SAGITA PUJIANTI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Kambing, Teluk Banten. Dibimbing oleh RIKA RAFFIUDIN dan SUSETIONO.
Meiofauna adalah kelompok hewan berukuran antara 0.063-1 mm. Meiofauna interstisiil hidup di rongga antar butir sedimen dan dalam distribusi verikal dapat ditemukan sampai dengan kedalaman 10 cm. Tujuan penelitian adalah mempelajari distribusi vertikal meiofauna interstisiil pada padang lamun dengan luasan tutupan berbeda di Pulau Kambing, Teluk Banten. Empat stasiun dipilih berdasarkan persentase kepadatan luasan tutupan lamun yaitu padat, sedang, rendah, dan tidak ditumbuhi lamun. Enhalus acoroides adalah spesies lamun yang mendominasi di P. Kambing. Kepadatan meiofauna meningkat di stasiun dengan tutupan lamun padat dibandingkan daerah tanpa ditumbuhi lamun. Kepadatan meiofauna interstisiil umumnya melimpah sampai dengan kedalaman 10 cm. Kepadatan meiofauna paling tinggi pada kedalaman 1-4 cm karena sedimen relatif stabil dan teroksigenisasi dengan baik. Hasil penelitian ini didapat sebelas taksa meiofauna di P. Kambing yaitu Nematoda, Copepoda, Polychaeta, Oligochaeta, Foraminifera, Ostracoda, Nauplii, Isopoda, Acari, Kinorhyncha, dan Tardigrada. Nematoda adalah taksa yang paling umum di tiap stasiun dan di tiap distribusi vertikal. Lima famili terbesar adalah Leptosomatidae, Linhomoeidae, Monhysteridae, Oxystominidae, dan Pelagonematoidae.
ABSTRACT
SAGITA PUJIANTI. Vertical Distributin of Meiofauna Interstitial in Seagrass Bed in Kambing Island, Banten Bay. Supervised by RIKA RAFFIUDIN dan SUSETIONO.
DISTRIBUSI VERTIKAL MEIOFAUNA INTERSTISIIL PADA PADANG
LAMUN, DI PULAU KAMBING, TELUK BANTEN
SAGITA PUJIANTI
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains pada
Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Menyetujui,
Pembimbing I Pembimbing II
Dr. Ir. Rika Raffiudin, M.Si. NIP 131999583
Drs. Susetiono, M.Sc. NIP 320005404
Mengetahui :
Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam Institut Pertanian Bogor
Dr. Drh. Hasim DEA. NIP 131578806
PRAKATA
Puji syukur penulis panjatkan kepada Allah SWT berkat rahmat serta rizki-Nya, dan rencana-Nya yang Maha Indah sehingga penulis bisa menyelesaikan karya tulis ini. Shalawat serta salam penulis panjatkan kepada Nabi Muhammad SAW, sang pembawa kebenaran hakiki.
Terima kasih penulis ucapkan kepada Dr. Ir. Rika Raffiudin, M.Si. dan Drs. Susetiono, M.Sc. atas saran serta bimbingannya selama penyusunan karya ilmiah ini. Ucapan terima kasih disampaikan kepada Prof. Ir. Alex Hartana atas diskusi dan peranan beliau sebagai penguji dan wakil komisi pendidikan dalam ujian skripsi.
Ungkapan terima kasih juga ditujukan kepada kepada Bapak Arifin dan Bapak Wawan Kiswara atas bantuannya selama di lapangan. Terima kasih juga penulis sampaikan kepada Pusat Penelitian Oseangrafi (P20), LIPI, Jakarta yang telah membantu selama penelitian.
Terima kasih kepada semua pihak yang membantu, kepada dosen-dosen Zoologi atas bimbingannya, kepada rekan seperjuangan Citra, Fajri, Iwa, dan Taufiq atas segala bantuannya, kepada Andri, Cicit, Dian, Mbem, Muley, Novan, Sarah, dan semua teman-teman Biologi 40 yang telah memberikan informasi, arahan, dan bantuan selama penelitian ini.
Terima kasih penulis ucapkan Terima kasih penulis ucapkan kepada Ayah, Ibu, Vina dan Lenggo serta seluruh keluarga besar atas segala doa dan kasih sayangnya.
Semoga karya tulis ini bermanfaat.
Bogor, Februari 2008
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 17 Desember 1985 dari ayah Syahdanur A.M. dan ibu Afriyanti Z. Sani. Penulis merupakan putri pertama dari tiga bersaudara.
Tahun 2003 penulis lulus dari SMA Negri 29 Jakarta dan pada tahun yang sama masuk IPB melalui jalur Undangan Seleksi Masuk IPB. Penulis memilih Program Studi Biologi, Jurusan Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam.
DAFTAR ISI
Halaman
DAFTAR TABEL...vii
DAFTAR GAMBAR ...vii
DAFTAR LAMPIRAN ... vii
PENDAHULUAN...1
Latar Belakang ...1
Tujuan ...1
Waktu dan Tempat ...1
BAHAN DAN METODE ...1
Bahan dan Alat ...1
Deskripsi Tempat Pengambilan Contoh ...1
Penentuan Stasiun ...2
Identifikasi Lamun ...2
Pengambilan Contoh Sedimen ...2
Pengukuran Parameter Lingkungan ...2
Ekstraksi Meiofauna...2
Analisis Total Berat Organik (TBO) ...2
Analisis Butiran Sedimen...2
Identifikasi Meiofauna ...3
Analisis Data ...3
HASIL...3
Kepadatan Meiofauna Tiap Stasiun dengan Perbedaan Luasan Tutupan Vegetasi ...3
Distribusi Vertikal Meiofauna...3
Parameter Lingkungan ...3
Deskripsi Meiofauna di P. Kambing ...6
Famili Nematoda di P. Kambing ...8
Klasifikasi dan Deskripsi Lima Famili Nematoda dengan Jumlah Tertinggi di P. Kambing...8
Identifikasi Lamun ...9
PEMBAHASAN ...9
Kepadatan dan Keragaman Meiofauna di Tiap Stasiun ...9
Kepadatan Meiofauna Tiap Stasiun dengan Perbedaan Luasan Tutupan E. acoroides...10
Distribusi Vertikal Meiofauna Tiap Stasiun...11
Nematoda Tiap Stasiun dengan Perbedaan Karakterisasi Sedimen...11
Peran Meiofauna di Komunitas Bentik ...12
SIMPULAN... 12
DAFTAR TABEL
Halaman
1 Rataan dan standar baku kepadatan taksa meiofauna tiap stasiun dengan perbedaan luasan tutupan E. acoroides di P. Kambing………...
4 2 Nilai hasil pengukuran parameter lingkungan tiap stasiun dengan perbedaan luasan
tutupan E. Acoroides di P. Kambing ………….………...………. 6 3 Persentase komposisi sedimen tiap stasiun dengan perbedaan luasan tutupan E. Acoroides
di P. Kambing …..……….………..…. 6
DAFTAR GAMBAR
Halaman
1 Rataan kepadatan meiofauna tiap stasiun dengan perbedaan luasan tutupan E. acoroides di P. Kambing ………..……….
4 2 Dendrogram hasil persamaan Bray-Curtis untuk distibusi vertikal meiofauna tiap stasiun
dengan perbedaan luasan tutupan E. acoroides di P. Kambing ……….………….. 5
3 Kepadatan taksa meiofauna pada distribusi vertikal...………. 5
4 Nematoda ………..… 6
5 Amphid Nematoda ... 6
6 Copepoda ... 7
7 Nauplii ………...……….……….. 7
8 Polychaeta ... 7
9 Foraminifera ... 7
10 Oligochaeta... 7
11 Ostracoda ... 7
12 Isopoda ... 8
13 Acari ... 8
14 Kinorhyncha ... 8
15 Tardigrada ... 8
DAFTAR LAMPIRAN
Halaman 1 Peta Teluk Banten dan P. Kambing ... 172 Kepadatan dan keragaman meiofauna di tiap stasiun dan kedalaman di P. Kambing Teluk Banten (individu) ... 18 3 Kepadatan dan keragaman meiofauna di tiap stasiun dan kedalaman di P. Kambing, Teluk Banten (individu tiap 10 cm2) ... 20 4 Beberapa famili Nematoda di P. Kambing ... 22
5 Tipe mulut Nematoda pada stasiun pengamatan 1-4 di P. Kambing ………... 22
Latar Belakang
Meiofauna adalah istilah yang digunakan untuk kelompok hewan yang berukuran antara 0.063-1 mm (Bouwman 1987, diacu dalam Susetiono 1999). Anggota meiofauna dapat digolongkan ke dalam dua kelompok, yaitu meiofauna tetap dan meiofauna sementara. Meiofauna tetap adalah hewan yang sepanjang siklus hidup ukuran tubuhnya selalu berada di dalam kisaran ukuran meiofauna (Coull 1988). Taksa meiofauna tetap adalah Gastrotricha, Gnathosmulida, Kinorhyncha, Loricifera, Tardigrada, Mystacocarida, dan beberapa wakil dari Rotifera, Nematoda, Poly-chaeta, Copepoda, Ostracoda, Turbellaria, A-cari, Hydrozoa, Nemertina, Entroprocta, Gas-tropoda, Aplacophora, Branchiopoda, Holo-thuroidea, Tunicata, Priapulida, Oligochaeta, dan Sipuncula (Coull 1988). Meiofauna sementara adalah hewan yang ketika juvenil, berdasarkan ukuran tubuhnya, dikelompokkan ke dalam meiofauna, tetapi ketika dewasa dikelompok ke dalam makrofauna. Juvenil dari makrofauna umumnya adalah meiofauna sementara (Coull 1988). Kata meiofauna berasal dari bahasa Yunani, yaitu meio yang berarti lebih kecil (Higgins & Thiel 1988). Ukuran meiofauna lebih kecil dari makro-fauna, yaitu hewan yang tersaring di saringan 1 mm (Higgins & Thiel 1988), tetapi lebih besar dari mikrofauna (Giere 1993).
