PEWARISAN KARAKTER KOMPONEN HASIL DAN
PEMANFAATAN SEGREGAN TRANSGRESIF
PERSILANGAN CABAI BESAR DAN KERITING DALAM
RANGKA PERBAIKAN HASIL
YUNANDRA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Pewarisan Karakter Komponen Hasil dan Pemanfaatan Segregan Transgresif Persilangan Cabai Besar dan Keriting dalam Rangka Perbaikan Hasil adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2016
Yunandra
RINGKASAN
YUNANDRA. Pewarisan Karakter Komponen Hasil dan Pemanfaatan Segregan Transgresif Persilangan Cabai Besar dan Keriting dalam Rangka Perbaikan Hasil. Dibimbing oleh MUHAMAD SYUKUR dan AWANG MAHARIJAYA.
Perakitan varietas cabai berdaya hasil tinggi merupakan salah satu cara untuk meningkatkan daya hasil cabai. Kegiatan seleksi memiliki peran yang sangat penting dalam rangka perakitan varietas cabai berdaya hasil tinggi. Seleksi menjadi efektif dan efisien ketika genotipe segregan transgresif berhasil diperoleh pada generasi awal sehingga dapat mempercepat perolehan galur-galur harapan berdaya hasil tinggi. Penelitian ini bertujuan untuk memperoleh informasi tentang pewarisan karakter komponen hasil pada tanaman cabai dan memperoleh kandidat galur harapan segregan transgresif homozigot pada awal generasi bersegregasi. Penelitian ini terdiri dari 3 percobaan, yaitu studi pewarisan karakter hasil dan komponen hasil cabai, seleksi genotipe cabai segregan transgresif, dan verifikasi genotipe cabai segregan transgresif pada populasi terseleksi.
Hasil penelitian menunjukkan bahwa karakter hasil dan komponen hasil cabai dikendalikan oleh gen-gen yang berada di dalam inti. Pewarisan karakter hasil dan komponen hasil cabai dikendalikan oleh aksi gen aditif dan non-aditif, dimana aksi gen non-aditif ini terdiri dari aksi gen dominan dan epistasis. Terdapat genotipe diduga segregan transgresif pada populasi F2 terseleksi untuk karakter umur berbunga, umur panen, panjang tangkai buah, panjang buah, jumlah buah, dan bobot buah per tanaman. Segregan transgresif terverifikasi pada karakter hasil dan komponen hasil populasi F3. Genotipe terverifikasi segregan transgresif untuk karakter bobot buah per tanaman sebanyak 10 genotipe F3 dari 20 genotipe F2 terpilih yang diduga segregan transgresif. Genotipe F3120005-3, F3120005-16, F3120005-74, F3120005-87, F3120005-120, F3120005-136, F3120005-146, F3120005-176, F3120005-184, dan F3120005-199 merupakan genotipe yang direkomendasikan sebagai kandidat galur harapan.
SUMMARY
YUNANDRA. Inheritance of Yield Component Characters and Transgressive Segregant Utilization in Big Chili and Curling Pepper Crosses for Yield Improvement. Supervised by MUHAMAD SYUKUR and AWANG MAHARIJAYA.
Varieties assembly of high yield pepper is one of the methods for increasing pepper productivity. Plant selections are important part in varieties assembly. Effective and efficient selections could be achieved when transgressive segregant genotypes successfully obtained on early generation thus shorten time to obtain promising high yield lines. This study aims to obtain genetic information about yield component characters inheritance in pepper and obtain new transgressive segregant homozygous candidate lines in early generation. This study consists of three experiments: observation of pepper yield component characters inheritance, pepper transgressive segregant genotypes selection, and verification pepper transgressive segregant genotypes in selected population.
Pepper yield component characters were controlled by additive and non-additive gene action, where non-non-additive gene action consists of dominant gene and epistatic. There were expected transgressive segregant genotypes in selected F2 population for flowering age, harvesting age, pedicel length, fruit length, number of fruit, and yield characters. Transgressive segregant verified in all yield component characters of F3 population. Transgressive segregants were verified for yield component characters in F3 population. Ten F3 genotypes from twenty selected F2 genotypes were suspected transgressive segregants for yield character. Genotypes of 3, 16, 74, 87, F3120005-120, F3120005-136, F3120005-146, F3120005-176, F3120005-184, and F3120005-199 were recommended as new candidate lines.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
PEWARISAN KARAKTER KOMPONEN HASIL DAN
PEMANFAATAN SEGREGAN TRANSGRESIF
PERSILANGAN CABAI BESAR DAN KERITING DALAM
RANGKA PERBAIKAN HASIL
YUNANDRA
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : Pewarisan Karakter Komponen Hasil dan Pemanfaatan Segregan Transgresif Persilangan Cabai Besar dan Keriting dalam Rangka Perbaikan Hasil
Nama : Yunandra
NIM : A253130021
Disetujui oleh Komisi Pembimbing
Prof Dr Muhamad Syukur, SP MSi Ketua
Dr Awang Maharijaya, SP MSi Anggota
Diketahui oleh
Ketua Program Studi
Pemuliaan dan Bioteknologi Tanaman
Dr Ir Yudiwanti Wahyu EK, MS
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MSc. Agr
PRAKATA
Puji syukur penulis panjatkan kepada Tuhan Yang Maha Esa yang telah memberikan kekuatan, rahmat, dan karunia-Nya, sehingga penulis dapat
menyelesaikan karya ilmiah ini yang berjudul “Pewarisan Karakter Komponen Hasil dan Pemanfaatan Segregan Transgresif Persilangan Cabai Besar dan Cabai
Keriting dalam Rangka Perbaikan Hasil”.
Pada kesempatan kali ini penulis menyampaikan terima kasih kepada: 1. Almarhum ayahanda Asrab dan ibunda Nursiam, S.Pd.I tercinta yang tiada
hentinya selalu mendoakan dan memberikan kasih sayang, pengorbanan, dukungan, dan motivasi. Abang Hendri SE dan kak Afrida Wahyuli SE, abang Syafriwan ST, abang Yudi Ansar Amd dan kak Dyaning Tyas Nugraheni SPd atas doa dan dukungan selama ini.
2. Prof. Dr. Muhamad Syukur, SP MSi dan Dr. Awang Maharijaya, SP MSi selaku dosen pembimbing atas segala arahan, saran, masukan, kesabaran, dan motivasi yang telah diberikan selama ini.
3. Dr. Willy Bayuardi Suwarno, SP MSi dan Dr. Ir. Yudiwanti Wahyu EK, MS selaku dosen penguji yang telah memberikan arahan dan masukan sehingga tesis ini menjadi lebih baik.
4. Direktorat Jenderal Pendidikan Tinggi (DIKTI) yang telah memberikan kesempatan melanjutkan studi magister melalui program beasiswa BPP-DN 2013.
5. Teman-teman Labdik Pemuliaan Tanaman: kak Abdul Hakim SP, kak Ady Daryanto SP MSi, kak Arya Widura SP MSi, Estriana Riti SP, Faradila Putri SP MSi, M. Ridha Alfarabi Istiqlal SP MSi, kak Dr. Marlina Mustafa SP MP, mbak Nura SP MSi, Pak Rudi Hermanto SP, mbak Siti Hapshoh SP MSi, mbak Siti Marwiyah SP MSi, Tustiah SP, dan mbak Zahratul Mila SP MSi atas bantuan tenaga, kerja sama, kebersamaan, dan ikatan kekeluargaan yang erat. 6. Teman-teman seperjuangan di program studi Pemuliaan dan Bioteknologi
Tanaman Angkatan 2013 atas persahabatan dan kekeluargaannya.
7. Mas Undang SP MSi dan Pak Darwa atas bantuan dan kerjasama selama penelitian.
8. Tiara Yudilastari, SP atas bantuan, kerjasama, dukungan, motivasi, doa, semangat, dan menjadi pendengar setia keluh kesah penulis.
Tiada yang sempurna pada karya manusia, mohon maaf atas segala kekurangan dan kesalahan dalam penulisan karya ilmiah ini. Semoga karya ini bermanfaat dalam pengembangan ilmu pengetahuan, khususnya bidang pemuliaan tanaman dan pertanian pada umumnya.
