua. Tidak ada penyesalan atas apa yang sudah terjadi dan pada akhirnya semua dapat terselesaikan juga walaupun harus melewati rintangan yang berliku.
Depok, September 2011
Penulis dilahirkkan di Jakarta pada tanggal 12 Juni 1984 dari Ayah H. Achwani dan Ibu Hj. Atika Nurkhasanah. Penulis merupakan anak sulung dari tiga bersaudara.
Tahun 2002, penulis lulus dari SMU Negeri 112 Jakarta dan pada tahun yang sama terdaftar sebagai mahasiswa di Fakultas Biologi Universitas Nasional Jakarta. Selama menempuh bangku kuliah, penulis mendapat beasiswa dari NEF (Nagao Environmental Foundation).
DAFTAR TABEL….……….
Faktor-Faktor yang Mempengaruhi Produksi Enterotoksin Stafilokoki (SE)
Mekanisme Aksi SE………...
Persiapan dan Pewarnaan Gram Kultur Bakteri S. aureus………..
Amplifikasi Gen SEA dan SEC1……….. SIMPULAN DAN SARAN………
Simpulan………. Saran………. DAFTAR PUSTAKA……… LAMPIRAN………...
7 Data cemaran S. aureus pada beberapa pangan... Faktor-faktor yang berkontribusi pada kasus keracunan pangan... Karakteristik utama enterotoksin stafilokoki (SE)... Persentase identitas asam amino pada SE yang berbeda... Support genetika pada beberapa gen penyandi SE... Faktor-faktor yang mempengaruhi produksi enterotoksin yang dihasilkan oleh S. aureus... Pasangan primer oligonukleotida yang digunakan untuk amplifikasi gen target... Produk sekuensing dari isolat-isolat lokal S. aureus dan isolat pembanding (S. aureus ATCC 25923) berdasarkan sekuen gen
parsial 16S rRNA dengan primer 63f, 1387r dan hasil contig……
Persen kemiripan isolat-isolat lokal S. aureus dan isolate pembanding (S. aureus ATCC 25923) yang dianalisis dengan beberapa galur S. aureus pada Bank Gen berdasarkan sekuen gen
parsial 16S rRNA dengan primer 1387r……….
Sumber isolat dan asal negara dari masing-masing galur S. aureus berdasarkan sekuen gen parsial
16S rRNA………....
Persen kemiripan isolat-isolat lokal S. aureus dan isolat pembanding (S. aureus ATCC 25923) yang dianalisis dengan beberapa galur S. aureus pada Bank Gen berdasarkan sekuen gen
parsial 16S rRNA………...
6 Morfologi S. aureus perbesaran 5000x... Mekanisme aksi enterotoksin stafilokoki (SE)...
Diagram alir pelaksanaan penelitian... Morfologi bakteri S. aureus hasil pewarnaan Gram………... Visualisasi total DNA genom isolat-isolat lokal S. aureus dan isolat luar S. aureus ATCC 25923. Sampel (a) dan (b) terdiri dari: ATCC 25923 (1), AS (2), NU1 (3), NU2 (4), NU3 (5), NU4 (6), NU5 (7), NU6 (8), NU7 (9), NU8 (10), NU9 (11), NU10 (12), NU11 (13), NU13 (14) dan
NU14 (15). M adalah DNA ladder 1 kb sebagai penanda DNA…………
Visualisasi DNA hasil amplifikasi fragmen gen 16S rRNA universal isolat lokal S. aureus. Sampel (a) dan (b) terdiri dari: ATCC 25923 (1), AS (2), NU1 (3), NU2 (4), NU3 (5), NU4 (6), NU5 (7), NU6 (8), NU7 (9), NU8 (10), NU9 (11), NU10 (12), NU11 (13), NU13 (14) dan NU14 (15). M adalah DNA ladder 1 kb sebagai penanda DNA dan K- sebagai
kontrol negatif……….
Visualisasi DNA hasil amplifikasi fragmen gen penyandi SEA isolat lokal S. aureus. Sampel terdiri dari: ATCC 25923 (1), AS (2), NU1 (3), NU3 (4), NU4 (5), NU5 (6), NU6 (7), NU7 (8), NU8 (9), NU9 (10), NU11 (11) dan NU13 (12). M adalah DNA ladder 100 bp sebagai penanda DNA dan K- sebagai kontrol negatif………...
68
69
76
84 Konsentrasi dan kemurnian DNA genom bakteri S. aureus………
Urutan basa (nukleotida) gen 16S rRNA menggunakan primer 63f untuk isolat-isolat lokal S. aureus dan isolat pembanding S. aureus 25923 yang disekuensing... Urutan basa (nukleotida) gen 16S rRNA menggunakan primer 1387r (reverse complement) untuk isolat-isolat lokal S. aureus dan isolat pembanding S. aureus 25923 yang disekuensing………... Urutan basa (nukleotida) gen 16S rRNA hasil contig menggunakan primer 63f dan 1387r untuk isolat-isolat lokal S. aureus dan isolat pembanding S. aureus 25923…………...……… 1
2
3
4
Latar Belakang
Keracunan pangan akibat kontaminasi bakteri patogen atau dikenal dengan istilah foodborne disease (FBD) merupakan permasalahan keamanan pangan yang menjadi perhatian dunia, tidak terkecuali Indonesia. Padahal dalam mengupayakan keamanan pangan tersebut, masing-masing negara telah membuat kebijakan seperti Undang-undang Perlindungan Konsumen, Peraturan Pemerintah tentang Pelabelan Pangan, Manajemen Keamanan Pangan dan berbagai panduan lainnya. Namun demikian, masih saja terdapat kejadian dimana cemaran pangan kurang dapat diantisipasi secara sempurna sehingga menimbulkan kekacauan, seperti temuan Staphylococcus aureus pada pangan siap santap.
Staphylococcus aureus adalah bakteri Gram positif, berbentuk bulat bergerombol seperti anggur dan tidak membentuk spora sehingga sangat mudah diinaktifkan dengan perlakuan panas. S. aureus merupakan bakteri yang umum terdapat pada manusia dan bersifat patogen yang dapat menyebabkan keracunan pangan. Keracunan yang disebabkan oleh bakteri ini tergolong dalam kasus intoksikasi, yaitu tertelannya enterotoksin yang dihasilkan oleh S. aureus pada pangan. Menurut Pelczar dan Chan (2005), gejala umum keracunan enterotoksin stafilokoki berupa mual, pusing, muntah dan diare. Enterotoksin stafilokoki dapat menyebabkan keracunan pada dosis yang sangat rendah, yaitu 0.1-1 µg/kg (ICSMF, 1996). Gejala keracunan dapat terlihat 30 menit hingga 8 jam setelah mengonsumsi makanan yang mengandung toksin tersebut (Blackburn dan Mc Clure, 2002).
food-borne disease. Hal ini terutama terjadi pada negara berkembang, seperti
Indonesia dan tidak menutup kemungkinan juga terjadi pada negara maju.
Berdasarkan laporan the Centers for Disease Control and Prevention (CDC) (1989) menyatakan bahwa pada tahun 2001 hingga 2005 rata-rata setiap tahun terjadi 327 kasus keracunan pangan akibat bakteri di Amerika Serikat, 15% diantaranya disebabkan oleh enterotoksin stafilokoki. Sementara itu pada tahun 2006, terjadi 295 kasus keracunan pangan akibat bakteri dan 9.8% diantaranya juga disebabkan oleh enterotoksin stafilokoki (SE). Tidak hanya kasus tersebut, di Amerika Serikat yang tergolong negara maju, foodborne disease diduga bertanggung jawab terhadap 76 juta kasus sakit, 325,000 kasus rawat inap dan 5,000 kasus kematian per tahunnya (Jaykus, 2003).
Kejadian Luar Biasa (KLB) lainnya yang pernah dilaporkan antara lain di Osaka Jepang pada tahun 2000, yaitu produk susu skim bubuk dan susu rekonstitusi ditemukan mengandung enterotoksin stafilokoki A (SEA). SEA yang terkandung mencapai 80 ng. Selanjutnya di Amerika Serikat, pada produk susu coklat cair mengandung 200 ng atau kurang SEA (Ikeda et al., 2005).