Meiofauna dapat ditemukan hampir di semua habitat air, mulai dari air tawar, payau sampai dengan laut. Di wilayah laut, meiofauna dapat ditemukan mulai dari daerah intertidal sampai dengan laut dalam. Meio-fauna juga dapat ditemukan diberbagai sedi-men (interstisiil) mulai dari lumpur sampai dengan kerikil kasar. Meiofauna yang hidup di sedimen dapat melakukan distribusi vertikal. Organisme yang hidup di sedimen laut disebut sebagai organisme bentik atau bentos (Brusca & Brusca 1990). Beberapa meiofauna di-temukan di atas sedimen, seperti pada daun makroalga, bagian tubuh hewan (celah koral, cacing tabung, duri Echinodermata), dan lamun (Coull 1988).
Lamun tumbuh di sedimen laut. Lamun termasuk kedalam divisi Spermatophyta; sub-divisi Angiosperma, dan mempunyai dua famili, yaitu Potamogetonaceae dan Hydro-charitaceae. Indonesia mempunyai 12 spesies lamun (Fortes 1990). Sebagian besar sedimen laut di dunia ditumbuhi lamun, kecuali di
Djamali 1995). Enhalus acoroides dan Thalassia hemprichii adalah dua spesies yang paling dominan di P. Kambing (Kiswara 2004).
Zona sebaran lamun di Teluk Banten mulai dari pantai sampai dengan tubir. Perbedaan zona sebaran lamun dijumpai dalam kom-posisi jenis lamun (vegetasi tunggal atau campuran) dan luasan tutupannya (jarang, sedang, tinggi atau sangat tinggi) (Kiswara 1992). Luasan tutupan lamun mempengaruhi kepadatan meiofauna.
Tujuan
Penelitian ini bertujuan meneliti distribusi vertikal meiofauna interstisiil pada padang lamun dengan luasan tutupan berbeda di Pulau Kambing, Teluk Banten.
Waktu dan Tempat
Penelitian ini dilaksanakan pada bulan Februari sampai dengan September 2007. Pengambilan contoh dilaksanakan pada bulan Mei 2007 di P. Kambing, Teluk Banten. Identifikasi dilakukan di Pusat Penelitian Oseangrafi (P20), LIPI, Jakarta.
BAHAN DAN METODE
Bahan dan Alat
Objek penelitian ini adalah meiofauna yang hidup di rongga antar butir sedimen (interstisiil). Bahan-bahan yang digunakan adalah contoh sedimen laut, formalin 4%, rose bengal, dan gliserol. Alat yang digunakan untuk penentuan stasiun adalah bingkai 50 x 50 cm2. Alat-alat yang digunakan untuk peng-ambilan contoh sedimen adalah core (p = 10 cm, d = 3 cm) dan (p = 30 cm, d = 5 cm) serta botol plastik. Alat-alat yang digunakan untuk menghitung parameter lingkungan adalah termometer, refraktometer, penguji tanah (soil tester), dan kertas tahan air (under water paper). Alat yang digunakan untuk identifi-kasi meiofauna adalah loop Irwin dan mikros-kop Leica ATC 2000 dan mikrosmikros-kop stereo Carton. Dokumentasi meiofauna menggu-nakan kamera Casio Exilim.
Deskripsi Tempat Pengambilan Contoh
175 km ke arah barat dari Jakarta. Teluk Banten dibatasi oleh Tanjung Potang di sebelah timur dan Tanjung Piatu di bagian barat. Teluk Banten terdiri dari beberapa Pulau, salah satunya adalah Pulau Kambing. Penduduk setempat menyebutnya Pulau Gedang. dasarkan luasan tutupan padang lamun. Empat stasiun tersebut berdasarkan kepadatan luasan tutupan padang lamun yaitu padat, sedang, rendah, dan tidak ditumbuhi lamun, berturut-turut untuk stasiun 1, 2, 3, dan 4. Kepadatan luasan tutupan padang lamun ditentukan menggunakan bingkai berukuran 50 x 50 cm2 (Kiswara & Djamali 1995).
Identifikasi Lamun
Identifikasi lamun dilakukan in situ. Idenitfikasi mengacu pada Seagrass of the World (Den hartog 1970). Ciri yang di-perhatikan adalah struktur anatomi rizoma, bentuk percabangan akar, pertulangan dan dentikulasi daun, bentuk ujung daun, dan ligula.
Pengambilan Contoh Sedimen
Contoh sedimen diambil dari empat stasiun. Pada tiap stasiun, pengambilan dilakukan sebanyak tiga kali. Sedimen diambil menggunakan core (p = 10 cm, d = 3 cm). Ketika core ditekan ke dalam sedimen diusahakan piston core sedekat mungkin dengan permukaan sedimen untuk mencegah pengadukan akibat tekanan dan tarikan core. Sedimen di dalam core dipotong menjadi lima bagian dengan ketebalan 2 cm (D1 = 1-2 cm, D2 = 3-4 cm, D3 = 5-6 cm, D4 = 7-8 cm, D5 = 9-10 cm). Selanjutnya, tiap bagian sedimen dimasukkan ke dalam botol plastik dan diawetkan dengan larutan formalin 4% dan di-warnai menggunakan rose bengal (Susetiono 1995).
Pengukuran Parameter Lingkungan
Pengukuran suhu air, pH sedimen, dan salinitas dengan refraktometer dilakukan in situ pada tiap stasiun. Hasil pengukuran di tulis di atas kertas anti air.
Ekstraksi Meiofauna
Tiap bagian sedimen dengan kedalaman tertentu dituang ke dalam gelas ukur berisi air tawar, lalu sedimen disaring dengan saringan berukuran 1 mm (Susetiono 1995), dengan tujuan memisahkan makrofauna. Sedimen yang lolos dari saringan 1 mm ditampung di dalam gelas ukur berisi air tawar. Air tawar yang digunakan sebelumnya disaring dengan saringan 0.032 mm untuk menghindari kontaminasi organisme air tawar. Sedimen di dalam gelas ukur, lalu didiamkan kira-kira 30 detik sampai pasir kasar tenggelam. Meiofauna dan pasir halus yang melayang-layang diatas per-mukaan air disaring dengan saringan 0.032 mm. Tahap ini diulang sebanyak tiga kali. Setelah itu, sedimen yang tertahan di saringan 0.032 mm di pindahkan ke cawan Petri (Pfannkuche & Thiel 1988).
Analisis Total Berat Organik (TBO)
Contoh sedimen diambil dari tiap stasiun menggunakan core (p = 30 cm, d = 5 cm). Setelah itu sedimen disimpan di dalam kantung plastik dan diberi label. Sedimen dianalisis menggunakan metode pengabuan yang dimulai dengan pemanasan selama 24 jam pada suhu 70o C. Setelah 24 jam, sedimen didinginkan lalu ditimbang dan didapat berat hasil pengeringan 70 oC (berat kering). Setelah ditimbang, sedimen dimasukkan ke dalam tanur untuk memasuki proses pengabuan selama 4 jam pada suhu 600o C. Setelah 4 jam, sedimen ditimbang. Selisih berat kering deng-an berat sedimen setelah pengabudeng-an me-rupakan jumlah TBO. TBO dinyatakan dalam persen.
Analisis Butiran Sedimen
Setelah itu ditimbang dan didapat berat kering. Berdasarkan ukurannya ada tiga ke-lompok sedimen, yaitu kerikil yang berukuran lebih besar dari 1 mm, pasir yang berukuran 0.063 sampai dengan 1 mm, dan lumpur yang berukuran lebih kecil dari 0.063 mm (Giere et. al 1988).
Identifikasi Meiofauna
Identifikasi meiofauna dilakukan sampai dengan tingkat taksa utama meiofauna. Identifikasi menggunakan mikroskop stereo, mikroskop cahaya, dan loop Irwin, dengan tujuan mengambil meiofauna dari dalam cawan Petri. Identifikasi menggunakan Intro-duction to the Study of Meiofauna (Higgins & Thiel 1988) dan Invertebrates (Brusca & Brusca 1990). Taksa dengan kepadatan ter-tinggi diidentifikasi sampai tingkat famili.
Analisis Data
Data biologi dianalisis menggunakan metode mulivariate, software PRIMER 5 (Plymouth Routines in Multivariate Eco-logical Research) (Deudero & Vincx 2000).
Kepadatan meiofauna ditulis dalam indi-vidu tiap 10 cm2. Jumlah meiofauna yang didapat dikonversikan ke luas permukaan 10 cm2 dengan cara mengkalikan individu dengan angka 1.4. Angka 1.4 didapat dengan membagi nilai 10 cm2 dengan luas core (Ø = 3 cm).
HASIL
Kepadatan Meiofauna Tiap Stasiun dengan Perbedaan Luasan Tutupan Vegetasi
Hasil penelitian ini didapatkan sebelas taksa meiofauna di P. Kambing, yaitu Nematoda, Copepoda, Polychaeta, Oligochae-ta, Foraminifera, Ostracoda, Nauplii, Isopoda, Acari, Kinorhyncha, dan Tardigrada (Lam-piran 2 & 3). Tujuh taksa pertama selalu ditemukan pada tiap stasiun (Tabel 1). Empat taksa lain, Isopoda, Acari, Kinorhyncha, dan Tardigrada, berturut-turut hanya ditemukan pada stasiun 1, 2, 3, dan 4 (Tabel 1). Nematoda adalah taksa yang mempunyai kepadatan tertinggi pada tiap stasiun (Tabel 1).