Bogor, Agustus 2016
DAFTAR ISI
DAFTAR TABEL xii
DAFTAR GAMBAR xii
1 PENDAHULUAN 1
1.1 Latar Belakang 1
1.2 Tujuan Penelitian 2
1.3 Hipotesis Penelitian 2
1.4 Ruang Lingkup Penelitian 3
2 TINJAUAN PUSTAKA 4
2.1 Botani dan Morfologi Cabai 4
2.2 Studi Pewarisan Karakter Hasil 4
2.3 Seleksi dan Kemajuan Seleksi 5
2.4 Segregan Transgresif 6
3 STUDI PEWARISAN KARAKTER HASIIL DAN KOMPONEN
HASIL CABAI 7
3.1 Pendahuluan 8
3.2 Bahan dan Metode 8
3.3 Hasil dan Pembahasan 12
3.4 Simpulan 23
4 SELEKSI GENOTIPE CABAI SEGREGAN TRANSGRESIF 24
4.1 Pendahuluan 25
4.2 Bahan dan Metode 25
4.3 Hasil dan Pembahasan 26
4.4 Simpulan 30
5 VERIFIKASI SEGREGAN TRANSGRESIF PADA POPULASI
TERSELEKSI 31
5.1 Pendahuluan 32
5.2 Bahan dan Metode 32
5.3 Hasil dan Pembahasan 34
5.4 Simpulan 41
6 PEMBAHASAN UMUM 42
7 SIMPULAN UMUM DAN SARAN 45
7.1 Simpulan 45
7.2 Saran 45
DAFTAR PUSTAKA 46
DAFTAR TABEL
3.1 Klasifikasi derajat dominansi berdasarkan potensi rasio (hp) 10 3.2 Nilai tengah dan ragam karakter Nilai tengah dan ragam
karakter umur berbunga, umur panen, bobot per buah, panjang tangkai
buah, dan panjang buah cabai 12
3.3 Nilai tengah dan ragam karakter diameter buah, tebal daging buah,
jumlah buah, dan bobot buah per tanaman cabai 13 3.4 Uji efek maternal populasi F1 dan F1R pada karakter komponen
hasil cabai 17
3.5 Jumlah gen pengendali (n) dan derajat dominansi (hp) pada
karakter komponen hasil cabai 18
3.6 Uji skala pada karakter komponen hasil cabai 19 3.7 Uji skala gabungan pada karakter umur berbunga, umur panen,
bobot per buah, panjang tangkai buah, dan panjang buah cabai 20 3.8 Uji skala gabungan pada karakter diameter buah, tebal daging
buah, jumlah buah, dan bobot buah per tanaman cabai 20 3.9 Perkiraan efek gen pada karakter umur berbunga, umur panen,
bobot per buah, panjang tangkai buah, dan panjang buah cabai 21 3.10 Perkiraan efek gen pada karakter umur berbunga, umur panen,
bobot per buah, panjang tangkai buah, dan panjang buah cabai 22 3.11 Komponen ragam dan heritabilitas pada karakter komponen
hasil cabai 23
4.1 Korelasi pearson karakter komponen hasil terhadap bobot buah
per tanaman cabai 27
4.2 Nilai indeks seleksi karakter panjang buah, jumlah buah per
tanaman, bobot per buah, dan bobot buah per tanaman cabai 28 4.3 Segregan transgresif pada 300 tanaman F2 cabai 29 4.4 Segregan transgresif pada 30 tanaman F2 cabai terseleksi 29 5.1 Kemajuan seleksi pada karakter komponen hasil cabai 34 5.2 Heritabilitas dalam arti luas dan sempit (realized) karakter
komponen hasil pada populasi F3 cabai 35
5.3 Nilai tengah dan ragam populasi F3 karakter umur berbunga,
umur panen, dan bobot per buah cabai 37
5.4 Nilai tengah dan ragam populasi F3 karakter diameter buah,
jumlah buah, dan panjang buah cabai 39
5.5 Nilai tengah dan ragam populasi F3 karakter panjang tangkai
buah, tebal daging buah, dan bobot buah per tanaman cabai 40
DAFTAR GAMBAR
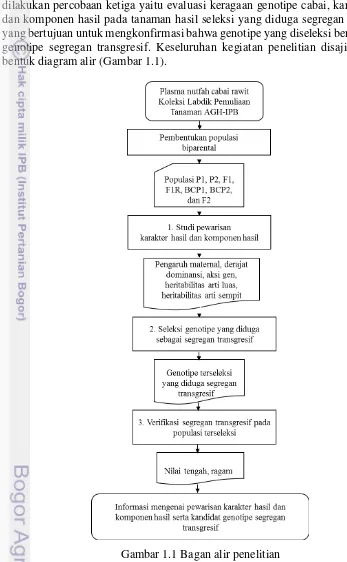
1.1 Bagan alir penelitian 3
3.1 Skema persilangan pada cabai 9
3.2 Sebaran data populasi F2 karakter umur berbunga dan umur panen cabai 14 3.3 Sebaran data populasi F2 karakter bobot per buah, panjang buah,
dan diameter buah cabai 15
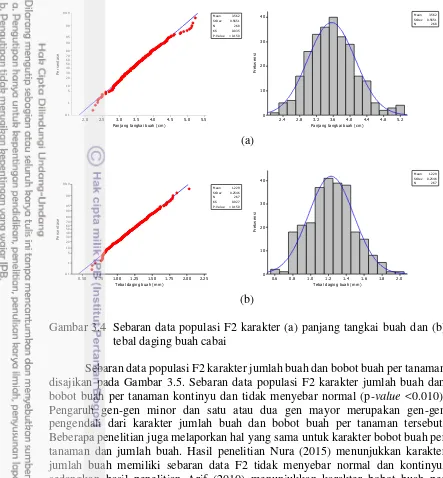
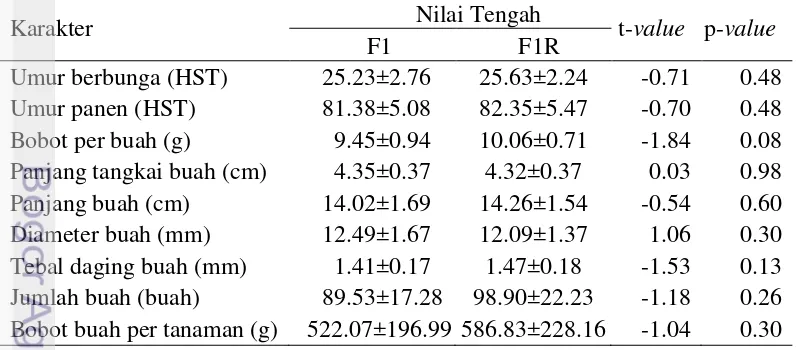
3.4 Sebaran data populasi F2 karakter panjang tangkai buah dan tebal
daging buah cabai 16
3.5 Sebaran data populasi F2 karakter jumlah buah dan bobot buah per
1
PENDAHULUAN
1.1 Latar Belakang
Cabai (Capsicum annuum L.) banyak ditanam di seluruh dunia baik sebagai rempah-rempah maupun sebagai tanaman sayur dan cabai juga termasuk salah satu sayuran solanaceae kedua yang paling penting di dunia setelah tomat (Hasanuzzaman dan Golam 2011). Produktivitas cabai di Indonesia masih tergolong rendah, yakni sebesar 8.35 ton ha-1 pada tahun 2014 (BPS 2015) sedangkan potensi hasil yang dapat dicapai adalah 18-24.2 ton ha-1 (Rubatzky dan Yamaghuci 1999). Rendahnya produktivitas cabai di Indonesia dikarenakan banyak faktor, salah satunya adalah petani belum banyak menggunakan varietas unggul berdaya hasil tinggi. Varietas unggul cabai bisa didapat dari serangkaian program pemuliaan tanaman yang tepat.
Keragaman genetik merupakan faktor yang sangat berpengaruh terhadap berhasilnya suatu program pemuliaan tanaman. Sumber keragaman genetik didapat dari introduksi, hibridisasi, mutasi maupun rekayasa genetika. Hibridisasi atau persilangan dua galur murni merupakan teknik yang paling umum digunakan dalam kegiatan pemuliaan tanaman. Tetua galur murni atau dalam keadaan homozigot akan menghasilkan turunan F1 heterozigot yang seragam dan segregasi akan muncul pada generasi F2 (Sofiari dan Kirana 2009). Kegiatan seleksi dapat dilakukan pada populasi F2 yang bersegregasi tersebut.
Informasi genetik merupakan hal yang penting dalam menyeleksi hasil persilangan untuk mendapatkan varietas unggul, terutama informasi pewarisan karakter kualitatif dan kuantitatif. Analisis pewarisan karakter kualitatif dan kuantitatif bertujuan untuk mengetahui jumlah gen yang mengendalikan karakter tersebut, aksi gen yang mengendalikan, dan informasi genetik lainnya (Arif et al. 2011). Melalui informasi pewarisan karakter yang digunakan, kegiatan seleksi akan menjadi lebih efektif dan efisien. Seleksi pada beberapa karakter komponen hasil lebih efektif daripada hanya seleksi pada satu karakter hasil per tanaman. Hal ini dikarenakan produktivitas cabai sangat dipengaruhi oleh karakter komponen hasil seperti bobot buah, diameter buah, panjang buah, panjang tangkai buah, tebal daging buah dan jumlah buah.
Langkah pertama yang dilakukan dalam kegiatan pemuliaan tanaman yaitu memilih tetua persilangan. Selanjutnya untuk memprediksi frekuensi segregan transgresif dapat dilakukan dengan persilangan dialel. Informasi daya gabung umum dari persilangan dialel dengan asumsi tidak ada linkage, tidak ada epistasis dan tidak ada efek maternal dapat diketahui aksi gen aditif (Kuczyñska et al. 2007). Aksi gen aditif berhubungan dengan lokus homozigot pada tanaman menyerbuk sendiri (Griffing 1956), sehingga frekuensi segregan transgresif dapat diprediksi. Kandidat tetua dengan daya gabung umum baik ini dipilih sebagai tetua yang digunakan sebagai tetua untuk persilangan biparental.
pada sifat-sifat kuantitatif dari zuriat hasil persilangan dua tetua yang memiliki jangkauan sebaran yang melampaui jangkauan sebaran kedua tetuanya (Poehlman dan Sleper 1996). Seleksi pada zuriat yang memiliki jangkauan sebaran melampaui jangkauan sebaran kedua tetuanya, belum tentu menghasilkan turunan yang keragaannya sama dengan tetuanya. Hal ini dikarenakan adanya pengaruh genotipe dan interaksi genotipe dengan lingkungan yang akan mengaburkan penarikan kesimpulan mengenai nilai fenotipe tanaman (Jambormias dan Riry 2009). Oleh karena itu proses seleksi memakan waktu yang lama hingga generasi F6 atau F7 untuk mendapatkan galur harapan.
Metode seleksi berbasis indeks merupakan salah satu metode seleksi yang cukup populer dilakukan pada generasi F2. Menurut Sumarno dan Zuraida (2006) metode seleksi berbasis indeks dapat menghasilkan genotipe-genotipe terbaik untuk karakter-karakter yang dilibatkan dalam penyusunan indeks. Seleksi akan memberikan respon yang baik apabila menggunakan kriteria seleksi yang tepat. Karakter yang digunakan sebagai penyusun indeks merupakan karakter-karakter yang memberikan pengaruh cukup besar pada peningkatan daya hasil. Individu-individu terpilih pada generasi F2 dengan nilai heritabilitas yang tinggi ketika digalurkan menjadi generasi F3 diharapkan mengalami kemajuan seleksi yang positif. Jambormias et al. (2011) menyatakan bahwa generasi F3 dapat menyediakan informasi kekerabatan dalam bentuk informasi antar famili dan dalam famili. Informasi antar famili dan dalam famili ini berguna dalam menentukan suatu genotipe telah seragam atau masih bersegregasi. Genotipe dikatakan segregan transgresif apabila memiliki ragam dalam famili yang rendah. Indikasi ragam dalam famili rendah dapat dilihat dari ragam genotipe F3 lebih rendah atau sama dengan ragam tetua. Tetua merupakan galur murni yang apabila ditanam tingkat keragamannya rendah sehingga dapat dijadikan indikator dalam menentukan genotipe F3 telah seragam atau masih bersegregasi.