KLB akibat S. aureus di Indonesia sendiri pernah terjadi di beberapa daerah. Di Tabanan Bali pada tahun 2004, 159 orang dilaporkan keracunan akibat mengonsumsi nasi bungkus dan hasil pengujian laboratorium BPOM menunjukkan bahwa sisa nasi bungkus dan muntahan korban mengandung mikroba patogen S. aureus. Bakteri patogen ini juga ditemukan pada jajanan jeli yang mengakibatkan 20 orang murid di Gresik keracunan. Pada tahun yang sama juga terjadi kasus keracunan di Bandar Lampung karena mengonsumsi campuran nasi dan ikan tongkol. Sementara itu pada tahun 2007, terjadi keracunan makanan pada 36 orang setelah mengonsumsi camilan di sebuah hotel di Padang. Hasil uji klinis laboratorium menunjukkan sampel makanan positif mengandung S. aureus (Gentina et al., 2008).
Keracunan pangan oleh S. aureus disebabkan oleh enterotoksin. Enterotoksin stafilokoki merupakan kelompok protein globular rantai tunggal yang bersifat antigenik dengan berat molekul rendah, yaitu 26,900-29,600 dalton.
larut air dan stabil terhadap panas (termostabil) serta kaya akan residu lisin, asam aspartat, asam glutamat, dan tirosin (Le Loir et al., 2003). Terdapat 14 jenis SE yang telah diidentifikasi, yaitu SEA SEB, SEC, SED, SEE, SEG, SEH, SEI, SEJ, SEK, SEL, SEM, SEN, dan SEO. Namun hanya SEA, SEB, SEC, SED, dan SEE yang hingga saat ini dapat dideteksi dengan peralatan komersial (Ikeda et al., 2005). Hanya sedikit galur yang menghasilkan SEB, proporsinya 1:10 dengan galur penghasil SEA (Bennett dan Amos, 1982). Selain SEA, enterotoksin stafilokoki yang paling sering mengontaminasi produk pangan antara lain SEC dan SED. Menurut Pelisser et al., (2009), pada produk keju dan daging ditemukan 10 galur S. aureus yang mengandung SEA dan 12 galur mengandung SED. Sedangkan pada produk susu mentah dan pasteurisasi terdapat 16 galur S. aureus yang mengandung SEA dan 8 galur yang mengandung SEC (Rall et al., 2008). Menurut Maslankova et al., (2009), pada produk olahan susu berturut-turut ditemukan S. aureus yang mengandung SEC 24.1%, SEB 13.9% dan SED 10.1%. Berdasarkan perbandingan sekuen asam amino, SEA berada satu kelompok dengan SED, sedangkan SEB berada satu kelompok dengan SEC1 (Argudin et al., 2010).
Enterotoksin stafilokoki sangat stabil terhadap enzim proteolitik, seperti pepsin dan tripsin, sehingga toksin ini tetap aktif di saluran pencernaan. Enterotoksin ini juga tahan terhadap kimotripsin, renin, dan papain. Meskipun demikian, SEB dan SEC1 dapat dipotong pada loop sistein oleh tripsin. SEB dapat dihancurkan oleh pepsin pada pH 2, tetapi SEB menjadi resisten terhadap pepsin pada pH yang lebih tinggi, dimana kondisi tersebut adalah kondisi normal di dalam lambung setelah seseorang mengonsumsi makanan (Bhatia dan Zahoor, 2007).
(masing-masing 1 isolat); SEC dan SEE (2 isolat); SEB dan SEC (4 isolat). Kombinasi 3 jenis gen penyandi SE ditemukan masing-masing sebanyak 1 isolat untuk SEB, SEC dan SEI; SEC, SEE dan SEI serta SEC, SEG dan SEI. Data mengenai keberadaan bakteri S. aureus di Indonesia berdasarkan gen penyandi SE yang berasal dari produk-produk pangan tradisional siap santap ataupun produk-produk olahan pangan lainnya belum pernah dilaporkan.
Laporan resmi tentang kasus keracunan stafilokoki di Indonesia masih terbatas jumlahnya. Data yang tersedia umumnya hanya menyatakan lokasi kejadian dan jumlah korban, namun tidak sampai pada menjelaskan penyebabnya. Hal ini menyebabkan monitor dan evaluasi apabila terjadi suatu kejadian luar biasa (KLB) berdasarkan asal-muasal keracunan (etiologi), sumber makanan dan tempatnya pun belum terlaksana dengan baik. Badan Pengawasan Obat dan Makanan (BPOM) melaporkan bahwa kasus keracunan pangan di Indonesia paling banyak terjadi pada pangan olahan yang melibatkan industri jasa boga dan industri jasa katering. Data mengenai prevalensi isolat lokal S. aureus dan keracunan stafilokoki yang ditimbulkannya pada produk pangan tradisional santap belum tersedia. Oleh karena itu perlu diketahui frekuensi keberadaan isolat lokal S. aureus dalam produk pangan tradisional siap santap dan toksin yang dihasilkannya guna dijadikan acuan dalam penetapan manajemen risiko baik di tingkat produsen maupun konsumen.
Tujuan Penelitian Penelitian ini bertujuan untuk :
(1) Mengidentifikasi isolat-isolat lokal S. aureus berdasarkan gen 16S rRNA, sehingga dapat diketahui persentase tingkat kemiripan antar isolat-isolat tersebut.
Manfaat Penelitian
Hasil yang diperoleh dari penelitian ini diharapkan dapat menambah informasi (database) tentang S. aureus yang ada di Indonesia untuk melakukan kajian risiko S. aureus isolat lokal yang ada di Indonesia.
Hipotesis
TINJAUAN PUSTAKA
Karakteristik Staphylococcus aureus
Staphylococcus aureus adalah bakteri koki Gram positif dan jika diamati di
bawah mikroskop akan tampak dalam bentuk bulat tunggal atau berpasangan, atau berkelompok seperti buah anggur seperti yang terlihat pada Gambar 1. Staphylococcus aureus termasuk dalam famili Staphylococcaceae, berukuran
diameter 0.5-1.5 µm dan membentuk pigmen kuning keemasan. Bakteri ini tidak membentuk spora, bersifat aerob atau anaerob fakultatif, non-motil, koagulase dan katalase positif, mampu memfermentasi mannitol serta mampu menjalankan dua macam metabolisme yaitu respirasi maupun fermentasi.
Gambar 1 Morfologi S. aureus perbesaran 5000x (Todar, 2008)
S. aureus mampu memanfaatkan berbagai komponen organik sebagai nutrisi
untuk pertumbuhannya. Asam-asam amino dibutuhkan sebagai sumber nitrogen, sedangkan tiamin dan asam nikotinat paling dibutuhkan diantara vitamin B lainnya. Apabila S. aureus ditumbuhkan pada kondisi cenderung anaerob, maka urasil sangat dibutuhkan. Sedangkan untuk kondisi aerob dan produksi enterotoksin, maka monosodium glutamat berperan sebagai sumber C, N dan energi. Arginin merupakan asam amino esensial yang dibutuhkan untuk produksi enterotoksin B (Bennet dan Monday dalam Militois dan Bier, 2003; Jay, 2000).
S. aureus termasuk ke dalam kelompok bakteri mesofilik, namun terdapat
pertumbuhan 30-37oC. Kisaran pH pertumbuhan antara 4.5 hingga 9.3, dengan pH optimum 7.0-7.5 (Bennet dan Monday, 2003). Berdasarkan aktivitas air (aw), stafilokoki mampu tumbuh pada kadar aw yang lebih rendah dibandingkan dengan bakteri nonhalofilik lainnya. Pertumbuhan stafilokoki tetap terjadi pada aw 0.83 yang merupakan kondisi di bawah ideal untuk pertumbuhan kebanyakan bakteri. Kebanyakan galur-galur S. aureus mempunyai toleransi tinggi terhadap konsentrasi garam dan gula. Bakteri ini masih dapat bertahan hidup pada konsentrasi natrium klorida lebih dari 15% dan memiliki toleransi tinggi terhadap komponen-komponen seperti telurit, merkuri klorida, neomycin, polymixin dan sodium azida, yang semuanya dapat digunakan sebagai media selektif S. aureus (Le Loir et al., 2003). Faktor-faktor yang mempengaruhi pertumbuhan S. aureus dapat dilihat pada Tabel 1.