Kepadatan meiofauna umumnya menurun dari stasiun 1 ke 4 (Gambar 1). Kepadatan tertinggi didapat pada stasiun 1 (2205.71 tiap 10 cm2), yaitu stasiun dengan luasan tutupan lamun terpadat dan kepadatan terendah pada stasiun 4 (1210.52 tiap 10 cm2), yaitu stasiun tanpa ditumbuhi lamun (Gambar 1).
Distribusi Vertikal Meiofauna
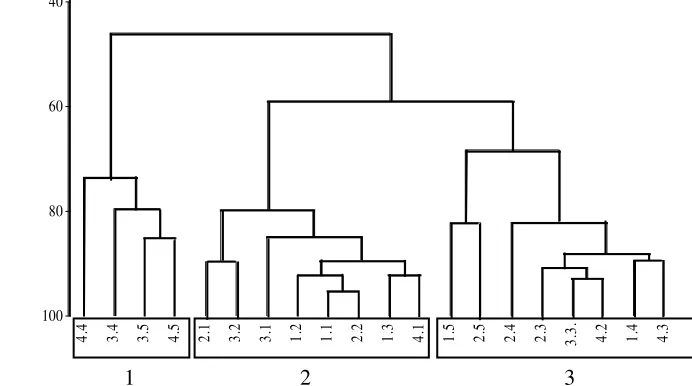
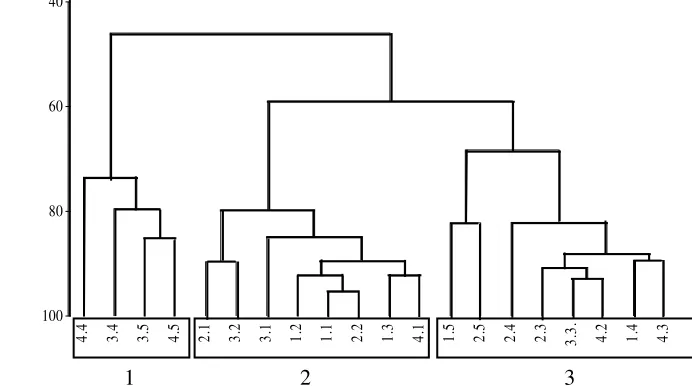
Hasil dari perhitungan nilai persamaan menggunakan persamaan Bray-Curtis ditam-pilkan dalam bentuk dendrogram (Gambar 2). Angka pertama pada ujung dendrogram me-nunjukkan jenis stasiun (1, 2, 3, dan 4). Empat stasiun tersebut berdasarkan kepadatan tutu-pan lamun yaitu padat, sedang, rendah, dan tidak ditumbuhi lamun, berturut-turut untuk stasiun 1, 2, 3, dan 4. Angka kedua menun-jukan kedalaman sedimen, yaitu D1 = 1-2 cm, D2 = 3-4 cm, D3 = 5-6 cm, D4 = 7-8 cm, D5 = 9-10 cm (1, 2, 3, 4, dan 5).
Kelompok 1 terdiri dari 4.4, 3.4, 3.5, dan 4.5 .(Gambar 2). Kepadatan meiofauna dalam kelompok ini berkisar antara 20-50 individu tiap 10 cm2 (Gambar 2). Kelompok 2 terdiri dari 2.1, 3.2, 3.1, 1.2, 1.1, 2.2, 1.3, dan 4.1. Kepadatan meiofauna dalam kelompok ini berkisar antara 300-700 individu tiap 10 cm2 (Gambar 2). Kelompok 3 terdiri dari 1.5, 2.5, 2.4, 2.3, 3.3, 4.2, 1.4, dan 4.3. Kepadatan meiofauna dalam kelompok ini berkisar antara 60-200 individu tiap 10 cm2 (Gambar 2).
Meiofauna ditemukan sampai dengan ke-dalaman 10 cm (distribusi vertikal) (Gambar 3). Kepadatan meiofauna mengalami pe-nurunan seiring dengan menurunnya keda-laman (Gambar 3). Kepadatan meiofauna ter-tinggi terdapat pada kedalaman 1-2 cm (D1) (Gambar 3).
Parameter Lingkungan
Annelida - Polychaeta - - 16.8 ± 3.2 18.2 ± 8.5 17.3 ± 6.5 8.4 ± 3.2
Oligochaeta - - 7.5 ± 3.6 10.7 ± 4.9 10.7 ± 5.2 8.9 ± 0.5
Arthropoda Crustacea Maxillopoda Copepoda - 560.0± 173.3 329.47± 73.0 549 ± 209.6 272.1 ± 46.2
Ostracoda - 1.4 ± 1.4 1.4 ± 0.8 0.9 ± 0.9 1.4 ± 1.40
Malacostraca - Isopoda 0.5 ±0.5 0 0 0
Larva: Nauplii 75.6 ± 30.9 48.5 ± 19.1 35.9 ± 17.8 25.7 ± 4.1
Cheliceriformes Chelicerata - Acari 0 0.5 ±0.5 0 0
Kinorhyncha - - - - 0 0 0.5 ± 0.5 0
Nematoda - - - - 1520.9 ± 378.0 937.5 ± 326.6 707.5 ± 112.4 874.1 ± 232.8
Sarcomastigophra Sarcodina Rhizopoda; Foraminifera - - 17.3 ± 7.5 6.5 ±2.0 14.0 ± 4.3 19.6 ± 3.7
Tardigrada - - - - 0 0 0 0.5 ± 0.5
0 500 1000 1500 2000 2500 3000
1 2 3 4
Stas iun
R
at
aa
n
k
ep
ad
at
an
m
ei
o
fa
u
n
a
(i
n
d
iv
id
u
t
ia
p
1
0
c
m
2 )
Gambar 1 Rataan kepadatan meiofauna tiap stasiun dengan perbedaan luasan tutupan E. acoroides, di P. Kambing.
Gambar 2 Dendrogram hasil persamaan Bray-Curtis untuk distibusi vertikal meiofauna tiap stasiun dengan perbedaan luasan tutupan E. acoroides, di P. Kambing. Angka pertama pada ujung cabang dendrogram menunjukan jenis stasiun (1, 2, 3, dan 4), sedangkan angka kedua menunjukan kedalaman sedimen (D1-D5) (1, 2, 3, 4, dan 5).
4
.4 3.4 3.5 4.5 2.1 3.2 3.1 1.2 1.1 2.2 1.3 4.1 1.5 2.5 2.4 2.3
3
.3
.
4
.2 1.4 4.3
100 80 60 40
S
im
il
ar
it
as
B
ra
y
-C
u
rt
is
1
2
3
0 500 1.000 1.500 2.000 2.500
1 2 3 4 5
Nematoda Copepoda Polychaeta Foraminifera Oligochaeta Ostracoda Nauplii Isopoda Acari Kinorhyncha Tardigrada
Gambar 3 Kepadatan taksa meiofauna pada distribusi vertikal. D1 = 1-2 cm, D2 = 3-4 cm, D3 = 5-6 cm, D4 = 7-8 cm, D5 = 9-10 cm.
P
er
sa
m
aa
n
B
ra
y
-C
u
rt
is
K
ed
al
am
an
Deskripsi Meiofauna di Pulau Kambing Nematoda
Nematoda mempunyai karakter tubuh triploblastik, bilateral, pseudoselomata, tidak bersegmen, tubuh ditutupi kutikula, pada kutikula terkadang dijumpai seta (Gambar 4), memiliki organ sensor dibagian kepala (amphid) (Gambar 5) dan di bagian ekor (phasmid), habitat di air laut, air tawar, da-ratan, beberapa hidup bebas dan beberapa parasit panjang Nematoda marin umumnya berkisar antara 1 sampai dengan 3 mm (Brus-ca & Brus(Brus-ca 1990).
Gambar 4 Nematoda.
Gambar 5 Amphid Nematoda.
Copepoda
Tubuh terbagi tiga tagmata, yaitu losome, metasome, dan urosome, cepha-losome terdiri dari kepala dan segmen pertama throrax (maxiliped). Kepala dileng-kapi dengan antenula (uniramus), antena (uni atau biramus), mandibula, maxilula (maksila pertama), maxila (maksila kedua). Thorax terdiri dari enam segmen, dilengkapi dengan lima pasang pereopod. Abdomen terdiri dari lima segmen, termasuk telson, kaudal rami berkembang dengan baik panjang berkisar antara 0.4-1 mm (Gambar 6) (Brusca & Brusca 1990; Wells 1988).
Tabel 3 Persentase komposisi sedimen tiap stasiun dengan perbedaan luasan tutupan E. acoroides di P. Kambing
Komposisi Sedimen (%) Diameter Sedimen
(mm) Jenis Sedimen Stasiun 1 Stasiun 2 Stasiun 3 Stasiun 4
> 1 Kerikil 13.46 7.19 21.89 24.46
0.063-1 Pasir 66.25 76.37 67.59 56.00
< 0.063 Lumpur 20.29 16.44 10.52 19.54
Seta Kutikula
100 µm
Tabel 2 Nilai hasil pengukuran parameter lingkungan tiap stasiun dengan perbedaan luasan tutupan E. acoroides di P. Kambing
Parameter lingkungan Stasiun 1 Stasiun 2 Stasiun 3 Stasiun 4
Luasan tutupan lamun (%) 60 45 10 0
TBO (%) 28.27 24.68 21.64 25.41
Salinitas (‰) 30 30 30 30
Suhu air (OC) 32 33 32 33
pH sedimen 7 7 7 7
Amphid
Gambar 6 Copepoda
Nauplii
Nauplii adalah larva dari subsubfilum Crustacea. Tiap perkembangannya terdiri dari 5 sampai dengan 6 tahap. Tiap tahapan per-kembangan dibedakan dari penambahan pada segmen prosome, yaitu segmen gabungan cephalosome dan metasome, dan urosome. Serta perkembangan alat-alat tambahan, panjang berkisar antara 0.1-0.5 mm (Gambar 7) (Wells 1988).