1.2 Tujuan Penelitian
Penelitian ini bertujuan untuk:
1. Memperoleh informasi tentang pewarisan karakter hasil dan komponen hasil pada tanaman cabai.
2. Memperoleh kandidat galur harapan segregan transgresif homozigot pada awal generasi bersegregasi.
1.3 Hipotesis Penelitian
Hipotesis yang diajukan pada penelitian ini adalah:
1. Terdapat pola pewarisan aditif dan dominan pada persilangan cabai besar dan keriting
2. Terdapat minimal satu kandidat genotipe cabai homozigot bersifat segregan transgresif
1.4 Ruang Lingkup Penelitian
Penelitian ini terdiri atas tiga percobaan. Percobaan pertama, pewarisan karakter hasil dan komponen hasil yang bertujuan untuk mendapatkan informasi genetik tentang pewarisan karakter hasil dan komponen hasil pada tanaman cabai. Kemudian dilanjutkan dengan percobaan kedua, seleksi genotipe cabai yang diduga sebagai segregan transgresif yang bertujuan untuk memperoleh genotipe dengan jangkauan sebaran yang melebihi jangkauan sebaran kedua tetuanya. Selain itu juga dilakukan percobaan ketiga yaitu evaluasi keragaan genotipe cabai, karakter hasil dan komponen hasil pada tanaman hasil seleksi yang diduga segregan transgresif yang bertujuan untuk mengkonfirmasi bahwa genotipe yang diseleksi benar sebagai genotipe segregan transgresif. Keseluruhan kegiatan penelitian disajikan dalam bentuk diagram alir (Gambar 1.1).
2
TINJAUAN PUSTAKA
2.1 Botani dan Morfologi Cabai
Cabai (Capsicum annuum L.) merupakan tanaman khas yang memiliki kandungan capsaicin. Centre of origin cabai adalah daerah selatan dari Meksiko yang menyebar hingga Columbia. Capsicum annuum L. merupakan cabai yang paling banyak di tanam. Cabai termasuk tanaman semusim (annual) berbentuk perdu. Perakaran cabai merupakan akar tunggang yang terdiri atas akar utama (primer) dan akar lateral (sekunder) dengan jumlah yang banyak (Poulos, 1993), dimana dari akar lateral tersebut keluar serabut-serabut akar (akar tersier). Akar tanaman cabai bisa tumbuh mencapai kedalaman satu meter (Rubatzky dan Yamaguchi, 1999). Sebagian besar batang cabai menjadi berkayu pada pangkal batang yang umumnya tumbuh tegak, sangat bercabang, dan tinggi 0.5 – 1.5 m. diameter batang cabai dapat mencapai hingga 1 cm. Warna batang beragam dari hijau, hijau kecoklatan dan sering juga terdapat semburat ungu pada bagian buku (Poulos 1993).
Daun cabai memiliki bentuk yang bervariasi, mulai dari lancip sampai dengan bulat telur dengan ujung daun yang lancip dan tepinya yang rata. Warna daun cabai bervariasi, dari mulai hijau, hijau tua, sampai hijau keunguan. Ukuran daun bervariasi dengan helaian daun lanset dan bulat telur lebar (Rubatzky dan Yamaguchi 1997). Tanaman cabai berbunga pada bagian aksilar buku percabangan utama yang kemudian terbentuk bunga pada setiap buku berikutnya (OECD 2006). Bunga cabai termasuk bunga hermafrodit. Petal bunga Capsicum annuum terdiri atas 5-7 helai yang umumnya berwarna putih atau ungu. Bunga cabai memiliki 3 orientasi arah tumbuh yakni: ke bawah, intermediet, dan tegak ke atas (Bosland and Votava 1999).
Buah cabai dapat diklasifikasikan sebagai buah non-klimaterik (Lounds et al. 1993). Warna buah cabai bervariasi, yaitu: hijau, kuning, atau bahkan ungu ketika muda dan kemudian berubah menjadi merah, jingga, atau campuran bersamaan dengan meningkatnya umur buah. Falusi dan Morakinyo (1994) menjelaskan bahwa terdapat berbagai variasi bentuk buah pada Capsicum annuum
L. Bentuk buah cabai, mulai dari pendek, panjang, bulat, oval, sampai keriting. Buah cabai juga memiliki buah tunggal yang tumbuh pada buku, namun juga lebih dari satu (fasiculate) tumbuh dalam satu buku (Kusandriani 1996).
2.2 Studi Pewarisan Karakter Hasil
mengindikasikan karakter tersebut dikendalikan oleh adanya pengaruh gen-gen minor dan satu atau dua gen mayor (Falconer dan Mackay 1996).
Analisis rata-rata generasi merupakan salah satu analisis yang sering digunakan untuk mempelajari pewarisan suatu karakter tanaman. Analisis rata-rata generasi merupakan alat yang dapat digunakan untuk mengkaji model efek genetik lain yang diluar model aditif-dominan (Derera dan Musimwa 2015). Analisis rata-rata generasi ini dapat digunakan untuk mempelajari pola pewarisan karakter kuantitatif dan kualititatif tanaman, pewarisan ketahanan tanaman terhadap cekaman abiotik maupun biotik, ataupun pewarisan ketahanan tanaman terhadap serangan hama dan penyakit.
Analisis rata-rata generasi menggunakan populasi yang terdiri dari dua populasi tetua, satu populasi F1, dua populasi silang balik (Backcross) dan populasi F2. Pengujian pada analisis rata-rata generasi terbagi menjadi dua, yaitu: uji skala individu dan uji skala gabungan. Uji skala individu digunakan model tiga parameter yang dijelaskan oleh Mather dan Jinks (1982), yaitu: A=2BCP1-P1-F1; B=2BCP2-P2-F1; C=4F2-2F1-P1-P2 untuk menduga kesesuaian model aditif-dominan. Apabila hasil uji skala individu tidak ada sesuai dengan model aditif-dominan maka dilanjutkan dengan uji skala gabungan (m = ½P1 + ½ P2 + 4F2 - 2BCP1 - 2BCP2; [d] = ½P1 - ½P2; [h]=6BCP1 + 6BCP2 - 8F2 - F1 - 1½P1 - 1½P2; [i]= 2BCP1 + 2BCP2 - 4F2; [j]=2BCP1 - P1 – 2BCP2 + P2; [l]=P1 + P2 + 2F1 + 4F2 – 4BCP1 – 4BCP2) untuk menambahkan kontribusi dari epistasis (interaksi non-alelik). Uji ini menghasilkan dugaan untuk tiga parameter mean (m), efek aditif [d], efek dominan [h] selain itu juga menghasilkan dugaan tiga parameter epistasis interaksi aditif x aditif [i], interaksi aditif x dominan [j] dan interaksi dominan x dominan [l].
Hasil uji skala gabungan dapat menjelaskan tipe aksi gen yang mengendalikan suatu karakter. Apabila nilai [h] dan [l] signifikan serta memiliki tanda yang sama disebut sebagai tipe komplementasi, sedangkan apabila nilai [h] dan [l] signifikan serta memiliki tanda yang berbeda disebut sebagai tipe duplikasi (Mather dan Jinks, 1982). Masing-masing tipe aksi gen dapat dimanfaatkan pada program pemuliaan tanaman. Aksi gen komplementasi dapat dimanfaatkan dalam merakit suatu varietas hibrida (Hasanuzzaman dan Golam 2011). Hal ini dikarenakan efek gen non-aditif lebih besar pada karakter tersebut. Namun apabila ingin melakukan kegiatan seleksi dapat dilakukan pada generasi lanjut, dimana tingkat homozigositas sudah cukup tinggi. Sedangkan apabila aksi gen duplikasi kurang tepat dimanfaatkan untuk merakit suatu varietas hibrida, hal ini dikarenakan mengurangi manfaat yang terjadi dari heterozigositas karena pembatalan dominasi dan efek epistasis (Dhall dan Hundal 2006).
2.3 Seleksi dan Kemajuan Seleksi
Pada tanaman menyerbuk sendiri untuk populasi heterozigot homozigot dapat menggunakan seleksi massa dan untuk populasi bersegregasi dapat menggunakan seleksi pedigree, seleksi bulk, single seed descent, dan seleksi silang balik. Sedangkan pada tanaman menyerbuk silang, metode seleksi yang digunakan berupa seleksi massa, seleksi ear to raw, dan seleksi berulang (seleksi berulang fenotipe, seleksi berulang daya gabung umum, seleksi berulang untuk daya gabung khusus, dan seleksi berulang resiprokal) (Sleper dan Poehlman 2006).
Menurut Baihaki (2000) kemajuan seleksi didasarkan pada perubahan rata-rata penampilan yang dicapai suatu populasi dalam setiap siklus seleksi. Nilai harapan kemajuan seleksi berguna menduga berapa besar pertambahan nilai sifat tertentu akibat seleksi dari nilai rata-rata populasi. Kemajuan seleksi sangat tergantung pada nilai heritabilitas populasi tanaman dan intensitas seleksi yang digunakan. Semakin tinggi nilai heritabilitas dan intensitas seleksi, maka kemajuan seleksi semakin tinggi. Nilai kemajuan seleksi pada generasi awal pada umumnya lebih tinggi dibandingkan dengan nilai kemajuan seleksi pada generasi lanjut. Hal ini dikarenakan pada generasi awal tingkat keragaman genetik dan nilai heritabilitas masih tinggi. Intensitas seleksi merupakan jumlah individu yang akan diseleksi dari suatu populasi. Pada populasi dengan tingkat keragaman yang tinggi sebaiknya digunakan intensitas seleksi yang tinggi pula. Sebaliknya, apabila tingkat keragaman rendah sebaiknya intensitas seleksi yang digunakan tidak terlalu tinggi.
2.4 Segregan Transgresif
Segregan transgresif merupakan individu-individu hasil segregasi yang memiliki keragaan di luar rentang keragaan tetuanya (Sleper dan Poehlman 2006). Keragaan yang berada diluar rentang tetuanya akibat dari berkumpulnya gen-gen homozigot dominan pada suatu genotipe turunan. Sehingga apabila genotipe segregan transgresif ini dilanjutkan, segregasi pada turunannya sudah tidak terlalu tinggi. Pada tanaman menyerbuk sendiri tingkat homozigositas cukup tinggi membutuhkan waktu yang cukup lama hingga 6-7 generasi. Hal ini dikarenakan, telah terjadi fiksasi pada berbagai lokus sehingga populasi telah menjadi homogen homozigot. Modifikasi dari seleksi dapat dilakukan untuk memotong waktu yang lama tersebut. Salah satu teknik dapat digunakan adalah dengan memilih tanaman-tanaman F2 yang berada pada posisi kanan atau kiri kurva normal. Segregan transgresif membentuk dua gugus segregan dalam spektrum sebaran, yaitu lebih kecil dari sebaran tetua dengan keragaan rendah dan lebih besar dari sebaran tetua dengan keragaan tinggi (Jambormias dan Riry 2009).