Tabel 1 Faktor-faktor yang mempengaruhi pertumbuhan S. aureus
Faktor Pengaruh
Pertumbuhan
Optimum Kisaran
Suhu 37°C 4 –48°C
pH 6.0-7.0 4.0-9.8
aw 0.98≥0.99 0.83≥0.99
Atmosfer Aerobik Anaerobik hingga aerobik Natrium Klorida 0.5-0.4% 0-20%
Adam dan Moss (1995)
S. aureus mampu memproduksi sejumlah besar enzim-enzim ekstraseluler,
dapat memecah DNA maupun RNA untuk menghasilkan produk fosfomononukleotida.
Tabel 2 Karakteristik spesies stafilokoki
Sifat S. aureus S. intermedius S. hyicus S. epidermidis
Pigmen +a − − −
Koagulase + + ± −
Dnase + + ± −
Hemolisis + + − ±
Mannitol (an) + − − −
Acetoin + − − +
Gumpalan + + + −
Hyaluronidase + − ± −
Lysostaphyn ST ST ST SR
a
>90%.
an:kondisi anaerobik; ST: sensitivitas tinggi; SR: sensitivitas rendah
Bennet dan Monday (2003)
Staphylococcus aureus tidak membentuk spora sehingga pertumbuhan oleh
S. aureus di dalam makanan dapat segera dihambat dengan perlakuan panas.
Namun, kontaminasi S. aureus tetap menjadi salah satu penyebab utama foodborne disease (FBD) karena S. aureus dapat mengkontaminasi produk
makanan selama persiapan dan pengolahan. Bakteri ini sendiri ditemukan di dalam saluran pernapasan, permukaan kulit dan rambut hewan berdarah panas
termasuk manusia, Lebih dari 30-50% populasi manusia adalah “carrier” S. aureus (Le Loir et al., 2003).
Aeromonas, Bacillus, Pseudomonas, S. epidermidis, kelompok Enterobactericeae,
Lactobacillaceae, kelompok Enterococci dan Streptococcus (Jay, 1996).
Staphylococcus aureus memliki beberapa jenis faktor virulensi yang mendukung terjadinya penyakit pada tubuh manusia, salah satunya adalah protein permukaan yang membantu kolonisasi pada jaringan inang. Untuk mendukung penyebarannya pada jaringan, bakteri ini menghasilkan invasin, seperti leukosidin, kinase dan hyaluronidase. Leukosidin adalah sitotoksin yang dapat membunuh leukosit, sedangkan hyaluronidase adalah enzim yang dapat mendegradasi asam hyaluronat sehingga meningkatkan permeabilitas jaringan (Todar, 2008).
Pada tubuh manusia terdapat faktor virulensi S. aureus yang mampu menghambat fagositosis, yaitu protein A dan kapsul. Protein A adalah protein permukaan yang mengikat molekul IgG. Bakteri ini juga menghasilkan senyawa biokimia yang menjadi pertahanannya terhadap fagosit, yaitu karotenoid dan katalase (Todar, 2008).
Kontaminasi S. aureus dalam Pangan
Stafilokoki dapat ditemukan di udara, debu, limbah, air, susu, makanan, atau peralatan makan, permukaan lingkungan, manusia, dan hewan. Manusia dan hewan adalah reservoir utama. Stafilokoki juga dapat ditemukan lebih dari 50% di saluran pernapasan, tenggorokan dan di permukaan rambut dan kulit seseorang yang sehat sekalipun.
Keberadaan S. aureus dalam bahan pangan erat kaitannya dengan sanitasi pekerja serta kebersihan lingkungan dan peralatan pengolahan (Stewart et al., 2002). Pangan yang dilaporkan dalam berbagai kejadian luar biasa S. aureus umumnya diolah dengan proses pemotongan, pemarutan, dan pengilingan yang melibatkan pekerja yang terkontaminasi. S. aureus terdapat luas di alam dan pada bahan baku pangan sehingga penanganan yang kurang tepat dapat meningkatkan risiko keracunan pangan akibat S. aureus (Robinson et al., 2000).
tinggi). Hal ini berkaitan dengan sifat S. aureus yang merupakan kompetitor lemah dalam ekosistem mikrobial yang kompleks sehingga adanya bakteri patogen dan pembusuk lain dapat menghambat pertumbuhannya. Bakteri psikrotropik sebagai contohnya dapat menghambat pertumbuhan S. aureus pada penyimpanan suhu rendah (refrigerasi). Selain itu pada proses fermentasi, bakteri asam laktat dapat memproduksi beberapa senyawa yang mampu menghambat pertumbuhan S. aureus seperti asam laktat, hidrogen peroksida dan bakteriosin (Ash, 2000).
US FDA (1999) menyatakan keberadaan S. aureus dalam pangan dapat disebabkan oleh beberapa hal seperti proses pengolahan pangan yang tidak tepat dari segi suhu dan waktu, suhu penyimpanan pangan yang salah yaitu kurang panas (60°C atau 140°F) atau kurang dingin (7.2°C atau 45°F), dan adanya kontaminasi silang dari bahan baku mentah maupun pekerja. Penjelasan mengenai konsekuensi yang ditimbulkan apabila proses pengolahan dan penanganan pangan tidak dilakukan dengan tepat dapat dilihat pada Tabel 3.
Jalur masuknya S. aureus kedalam bahan pangan biasanya melalui jaringan kulit atau selaput lendir yang terluka seperti terpotong benda tajam, luka bakar, gigitan serangga, pengelupasan kulit, atau penyakit kulit lain. Oleh sebab itu, pekerja dengan luka pada kulit tidak diperbolehkan mengolah makanan terlebih disertai dengan praktik sanitasi yang buruk yang dapat memperbanyak jumlah S. aureus (Schaechter et al., 1993).
Apabila S. aureus terkontaminasi ke dalam bahan pangan yang mengandung nutrisi yang menunjang bagi pertumbuhannya, jumlah S. aureus akan bertambah dengan laju pertumbuhan yang cepat. Bahan pangan yang menyediakan nutrisi yang menunjang pertumbuhan S. aureus adalah bahan pangan dengan kadar protein yang tinggi seperti daging dan produk olahannya, unggas dan produk olahannya, telur dan produk olahannya, salad yang mengandung telur, tuna, ayam, kentang dan makaroni, produk bakery, serta susu dan produk olahannya (US FDA, 1999). Hal ini disebabkan adanya 11 asam amino yaitu valin, leusin, threonin, phenilalanin, tirosin, sistein, metionin, lisin, prolin, histidin dan arginin
Tabel 3 Sumber, faktor resiko, dan konsekuensi keberadaan S. aureus dalam rantai pangan
Sumber Resiko Konsekuensi jika
resiko tidak terkontrol
(hingga 20%). Cara yang dianjurkan untuk mengontrol atau mencegah adanya S. aureus pada pangan yang sudah diolah adalah mendinginkannya hingga suhu
20°C, melarang orang sakit untuk menangani makanan, menggunakan sarung tangan disposable dalam proses penanganan makanan, melakukan refrigerasi atau pendinginan pada makanan hingga <10°C dan meminimalkan terjadinya kontaminasi silang dari bahan mentah ke makanan matang ataupun dari lingkungan kerja yang kotor serta peralatan yang kotor ke makanan matang (Jay, 2000).
Harmayani et al., (1996) mengevaluasi total bakteri dan S. aureus pada beberapa sampel pangan menggunakan media Baird-Parker Agar (BPA) yang dilanjutkan dengan uji koagulase. Dalam penelitian tersebut ditemukan bahwa karkas ayam mentah yang digunakan sebagai bahan dasar pembuatan ayam panggang bumbu sate memiliki total bakteri sebanyak 6.5 x 107 CFU/g dan total S. aureus 7.3 x 105 CFU/g. Karkas ayam mentah diproses melalui tahap pencucian dan perebusan. Pada akhir tahap perebusan, ternyata total bakteri menurun menjadi 1.7 x 106 CFU/g dan total S. aureus <103 CFU/g. Setelah pembakaran,
total S. aureus berkurang lagi menjadi 5.0 x 102 CFU/g. Namun populasi S. aureus menjadi 1.5 x 104 CFU/g selama proses pengangkutan dan penyimpanan
pada suhu ruang selama 7.5 jam. Jumlah ini telah melewati batas maksimum cemaran S. aureus yang telah ditetapkan oleh Badan Pengawas Obat dan Makanan (BPOM), yaitu 0-5 x 103 sel/g.