Gambar 7 Nauplii.
Polychaeta
Tubuh lunak, tubuh terbagi-bagi menjadi segmen berukuran sama, segmen tubuh mempunyai banyak seta, parapodia ber-kembang dengan baik, prostomium dan peristomiun tanpa klitelum, panjang berkisar antara 1 sampai dengan 3 m (Brusca & Brusca 1990) (Gambar 8).
Gambar 8 Polychaeta.
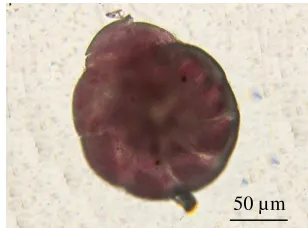
Foraminefera
Cangkang tersusun dari kalsium karbonat, cangkang terdiri dari kamar-kamar, tidak mempunyai organel lokomotor yang perma-nen, ukuran berkisar antara 0.1-0.6 mm (Gambar 9) (Brusca & Brusca 1990).
Gambar 9 Foraminifera.
Oligocaheta
Tubuh bersegmen, seta beberapa, tidak ada parapodia, mempunyai klitelum, panjang berkisar antara 1 mm-1.5 mm (Gambar 10).
Gambar 10 Oligochaeta.
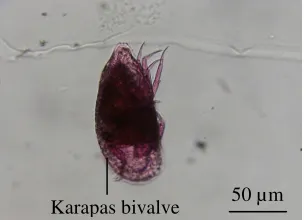
Ostracoda
Segmen tubuh tereduksi, karapas tangkup dua (bivalve), karapas menutupi badan dan kepala, mempunyai antenula, antena, man-dibula, maksilula, maksila, mempunyai kaudal rami, panjang berkisar antara 0.1-0.5 mm (Gambar 11).
Gambar 11 Ostracoda.
Karapas bivalve 50 µm 50 µm
Seta
Klitelum 100 µm
Parapodia
Seta 200 µm
50 µm Antena
Metosome
Cephalosome
Pereopod Kaudal rami
Isopoda
Tidak mempunyai karapas, mempunyai dua pasang antena, mempunyai tujuh pasang pereopod, badan datar dorsoventral. Ekor terdiri dari telson dan uropod, telson ber-gabung bengan 1-6 pleonit membentuk pleo-telson, tidak mempunyai kaudal rami, panjang berkisar antara 1-1.5 mm (Gambar 12).
Gambar 12 Isopoda.
Acari
Termasuk kedalam Kelas Chelicerata, Ordo Acari. Tubuh terbagi menjadi gnatho-soma (anterior) dan idiognatho-soma (posterior), gna-thosoma mungkin hampir sama panjang dengan idiosoma, tapi biasanya lebih pendek, mempunyai empat pasang kaki, dua pasang menghadap ke depan, dua pasang menghadap ke belakang, panjang berkisar antara 0.1-0.5 mm (Gambar 13).
Gambar 13 Acari.
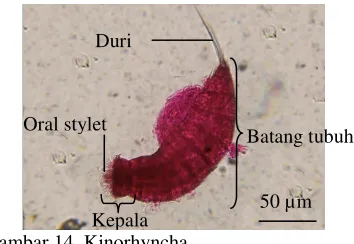
Kinorhyncha
Simetri bilateral, tubuh terbagi menjadi kepala dan batang tubuh, segmen tubuh jelas, mempunyai kerucut oral (oral cone) pada kepala dan dikelilingi oleh duri yang tersusun melingkar (oral stylet), mempunyai duri pada ujung, panjang berkisar antara 0.1-0.5 mm (Gambar 14).
Gambar 14 Kinorhyncha.
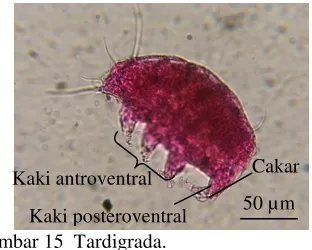
Tardigrada
Simetri bilateral, tubuh dibagi ke dalam lima bagian yaitu kepala, tiga segmen yang tiap segmennya dilengkapi dengan sepasang kaki anteroventral, dan satu segmen terakhir dengan sepasang kaki posteroventral, tiap kaki mempunyai cakar atau kuku, panjang berkisar antara 0.1-0.5 mm (Gambar 15).
Gambar 15 Tardigrada.
Famili Nematoda di P. Kambing
Nematoda adalah taksa yang mendominasi di tiap stasiun dan di tiap distribusi vertikal. Pada penelitian ini ditemukan 33 famili Nematoda (Lampiran 4). Lima famili terbesar adalah Leptosomatidae, Linhomoeidae, Mon-hysteridae, Oxystominidae, dan Pelago-nematidae. Nematoda diidentifikasi berda-sarkan permukaan kutikula, bentuk amphid, buccal cavity dan alat tambahan (Platt & Warwick 1988).
Klasifikasi dan Deskripsi Lima Famili Nematoda dengan Jumlah Tertinggi di P. Kambing
Filum: Nematoda: Kelas: Adenophorea: Ordo: Enoplida
Amphid samar. Integumen halus atau beranulasi (berbentuk seperti cincin).
Duri
Oral stylet
50 µm Batang tubuh
Kepala
Kaki antroventral Cakar Kaki posteroventral 50 µm Antena Pereopod 200 µm
Telson & uropod
Gnathosoma
Idiosoma
Esofagus silinder, jarang membentuk bulb yang membengkak. Bentuk buccal cavity bervariasi. Terdapat atau tidak terdapat seta pada kepala. Seta pada badan umumnya hadir. Terdapat atau tidak terdapat kelenjar kaudal (Chitwood 1933).
Subordo: Enoplina
Kutikula halus. Terdapat penebalan pada bagian kepala, membentuk struktur seperti helm atau topi. Seta pada kepala berkembang dengan baik. Amphid berukuran sedang. Buccal cavity kompleks, dengan satu atau tiga gigi berukuran besar, jarang simpel. Bagian anterior esofagus mencapai bagian depan stoma. Organ tambahan berbentuk tubulifrom (Chitwood 1933).
Superfamili: Leptosomatoidea
Stoma simpel. Gigi, jika hadir, umumnya berukuran tidak sama. Esofagus muskular (Filipjev 1916).
Famili: Leptosomatidae
Kapsul pada kepala samar. Stoma sempit. Gigi berukuran kecil (Filipjev 1916).
Subordo: Oncholaimina
Kutikula halus. Seta kepala pendek. Stoma selalu besar, berbentuk seperti laras senapan. Mempunyai tiga gigi berukuran besar. Esofagus dimulai dari dasar stoma. Organ tambahan berbentuk papil atau tombol (de Coninck 1965; & Blaxter 2000).
Famili: Pelagonematoidae
Stoma tidak bergigi atau gigi berukuran kecil, esofagus silinder (de Coninck 1965, diacu dalam Ley & Blaxter 2000).
Subordo: Tripylina
Kutikula halus, jarang beranulasi. Seta pada kepala berukuran kecil atau berbentuk papil. Amphid berukuran kecil. Buccal cavity simpel, berbentuk corong, sempit. Esofagus silinder (Andrassy 1974; Ley & Blaxter 2000).
Superfamili: Oxystomonoidea
Stoma hampir absen. Kepala sempit dan membulat. Seta kepala kecil, sering berbentuk papil. Esofagus melebar secara bertahap. Badan sangat ramping. Organ tambahan berbentuk papil, tombol, atau seta (Filipjev 1918).
Famili: Oxystominidae
Amphid sangat besar dan jika kecil bentuknya beragam. Organ tambahan sangat kecil (Filipjev 1918).
Filum: Nematoda: Kelas: Adenophorea: Ordo: Monhysterida
Seta pada kepala berjumlah 4, 4+6, atau lebih. Kutikula halus atau anulasi, tanpa punctuation. Amphid berbentuk sirkular atau subspriral, kecuali spiral. Buccal cavity umumnya kecil dan tidak terlindungi, jarang luas dan bergigi. Esofagus silinder, jarang mempunyai bulb yang membesar (de coninck & Chuurmans-Stekhoven 1933).
Famili: Monhysteridae
Kutikula halus. Bagian terminal kaudal jarang berseta (de Man 1876; Ley & Blaxter 2000).
Famili Linhomoeidae
Esofagus biasanya dengan bulb (Filipjev 1922).
Identifikasi Lamun
Spesies lamun yang ditemukan di Pulau Kambing adalah Enhalus acoroides. E acoroides mempunyai ciri-ciri rizoma merambat dan berdimeter lebih dari 1 cm. Akar seperti tali, tidak bercabang, panjang 10-20 cm dan tebal 3-5 mm. Tanpa ligula, daun tebal dan kaku, panjang 30-150 cm, lebar 1.5-2 cm, ujung daun membulat atau tumpul, tepi daun tebal dan licin (Lampiran 6).
PEMBAHASAN
Kepadatan dan Keragaman Meiofauna di Tiap Stasiun
Kino-rhyncha, Acari, Amphipoda, Nauplii, Isopoda, dan Turbelaria.
Nematoda adalah taksa yang paling umum ditemukan di tiap stasiun (Tabel 1). Penelitian sebelumnya melaporkan bahwa Nematoda adalah taksa yang paling umum ditemukan pada padang lamun di Krabi, Thailand (Monthum & Aryuthaka 2006). Hasil ini mungkin karena Nematoda mempunyai kelenjar kaudal. Kelenjar kaudal mem-produksi bahan perekat yang dikeluarkan melalui struktur khusus di ujung ekor atau spineret, sehingga Nematoda dapat me-rekatkan diri pada butir sedimen (Platt & War-wick 1988).