3
STUDI PEWARISAN KARAKTER HASIL DAN
KOMPONEN HASIL CABAI
Abstrak
Pewarisan karakter komponen hasil cabai dipelajari dari populasi yang berasal dari persilangan cabai keriting (IPB C120) dan cabai besar (IPB C5), antara lain F2, F1, silang balik ke tetua termasuk kedua tetua. Uji skala individu (satu atau lebih skala A, B, dan C) dan uji skala gabungan yang signifikan mengindikasikan adanya aksi gen epistasis. Model aditif-dominan dengan pengaruh interaksi aditif-aditif dan interaksi aditif-dominan sesuai untuk karakter umur panen, bobot per buah, dan diameter buah. Model aditif-dominan dengan pengaruh interaksi aditif-aditif dan interaksi dominan-dominan sesuai untuk karakter tebal daging buah, jumlah buah, dan bobot buah per tanaman. Model aditif-dominan dengan pengaruh interaksi aditif-dominan dan interaksi dominan-dominan sesuai dengan karakter umur berbunga, panjang tangkai buah, dan panjang buah. Heritabilitas dalam arti luas termasuk kategori tinggi untuk karakter umur berbunga, umur panen, bobot per buah, panjang buah, diameter buah, jumlah buah, bobot buah per tanaman serta kategori sedang untuk karakter panjang tangkai buah, dan tebal daging buah. Heritabilitas dalam arti sempit termasuk dalam kategori tinggi hanya pada karakter panjang buah. Kategori sedang untuk karakter umur panen, bobot buah, panjang tangkai buah, diameter buah, tebal daging buah, serta kategori rendah untuk karakter umur berbunga, jumlah buah, dan bobot buah per tanaman.
Kata kunci: epistasis, heritabilitas, komponen hasil, pewarisan
Abstract
Pepper yield component characters inheritance was studied in population of curling (IPB C120) and big chili pepper (IPB C5) crosses which consists of F2, F1, and first back crosses generation including both parents. The significant scaling test (one or more scales in A, B and C) and joint scaling test indicated the presence of epistasis gene action. Influence of additive-additive and additive-dominant interaction model was found suitable in harvesting age, fruit weight, and fruit diameter characters. Influence of additive-additive and dominant-dominant interanction model was found suitable for pericarp thickness, number of fruit per plant and yield. Influence additive-dominant and dominant-dominant model was suitable for flowering age, pedicel length and fruit length characters. High broad-sense heritability were found in flowering age, harvesting age, fruit weight, fruit length, fruit diameter, number of fruit per plant, and yield characters while medium broad-sense heritability were found in pedicel length and pericarp thickness characters. High narrow-sense heritability only found in for fruit length character while medium narrow-sense heritability were found in harvesting age, fruit weight, pedicel length, fruit diameter, and pericarp thickness characters. Low narrow-sense heritability were found in flowering age, number of fruit per plant and yield charater.
3.1 Pendahuluan
Cabai (Capsicum annuum L.) termasuk sayuran solanaceae kedua yang penting di dunia setelah tomat (Hasanuzzaman dan Golam 2011). Kebutuhan akan cabai semakin meningkat setiap tahunnya. Peningkatkan produktivitas cabai dapat dilakukan dengan cara menggunakan varietas unggul berdaya hasil tinggi. Varietas unggul cabai bisa didapat dari serangkaian program pemuliaan tanaman yang tepat.
Keragaman genetik merupakan faktor yang berpengaruh terhadap berhasilnya suatu program pemuliaan tanaman. Informasi genetik merupakan hal yang penting dalam menyeleksi hasil persilangan terutama informasi pewarisan karakter kualitatif dan kuantitatif. Analisis pewarisan karakter kualitatif dan kuantitatif bertujuan untuk mengetahui jumlah gen yang mengendalikan karakter tersebut, aksi gen yang mengendalikan, dan informasi genetik lainnya (Arif et al. 2011).
Selain itu, pola pewarisan, variabilitas genetik dan heritabilitas suatu karakter merupakan parameter genetik penting yang berkaitan dengan proses seleksi dan penggabungan karakter-karakter penting dalam suatu genotipe (Alia et al. 2004). Analisis rata-rata generasi merupakan alat yang dapat digunakan untuk mengkaji model efek genetik lain yang diluar model aditif-dominan (Derera dan Musimwa 2015). Penjelasan mengenai aksi gen aditif dan dominan dapat diperoleh dari penilaian perbandingan komponen linier yang terdiri dari aditif [d], dominan [h], aditif x aditif [i], aditif x dominan [j], dan dominan x dominan [l]. Studi pewarisan karakter hasil dan komponen hasil menjadi penting dalam memaksimalkan penggunaan potensi genetik dalam program pemuliaan yang efektif. Penelitian ini bertujuan untuk memperoleh informasi tentang pewarisan karakter hasil dan komponen hasil pada tanaman cabai.
3.2 Bahan dan Metode
Penelitian ini dilaksanakan pada bulan September 2013-Juli 2014 (pembentukan populasi) dan Agustus-Desember 2014 (pengujian). Pembentukan populasi dilakukan di perumahan IPB Alam Sinar Sari, sedangkan untuk pengujian dilakukan di Kebun Percobaan Leuwikopo, IPB.
Bahan tanaman yang digunakan adalah tetua cabai keriting (IPB C120) dan tetua cabai besar (IPB C5), turunan pertama (F1), turunan pertama resiprokal (F1R) masing-masing ditanam sebanyak 40 tanaman, backcross ke tetua betina (BCP1) sebanyak 100 tanaman, backcross ke tetua jantan (BCP2) sebanyak 100 tanaman, dan turunan kedua (F2) sebanyak 300 tanaman. Pembentukan populasi dilakukan dengan melakukan persilangan buatan dan selfing. Populasi yang dibentuk adalah F1, F1R, BCP1, dan BCP2. Populasi P1, P2, F2 diperoleh dari selfing, sedangkan populasi F1, F1R, BCP1, dan BCP2 diperoleh dari persilangan buatan.
Pembentukan populasi dilakukan dengan melakukan persilangan buatan dan
selfing mengikuti skema persilangan pada Gambar 3.1. Populasi yang dibentuk
Gambar 3.1 Skema persilangan pada cabai
Persilangan pembentukan populasi akan dilaksanakan secara buatan di Perumahan IPB Alam Sinar Sari. Benih cabai disemai pada tray semai dengan media tanam campuran media tanam dan pupuk kandang (1:1). Setelah bibit berumur 40 hari (atau memiliki 4-5 helai daun) dipindahkan ke polibag (ukuran 40 cm x 35 cm) berisi 10 kg campuran topsoil dan pupuk kandang (2:1). Pemupukan dengan NPK 16-16-16 dengan cara di kocor 5 g L-1 air sebanyak 250 ml per tanaman dan pupuk daun 5 g L-1 air diberikan seminggu sekali. Pengendalian hama dan penyakit menggunakan insektisida, fungisida, akarisida yang diaplikasikan jika diperlukan.
Emaskulasi dan penyerbukan dilakukan pagi hari pada pukul 06.00-09.00 WIB. Emaskulasi menggunakan pinset yang telah disterilkan dengan alkohol 70%, pada saat bunga belum mekar untuk tetua betina reseptif. Tepung sari diambil dari bunga tetua jantan yang telah antesis. Selanjutnya tepung sari ditempelkan ke stigma betina yang reseptif. Bunga yang telah diserbuki ditutup dengan selotif dan diberi label yang berisi informasi genotipe tetua persilangan dan tanggal persilangan. Selfing dilakukan dengan menyungkup tanaman cabai, agar terhindar dari kontaminasi serbuk sari lain.
Karakter yang diamati adalah karakter komponen hasil, yaitu:
1. Umur berbunga (HST), dihitung saat tanaman telah memiliki minimal 1 bunga yang mekar sempurna.
2. Umur panen (HST), dihitung saat tanaman telah memiliki minimal 1 buah sudah layak panen.
3. Bobot buah (g), diambil rata-rata 5 buah setiap tanaman dari panen kedua dan diukur menggunakan timbangan analitik.
4. Diameter buah (mm), diambil rata-rata 5 buah setiap tanaman mulai dari panen kedua dan diukur mengunakan jangka sorong digital.
5. Tebal daging buah (mm), diambil rata-rata 5 buah setiap tanaman mulai dari panen kedua. Buah dibelah secara melintang dan diukur tebal daging buahnya menggunakan jangka sorong digital.
6. Panjang buah (cm), diambil rata-rata 5 buah setiap tanaman mulai dari panen kedua dan diukur dari pangkal sampai ujung buah.
7. Panjang tangkai buah (cm), diambil rata-rata 5 buah setiap tanaman mulai dari panen kedua dan diukur dari pangkal sampai ujung tangkai buah.
8. Jumlah buah per tanaman (buah), dihitung setiap kali panen dengan menjumlahkan jumlah buah tiap panen selama 8 minggu.
9. Bobot buah total per tanaman (g/tan), dihitung dengan menjumlahkan bobot buah tiap panen selama 8 minggu.
♀ P1 x P2 ♂
P1 x F1 x P2
BCP1 F2 BCP2
♀ P2 x P1 ♂
Analisis data dilakukan mengacu pada Limbongan et al. (2008) dan Arif et al. (2012) yaitu:
1. Uji normalitas pada populasi F2
Uji normalitas dilakukan untuk mengetahui sebaran populasi F2, untuk mengetahui aksi gen dari karakter-karakter yang diamati. Uji normalitas menggunakan metode Kolmogorov-smirnov.