Tabel 4 Data cemaran S. aureus pada beberapa pangan
Karkas Ayam 5.15 c Harmayani et al., (1996)
Ayam Goreng 2.64 d Sari (2010)
Ayam Kecap 3.22 d
Ayam Opor 3.66 d
Ayam Balado 2.36 d
* Dengan media Vogel-Johnson Agar (VJA), sampel diambil jam 11 siang ** Dengan media Baird-Parker Agar (BPA), sampel diambil 2-3 jam sejak penjaja mulai berjualan
Perilaku S. aureus dalam Pangan
Bakteri S. aureus yang tidak membentuk spora adalah termasuk bakteri yang mudah diinaktivasi oleh panas. Ketahanan panas lebih tinggi terutama pada pangan dengan aktivitas air tinggi (Stewart, 2003). Jika dibandingkan dengan bakteri lainnya S. aureus memiliki ketahanan panas yang cukup tinggi pada suhu 62.8°C. S. aureus lebih tahan terhadap pemanasan pada heating menstruum susu dengan suhu 62.8°C jika dibandingkan dengan bakteri nonspora lainnya seperti, E. coli, Campylobacter jejuni, Streptococcus. faecalis, dan Lactobacillus lactis.
Akan tetapi, S. aureus tidak lebih tahan panas dibandingkan dengan spora bakteri seperti spora Bacillus cereus, dan Clostridium botulinum.
0.39 menit. Nilai Z yang dihasilkan sebesar 6.04°C. Sedangkan S. aureus 196E mempunyai D60 sebesar 3.44 menit dan D65.6 sebesar 0.28 menit. Nilai Z yang diperoleh sebesar 5.10°C. Dari percobaan ini, diketahui bahwa bakteri ini bisa direduksi dengan pasteurisasi pada suhu 63°C selama 15 menit.
Walker dan Harmon (1966) juga menyelidiki ketahanan panas strain S. aureus pada susu murni, susu skim, whey keju ceddar, dan fosfat buffer. Strain
yang diujikan dalam penelitian ini meliputi isolat 161-C, S-1, B-120, dan S-18. Isolat B-120, dan S-18 hanya diujikan pada heating menstruum fosfat dan susu murni. Suhu perlakuan dalam percobaan berkisar antara 52-62°C. Nilai D hasil percobaan Walker dan Harmon berkisar antara 0.20-3.50 menit untuk isolat 161-C. Kisaran D-value untuk isolat S-1, B-120, dan S-18 berturut-turut sebesar 0.15-3.0 menit, 0.40-1.50 menit, dan 0.50-2.55 menit.
Eden et al., (1977) mempelajari ketahanan panas strain Staphylococcus aureus yang diisolasi dari susu mentah dengan metode tabung kapiler. Heating
menstruum yang digunakan adalah susu skim dengan jumlah mikroba awal
1.0 x 109 CFU/ml. Pemanasan dilakukan pada kisaran suhu 50-75oC. Nilai Z S. aureus sebesar 9.4oC. Nilai D yang dihasilkan dari percobaan Eden et al., (1977) berkisar antara 0.02-9.96 menit.
Ketahanan S. aureus dalam susu kambing dipelajari oleh Parente dan Mazzatura (1991). Dalam percobaan ini digunakan isolat BP3 dan isolat 237. Metode percobaan menggunakan metode tabung kapiler dengan jumlah mikroba awal >1.0 x 109 CFU/ml. Suhu yang digunakan berkisar antara 55-68 oC. Nilai D isolat BP3 berkisar antara 0.03-3.30 menit sedangkan isolat 237 memiliki D-value sekitar 0.01-10.60 menit. Nilai Z sebesar 4.83 ± 0.06 untuk isolat BP3 dan 4.50 ± 0.05 untuk isolat 237.
El-Banna et al., (1983) menunjukkan bahwa strain S. aureus yang tumbuh di bawah kondisi stress memilki ketahanan panas lebih tinggi dibandingkan dengan yang tumbuh pada lingkungan yang mendukung pertumbuhannya. Nilai D kultur S. aureus yang tumbuh pada suhu 37oC memiliki D60 sebesar 2.73 menit, sebaliknya yang tumbuh pada suhu 45 oC memiliki D60 sebesar 12.6 menit.
mengetahui pengaruh pembekuan terhadap ketahanan panas S. aureus. Isolat S. aureus positif koagulase diujikan pada media pemanas TSB kemudian
dicawankan pada media Baird Parker Agar (BPA), dan media campuran antara Trypticase Soy Agar (TSA) dengan media BPA. Dua perlakuan diujikan pada
isolat. Perlakuan pertama, isolat diuji ketahanan panasnya secara langsung. Perlakuan kedua, isolat terlebih dahulu diberi perlakuan pendinginan kemudian dipanaskan. Hasil dari dua perlakuan kemudian dibandingkan untuk mengetahuai pengaruh perlakuan pendinginan awal terhadap ketahanan panas. Berdasarkan percobaan Kennedy diperoleh kesimpulan bahwa perlakuan pembekuan pendahuluan tidak menghasilkan nilai yang berbeda secara signifikan.
Penyakit karena S. aureus
Kontaminasi S. aureus pada makanan dapat menyebabkan keracunan (intoksikasi). Hal ini disebabkan karena bakteri tersebut mampu menghasilkan toksin yang berupa enterotoksin di dalam saluran pencernaan. Enterotoksin dapat diproduksi apabila kondisi lingkungan mendukung untuk pertumbuhan dan perkembangan bakteri tersebut, seperti pH dan suhu (Miliotis dan Bier, 2003).
Gejala awal dari keracunan makanan akibat enterotoksin stafilokoki umumnya berlangsung selama 2-6 jam atau pada masa inkubasi 30 menit sampai 7 jam setelah mengkonsumsi makanan yang mengandung enterotoksin (atau rata-rata terjadi pada 2-4 jam), tergantung dari ketahanan individu masing-masing.
Gejala keracunan ditandai dengan mual, muntah, kram perut dan prostrasi. Pada kasus yang lebih serius ditandai dengan pusing, kram otot dan perubahan transient pada tekanan darah. Pemulihan umumnya terjadi selama 2-3 hari atau lebih. Kematian akibat enterotoksin stafilokoki jarang terjadi (Miliotis dan Bier, 2003).
berbeda tergantung dari jumlah makanan yang dimakan dan kerentanan terhadap toksin.
Dosis Infeksi S. aureus
Staphylococcus aureus menghasilkan enterotoksin yang menyebabkan
gastroenteritis. Jumlah sel yang diperlukan oleh Staphylococcus aureus untuk menghasilkan racun yang cukup sehingga bersifat meracuni adalah 106 CFU/g (Buckle et al., 1987; Jay, 2000). Namun berdasarkan hasil penelitian yang dilakukan oleh Harmayani et al., (1996), enterotoksin belum dapat terdeteksi pada total populasi S. aureus mencapai >106 CFU/g. Pada kasus-kasus keracunan makanan, biasanya jumlah S. aureus mencapai 108 CFU/g atau lebih.
Shapton dan Shapton (1993) menyatakan bahwa populasi Staphylococcus aureus yang diperlukan untuk menghasilkan toksin adalah 5 x 106 CFU/g, dimana toksin yang dihasilkan bersifat tahan panas. Oleh karena itu, walaupun bakterinya sudah mati karena pemanasan kemungkinan toksinnya masih tetap dapat bertahan. Menurut Ray dan Bhunia (2008) keracunan Staphylococcus aureus disebakan karena terkonsumsinya toksin dalam jumlah 100-200 ng yang dihasilkan oleh 106-107 CFU/ml atau CFU/g dalam 30 g/ml makanan. Pada individu yang memiliki sensitivitas tinggi, dosis 100-200 ng sudah dapat menyebabkan penyakit (Miliotis dan Bier, 2003).
US FDA (2001) menyatakan bahwa jumlah toksin Staphylococcus aureus
yang diperlukan untuk menyebabkan keracunan pangan sebesar 1.0 g. Pada level
ini dicapai jumlah bakteri sebanyak 1.0 x 105 CFU/g atau CFU/ml.
Keracunan Makanan oleh Stafilokoki
(Ray dan Bhunia, 2008). Kemampuan galur Staphylococcus aureus untuk tumbuh dan memproduksi enterotoksin pada kisaran kondisi lingkungan yang luas, ketahanan panas toksin, dan penanganan yang salah menjadi penyebab utama kasus keracunan pangan di berbagai dunia (Ray dan Bhunia, 2008).