Keragaman meiofauna interstisiil di laut dalam umumnya lebih tinggi dibandingkan, meiofauna interstisiil di laut dangkal. Penyebab keragaman ini karena beberapa faktor di laut dalam seperti predasi atau gangguan. Faktor-faktor tersebut menekan terjadi dominansi (Coull 1988).
Empat taksa, yaitu Isopoda, Kinorhyncha, Acari, dan Tardigrada berturut-turut hanya ditemukan satu individu pada stasiun 1, 2, 3, dan 4 (Tabel 1). Acari ditemukan pada stasiun 2 dengan kepadatan 0.5 individu tiap 10 cm2 0.5 individu tiap 10 cm2 (Lampiran 3). Stasiun 3 mempunyai kandungan kerikil dan pasir yang tinggi (Tabel 3). Giere (1993) me-laporkan umumnya di dalam sedimen, Kinorhycha hanya ditemukan 15 individu tiap 10 cm2. Kinorhyncha umumnya ditemukan di sedimen berpasir atau kerikil pada lapisan atas sedimen (0-3 cm) (Giere 1993). Tardigrada ditemukan pada stasiun 4 (Tabel 1). Stasiun 4 mempunyai kandungan kerikil tertinggi (Tabel 3). Tardigrada mempunyai cakar di tiap kaki yang berfungsi untuk mencengkram butir sedimen (Brusca & Brusca 1990; Giere 1993).
Seluruh stasiun pengamatan mempunyai nilai parameter lingkungan yang tidak berbeda (Tabel 2). Nilai parameter lingkungan tidak mempengaruhi kepadatan meiofauna (Tabel 1 dan 2). Hasil ini sesuai dengan penelitian sebelumnya mengenai kepadatan meiofauna di daerah bakau, yaitu hubungan antara kepadatan meiofauna dengan suhu tidak berbeda nyata (Olafsson 1995). Tetapi, untuk beberapa taksa, suhu mempengaruhi kepadatan meiofauna. Suhu dan persediaan makanan adalah faktor utama yang
menentukan perkembangan Copepoda (Giere 1993). Copepoda merupakan anggota dari subsubfilum Crustacea yang mempunyai tahap Nauplii dalam perkembangannya (Wells 1988). Perkembangan Copepoda dengan enam tahap Nauplii dan lima tahap copepodite dipengaruhi oleh suhu tahunan (Giere 1993). Nauplii yang ditemukan pada penelitian ini menandakan pada stasiun pengematan siklus reproduksi Crustacea masih dapat berlangsung (Tabel 1). Meiofauna dari subfilum Crustacea umumnya lebih cepat terpengaruh oleh polusi dibandingkan Nematoda karena anggota subfilum Crustacea, tidak toleran terhadap lingkungan dengan kandungan oksigen rendah (Coull 1988; Susetiono 1999). Pada Nematoda, jumlah produksi telur dipengaruhi oleh suhu (Platt & Warwick 1988).
Kepadatan Meiofauna Tiap Stasiun dengan Perbedaan Luasan Tutupan E. acoroides
Lamun di P. Kambing, didominasi oleh E. acoroides, Famili: Hydrocharitaceae, Genus: Enhalus (Den Hartog 1970). Pada penelitian ini kepadatan meiofauna tertinggi diperoleh pada stasiun 1, 2, 3, dan 4 berturut-turut untuk stasiun luasan tutupan padang lamun padat, sedang, rendah, dan daerah tanpa ditumbuhi lamun (Gambar 1). Hasil ini memperkuat penelitian sebelumnya yang dilakukan di padang lamun Krabi, Thailand Monthum & Aryuthaka (2006), yaitu kepadatan meiofauna di daerah yang bervegetasi lebih tinggi dibandingkan daerah tanpa vegetasi. Per-tumbuhan daun dan sistem perakaran yang padat, dapat memperlambat gerakan air yang disebabkan oleh arus dan ombak serta menyebabkan perairan disekitarnya tenang (Azkab 2006).
Distribusi Vertikal Meiofauna Tiap Stasiun
Kepadatan lamun dan kedalaman sedimen dapat mempengaruhi distribusi vertikal meio-fauna (Gambar 2). Tetapi dalam distribusi vertikal, kedalaman sedimen lebih mem-pengaruhi kepadatan meiofauna dibandingkan kepadatan lamun (Gambar 2). Distribusi vertikal dikontrol oleh redox potential discontinuity (RPD) yaitu batas antara sedimen aerobik dan anaerobik. RPD berhubungan dengan kandungan oksigen (Coull 1988).
Kelompok 1 mempunyai kepadatan meiofauna terendah berkisar antara 20-50 individu tiap 10 cm2 (Gambar 2). Kelompok 1 beranggotakan stasiun 3 dan 4 dan tiap stasiun mempunyai kedalaman sedimen antara 7-10 cm (D4-D5). Kelompok 2 mempunyai kepadatan meiofauna tertinggi, berkisar antara 300-700 individu tiap 10 cm2 (Gambar 2). Kelompok 2 umumnya beranggotakan stasiun 1 sampai dengan stasiun 3, kecuali satu stasiun 4. Tetapi, tiap stasiun umumnya dengan kedalaman sedimen 1-4 cm (D1-D2), kecuali 1.3 yaitu stasiun 3, kedalaman sedimen 5-6 cm (D3) (Gambar 2). Meiofauna banyak ditemui pada kedalaman sedimen 1-6 cm (D1-D3) (Gambar 2) karena pada kedalaman 2-4 cm sedimen relatif stabil dan teroksigenisasi dengan baik (Susetiono 1995)
Meiofauna ditemukan sampai dengan kedalaman sedimen 10 cm (distribusi vertikal) (Gambar 3). Komposisi sedimen pada tiap stasiun di P. Kambing didominasi oleh pasir (Tabel 3). Distibusi vertikal meiofauna bergantung atas komposisi sedimen, terutama pada pantai yang tidak terpolusi (Perkins 1974; Henning et al. 1982). Sebagian besar meiofauna menyukai hidup di sedimen berdiameter sedang (0.125-0.5 mm) karena sedimen tersebut teroksigenisasi dengan baik (Henning et al. 1982). Pada habitat dengan sedimen berlumpur, meiofauna dapat ditemukan pada lapisan lebih dalam (Coull 1988). Pada habitat berlumpur meiofauna sebagian besar ditemukan di lapisan atas sedimen yang teroksigenisasi (Coull 1988).
Kepadatan meiofauna dalam distribusi vertikal, umumnya menurun dengan me-ningkatnya kedalaman (Gambar 3). Hasil penelitian ini memperkuat simpulan Susetiono (1996) bahwa semakin dalam lapisan sedimen, kepadatan meiofauna semakin rendah.
Nematoda adalah taksa paling umum ditemukan dalam distribusi vertikal dan di setiap kedalaman pada tiap stasiun pengamatan (Gambar 3). Hasil ini disebabkan
karena Nematoda sangat toleran terhadap lingkungan yang rendah oksigen Susetiono (1995). Oleh karena itu, Nematoda mempunyai kesempatan lebih besar untuk bermigrasi ke bagian sedimen yang lebih dalam dibandingkan taksa meiofauna lain (Susetiono 1999).
Copepoda adalah taksa kedua yang paling umum dalam distribusi vertikal. Kepadatan Copepoda menurun drastis pada kedalam 7-10 cm (Gambar 3). Copepoda banyak ditemukan pada lapisan atas sedimen karena Copepoda tidak toleran terhadap lingkungan yang anaerobik atau miskin oksigen (Susetiono 1999; Perkins 1974).
Distibusi vertikal dapat dipengaruhi oleh desikasi (pengeringan). Ketika surut, pasir menjadi kering. Oleh karena itu, meiofauna akan bermigrasi ke lapisan bawah ketika surut dan ke atas ketika pasang (Coull 1988). Cahaya juga dapat mempengaruhi distribusi vertikal pada Nematoda yang mungkin berhubungan dengan migrasi diatom, yang merupakan makanan Nematoda (Castel et al. 1989).
Nematoda Tiap Stasiun dengan Perbedaan Karakteristik Sedimen
Pada stasiun 1, 2, 3, dan 4 berturut-turut ditemukan 18, 22, 22, dan 26 famili Nematoda (Lampiran 4). Stasiun 4 mempunyai kera-gaman tertinggi (Lampiran 4). Kerakera-gaman Nematoda dipengaruhi oleh komposisi sedi-men. Stasiun 4 mempunyai kandungan kerikil tertinggi (Tabel 3). Hasil ini sesuai dengan penelitian sebelumnya bahwa keragaman Nematoda tinggi pada sedimen kasar (Gour-bault et al. 1995; Steyaert et al. 1999).
Termasuk tipe makan herbivor (Perkins 1974). Tipe 2B adalah buccal cavity yang mempunyai pelindung yang besar dan kuat. Sebagian besar predator. Mangsa ditelan secara utuh. Termasuk tipe makan karnivora atau omnivora (Perkins 1974).
Nematoda yang mempunyai tipe makan 1B banyak pada stasiun 1 dan 2. Tipe Stasiun 1 dan 2 adalah stasiun dengan kandungan kerikil rendah (Tabel 3), sedangkan kan-dungan pasir dan lumpur tinggi (Tabel 3). Steyaert et al. (1999) melaporkan bahwa semakin halus kandungan sedimen, jumlah Nematoda dengan tipe makan deposit tidak selektif semakin (1B) meningkat. Hasil ini mungkin disebabkan bahan organik yang berasosiasi dengan butiran sedimen halus (Steyaert et al. 1999). Nematoda dengan tipe makan 2B atau karnivora banyak terdapat pada stasiun 3 yaitu daerah dengan penutupan rendah dan 4 yaitu daerah tanpa penutupan lamun. Hasil ini sesuai dengan laporan (Gourbault et al. 1998) bahwa jumlah Nematoda karnivora meningkat pada pantai yang terbuka dan mempunyai kandungan organik rendah.