2. Efek maternal
Uji efek maternal menggunakan pengujian dua populasi dengan uji t-student
pada nilai tengah populasi F1 dan F1R. Rumus uji-t mengacu pada (Steel dan Torrie 1981):
= µ − µ− Keterangan
µF1= Nilai Tengah populasi F1 S F1 = Simpangan baku populasi F1 µF1R= Nilai Tengah populsi F1R S F1R = Simpangan baku populasi F1R 3. Jumlah gen pengendali karakter
Jumlah gen pengendali karakter (menunjukkan jumlah kelompok gen yang terdiri dari beberapa gen minimal dari setiap kelompok) mengacu pada Mather dan Jinks (1982):
� = µ� − µ� Keterangan:
n = Jumlah faktor efektif (gen pengendali) VE = ragam lingkungan VF2 = ragam populasi F2 µP1 = Nilai tengah P1 H = 4((VBCP1+VBCP2)-(VF2-VE)) µP2 = Nilai tengah P2 4. Besaran nilai derajat dominansi
Pendugaan besaran nilai dominansi menggunakan analisis potensi rasio (hp) menurut Petr dan Frey (1966):
ℎ� =µ � − µ �µ − µ � Keterangan:
µF1 = Nilai rata-rata F1
µMP = Nilai tengah/mid parent kedua tetua µHP = Nilai tengah tetua tertinggi (high parent)
Derajat dominansi diduga dengan menggunakan tabel klasifikasi derajat dominansi berdasarkan potensi rasio pada Tabel 3.1.
Tabel 3.1 Klasifikasi derajat dominansi berdasarkan potensi rasio (hp) Nilai hp Derajat Dominansi
hp < -1 Overdominan
hp = -1 Resesif sempurna
-1 < hp < 0 Resesif parsial
hp = 0 Tidak ada dominansi/aditif 0 < hp < 1 Dominan parsial
hp = 1 Dominan sempurna
5. Kelayakan model genetik
Pendugaan kelayakan model genetik mengacu pada Mather dan Jink (1982). Uji skala individu digunakan model tiga parameter yang dijelaskan oleh Mather and Jinks (1982), yaitu: A=2BCP1-P1-F1; B=2BCP2-P2-F1; C=4F2-2F1-P1-P2. untuk menduga kesesuaian model aditif-dominan. Apabila hasil uji skala individu tidak ada sesuai dengan model aditif-dominan maka dilanjutkan dengan uji skala gabungan (m = ½P1 + ½ P2 + 4F2 - 2BCP1 - 2BCP2; [d] = ½P1 - ½P2; [h]=6BCP1 + 6BCP2 - 8F2 - F1 - 1½P1 - 1½P2; [i]= 2BCP1 + 2BCP2 - 4F2; [j]=2BCP1 - P1 – 2BCP2 + P2; [l]=P1 + P2 + 2F1 + 4F2 – 4BCP1
– 4BCP2) untuk menambahkan kontribusi dari epistasis (interaksi non-alelik). Uji ini menghasilkan dugaan untuk tiga parameter mean (m), efek aditif [d], efek dominan [h] selain itu juga menghasilkan dugaan tiga parameter epistasis interaksi aditif x aditif [i], interaksi aditif x dominan [j] dan interaksi dominan x dominan [l].
6. Komponen ragam
Komponen ragam yang dihitung adalah ragam fenotipe (VF2), lingkungan (VE), genotipe (VG), silang balik (backcross) (VBC), ragam aditif (VA), dan ragam dominan (VD). Ragam fenotipe berasal dari ragam F2.
7. Pendugaan nilai heritabilitas
Pendugaan nilai heritabilitas terdiri dari nilai heritabilitas dalam arti luas mengacu pada Allard (1960); Syukur et al. (2015) dan heritabilitas dalam arti sempit megacu pada Warner (1952).
ℎ � = � − � + �� + �� /�
ℎ�� = � − � � + � �
� Keterangan:
3.3Hasil dan Pembahasan
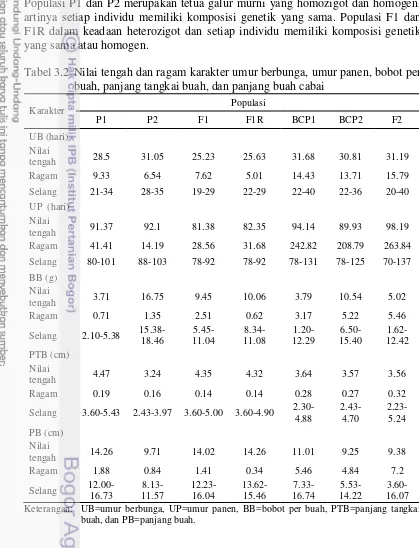
Nilai tengah, ragam, dan selang data populasi biparental untuk karakter umur berbunga, umur panen, bobot per buah, panjang tangkai buah, dan panjang buah disajikan pada Tabel 3.2. Populasi F2 merupakan populasi dengan tingkat keragaman tertinggi, diikuti oleh populasi BCP1 dan BCP2. Populasi P1, P2, F1 dan F1R lebih seragam dibanding BCP1, BCP2, dan F2. Keragaman tertinggi pada populasi F2 dikarenakan populasi ini merupakan segregasi yang paling maksimal. Populasi P1 dan P2 merupakan tetua galur murni yang homozigot dan homogen, artinya setiap individu memiliki komposisi genetik yang sama. Populasi F1 dan F1R dalam keadaan heterozigot dan setiap individu memiliki komposisi genetik yang sama atau homogen.
Tabel 3.2 Nilai tengah dan ragam karakter umur berbunga, umur panen, bobot per buah, panjang tangkai buah, dan panjang buah cabai
Karakter Populasi
Selang 3.60-5.43 2.43-3.97 3.60-5.00 3.60-4.90 2.30-4.88 Keterangan: UB=umur berbunga, UP=umur panen, BB=bobot per buah, PTB=panjang tangkai
Selang data BCP2 untuk karakter bobot per buah memiliki nilai maksimum yang melebihi nilai maksimum pada populasi F2. Populasi BCP2 merupakan hasil silang balik ke tetua P2 yang memiliki bobot buah jauh lebih tinggi dibandingkan tetua P1, hal inilah yang mengakibatkan nilai maksimum BCP2 lebih tinggi dibandingkan F2. Tingkat keragaman karakter bobot per buah pada populasi F2 tetap lebih tinggi daripada populasi BCP2 yakni sebesar 5.46 dibandingkan ragam populasi BCP2 sebesar 5.22 (Tabel 3.2).
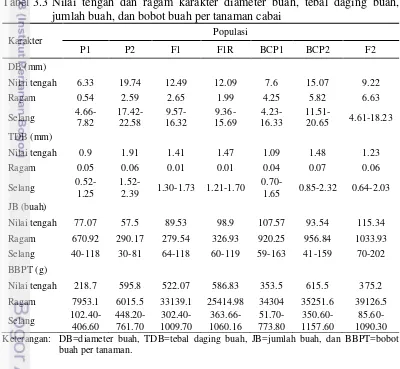
Nilai tengah, ragam, dan selang data populasi biparental untuk karakter diameter buah, tebal daging buah, jumlah buah, dan bobot buah per tanaman disajikan pada Tabel 3.3. Populasi F2 merupakan populasi dengan tingkat keragaman tertinggi, diikuti oleh populasi BCP1 dan BCP2. Populasi P1, P2, F1 dan F1R lebih seragam dibanding BCP1, BCP2, dan F2. Selang data BCP2 pada karakter diameter buah, tebal daging buah, dan bobot buah per tanaman nilai maksimumnya melebihi selang data F2. Hal ini diduga akibat dari silang balik ke tetua P2 yakni IPB C5 (Cabai besar) yang nilai tengahnya tinggi, sehingga nilai maksimumnya lebih baik daripada F2. Populasi F2 pada karakter-karakter tersebut tetap memiliki tingkat keragaman yang lebih tinggi dibandingkan populasi BCP2 walaupun dari nilai maksimumnya lebih baik pada populasi BCP2.
Tabel 3.3 Nilai tengah dan ragam karakter diameter buah, tebal daging buah, jumlah buah, dan bobot buah per tanaman cabai
Karakter Populasi Keterangan: DB=diameter buah, TDB=tebal daging buah, JB=jumlah buah, dan BBPT=bobot
Umur berbunga (HST)
3.3.1 Uji Normalitas pada Populasi F2
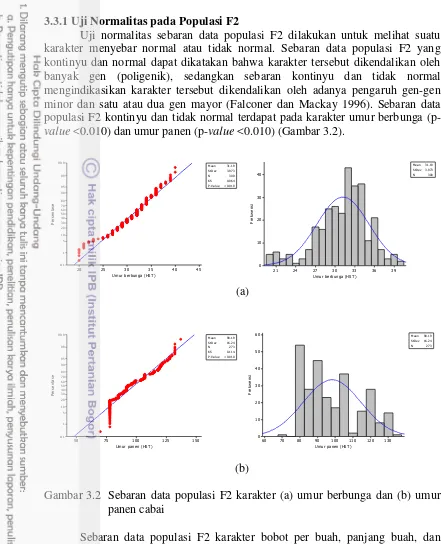
Uji normalitas sebaran data populasi F2 dilakukan untuk melihat suatu karakter menyebar normal atau tidak normal. Sebaran data populasi F2 yang kontinyu dan normal dapat dikatakan bahwa karakter tersebut dikendalikan oleh banyak gen (poligenik), sedangkan sebaran kontinyu dan tidak normal mengindikasikan karakter tersebut dikendalikan oleh adanya pengaruh gen-gen minor dan satu atau dua gen mayor (Falconer dan Mackay 1996). Sebaran data populasi F2 kontinyu dan tidak normal terdapat pada karakter umur berbunga
(p-value <0.010) dan umur panen (p-value <0.010) (Gambar 3.2).
(a)
(b)
Gambar 3.2 Sebaran data populasi F2 karakter (a) umur berbunga dan (b) umur panen cabai
Sebaran data populasi F2 karakter bobot per buah, panjang buah, dan diameter buah disajikan pada Gambar 3.3. Sebaran data populasi F2 kontinyu dan normal terdapat pada karakter panjang buah (p-value >0.150) dan tebal daging buah
(p-value >0.150). Hal ini dapat diartikan karakter panjang buah dikendalikan oleh
banyak gen (poligenik). Karakter bobot per buah dan diameter buah memiliki sebaran yang tidak normal (p-value <0.010) dan kontinyu. Sebaran kontinyu dan tidak normal mengindikasikan karakter bobot per buah dan diameter buah dikendalikan oleh adanya pengaruh gen-gen minor dan satu atau dua gen mayor.
(a)
(b)
(c)
Gambar 3.3 Sebaran data populasi F2 karakter (a) bobot per buah (b) panjang buah dan (c) diameter buah cabai
Sebaran data populasi F2 karakter panjang tangkai buah dan tebal daging buah disajikan pada Gambar 3.4. Karakter panjang tangkai buah dan tebal daging buah memiliki sebaran yang normal (p-value <0.150) dan kontinyu. Sebaran kontinyu dan normal mengindikasikan karakter bobot per buah dan diameter buah dikendalikan oleh banyak gen (poligenik) (Falconer dan Mackay 1996).