Tabel 5 Faktor-faktor yang berkontribusi pada kasus keracunan pangan
Faktor Persentase
Faktor yang berhubungan dengan pertumbuhan mikroba
Penyimpanan makanan pada suhu ruang 43
Suhu pembekuan yang tidak tepat 32
Penyiapan makanan yang terlalu lama saat penyajian 41 Holding pada suhu panas yang tidak cukup 12 Thawing yang tidak tepat 4 Penyajian makanan dalam jumlah yang terlalu banyak 22 Faktor yang berhubungan dengan ketahanan mikroba
Pemanasan ulang yang tidak tepat 17
Pemanasan yang tidak cukup 13
Pembersihan Peralatan pengolahan yang tidak tepat 7
Sumber yang tidak aman 5
Kontaminasi makanan kaleng 2
Forsythe (2000)
Pada tahun 1989 di Starkville, Mississippi, terjadi kasus keracunan pangan S. aureus yang disebabkan karena konsumsi jamur kaleng (CDC, 1989). Sebanyak
22 orang mengalami gastroenteris selama beberapa jam setelah memakan makanan di kafetaria kampus. Gejala keracunan yang terjadi meliputi mual-mual, muntah, diare, dan kejang perut. Sebanyak sembilan orang korbannya dilarikan ke rumah sakit. Setelah diidentifikasi, ditemukan adanya enterotoksin stafilokoki A pada sampel jamur dalam omlet bar.
sama. Pada ham yang tersisa, muntahan pasien, serta hidung dan tenggorokan pasien ditemukan S. aureus (Bergdoll, 1992)
Pada bulan September 1997, terjadi kasus keracunan pangan di Florida (USA) karena konsumsi ham yang terkontaminasi toksin S. aureus. Sebanyak 31 orang dari 125 orang yang mengikuti pesta mengalami keracunan. Gejala keracunan yang terlihat meliputi mual (94%), muntah (89%), diare (72%), berkeringat (61 %), menggigil (44 %), lesu (39 %), kram otot (28 %), pusing kepala (11%) dan demam (11 %). Gejala keracunan berlangsung selama 3-6 jam setelah mengkonsumsi ham dan berakhir setelah 24 jam. Ternyata, sehari sebelum pesta, sebanyak 8 kg ham mentah dan packed ham dipanggang selama 1.5 jam pada suhu 204°C. Setelah dipanggang, ham diiris dengan slicer yang tidak bersih. Ham yang telah dipotong ditempatkan di wadah plastik yang dilapisi alumunium foil, dan disimpan selama 6 jam dalam lemari pendingin. Di hari selanjutnya ham disajikan dalam keadaan dingin. Kemungkinan ham terkontaminasi S. aureus selama pemotongan dengan slicer (Bergdoll, 1992).
Pada tahun 1996, di Institut Robert Koch, Wernigerode, Jerman dilaporkan terjadi kasus keracunan pangan yang disebabkan oleh konsumsi Schwarzwalder Schinken. Produk ham (sekurang-kurangnya pada 6 batch berbeda) diketahui
terkontaminasi oleh enterotoksin S. aureus. Investigasi lebih lanjut memberikan beberapa kesimpulan bahwa kontaminasi terjadi karena rendahnya praktik higiene pada area produksi; produk ham terkontaminasi S. aureus dalam jumlah yang cukup tinggi dan hampir semua isolat yang diuji menghasilkan enterotoksin (Bergdoll, 1992).
tersebut tidak menggunakan sistem pembersih dan disinfeksi otomatis. S. aureus dalam jumlah besar terdeteksi di bagian pipa pengolahan. Hal ini terjadi karena pipa pengolahan tidak dibersihkan selama 3 minggu (Bergdoll, 1992).
Bulan Maret dan April tahun 2002, kasus keracunan pangan akibat S. aureus terjadi di Australia. Kasus ini mengakibatkan sebanyak 250 orang
menjadi korban. Sekitar 600 orang berpartisipasi dalam kegiatan di Imam Ali Islamic Centre, Victoria. Pada tempat tersebut disajikan makanan yang terdiri atas nasi, kentang, dan daging. Beberapa orang langsung mengkonsumsi makanan tersebut dan sebagian lainnya membawa makanannya ke rumah. Beberapa orang yang memakan makanannya di rumah mengalami keracunan. Lebih dari 100 pasien dilarikan ke rumah sakit (Bergdoll, 1992).
Sementara di Indonesia tidak semua kasus keracunan pangan dilaporkan. Menurut Kusumaningrum (2009), nasi bungkus pernah beberapa kali dilaporkan, seperti kasus keracunan di Tasikmalaya tahun 2009 yang memakan korban sebanyak 148 orang. Pada nasi bungkus tersebut ditemukan S. aureus. Pada tahun 2005, sebanyak siswa di Sanggau, Kalimantan Barat juga keracunan setelah mengonsumsi nasi bungkus yang kemudian ditemukan mengandung S. aureus dan Streptococcus faecalis.
Pada tahun 2007 terjadi keracunan makanan pada 36 orang setelah mengonsumsi camilan yang terbuat dari beras ketan di sebuah hotel di Kota Padang. Hasil uji klinis di laboratorium menunjukkan positif S. aureus (Gentina et al., 2008).
Metode Deteksi S. aureus
Beberapa metode yang dapat digunakan untuk mendiagnosis keberadaan S. aureus meliputi metode seleksi media, platting secara langsung dan isolasi pada
media pengayaan. Metode isolasi pada media pengayaan dan platting langsung merupakan metode yang sering digunakan untuk mendeteksi dan menghitung S. aureus di dalam makanan. Prosedur pengayaan ada yang selektif dan non selektif. Pengayaan non selektif bertujuan untuk menunjukkan adanya pertumbuhan sel yang luka (injured) dan dihambat oleh adanya komponen toksik yang terdapat pada media pengayaan selektif. Penghitungan dengan metode isolasi pada media pengayaan atau khususnya media selektif, dapat dilakukan dengan menggunakan metode Most Probable Number (MPN) atau Angka Paling Mungkin. Metode MPN dapat digunakan sebagai analisis rutin pada produk yang jumlah S. aureus-nya rendah dan pada makanan yang mengandung populasi kompetitor tinggi. Metode platting secara langsung cocok digunakan untuk analisis makanan yang mengharapkan jumlah S. aureus mencapai lebih dari 100 sel/g. Hal-hal yang perlu diperhatikan dalam melakukan metode ini adalah peralatan, media, reagen, persiapan sampel dan prosedur isolasi serta penghitungan S. aureus (US FDA, 2001).
Media diagnosis yang digunakan untuk mendeteksi dan menghitung Staphylococcus aureus harus memperhatikan beberapa sifat dari bakteri tersebut antara lain yaitu kemampuannya untuk tumbuh pada konsentrasi NaCl sebesar 7.5 atau 10%, lithium klorida sebesar 0.01-0.05% dan glisin sebesar 0.12 - 1.26% atau 40 ng/mL polimiksin; kemampuannya mereduksi potasium telurit dan membentuk koloni hitam secara aerobik maupun anaerobik; bentuk, ukuran dan pigmen koloni; aktivitas koagulase dan produksi asam pada medium padat; kemampuannya menghidrolisis kuning telur; produksi termonuklease, aseton,
-galaktosidase, fosfatase dan toksin α (hemolisin) dan dapat tumbuh pada suhu 42-430C di atas agar selektif (Bergdoll, 1990).
mengkonfirmasi koloni tunggal Staphylococcus aureus yaitu agar darah dan Brain Heart Infusion (BHI) yang diinkubasi pada suhu 370C dengan shaking (US FDA, 2001).
Studi tentang karakterisasi molekular S. aureus telah banyak dilakukan, baik untuk deteksi dan untuk identifikasi klaster S. aureus, mempelajari hubungan genetika isolat-isolat yang berasal dari kasus klinis ataupun dari pangan, menjelaskan keragaman filogenetiknya maupun untuk membuat primer PCR yang tepat untuk mendeteksi keberadaan bakteri ini.