Lima famili Nematoda terbesar yang ditemukan di Pulau Kambing telah dilaporkan di beberapa habitat diseluruh dunia. Famili Monhysteridae, Oxystominidae, dan Lin-homoeidae telah dilaporkan pada sedimen di laut Weddell, Antartika (Vanhove et al. 1999) dan di Laut Baltic pada kedalaman 37 m (Olafsson & Elmgren 1997).
Deontostoma timmerchioi dari famili Leptosomatidae, ditemukan di daerah Antar-tika di ruang interspikular spons (Hope 1974). Genus Odontobius dari famili Monhysteridae, ditemukan hidup pada balin dari paus balin (Lorenzen 1986). Genus Sphaerotheristus dari famili Monhysteridae, dilaporkan terdapat di timur Pakistan (Timm 1968). Famili Lin-homoeidae telah dilaporkan di Pulau Socorro, Colina Mexico (de Jesus 2007). Genus Thalassogenus dari famili Pelagonematidae, mempunyai distribusi cukup luas yaitu di India, Papua Niugini, Pulau Samoa dan Amerika tengah (Loof & Zullini 2000).
Pada penelitian ini tipe makant yang paling umum ditemukan berturut-turut adalah 1B, 2A, 1A, dan 2B (Lampiran 5). Hasil ini ber-beda dengan laporan Susetino (1999) pada padang lamun E. Acoroides di Pulau Lombok yaitu di dalam sedimen tipe makan tipe makan yang paling umum berturut-turut adalah 1A, 2A, 1B, dan 2B.
Peran Meiofauna di Komunitas Bentik
Meiofauna di dalam komunitas bentik berperan sebagai sumber makanan untuk sesama meiofauna, biodegradasi bahan organik, makanan untuk tingkat tropik yang lebih tinggi, dan bioindikator lingkungan (Coull 1988). Meiofauna, terutama Copepoda, adalah sumber makanan utama untuk beraneka ragam predator, terutama juvenil ikan (Coull 1990). Crustacea, ikan-ikan kecil, dan Polychaeta juga biasa memangsa Nematoda (Giere 1993).
Sebagai bioindikator, kelimpahan dan keragaman taksa utama meiofauna sering digunakan sebagai acuan (Susetiono 2000). Meiofauna sangat berguna untuk penelitian lingkungan karena jumlahnya yang banyak, tidak berpindah-pindah habitat, waktu generasi pendek, dan tidak adanya tingkat larva plantonik (Coull 1988). Oleh karena itu, jika terjadi polusi meiofauna tidak bisa migrasi dan menghindar, sehingga mereka harus beradaptasi atau mati (Gray 1982).
Rasio Nematoda / Copepoda (N / C) dapat dijadikan bioindikator polusi pada komunitas bentik (Susetiono 2000). Penggunaan Nematoda dan Copepoda dalam rasio mungkin karena kedua taksa tersebut umum mendominasi di sedimen. Monthum & Aryuthaka (2006) dan Ansari & Parulekar (1994) melaporkan Nematoda dan Copepoda adalah dua taksa yang umum ditemukan di sedimen.
Pada penelitian ini, rasio N / C lebih dari 1:1 (Tabel 1). Hasil ini mengindikasikan mungkin pernah terjadi gangguan di stasiun pengamatan sebagai akibat dari aktifitas industri sekitar pantai. Pantai yang tidak terpolusi, rasio N / C mendekati satu (Henning et al. 1982).
SIMPULAN
terbesar adalah Leptosomatidae, Linho-moeidae, Monhysteridae, Oxystominidae, dan Pelagonematoidae.
DAFTAR PUSTAKA
Andrassy I, 1974. A nematodák evolúcioja és rendszerezése. Magyar Tudomanyos Aka-demia Biologiai Tudomanyok Osztályanak Közlemenyei 17: 13-58.
Ansari ZA, Parulekar AH. 1994. Meiobenthos in the sediment of seagrass meadows of Lakshadweep, Arabian Sea. Vie Milieu 44:185-190
Armonies W. 1988. Active emergence of meiofauna from intertidal sediment. Mar Ecol Prog Ser 43:151-159
Azkab MH. 2006. Ada apa dengan lamun. Oseana XXXI:45-55.
Bouwman LA. 1987. Meiofauna. Di dalam: Baker JM, Wolff WJ, (ed). Biological Survey of Estuaries and Coast. Cam-bridge: Cambridege Univ Pr. hlm 140-156. Brusca RC, Brusca GJ. 1990. Invertebrates Sinauer Associates, Massachusetts: Inc. Sunderland.
Castel J, Labourg PJ, Escaravage V, Auby I, Garcia M.E. 1989. Influence of seagrass beds and oyster parks on the abundace and biomass patterns of meio- and macro-benthos on tidal flats. Estuar Coast Shelf Sci 28:71-85.
Chitwood BG. 1933. A revised classification of the Nematoda. J Parasitol 20:131. Coull BC. 1988. Ecology of marine
meiofauna. Di dalam: Higgins RP, Thiel H, (ed). Introduction to the Study of Meiofauna. London: Smithsonian Ins-titution Pr. hlm 18-23.
Coull BC. 1990. Are members of the meiofauna food for higher trophic levels. Transac Am Micros Soc. 109:233-246. de Coninck LAP. 1965. Systematique des
nematodes. Di dalam: Grasse PP, (ed). Traite de zoologie, anatomie, syste-matique, biologie. Paris: Masson. hlm 586-731
de Coninck LAP, and Chuurmans-Stekhoven JH Jr. 1933. The freeliving marine nemas of the Belgian coast. II: with general remarks on the structure and the systems of nemas. Mém Mus R Hist Nat Belg 58:1-163.
de Jesus NA. 2007. Littoral free living nematode fauna of Socorro Island, Colima, Mexico. Hidrobiologica 17:61-66.
de Man JG. 1876. Onderzoekingen over vrij in de aarde levende nematoden. Tijdschrift der Nederlandsche Dierkundige Veree-niging 2: 78-196
Den Hartog C. 1970. The Seagrass of the World. Amsterdam: North Holland Publi. Deudero S, Vincx M. 2000. Sublittoral
meiobenthic assemblages from disturbed and non-disturbed sediment in the Balearics. Sci Mar 64:285-293.
Edgar GJ. 1999. Experimental analysis of structural versus trophic importance of seagrass beds: effects on macrofaunal and meiofaunal invertebrates. Vie et milieu 49:239-248.
Filipjev IN. 1916. Les Nématodes libres contenus dans les collections du Museé zoologique de l’Académie impérale des Sciences de Petrograd. Ann Mus Zool Acad Sci 21: 59-116.
Filipjev IN. 1918. Free-living marine Nematodes of the Sevastopol area (in Russian). Trudy Osob Zool Lab Sevastop. Biol Sta 4: 1-350.
Filipjev IN. 1922. Encore sur les Nématodes libres dela Mer noire. Tr Stavrop S-kh Inst 1: 84-184.
Fortes MD. 1990. Seagrass; A Resource Unknown in The ASEAN Region. Manila: International center for living aquatic resources management.
Giere O, Eleftheriou A, Murison DJ. 1988. Abiotic Factor. Di dalam: Higgins R.P, Thiel H, (ed). Introduction to the Study of Meiofauna. London: Smithsonian Institu-tion Pr. hlm 61-78.
Giere O. 1993. Meiobenthology: The Micros-copic Fauna in Aquatic Sedimens. Berlin: Spinger-Verlag
Gourbault A, Warwick RM, Helleouet MN. 1995. A survey of intertidal meiobenthos (especially Nematoda) in coral sandy beaches pf Moorea (French Polynesia). Bull Mar Sci 57:476-488.
Gourbault A, Warwick R.M, Helleouet MN. 1998. Spatial and temporal variability in the composition and structure of meio-benthic assemblages (especially nema-todes) in tropical beaches (Guadeloupe, FWI). Biol Mar 39:29-39.
Gray JS. 1982. Effects pollutants on marine ecosystems. Neth J Sea Res 16: 424-443. Hemminga A, Duarte CM. 2000. Seagrass
Ecology. Cambridge: Cambridge Univ Pr. Henning HF, Fricke KO, Greenwood PJ,
sandy beaches. Envir Monitoring Assess 1:337-344.
Higgins RP, Thiel H. 1988. Prospectus. Di dalam: Higgins R.P, Thiel H, (ed). Introduction to the Study of Meiofauna. London: Smithsonian Institution Pr. hlm 11-13.
Hope WD. 1974. Deontostoma timmerchioi n. sp., a New Marine Nematode (Leptoso-matidae) from Antarctica, with a Note on the structure and Possible Function of the ventromedian supplement. Transac Am Micros Soc 93: 314-324.
Kiswara W, Djamali A. 1995. Seagrass community and associated biota in Banten West Java- Indonesia: Problem and research priorities. First workshop on marine and coastal research; Jakarta 1-4 Mei 1995. Netherlands: Netherlands Oraganization for Scientific Research-now. hlm 92-107.
Kiswara W. 1992. Community structure and biomass distribution of seagrasses at Banten Bay, West Java-Indonesia. Di dalam: Chou L.M dan C.R. Wilkinson, (ed). Marine Science: Living Coastal Resources. Proceedings of Third ASEAN Science and Technology Week Confe-rence; Singapore, 21-23 September 19-92. Singapore: National Univ of Singa-pore. hlm 241-250.
Kiswara W. 2004. Kondisi Padang Lamun (Seagrass) di Perairan Teluk Banten 1998-2001. Jakarta: Lembaga Penelitian Oseanografi.