(a)
(b)
Gambar 3.4 Sebaran data populasi F2 karakter (a) panjang tangkai buah dan (b) tebal daging buah cabai
Sebaran data populasi F2 karakter jumlah buah dan bobot buah per tanaman disajikan pada Gambar 3.5. Sebaran data populasi F2 karakter jumlah buah dan bobot buah per tanaman kontinyu dan tidak menyebar normal (p-value <0.010). Pengaruh gen-gen minor dan satu atau dua gen mayor merupakan gen-gen pengendali dari karakter jumlah buah dan bobot buah per tanaman tersebut. Beberapa penelitian juga melaporkan hal yang sama untuk karakter bobot buah per tanaman dan jumlah buah. Hasil penelitian Nura (2015) menunjukkan karakter jumlah buah memiliki sebaran data F2 tidak menyebar normal dan kontinyu, sedangkan hasil penelitian Arif (2010) menunjukkan karakter bobot buah per tanaman memiliki sebaran data F2 tidak menyebar normal dan kontinyu.
(a)
(b)
Gambar 3.5 Sebaran data populasi F2 karakter (a) jumlah buah dan (b) bobot buah per tanaman cabai
3.3.2 Efek Maternal
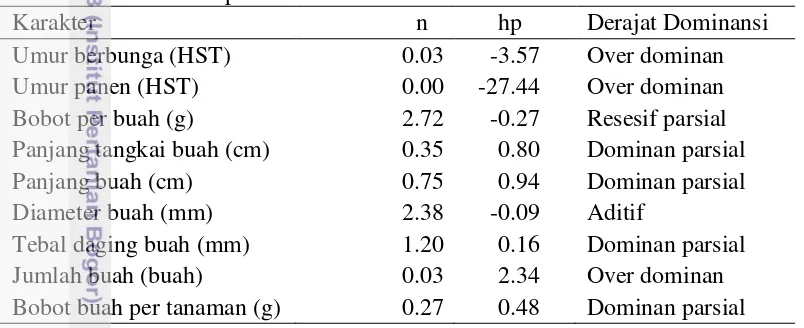
Uji efek maternal dilakukan untuk menduga suatu karakter dipengaruhi oleh gen yang berada di luar inti atau ekstrakromosomal. Uji efek maternal menunjukkan tidak adanya perbedaan yang nyata pada semua karakter komponen hasil cabai antara F1 dan F1R (Tabel 3.4). Hal ini menunjukkan bahwa tidak ada pengaruh tetua betina terhadap pewarisan karakter-karakter komponen hasil cabai. Hal ini dapat juga diartikan bahwa gen-gen dalam inti yang mengendalikan karakter komponen hasil pada penelitian ini.
Tabel 3.4 Uji efek maternal populasi F1 dan F1R pada karakter komponen hasil Bobot buah per tanaman (g) 522.07±196.99 586.83±228.16 -1.04 0.30
Beberapa penelitian juga menunjukkan hal yang sama seperti pada karakter umur berbunga, diameter buah, panjang buah (Hilmayanti et al. 2006); karakter umur panen dan bobot per buah (Arif et al. 2012) tidak terdapat pengaruh tetua betina. Pewarisan karakter pada penelitian ini tidak perlu dipisahkan antara F1 dan F1R, karena F2 yang dihasilkan dari F1 ataupun F1R segregasinya tidak berbeda. 3.3.3 Jumlah Kelompok Gen Pengendali Karakter dan Derajat Dominansi
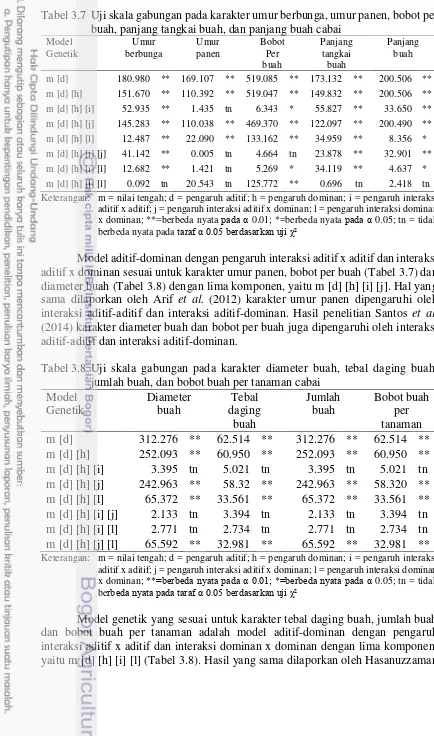
Jumlah gen-gen efektif yang mengendalikan umur berbunga, umur panen, panjang tangkai buah, panjang buah, jumlah buah dan bobot buah per tanaman adalah 1 kelompok gen efektif. Arif et al. (2012) melaporkan karakter umur berbunga jumlah gen-gen efektif yang mengendalikan sebanyak 1 kelompok gen, sedangkan bobot per buah dilaporkan dikendalikan oleh 18 kelompok gen pengendali. Perbedaan jumlah kelompok gen pengendali pada karakter bobot per buah diduga karena populasi yang digunakan berbeda. Jumlah gen-gen efektif karakter tebal daging buah dan diameter buah dikendalikan oleh 2 kelompok gen efektif, sedangkan karakter bobot per buah dan diameter buah dikendalikan oleh 3 kelompok gen efektif (Tabel 3.5).
Tabel 3.5 Jumlah kelompok gen pengendali (n) dan derajat dominansi (hp) pada karakter komponen hasil cabai
Karakter n hp Derajat Dominansi
Umur berbunga (HST) 0.03 -3.57 Over dominan
Umur panen (HST) 0.00 -27.44 Over dominan
Bobot per buah (g) 2.72 -0.27 Resesif parsial Panjang tangkai buah (cm) 0.35 0.80 Dominan parsial
Panjang buah (cm) 0.75 0.94 Dominan parsial
Diameter buah (mm) 2.38 -0.09 Aditif
Tebal daging buah (mm) 1.20 0.16 Dominan parsial
Jumlah buah (buah) 0.03 2.34 Over dominan
3.3.4 Kelayakan Model Genetik 3.3.4.1 Uji Skala Individu
Uji skala individu yang berbeda nyata pada A, B dan C baik itu satu atau lebih mengindikasikan bahwa pada penelitian ini terdapat epistasis atau interaksi antar lokus (Tabel 3.6). Nilai A semua karakter menunjukkan nilai yang berbeda sangat nyata kecuali bobot buah per tanaman, artinya model aditif-dominan tidak sesuai untuk model genetik. Walaupun nilai A dan B karakter bobot buah per tanaman tidak berbeda nyata, model aditif-dominan tidak dapat digunakan karena nilai C karakter ini berbeda nyata. Apabila nilai C signifikan mengindikasikan adanya tipe interaksi [l] yaitu pengaruh interaksi dominan x dominan. Menurut Hill et al. (1998) apabila model aditif dominan tidak sesuai maka perlu dilakukan uji ke model lainnya yang melibatkan interaksi gen antar lokus.
Tabel 3.6 Uji skala pada karakter komponen hasil cabai
Karakter A B C
Umur berbunga (HST) 9.629 ** 5.543 ** 14.760 ** Umur panen (HST) 13.409 ** 9.166 ** 45.718 ** Bobot per buah (g) -4.842 ** -2.588 ** -18.959 ** Panjang tangkai buah (cm) -1.531 ** -0.458 ** -2.161 ** Panjang buah (cm) -6.741 ** -4.960 ** -13.325 ** Diameter buah (mm) -3.582 ** -2.044 ** -14.073 ** Tebal daging buah (mm) -0.180 ** -0.057 tn -0.749 ** Jumlah buah (buah) 37.426 ** 39.388 ** 146.428 ** Bobot buah per tanaman (g) 2.461 tn 39.658 tn -213.872 * Keterangan: **= berbeda nyata pada α 0.01, *= berbeda nyata pada α 0.05, dan tn= tidak
berbeda nyata pada α 0.05
3.3.4.2 Uji Skala Gabungan
Parameter model genetik menurut Mather dan Jinks (1982) digunakan untuk menguji tipe interaksi antar lokus. Model genetik yang diduga paling sesuai adalah ketika nilai χ2 tidak berbeda nyata. Namun apabila lebih dari satu nilai χ2 yang tidak berbeda nyata, untuk menentukan model yang sesuai dapat dilakukan dengan melihat nilai duga yang terkecil. Model genetik yang sesuai untuk karakter umur berbunga, panjang tangkai buah dan panjang buah adalah model aditif-dominan dengan pengaruh interaksi aditif x dominan dan interaksi dominan x dominan dengan lima komponen, yaitu m [d] [h] [j] [l] (Tabel 3.7). Model aditif-dominan dengan pengaruh interaksi aditif x dominan dan interaksi dominan x dominan umur berbunga dan panjang buah sama dengan hasil penelitian Hasanuzzaman dan Golam (2011). Sedangkan Patil (2011) juga menghasilkan model aditif-dominan dengan pengaruh interaksi aditif x dominan dan interaksi dominan x dominan untuk karakter panjang buah.
Tabel 3.7 Uji skala gabungan pada karakter umur berbunga, umur panen, bobot per buah, panjang tangkai buah, dan panjang buah cabai
Model Keterangan: m = nilai tengah; d = pengaruh aditif; h = pengaruh dominan; i = pengaruh interaksi
aditif x aditif; j = pengaruh interaksi aditif x dominan; l = pengaruh interaksi dominan x dominan; **=berbeda nyata pada α 0.01; *=berbeda nyata pada α 0.05; tn = tidak berbeda nyata pada taraf α 0.05 berdasarkan uji χ²
Model aditif-dominan dengan pengaruh interaksi aditif x aditif dan interaksi aditif x dominan sesuai untuk karakter umur panen, bobot per buah (Tabel 3.7) dan diameter buah (Tabel 3.8) dengan lima komponen, yaitu m [d] [h] [i] [j]. Hal yang sama dilaporkan oleh Arif et al. (2012) karakter umur panen dipengaruhi oleh interaksi aditif-aditif dan interaksi aditif-dominan. Hasil penelitian Santos et al.
(2014) karakter diameter buah dan bobot per buah juga dipengaruhi oleh interaksi aditif-aditif dan interaksi aditif-dominan.