Shehata (2008) melakukan isolasi S. aureus yang bersumber dari kasus klinis seperti urin dan pangan seperti daging masak, susu sapi serta susu kacang kedelai. Isolat kemudian diidentifikasi secara molekular dengan metode Random Amplified Polymorphic DNA (RAPD)-PCR menggunakan jumlah DNA primer sebanyak 11 buah, yang terdiri dari 105 buah pita, dimana 59 pita penandanya bersifat polimorfik dengan ukuran 200-2500 pasang basa. Hasilnya adalah bahwa isolat yang diuji memiliki 70% koefisien persamaan. Metode ini umumnya digunakan untuk jumlah sampel yang relatif besar, dan menggunakan DNA primer yang spesifik serta bervariasi dengan sekuen oligonukleotida acak.
Staphylococcus aureus memiliki beberapa faktor virulensi yang menyebabkan terjadinya penyakit. Hal ini dapat dideteksi dan diidentifikasi secara molekular juga, yaitu dengan cara mengamplifikasi gen yang diinginkan melalui teknik Polimerase Chain Reaction (PCR). Rall et al., (2008) mendeteksi gen penghasil enterotoksin stafilokoki yang berasal dari susu sapi mentah dan susu sapi pasteurisasi. Berdasarkan penelitian tersebut, 57 galur S. aureus hasil isolasi, sebesar 39 isolat (68.4%) positif mengandung satu atau lebih gen SE. Enterotoksin stafilakoki yang paling ditemukan yaitu SEA (41%) dan SEA (20.5%).
stabil terhadap panas dan gen hsp60 (heat shock protein 60 kDa). Hasilnya menunjukkan bahwa kedua gen tersebut lebih dapat membedakn secara efektif genus dan spesies bakteri Staphylococcus (Stackebrandt dan Goebel, 1994).
Enterotoksin Stafilokoki (SE)
Enterotoksin stafilokoki (SE) adalah kelompok protein dengan rantai tunggal dan bersifat antigenik dengan berat molekul sekitar 26,000-29,000 yang diproduksi oleh beberapa jenis stafilokoki terutama oleh S. aureus, tetapi bisa juga oleh S. intermedius, S. hyicus, S. xylosus dan S. epidermis. Enterotoksin termasuk protein netral dengan nilai isoelektrik sebesar 7 – 8.6. Zat ini resisten terhadap enzim-enzim proteolitik, seperti tripsin dan pepsin, hal inilah yang memungkinkan bagi enterotoksin untuk dapat masuk ke dalam saluran pencernaan menuju sisi aksinya. Toksin ini sangat stabil dalam pemanasan, dan membuat enterotoksin menjadi titik potensial bahaya kesehatan dalam pembuatan makanan kaleng (Monday dan Bennet, 2003).
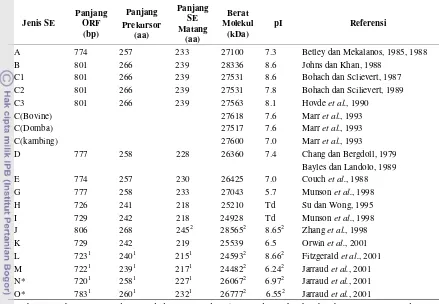
Tabel 6 Karakteristik utama SE
*Sebelumnya bernama SEK dan SEL di dalam Jarraud et al., 2001, dinamakan kembali berturut-turut menjadi SEM dan SEO, di dalam catatan koreksi dipublikasi di dalam J. immunol 166 : 4260 (2001)
1
Panjang SE matang ditentukan oleh penulis setelah Henrik Nielson, Jacob Engelbrecht, Seren Branak dan Gunnar von Heijne : Identification of prokayotic and eukaryotic signal peptides and prediction of their cleavage sites. Protein Eng 10 :1-6 (1997)
2
Berat molekul dan titik isoelektrik SE matang ditentukan oleh penulis menggunakan software MWCALC, Infobiogen, http://www.infobiogen.fr/service/analyseq/cgi-bin/mwcalc_in.pI
Td, tidak ditentukan Le Loir et al. (2003)
dan ini terjadi pada empat bentangan sekuen utama yang berlokasi baik di tengah maupun di C terminal.
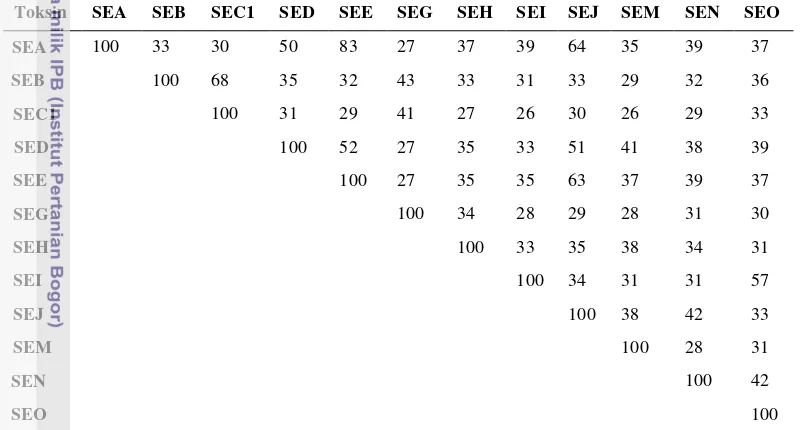
Tabel 7 menampilkan persentase identitas asam amino dari beberapa jenis enterotoksin stafilokoki yang menunjukkan seberapa besar kemiripan antar mereka.
Tabel 7 Persentase identitas asam amino pada SE yang berbeda
Toksin SEA SEB SEC1 SED SEE SEG SEH SEI SEJ SEM SEN SEO
SEA 100 33 30 50 83 27 37 39 64 35 39 37
EB SEB 100 68 35 32 43 33 31 33 29 32 36
EC1 SEC1 100 31 29 41 27 26 30 26 29 33
SED 100 52 27 35 33 51 41 38 39
EE SEE 100 27 35 35 63 37 39 37
SEG 100 34 28 29 28 31 30
E SEH 100 33 35 38 34 31
EI SEI 100 34 31 31 57
EJ SEJ 100 38 42 33
EM SEM 100 28 31
E SEN 100 42
EO SEO 100
Setelah Jarraud et al., 2001. Penamaan dikoreksi oleh penulis setelah catatan koreksi dipublikasi di dalam J. Immunol. 166:4260. Amino acid sequences of the precursors were compared using Blast2 sequence (open gap of 11 and extension gap penalties of 1)
Le Loir et al. (2003)
ketergantungan pada gen agr, sedangkan sea dan sej tidak bergantung pada gen agr. Penelitian yang dilakukan oleh Zhang et al. (1998) menunjukan bahwa SEB
yang menyerupai toxic shock syndrome toxins (TSST) tidak mempunyai regulator untuk transkripsi gen eksoprotein dan karenanya SEB bergantung pada sistem agr. Ekspresi gen agr sangat berkaitan dengan quorum sensing, produksi SE yang
bergantung pada agr di dalam komponen makanan bergantung pada kemampuan S. aureus untuk meningkatkan densitas sel yang tinggi (diperkirakan 106 CFU/g) di dalam makanan, dan faktor lingkungan memainkan peran penting di dalam ekspresi gen SE (Betley dan Mekalanos, 1985). Tabel 8 menampilkan support genetik pada beberapa gen enterotoksin stafilokoki.
Tabel 8 Support genetika pada beberapa gen penyandi SE
Shafer dan Landolo, 1978; Shalita et al., 1977; Altboum et al., 1985
sec1 Plasmid Altboum et al., 1985 secbov Phatogenicity island Fitzgerald et al., 2001 sed plasmid (pIB485) Bayles dan Iandolo, 1989 see fage cacat Couch et al., 1988
seg Enterotoxin gene cluster (egc), kromosom
Jarraud et al., 2001
sei egc, kromosom Jarraud et al., 2001 sej plasmid (pIB485) Zhang et al., 1998 sek Phatogenicity island Orwin et al., 2001 sel Phatogenicity island Fitzgerald et al., 2001 sem egc, kromosom Jarraud et al., 2001
Enterotoksin stafilokoki merupakan salah satu faktor virulensi yang dimiliki oleh S. aureus. Faktor virulensi inilah yang menyebabkan bakteri tersebut bersifat patogen, terutama jika sudah mengontaminasi produk pangan. Dampaknya tidak lain adalah keracunan pangan.