Ley PD, Blaxter M. 2000. Systematic position and phylogeny . Di dalam: Lee DL, (ed). The Biology of Nematodes. New York: Taylor & Francis
Loof PA, Zullini A. 2000. A new species of the genus Thalassogenus Andrássy, 1973 and the systematic position of the genus (Nematoda: Oncholaimina). Ann Zool 50:263-266.
Lorenzen S. 1986. Odontobius (Nematoda, Monhysteridae) from the baleen plates of whales and its relationship to Gamma-rinema living on crustaceans. Zool Scrip 15:101-106
Monthum Y, Aryuthaka C. 2006. Spatial distributin of meiobenthic community in Tha Len seagrass bed, Krabi Province, Thailand. Coast Mar Sci 30:146-153. Olafsson E, Elmgren R. 1997. Seasonal
dynamics of sublittoral meiobenthos in relation to phytoplankton sedimentation in the baltic sea. Estue Coast Shelf Scei 45:149-164.
Perkins EJ. 1974. The Biology of Estuaries and Coastal Waters. New York: Academic Pr.
Pfannkuche O, Thiel H. 1988. Sample Processing. Di dalam: Higgins R.P, Thiel H, (ed). Introduction to the Study of Meiofauna. London: Smithsonian Institu-tion Pr. hlm 134-145.
Platt HM, Warwick R.M. 1988. Free Living Marine Nematodes. London: The Linnean of Society.
Susetiono. 1995. Struktur komunitas biologi padang lamun di pantai Selatan Lombok dan kondisi lingkungannya. Jakarta: Pusat Penelitian dan Pengembangan Oseanologi. Lembaga Ilmu Pengetahuan Indonesia. hlm 79-95.
Susetiono. 1996. Meiofauna of seagrass bed in Kuta Bay, Lombok, Indonesia. National Olympics Memorial Youth Center. Proceedings of Seventh Joint Seminar on dinamycs at monotypic seagrass Enhalus acoroides Kuta Bay, Lombok Island. Di dalam: Saadon M.N, Abdullah S.A, Sheriff S.MD, Afirrin N.A, (ed). Proceedings of The Tenth JSPS Joint Seminar on Marine and Fishery Sciences; Malaysia, 29 November-1 Desember 1999. Malaysia: Kolej Universiti Tengganu. hlm 100-117.
Susetiono . 2000. The use of meiofauna for environmental monitoring tool. Prosiding Seminar Kelautan; Jakarta 29-30 Maret 2000. Jakarta: Peranan Ilmu Alamiah Da-lam Eksplorasi Eksploitasi, dan Pelestarian Sumber Daya Laut. hlm 255-262.
Steyaert M, Garner N, Dirk VG, Vincx M. 1999. Nematode communities from the North Sea: environmental controls on species diversity and vertical distribution within the sediment. J Mar Biol Ass UK 79:253-264.
Lampiran 1 Peta Teluk Banten dan P. Kambing
Lampiran 2 Kepadatan dan keragaman meiofauna di tiap stasiun dan kedalaman di P. Kambing, Teluk Banten (Individu)
Lokasi Ne Co Poly For Oli Os Na Iso
S1U1D1 455 199 5 3 0 0 32 0
S1U1D2 183 21 2 3 0 0 2 0
S1U1D3 56 14 0 8 0 0 2 0
S1U1D4 43 14 1 1 1 0 0 0
S1U1D5 47 5 0 0 0 0 0 0
S1U2D1 364 148 1 10 0 2 50 1
S1U2D2 379 291 11 8 0 1 30 0
S1U2D3 707 175 2 0 6 0 16 0
S1U2D4 119 27 0 0 4 0 2 0
S1U2D5 56 5 2 2 0 0 0 0
S1U3D1 158 40 1 0 2 0 1 0
S1U3D2 298 212 3 0 2 0 14 0
S1U3D3 281 32 8 0 1 0 10 0
S1U3D4 92 12 0 2 0 0 3 0
S1U3D5 21 5 0 0 0 0 0 0
Lokasi Ne Co Poly For Oli Os Na Aca
S2U1D1 132 86 8 0 0 0 7 0
S2U1D2 641 157 8 4 1 0 37 0
S2U1D3 110 23 5 1 0 0 6 0
S2U1D4 80 13 4 0 1 0 2 0
S2U1D5 28 7 0 0 0 0 1 0
S2U2D1 347 107 4 0 2 0 2 0
S2U2D2 248 129 5 2 0 1 28 0
S2U2D3 87 38 0 0 2 1 5 0
S2U2D4 59 6 0 0 1 0 1 0
S2U2D5 61 9 0 0 2 0 7 0
S2U3D1 50 49 4 0 6 0 3 0
S2U3D2 71 44 0 3 3 1 7 1
S2U3D3 35 27 1 0 4 0 1 0
S2U3D4 25 4 0 1 0 0 0 0
S2U3D5 35 7 0 3 1 0 2 0
Lokasi Ne Co Poly For Oli Os Na Kinor
S3U1D1 255 212 15 1 7 0 23 1
S3U1D2 91 52 2 1 5 0 3 0
S3U1D3 56 10 0 2 2 0 1 0
S3U1D4 23 2 0 0 0 0 0 0
S3U1D5 15 2 0 4 1 0 0 0
S3U2D1 266 505 12 0 1 0 16 0
S3U2D2 80 126 11 0 1 1 22 0
S3U2D3 51 48 4 3 0 1 6 0
S3U2D4 20 6 0 9 1 0 3 0
S3U2D5 12 4 0 4 0 0 0 0
S3U3D1 327 150 2 0 2 0 2 0
S3U3D2 212 27 1 1 3 0 1 0
S3U3D3 97 8 0 5 0 0 0 0
Lampiran 2 (Lanjutan)
Lokasi Ne Co Poly For Oli Os Na Kinor
S3U3D5 6 2 0 0 0 0 0 0
Lokasi Ne Co Poly For Oli Os Na Tar
S4U1D1 206 138 8 1 4 0 12 0
S4U1D2 82 12 1 2 2 1 2 0
S4U1D3 20 79 1 0 0 0 0 0
S4U1D4 18 0 0 2 0 1 0 0
S4U1D5 13 6 0 5 0 1 0 0
S4U2D1 328 147 4 0 2 0 13 0
S4U2D2 141 33 2 4 0 0 6 0
S4U2D3 100 35 0 2 1 0 5 0
S4U2D4 42 1 0 5 3 0 0 0
S4U2D5 8 3 0 2 0 0 0 1
S4U3D1 695 83 1 5 4 0 16 0
S4U3D2 100 30 1 0 1 0 0 0
S4U3D3 88 14 0 0 1 0 1 0
S4U3D4 23 1 0 11 1 0 0 0
S4U3D5 9 1 0 3 0 0 0 0
Keterangan: Ne = Nematoda, Co = Copepoda, Poly = Polychaeta, For = Foraminifera, Oli = Oligochaeta, Os = Ostracoda, Na= Nauplii, Iso = Isopoda, Aca = Acari, Kinor = Kinorhyncha, Tar = Tardigrada, S = stasiun, U = ulangan, D = kedalaman (DD11==11--22 c
Lampiran 3 Kepadatan dan keragaman meiofauna di tiap stasiun dan kedalaman di P. Kambing, Teluk Banten (individu tiap 10 cm2)
Lokasi Ne Co Poly For Oli Os Na Iso
S1U1D1 637 278.6 7 4.2 0 0 44.8 0
S1U1D2 256.2 29.4 2.8 4.2 0 0 2.8 0
S1U1D3 78.4 19.6 0 11.2 0 0 2.8 0
S1U1D4 60.2 19.6 1.4 1.4 1.4 0 0 0
S1U1D5 65.8 7 0 0 0 0 0 0
S1U2D1 509.6 207.2 1.4 14 0 2.8 70 1.4
S1U2D2 530.6 407.4 15.4 11.2 0 1.4 42 0
S1U2D3 989.8 245 2.8 0 8.4 0 22.4 0
S1U2D4 166.6 37.8 0 0 5.6 0 2.8 0
S1U2D5 78.4 7 2.8 2.8 0 0 0 0
S1U3D1 221.2 56 1.4 0 2.8 0 1.4 0
S1U3D2 417.2 296.8 4.2 0 2.8 0 19.6 0
S1U3D3 393.4 44.8 11.2 0 1.4 0 14 0
S1U3D4 128.8 16.8 0 2.8 0 0 4.2 0
S1U3D5 29.4 7 0 0 0 0 0 0
Lokasi Ne Co Poly For Oli Os Na Aca
S2U1D1 184.8 120.4 11.2 0 0 0 9.8 0
S2U1D2 897.4 219.8 11.2 5.6 1.4 0 51.8 0
S2U1D3 154 32.2 7 1.4 0 0 8.4 0
S2U1D4 112 18.2 5.6 0 1.4 0 2.8 0
S2U1D5 39.2 9.8 0 0 0 0 1.4 0
S2U2D1 485.8 149.8 5.6 0 2.8 0 2.8 0
S2U2D2 347.2 180.6 7 2.8 0 1.4 39.2 0
S2U2D3 121.8 53.2 0 0 2.8 1.4 7 0
S2U2D4 82.6 8.4 0 0 1.4 0 1.4 0
S2U2D5 85.4 12.6 0 0 2.8 0 9.8 0
S2U3D1 70 68.6 5.6 0 8.4 0 4.2 0
S2U3D2 99.4 61.6 0 4.2 4.2 1.4 2.8 1.4
S2U3D3 49 37.8 1.4 0 5.6 0 1.4 0
S2U3D4 35 5.6 0 1.4 0 0 0 0
S2U3D5 49 9.8 0 4.2 1.4 0 2.8 0
Lokasi Ne Co Poly For Oli Os Na Kinor
S3U1D1 357 296.8 21 1.4 9.8 0 32.2 1.4
S3U1D2 127.4 72.8 2.8 1.4 7 0 4.2 0
S3U1D3 78.4 14 0 2.8 2.8 0 1.4 0
S3U1D4 32.2 2.8 0 0 0 0 0 0
S3U1D5 21 2.8 0 5.6 1.4 0 0 0
S3U2D1 372.4 707 16.8 0 1.4 0 22.4 0
S3U2D2 112 176.4 1.4 0 1.4 1.4 30.8 0
S3U2D3 71.4 67.2 5.6 4.2 0 1.4 8.4 0
S3U2D4 2.8 8.4 0 12.6 1.4 0 4.2 0
S3U2D5 16.8 5.6 0 5.6 0 0 0 0
S3U3D1 457.8 210 2.8 0 2.8 0 2.8 0
S3U3D2 296.8 37.8 1.4 1.4 4.2 0 1.4 0
S3U3D3 135.8 11.2 0 7 0 0 0 0
Lampiran 3 (Lanjutan)
Lokasi Ne Co Poly For Oli Os Na Kinor