Tabel 3.8 Uji skala gabungan pada karakter diameter buah, tebal daging buah, jumlah buah, dan bobot buah per tanaman cabai
Model Keterangan: m = nilai tengah; d = pengaruh aditif; h = pengaruh dominan; i = pengaruh interaksi
aditif x aditif; j = pengaruh interaksi aditif x dominan; l = pengaruh interaksi dominan x dominan; **=berbeda nyata pada α0.01; *=berbeda nyata pada α 0.05; tn = tidak
berbeda nyata pada taraf α0.05 berdasarkan uji χ²
dan Golam (2011) pada karakter buah dan penelitian Navhale et al. (2014) untuk karakter jumlah buah dan bobot buah per tanaman.
Pendugaan parameter model genetik yang cocok mengacu pada Navhale et al. (2014) dengan melihat nilai karakter yang berbeda nyata seperti: m, [d], [h], [i], [j], dan [l] mengindikasikan adanya aksi gen aditif, dominan dan epistasis serta interaksinya. Apabila nilai [h] dan [l] signifikan serta memiliki tanda yang sama disebut sebagai tipe komplementasi, sedangkan apabila nilai [h] dan [l] signifikan serta memiliki tanda yang berbeda disebut sebagai tipe duplikasi (Mather and Jinks, 1982). Hasil penelitian menunjukkan karakter umur berbunga, panjang tangkai buah, dan panjang buah (Tabel 3.8) memiliki interaksi gen komplementasi. Adanya interaksi gen komplementasi ini program pemuliaan dapat dilanjutkan dengan memanfaatkan efek heterosis dalam merakit suatu varietas hibrida. Hal ini dikarenakan efek gen non-aditif lebih besar pada karakter tersebut. Menurut Hasanuzzaman dan Golam (2011) dengan adanya interaksi gen komplementasi seleksi pada generasi awal segregasi tidak akan memberikan kontribusi yang besar untuk perbaikan karakter-karakter ini. Sedangkan menurut Navhale et al. (2014) modifikasi seleksi bulk dianjurkan apabila terdapat interaksi gen komplementasi, dimana seleksi dilakukan setelah tangkat homozigositas sudah cukup tinggi. Tabel 3.9 Perkiraan efek gen pada karakter umur berbunga, umur panen, bobot per
buah, panjang tangkai buah, dan panjang buah cabai Efek Komplementasi - Duplikasi Komplementasi Komplementasi Keterangan: m = nilai tengah; d = pengaruh aditif; h = pengaruh dominan; i = pengaruh interaksi
aditif x aditif; j = pengaruh interaksi aditif x dominan; l = pengaruh interaksi dominan x dominan; **=berbeda nyata pada α 0.01; *=berbeda nyata pada α 0.05; tn = tidak berbeda nyata pada taraf α 0.05 berdasarkan uji t
Karakter bobot per buah (Tabel 39), tebal daging buah, dan bobot buah per tanaman (Tabel 3.10) memiliki interaksi gen duplikasi karena [h] dan [l] signifikan dengan arah tanda yang berbeda. Komplementasi akan meningkatkan heterosis, sedangkan duplikasi menyebabkan heterosis turun. Hal ini akan mengurangi manfaat yang terjadi dari heterozigositas karena pembatalan dominasi dan efek epistatis (Dhall dan Hundal, 2006). Pada kasus ini, menurut Sharma and sharma (1995) menunda seleksi akan lebih efektif untuk mendapatkan respon seleksi yang lebih baik.
agar efek gen dominan tidak hilang dengan harapan efek dominan tersebut terkumpul. Menurut Pathak et al. (2014) pemuliaan seleksi berulang resiprokal tampaknya menjadi metode terbaik karena akan memanfaatkan ketiga jenis efek gen aditif, dominan dan epistasis yang menghasilkan terkumpulnya rekombinan yang diinginkan pada generasi lanjut.
Tabel 3.10 Perkiraan efek gen pada karakter umur berbunga, umur panen, bobot per buah, panjang tangkai buah, dan panjang buah cabai
Efek
Keterangan: m = nilai tengah; d = pengaruh aditif; h = pengaruh dominan; i = pengaruh interaksi aditif x aditif; j = pengaruh interaksi aditif x dominan; l = pengaruh interaksi dominan x dominan; **=berbeda nyata pada α0.01; *=berbeda nyata pada α 0.05; tn = tidak
berbeda nyata pada taraf α 0.05 berdasarkan uji t
3.3.5 Komponen Ragam dan Heritabilitas
Nilai heritabilitas yang termasuk dalam kategori tinggi menunjukkan bahwa faktor genetik memiliki pengaruh yang lebih besar dibandingkan dengan lingkungan pada penampilan fenotipik suatu tanaman. Nilai heritabilitas tinggi khususnya heritabilitas arti sempit memainkan peran penting dalam meningkatkan efektivitas seleksi (Syukur et al. 2015). Heritabilitas arti luas pada semua karakter yang diuji berkisar antara 36.78% sampai 89.37% atau dapat diklasifikasikan dari sedang (25%<h<50%) hingga tinggi (>50%) (Tabel 3.11). Heritabilitas dalam arti luas pada semua karakter berada pada kisaran tinggi kecuali untuk karakter panjang tangkai buah dan tebal daging buah yang berada pada kisaran sedang. Beberapa penelitian cabai juga menunjukkan bahwa nilai heritabilitas dalam arti luas yang tinggi pada umur berbunga (Singh et al. 2014; Syukur and Rosidah 2014), umur panen (Nsabiera et al. 2013), bobot per buah dan panjang buah (Santos et al. 2014), diameter buah (Santos et al. 2014), jumlah buah (Sreelathakumary and Rajamony 2004) dan bobot buah per tanaman (Syukur et al. 2010).
umur berbunga tinggi, sedangkan diameter buah rendah. Hasil penelitian Ben-Chaim dan Paran (2000) serta Santos (2014) bobot per buah memiliki heritabilitas arti sempit yang tinggi. Nilai heritabilitas dalam arti sempit karakter umur berbunga dan bobot buah per tanaman berada pada kisaran rendah.
Tabel 3.11 Komponen ragam dan heritabilitas pada karakter komponen hasil cabai
Komponen UB UP BB PTB PB DB TDB JB BBPT
Ragam P1 9.33 41.41 0.71 0.19 1.88 0.54 0.05 670.92 7953.10 Ragam P2 6.54 14.19 1.35 0.16 0.84 2.59 0.06 290.17 6015.50 Ragam F1 7.62 28.56 2.51 0.14 1.41 2.65 0.01 279.54 33139.10 Ragam BCP1 14.43 242.82 3.17 0.28 5.46 4.25 0.04 920.25 34304.00 Ragam BCP2 13.71 208.79 5.22 0.27 4.84 5.82 0.07 956.84 35251.60 Ragam F2 15.79 263.84 5.46 0.32 7.20 6.63 0.06 1033.93 39126.50 Rasio aditif 0.43 0.32 0.64 0.56 0.70 0.68 0.91 0.31 0.37 Rasio non-aditif 0.57 0.68 0.36 0.44 0.30 0.32 0.09 0.69 0.63 h2
bs (%) 50.42 89.37 72.05 49.03 80.90 70.96 36.78 60.00 59.87 h2
ns (%) 21.76 28.83 46.24 27.25 56.84 48.13 33.33 18.45 22.23 Keterangan: UB=Umur berbunga, UP=Umur panen, BB=Bobot per buah, PTB=Panjang tangkai buah,
PB=Panjang buah, DB=Diameter buah, TDB=Tebal daging buah, JB=Jumlah buah dan BBPT=Bobot buah per tanaman.
3.4 Simpulan
4
SELEKSI GENOTIPE CABAI SEGREGAN TRANSGRESIF
Abstrak
Seleksi pada program pemuliaan tanaman dilakukan dalam rangka memilih kandidat galur yang berdaya hasil tinggi. Penelitian ini dilaksanakan pada bulan September 2013-Juli 2014 (pembentukan populasi) dan Agustus-Desember 2014 (pengujian). Pembentukan populasi dilakukan di perumahan IPB Alam Sinar Sari, sedangkan untuk pengujian dilakukan di Kebun Percobaan Leuwikopo, IPB. Penelitian bertujuan untuk memperoleh kandidat galur harapan segregan transgresif pada populasi F2 dengan memanfaatkan indeks seleksi terboboti. Hasil penelitian menunjukkan bahwa karakter panjang buah, jumlah buah, dan bobot per buah memiliki korelasi positif dan sangat nyata terhadap bobot buah per tanaman. Karakter panjang buah, jumlah buah, bobot per buah, dan bobot buah per tanaman digunakan sebagai dasar dalam pemilihan genotipe yang dilanjutkan ke generasi selanjutnya. Hasil seleksi indeks terpilih 30 genotipe F2, yakni F2120005-3, 14, 16, 22, 35, 36, 50, 56, 62, 70, 74, 87, 104, 115, 120, 125, 136, 141, 142, 145, 146, 147, 149, 176, 180, 181, 184, 185, 199, 215, dan 238. Terdapat genotipe segregan transgresif pada populasi F2 terseleksi untuk karakter umur berbunga, umur panen, panjang tangkai buah, panjang buah, jumlah buah, dan bobot buah per tanaman. Kata kunci: indeks seleksi, segregan transgresif, seleksi
Abstract
Selections in plant breeding program were conducted to choose high yield line candidates. The studies were conducted from September 2013 to July 2014 (population establishment) and from August 2014 to December 2014 (experiment). Population establishments took place in Alam Sinar Sari IPB Housing, whereas trials took place in Leuwikopo Experimental Field, IPB. This study aims to obtain transgressive segregant line candidates in F2 population by utilize selection weighted index. Results showed character of fruit length, number of fruit, and fruit weight had positive correlation and significant against yield. Character of fruit length, number of fruit, and fruit weight had positive and significant correlation toward yield. Character of fruit length, number of fruit, fruit weight, and yield were used as basis parameters for genotype selection for next generation. Thirty F2 genotype: F2120005-3, 14, 16, 22, 35, 36, 50, 56, 62, 70, 74, 87, 104, 115, 120, 125, 136, 141, 142, 145, 146, 147, 149, 176, 180, 181, 184, 185, 199, 215, and 238 were selected. There were selected segregan transgressive genotypes in F2 population for flowering age, harvesting age, pedicel length, fruit length, number of fruit, and yield characters.