Banyak variabel yang mempengaruhi jumlah produksi enterotoksin, oleh karena itu tidak ada hal dapat memprediksi dengan tepat jumlah dari S. aureus dalam masakan yang dibutuhkan untuk menyebabkan SFP (Staphylococcal food poisoning) ini. Faktor yang berkontribusi terhadap tingkat konsentrasi
enterotoksin telah secara luas dipelajari, antara lain komposisi masakan, temperatur, dan keberadaan dari inhibitor (Doyle et al., 1997).
produksinya. Suhu optimum untuk produksi enterotoksin sama dengan suhu optimum pertumbuhan dari bakteri S. aureus tersebut, sedangkan batas suhu untuk produksi enterotoksin sekitar 10ºC. Pada suhu ini produksi enterotoksin sangat lambat dan dalam satu penelitian dibutuhkan empat minggu untuk memproduksi enterotoksin (SEB) dari inokulasi sebanyak 106/g pada daging babi. Enterotoksin stafilokoki sangat resisten terhadap pembekuan dan pemanasan. Enterotoksin dapat bertahan pada semua jenis proses pasteurisasi komersial dan kadangkala dapat bertahan pula dalam proses sterilisasi pada makanan kaleng (Lund, 2000).
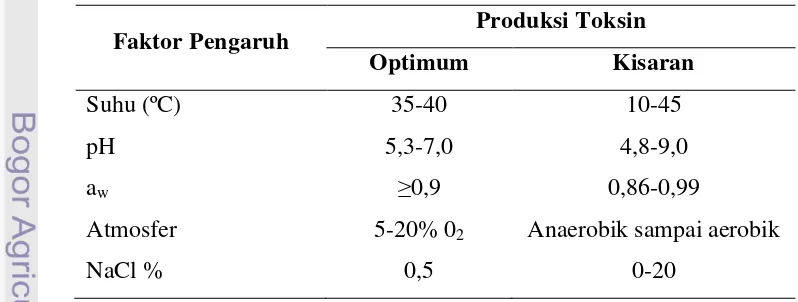
Enterotoksin Staphylococcus diproduksi pada kisaran pH yang lebih rendah dari pada nilai kisaran pH pertumbuhan bakterinya, dan batas pH untuk dapat memproduksi enterotoksin pada makanan asam adalah 5. Nilai pH optimum untuk produksi enterotoksin berada pada kisaran 7 - 8. Sementara itu, secara umum nilai aw optimum untuk produksi enterotoksin lebih tinggi dari pada batas aw pertumbuhan bakteri S. aureus tersebut (Lund, 2000). Sumber lain yang menyajikan data mengenai faktor-faktor atau kondisi yang dapat mempengaruhi produksi enterotoksin dapat dilihat pada Tabel 9.
Tabel 9 Faktor-faktor yang mampengaruhi produksi enterotoksin yang dihasilkan oleh S. aureus
Faktor Pengaruh Produksi Toksin
Optimum Kisaran
Suhu (ºC) 35-40 10-45
pH 5,3-7,0 4,8-9,0
aw ≥0,9 0,86-0,99
Atmosfer 5-20% 02 Anaerobik sampai aerobik
NaCl % 0,5 0-20
Adam dan Moss (1995)
diformulasikan dengan cara yang berbeda, dan disimpan dalam kondisi tertentu. (Lund, 2000).
Sistem regulator utama yang mengatur ekspresi gen faktor virulensi S. aureus adalah berupa aksesori gen regulator (agr) yang beraksi dengan
bergabung dengan aksesori regulator staphylococcal (sar). Staphylococcus aureus dalam memproduksi enterotoksin stafilokoki membutuhkan beberapa kondisi lingkungan yang mendukung seperti asam amino pada kebutuhan nutrisinya, konsentrasi garam dan tingkat keasaman. Staphylococcus aureus membutuhkan asam amino valin untuk pertumbuhannya dan arginin serta sistein untuk pertumbuhan sekaligus produksi enterotoksin stafilokoki. Untuk beberapa jenis enterotoksin stafilokoki seperti SEB dan SEC, keberadaan glukosa justru dapat menghambat produksi enterotoksi stafilokoki. Kondisi ini merupakan dampak dari proses metabolisme glukosa yang dapat mempengaruhi tingkat keasaman di lingkungan sekitarnya atau menurunkan pH . Hal ini juga sangat berpengaruh terhadap ekspresi gen agr, sebagai gen regulator utama yang mengatur ekspresi gen faktor virulensi (Le Loir et al., 2003).
Produksi enterotoksin stafilokoki akan optimal jika kondisi pH lingkungannya netral dan menurun pada kondisi pH yang terlalu asam (di bawah 5). Beberapa penelitian melaporkan bahwa pada pH 8,0 produksi SE sangat terhambat. Sedangkan pada pH 6,0 produksi SE akan berkurang sebanyak 50% dibandingkan dengan pH 7,0 (Supardi dan Sukamto, 1999). Pada umumnya produksi SEA kurang sensitif terhadap pH daripada SEB. Kultur medium yang dipertahankan pH-nya pada pH 7 menghasilkan lebih banyak SEB daripada kultur medium yang tidak dipertahankan pH-nya atau yang dipertahankan pH-nya pada kondisi asam. Menurut ICMSF (1996), SEA dapat dihasilkan pada aw yang lebih rendah daripada SEB. SEA dan SED umumnya dihasilkan pada kisaran kondisi pertumbuhan yang lebih luas daripada SEB. Produksi enterotoksin dapat dihambat dengan etil-4-hidroksibenzoat.
Penambahan sodium klorida yang cukup tinggi meningkatkan efek penghambatan tingkat keasaman. Enterotoksin stafilokoki tidak diproduksi pada konsentrasi garam di atas 12%. Pada SEB, SEC dan SED, pH yang tinggi atau bersifat basa juga mampu menurunkan produksi enterotoksin stafilokoki jenis tersebut sehingga ekspresi gen agr juga terhambat (Le Loir et al., 2003).
Staphylococcus aureus merupakan salah satu jenis bakteri yang tidak mampu untuk berkompetisi dengan bakteri lainnya. Kompetisi bakteri yang terjadi di dalam produk susu menunjukkan bahwa pertumbuhan S. aureus yang bertahan cukup sedikit dan enterotoksi stafilokoki yang diproduksi pun juga sedikit jumlahnya. Bakteri asam laktat (BAL) yang umumnya terdapat di dalam susu pertumbuhannya lebih bersifat dominan karena bakteri ini memproduksi beberapa senyawa yang mampu menghambat pertumbuhan S. aureus seperti asam laktat yang mampu menurunkan pH, oksigen peroksida dan substansi antimikrobial lainnya (bakteriosin) (Le Loir et al., 2003). Beberapa penelitian melaporkan bahwa spesies tertentu dari bakteri asam laktat dapat menghambat pertumbuhan S. aureus dan produksi enterotoksinnya, terutama golongan streptokoki dan
pediokoki misalnya Pediococcus cerevisiae. Sedangkan Lactobasili dan Leuconostoc tidak berpengaruh terhadap pertumbuhan sel dan hanya sedikit mempengaruhi produksi toksin. Bakteri-bakteri lain misalnya Serratia marcescens, E. coli, dan Streptococcus faecalis tidak berpengaruh terhadap
pertumbuhan sel S. aureus, tetapi menghambat produksi enterotoksin (Ikeda, et al., 2005).
Mekanisme Aksi SE
Pada mekanisme aksinya, enterotoksin stafilokoki yang dihasilkan oleh bakteri S. aureus memiliki sifat yang berbeda dalam memicu terjadinya kasus keracunan pangan. Enterotoksin tersebut ada yang bersifat memicu terjadinya muntah atau emetic activity dan ada yang bersifat superantigen. Namun mekanisme superantigen activity sudah jauh lebih baik karakterisasinya dibandingkan dengan emetic activity.
enterotoksin stafilokoki tipe I (SEI), yang meskipun memiliki struktur cystine loop tetapi SEI bersifat superantigen dan emetic (Le Loir et al., 2003; Pinchuk et
al., 2009).
Antigen sebagai sesuatu yang asing bagi tubuh, akan direspon oleh sistem imun yang ada, dalam hal ini adalah sel T. Sebagai senyawa asing, antigen akan berinteraksi dengan reseptor antigen sel T atau T-Cell Antigen Receptors (TCR) dan Major Histocompability (MHC) pada permukaan Antigen-Presenting Cells (APC) (Le Loir et al., 2003 dan Pinchuk et al., 2009).
TCR akan mengalami proses glikosilasi heterodimer membentuk rantai α dan serta rantai δ dan . Hal ini merupakan bagian dari tahap awal respon imun
selular dan sebagai hal utama yang berperan dalam meningkatkan spesifisitas respon imun tubuh. Namun untuk kasus keracunan pangan yang disebabkan oleh enterototoksin stafilokoki, SE sebagai superantigen akan berinteraksi dengan
Gambar 2 Mekanisme aksi SE (Pinchuk et al., 2009)
Konstruksi Pustaka Gen 16S rRNA
Polymerase Chain Reaction (PCR)
PCR atau polimerisasi berantai adalah teknik amplifikasi (perbanyakan) DNA spesifik dengan melakukan proses pemanjangan nukleotida dari primer yang merupakan pasangan komplementer dari utas DNA secara simultan. Proses pemanjangan nukleotida dilakukan oleh DNA polimerase berdasarkan cetakan DNA (Muladno, 2002; Yusuf, 2001).
Menurut Muladno (2002), tahap-tahap PCR meliputi tahap denaturasi, penempelan primer pada cetakan DNA (annealing) dan tahap pemanjangan primer melalui reaksi polimerisasi nukelotida (extention).
1. Denaturasi
Tahap ini merupakan tahap pengudaran DNA utas ganda menjadi DNA utas tunggal, dimana masing-masing untai dapat mencetak pasangannya (komplementer). Denaturasi berlangsung pada suhu 90-95oC.
2. Penempelan primer pada cetakan DNA (annealing)
Tahap ini merupakan tahap penempelan primer pada utas DNA cetakan yang telah terdenaturasi menjadi utas tunggal akibat kecocokan pasangan basa. Umumnya penempelan terjadi pada suhu 55-57oC untuk primer 20 mer dan 34-40oC untuk primer 10 mer. Suhu penempelan primer yang ideal umumnya adalah 5oC di bawah suhu leleh (Tm) dari tiap primer (Sambrook et a., 1989) 3. Pemanjangan primer DNA
Setelah primer menempel pada utas tunggal DNA cetakan, maka DNA polimerase akan mensintesis utas DNA yang baru berdasarkan utas DNA cetakan. DNA polimerase mulai mensintesis DNA dengan mengikatkan
deoksinukleotida pada ujung γ’-OH dari primer, sehingga arah pertumbuhan
utas DNA yang baru adalah 5’-P ke γ’-OH. Síntesis DNA atau pemanjangan primer ini dilakukan pada suhu cukup tinggi, yaitu sekitar 72oC supaya tahap berikutnya (denaturasi protein) relatif lebih mudah dan enzim Taq DNA polimerase dapat bekerja optimal.
suhu dan selang waktu yang dikehendaki untuk setiap siklus pada suatu reaksi. Banyaknya pengulangan sangat tergantung dari kemampuan DNA polimerase untuk mensintesis DNA dan biasanya berkisar antara 25 dan 40 siklus. Reaksi polimerisasi ini berantai atau berulang, maka dibutuhkan primer dalam jumlah realtif banyak. Efisiensi reaksi dapat dilakukan dengan perlakuan pra-PCR pada suhu 95oC selama 5 menit untuk mendenaturasi DNA cetakan yang ukurannya relatif besar. Setelah reaksi selesai, biasanya ditambahkan perlakuan pasca-PCR pada suhu 72oC selama 5 menit. Hasil amplifikasi dapat dilihat dengan melakukan migrasi di dalam gel (elektroforesis).
Menurut Sambrook et al., (1989) Kegagalan reaksi PCR selain karena tidak sempurnanya denaturasi atau suhu annealing yang terlalu tinggi, juga disebabkan oleh beberapa faktor lain diantaranya :
1. Konsentrasi DNA cetakan
Proses PCR tidak memerlukan DNA dengan tingkat kemurnian tinggi, namun amplifikasi akan terganggu apabila DNA cetakan masih banyak yang terkontaminasi dengan deterjen, EDTA maupun fenol. Konsentrasi DNA yang dibutuhkan adalah 10-100 ng untuk setiap reaksi.
2. Pemicu reaksi
Primer adalah rantai utas tunggal DNA yang pendek dan terdiri dari beberapa nukleotida. Umumnya terdiri atas 10-25 nukleotida (oligonukleotida). Primer yang biasa digunakan dalam percobaan adalah primer acak dan primer spesifik. Primer acak adalah primer yang susunan basa nukleotida seimbang sehingga dapat digunakan untuk analisis DNA dengan sampel yang belum diketahui susunan basa nukleotidanya. Primer spesifik adalah primer yang susunan basa nukleotidanya telah diketahui dan merupakan komponen dari utas DNA yang akan dianalisis.
3. Enzim Taq DNA Polimerase
pada suhu 75-80oC dan digunakan untuk membantu amplifikasi potongan primer dan proses pemanjangan DNA. Aktivitas enzim ini akan terhambat oleh adanya bufer fosfat, tetapi akan aktif apabila ditambahkan 10 mM tris dalam bufer pada suhu ruang dengan pH 8.3 (Sambrook et al. 1989). Taq DNA polimerase mulai aktif pada pH 8.2 - 9.0 dan suhu 65 - 72oC.
4. dNTP
dNTP yang digunakan berupa campuran dari keempat macam nukleotida yaitu dATP, dGTP, dTTP dan dCTP. Larutan stok dNTP bersifat netral pada pH sekitar 7.0. Konsentrasi dNTP yang digunakan berkisar antara 0.1 - 1.6 mM untuk setiap reaksi. dNTP masih bersifat stabil sampai proses siklus berulang 50 kali hanya berkurang 50% (Newton, 1995).
5. Mg2+
Mg2+ mempengaruhi aktivitas enzim Taq DNA polimerase karena ion Mg2+ berfungsi sebagai kofaktor yang dapat membentuk kelat dengan larutan EDTA. Ion ini berperan dalam kestabilan primer pada tahap penempelan primer. 6. Bufer
Bufer PCR terdiri atas larutan Tris-HCl dengan konsentrasi 10-50 mM dan pH 8.3 - 8,8 serta berperan dalam keberhasilan proses amplifikasi (Innis dan Gelfand, 1990). Proses penempelan primer pada bufer PCR dapat ditambahkan KCl dengan konsentrasi 50 mM.
Perunutan Basa Nukleotida (Sekuensing)
Sekuensing DNA adalah suatu proses penentuan urutan basa suatu DNA. Proses ini menggunakan prinsip reaksi polimerisasi DNA secara enzimatis. Reaksi yang dilakukan secara in vitro ini dikembangkan oleh Sanger dengan cara memasukkan satu nukleotida ddNTP (dideoksi nukleosidatrifosfat) yang berbeda ke dalam masing-masing 4 reaksi untuk menghentikan reaksi polimerisasi. Teknik dideoksi sekuensing hanya mampu membaca dengan teliti urutan basa sepanjang 400 sampai dengan 500 bp (Brown, 1992).
bioteknologi secara umum. Berikut adalah tahap-tahap yang dilakukan dalam sekuensing :
(i) Disiapkan empat macam reaksi polimerisasi DNA yang masing-masing mengandung primer DNA, dATP, dCTP, dGTP, dTTP, enzim DNA polimerase.
(ii) Selanjutnya ke dalam masing-masing tabung ditambahkan satu ddNTP yang berbeda dan reaksi dijalankan.
(iii)Hasil reaksi difraksionasi (dielektroforesis dengan gel poliakrilamida).
(iv) Fragmen DNA hasil pemisahan kemudian divisualiasaikan secara otomatis atau manual (Brown, 1992).
Analisis Keragaman Genetika
Keragaman mikroorganisme mudah dimengerti melalui sistem klasifikasi dan identifikasi berdasarkan filogenetik. Pendekatan yang umum digunakan dewasa ini adalah melalui analisis sekuen gen 16S rRNA. Gen 16S rRNA merupakan gen bagian dari DNA yang bersifat spesifik terhadap organisme prokariot, sehingga dapat dikatakan bahwa gen 16S rRNA juga dibentuk oleh gen 16S rDNA (Amann et al. 1994).
Sekuen gen 16S rRNA memiliki ukuran sekitar 1550 bp dan terdiri dari dua daerah, yaitu variable region dan conserved region. Conserved region umumnya menggunakan primer yang bersifat universal dengan ukuran sekitar 540 bp. Variable region merupakan daerah sekuen yang digunakan untuk membandingkan tingkat genus, spesies dan subspesies dalam taksonomi.