4.1 Pendahuluan
Seleksi pada program pemuliaan tanaman dilakukan dalam rangka memilih kandidat galur yang berdaya hasil tinggi. Menurut Syukur et al. (2011) seleksi akan efektif jika populasi tersebut mempunyai keragaman genetik yang luas dan heritabilitas yang tinggi, sehingga hal tersebut sangat bermanfaat dalam proses seleksi. Heritabilitas yang tinggi dapat diartikan penampilan fenotipik lebih dipengaruhi oleh genetik dibandingkan pengaruh lingkungan. Seleksi pada karakter daya hasil akan menjadi lebih efektif apabila diikuti dengan seleksi pada karakter komponen hasil lainnya. Hal ini dikarenakan produktivitas cabai sangat dipengaruhi oleh karakter komponen hasil seperti bobot buah, diameter buah, panjang buah, tebal daging buah dan jumlah buah. Metode seleksi pada beberapa karakter sekaligus dapat dilakukan dengan beberapa metode. Salah satu metode yang dapat dilakukan adalah metode indeks seleksi.
Menurut Sumarno dan Zuraida (2006) metode seleksi berbasis indeks dapat menghasilkan genotipe-genotipe terbaik untuk karakter-karakter yang dilibatkan dalam penyusunan indeks. Seleksi akan memberikan respon yang baik apabila menggunakan kriteria seleksi yang tepat. Karakter-karakter yang digunakan sebagai karakter penyusun indeks merupakan karakter-karakter yang memberikan pengaruh cukup besar pada peningkatan daya hasil. Seleksi dilakukan pada populasi F2 yang merupakan populasi dengan segregasi atau tingkat keragaman paling tinggi. Genotipe-genotipe hasil segregasi generasi F2 ini terdapat genotipe-genotipe yang bersifat segregan transgresif, yaitu segregasi gen pada sifat-sifat kuantitatif dari zuriat hasil persilangan dua tetua yang memiliki jangkauan sebaran yang melampaui jangkauan sebaran kedua tetuanya (Poehlman dan Sleper 1996). Genotipe-genotipe yang diduga segregan transgresif ini apabila dilanjutkan ke generasi selanjutnya akan menjadi galur harapan dengan ragam dalam famili yang rendah. Famili segregan transgresif ini diharapkan tidak perlu menunggu hingga 6-7 generasi untuk menjadi galur murni, cukup dengan 4-5 generasi sudah menjadi galur murni yang homogen homozigot. Penelitian ini bertujuan untuk memperoleh kandidat galur harapan segregan transgresif pada populasi F2 dengan memanfaatkan seleksi indeks terboboti.
4.2 Bahan dan Metode
Penelitian ini dilaksanakan bersamaan dengan percobaan sebelumnya pada bulan September 2013-Juli 2014 (pembentukan populasi) dan Agustus-Desember 2014 (pengujian). Pembentukan populasi dilakukan di perumahan IPB Alam Sinar Sari, sedangkan untuk pengujian dilakukan di Kebun Percobaan Leuwikopo, IPB.
Karakter yang diamati adalah karakter komponen hasil, yaitu:
1. Umur berbunga (HST), dihitung saat tanaman telah memiliki minimal 1 bunga yang mekar sempurna.
2. Umur panen (HST), dihitung saat tanaman telah memiliki minimal buah sudah layak panen.
3. Bobot buah (g), diambil rata-rata 5 buah setiap tanaman dari panen kedua dan diukur menggunakan timbangan analitik.
4. Diameter buah (mm), diambil rata-rata 5 buah setiap tanaman mulai dari panen kedua dan diukur mengunakan jangka sorong digital.
5. Tebal daging buah (mm), diambil rata-rata 5 buah setiap tanaman mulai dari panen kedua. Buah dibelah secara melintang dan diukur tebal daging buahnya menggunakan jangka sorong digital.
6. Panjang buah (cm), diambil rata-rata 5 buah setiap tanaman mulai dari panen kedua dan diukur dari pangkal sampai ujung buah.
7. Panjang tangkai buah (cm), diambil rata-rata 5 buah setiap tanaman mulai dari panen kedua dan diukur dari pangkal sampai ujung tangkai buah.
8. Jumlah buah per tanaman (buah), dihitung setiap kali panen dengan menjumlahkan jumlah buah tiap panen selama 8 minggu.
9. Bobot buah total per tanaman (g/tan), dihitung dengan menjumlahkan bobot buah tiap panen selama 8 minggu.
Seleksi dilakukan berdasarkan indeks seleksi terboboti dengan melihat korelasi antara karakter komponen hasil terhadap bobot buah per tanaman dengan rumus yang mengacu pada Falconer (1976):
I = a1Z1+ a2Z2 + a3Z3 + …. + anZn, dimana Zn = � − �̅ / √� Keterangan:
I = nilai indeks total suatu fenotipe a = pembobot masing-masing peubah Z = nilai fenotipe yang telah distandardisasi x = rataan peubah dari suatu genotipe
�̅ = rataan peubah dari suatu total seluruh genotipe
� = ragam
Pembobotan yang digunakan sebagai berikut: panjang buah (+1), jumlah buah (+2), bobot per buah (+1), dan bobot buah per tanaman (+3). Hasil seleksi berdasarkan indeks seleksi dilihat kembali nilai tengah semua karakter yang diamati. Apabila memiliki nilai tengah yang melebihi nilai tengah tetua tertinggi diduga sebagai genotipe segregan transgresif.
4.3 Hasil dan Pembahasan 4.3.1 Seleksi Berdasarkan Indeks
komponen hasil yang diamati memiliki korelasi yang positif dan sangat nyata terhadap bobot buah per tanaman, kecuali karakter umur berbunga dan umur panen (Tabel 4.1).
Tabel 4.1 Korelasi pearson karakter komponen hasil terhadap bobot buah per tanaman cabai
Karakter Koefisien korelasi
Umur berbunga (HST) -0.126 tn
Umur panen (HST) -0.092 tn
Bobot per buah (g) 0.601**
Panjang tangkai buah (cm) 0.457**
Panjang buah (cm) 0.509**
Diameter buah (mm) 0.561**
Tebal daging buah (mm) 0.309**
Jumlah buah (buah) 0.745**
Keterangan: ** = berkorelasi nyata pada α 0.01; * = berkorelasi nyata pada α 0.05; tn = tidak berkorelasi nyata pada α 0.05
Karakter bobot per buah, panjang buah, dan jumlah buah dipilih sebagai karakter dasar dalam menentukan genotipe yang akan dilanjutkan ke generasi selanjutnya. Pemilihan ketiga karakter ini sebagai dasar dalam seleksi indeks dikarenakan memiliki nilai koefisien korelasi yang lebih baik dibandingkan dengan karakter lainnya. Karakter diameter buah juga memiliki nilai koefisien korelasi yang cukup baik yakni sebesar 0.561 (Tabel 4.1), tetapi tidak digunakan sebagai karakter seleksi indeks. Hal ini dikarenakan karakter diameter buah pada penelitian ini bukan menjadi karakter utama dalam kegiatan seleksi. Berdasarkan nilai heritabilitas dan korelasi maka terpillih karakter bobot per buah, jumlah buah, dan panjang buah serta bobot buah buah per tanaman yang menjadi karakter seleksi indeks pada kegiatan seleksi.
Pembobotan yang dilakukan berbeda antar karakter yang dipilih. Karakter bobot buah per tanaman memiliki bobot yang paling tinggi, yakni diberi skor 3 karena penelitian ini bertujuan untuk mendapatkan genotipe dengan produktivitas yang tinggi. Jumlah buah per tanaman diberi skor 2 karena karakter ini memiliki korelasi yang paling tinggi dibandingkan karakter lainnya. Panjang buah dan bobot buah masing-masing diberi skor 1 karena diharapkan terpilih cabai keriting yang memiliki buah cukup panjang dan memiliki bobot lebih baik dari tetua cabai keriting.
Tabel 4.2 Nilai indeks seleksi karakter panjang buah, jumlah buah per tanaman, bobot per buah, dan bobot buah per tanaman cabai
Genotipe PB JB BB BBPT I
F2120005-3 2.14 1.07 2.09 2.46 13.73
F2120005-14 1.25 0.05 1.65 0.95 5.86
F2120005-16 1.25 0.87 1.81 2.27 11.62
F2120005-22 0.26 0.96 0.55 1.01 5.74
F2120005-35 1.24 0.74 0.60 1.99 9.30
F2120005-36 0.75 0.83 1.57 1.50 8.46
F2120005-50 1.22 0.66 0.34 1.04 5.99
F2120005-56 1.82 0.64 0.74 0.82 6.27
F2120005-62 2.14 4.15 1.39 3.61 22.65
F2120005-70 1.28 0.36 0.91 1.63 7.77
F2120005-74 0.40 1.43 0.61 1.51 8.41
F2120005-87 0.78 3.50 0.20 2.62 15.84
F2120005-104 0.67 0.33 1.41 1.43 7.04
F2120005-115 1.29 -0.29 2.22 1.58 7.66
F2120005-120 2.18 -0.76 2.48 -0.71 1.00
F2120005-125 1.38 -0.03 2.11 1.72 8.58
F2120005-136 1.91 1.71 1.71 2.40 14.24
F2120005-141 -0.08 2.19 -0.52 0.81 6.20
F2120005-142 0.82 1.15 1.18 2.38 11.46
F2120005-145 0.09 1.65 -0.41 1.00 5.99
F2120005-146 1.65 1.11 0.45 0.90 7.02
F2120005-147 0.28 3.05 -0.13 2.11 12.56
F2120005-149 0.89 1.33 0.77 1.35 8.35
F2120005-176 0.25 1.07 1.01 1.56 8.07
F2120005-180 1.38 1.86 1.57 2.21 13.29
F2120005-181 1.16 1.09 0.47 1.06 6.99
F2120005-184 1.63 0.53 0.77 1.19 7.02
F2120005-185 0.78 0.77 0.65 1.41 7.19
F2120005-199 0.59 1.37 0.77 1.91 9.82
F2120005-215 2.12 1.05 1.18 1.86 10.96
F2120005-238 0.90 1.89 0.38 0.41 6.28
Keterangan: PB=panjang buah, JB=jumlah buah, BB=bobot per buah, BBPT=bobot buah per tanaman, I=nilai indeks
4.3.2 Seleksi Genotipe Cabai Segregan Transgresif