S3U3D5 8.4 2.8 0 0 0 0 0 0
Lokasi Ne Co Poly For Oli Os Na Tar
S4U1D1 288.4 193.2 11.2 1.4 5.6 0 16.8 0
S4U1D2 114.8 16.8 1.4 2.8 2.8 1.4 2.8 0
S4U1D3 28 110.6 1.4 0 0 0 0 0
S4U1D4 25.2 0 0 2.8 0 1.4 0 0
S4U1D5 18.2 8.4 0 7 0 1.4 0 0
S4U2D1 459.2 205.8 5.6 0 2.8 0 18.2 0
S4U2D2 197.4 46.2 2.8 5.6 0 0 8.4 0
S4U2D3 140 49 0 2.8 1.4 0 7 0
S4U2D4 58.8 1.4 0 7 4.2 0 0 0
S4U2D5 11.2 4.2 0 2.8 0 0 0 1.4
S4U3D1 973 116.2 1.4 7 5.6 0 22.4 0
S4U3D2 140 42 1.4 0 1.4 0 0 0
S4U3D3 123.2 19.6 0 0 1.4 0 1.4 0
S4U3D4 32.2 1.4 0 15.4 1.4 0 0 0
S4U3D5 12.6 1.4 0 4.2 0 0 0 0
Keterangan: Ne = Nematoda, Co = Copepoda, Poly = Polychaeta, For = Foraminifera, Oli = Oligochaeta, Os = Ostracoda, Na= Nauplii, Iso = Isopoda, Aca = Acari, Kinor = Kinorhyncha, Tar = Tardigrada, S = stasiun, U = ulangan, D = kedalaman (DD11==11--22 c
Lampiran 4 Beberapa famili Nematoda di Pulau Kambing
Stasiun Famili
1 2 3 4
1 Araeolamidae 0 0 4 8
2 Belbollidae 2 0 1 0
3 Choanolaimoidae 9 1 0 7
4 Chromadoridae 48 58 24 3
5 Comesomatidae 2 25 8 27
6 Cyatholaimidae 27 7 0 3
7 Cylindrolaimidae 0 12 0 0
8 Desmodoridae 17 22 31 29
9 Desmoscolecidaepo 0 0 2 0
10 Dorylaimopsidae 0 0 1 0
11 Enchelidiidae 0 0 0 1
12 Enoplidae 0 0 1 3
13 Eurystominidae 0 0 0 1
14 Haliplectidae 0 2 0 0
15 Hypodontolaimidae 11 12 35 40
16 Ironidae 19 46 5 11
17 Lauratonematidae 0 0 0 6
18 Leptosomatidae 330 297 53 95
19 Linhomoeidae 49 21 81 31
20 Microlaimidae 0 0 0 2
21 Monhysteridae 48 30 29 61
22 Monocholaimidae 0 3 3 5
23 Meyliidae 0 4 3 0
24 Oncholaimidae 20 35 20 27
25 Oxystominidae 23 81 33 10
26 Pelagonematidae 28 39 55 80
27 Selachinematidae 14 9 1 5
28 Sphaerolaimidae 0 0 0 3
29 Thoracostomopsidae 0 0 0 2
30 Tripylidae 7 21 6 30
31 Tripyloididae 4 25 12 0
32 Xennelidae 0 1 0 4
33 Xyalidae 6 16 16 13
Total 664 767 424 507
Lampiran 5 Tipe mulut Nematoda pada stasiun pengamatan 1-4 di P. Kambing Stasiun 1 Stasiun 2 Stasiun 3 Stasiun 4
1 A 107 162 156 116
1 B 362 391 110 246
2 A 164 186 114 119
Lampiran 6 Kunci identifikasi Enhalus acoroides (Den Hartog 1970)
Famili Hydrocharitaceae
• Daun dibedakan ke dalam pelepah dan helai, tanpa ligula atau dengan petiole.
• Bunga dioecious (terkadang monoecious), dengan tiga bagian perian. Polen berbentuk bulatan, bebas, atau tersusun membentuk seperti untaian manik.
Genus Enhalus
• Rizoma tebal dan daun berbentuk seperti tali. Tulang daun terlihat jelas.
Spesies Enhalus acoroides
• Daun pipih
• Daun membujur seperti garis, biasanya panjang 30 – 150 cm, lebar 1.5 – 2 cm
• Daun tidak menyempit pada bagian bawah
• Ujung daun membulat atau tumpul, tepi daun tebal dan licin
SAGITA PUJIANTI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Kambing, Teluk Banten. Dibimbing oleh RIKA RAFFIUDIN dan SUSETIONO.
Meiofauna adalah kelompok hewan berukuran antara 0.063-1 mm. Meiofauna interstisiil hidup di rongga antar butir sedimen dan dalam distribusi verikal dapat ditemukan sampai dengan kedalaman 10 cm. Tujuan penelitian adalah mempelajari distribusi vertikal meiofauna interstisiil pada padang lamun dengan luasan tutupan berbeda di Pulau Kambing, Teluk Banten. Empat stasiun dipilih berdasarkan persentase kepadatan luasan tutupan lamun yaitu padat, sedang, rendah, dan tidak ditumbuhi lamun. Enhalus acoroides adalah spesies lamun yang mendominasi di P. Kambing. Kepadatan meiofauna meningkat di stasiun dengan tutupan lamun padat dibandingkan daerah tanpa ditumbuhi lamun. Kepadatan meiofauna interstisiil umumnya melimpah sampai dengan kedalaman 10 cm. Kepadatan meiofauna paling tinggi pada kedalaman 1-4 cm karena sedimen relatif stabil dan teroksigenisasi dengan baik. Hasil penelitian ini didapat sebelas taksa meiofauna di P. Kambing yaitu Nematoda, Copepoda, Polychaeta, Oligochaeta, Foraminifera, Ostracoda, Nauplii, Isopoda, Acari, Kinorhyncha, dan Tardigrada. Nematoda adalah taksa yang paling umum di tiap stasiun dan di tiap distribusi vertikal. Lima famili terbesar adalah Leptosomatidae, Linhomoeidae, Monhysteridae, Oxystominidae, dan Pelagonematoidae.
ABSTRACT
SAGITA PUJIANTI. Vertical Distributin of Meiofauna Interstitial in Seagrass Bed in Kambing Island, Banten Bay. Supervised by RIKA RAFFIUDIN dan SUSETIONO.
Latar Belakang
Meiofauna adalah istilah yang digunakan untuk kelompok hewan yang berukuran antara 0.063-1 mm (Bouwman 1987, diacu dalam Susetiono 1999). Anggota meiofauna dapat digolongkan ke dalam dua kelompok, yaitu meiofauna tetap dan meiofauna sementara. Meiofauna tetap adalah hewan yang sepanjang siklus hidup ukuran tubuhnya selalu berada di dalam kisaran ukuran meiofauna (Coull 1988). Taksa meiofauna tetap adalah Gastrotricha, Gnathosmulida, Kinorhyncha, Loricifera, Tardigrada, Mystacocarida, dan beberapa wakil dari Rotifera, Nematoda, Poly-chaeta, Copepoda, Ostracoda, Turbellaria, A-cari, Hydrozoa, Nemertina, Entroprocta, Gas-tropoda, Aplacophora, Branchiopoda, Holo-thuroidea, Tunicata, Priapulida, Oligochaeta, dan Sipuncula (Coull 1988). Meiofauna sementara adalah hewan yang ketika juvenil, berdasarkan ukuran tubuhnya, dikelompokkan ke dalam meiofauna, tetapi ketika dewasa dikelompok ke dalam makrofauna. Juvenil dari makrofauna umumnya adalah meiofauna sementara (Coull 1988). Kata meiofauna berasal dari bahasa Yunani, yaitu meio yang berarti lebih kecil (Higgins & Thiel 1988). Ukuran meiofauna lebih kecil dari makro-fauna, yaitu hewan yang tersaring di saringan 1 mm (Higgins & Thiel 1988), tetapi lebih besar dari mikrofauna (Giere 1993).
Meiofauna dapat ditemukan hampir di semua habitat air, mulai dari air tawar, payau sampai dengan laut. Di wilayah laut, meiofauna dapat ditemukan mulai dari daerah intertidal sampai dengan laut dalam. Meio-fauna juga dapat ditemukan diberbagai sedi-men (interstisiil) mulai dari lumpur sampai dengan kerikil kasar. Meiofauna yang hidup di sedimen dapat melakukan distribusi vertikal. Organisme yang hidup di sedimen laut disebut sebagai organisme bentik atau bentos (Brusca & Brusca 1990). Beberapa meiofauna di-temukan di atas sedimen, seperti pada daun makroalga, bagian tubuh hewan (celah koral, cacing tabung, duri Echinodermata), dan lamun (Coull 1988).
Lamun tumbuh di sedimen laut. Lamun termasuk kedalam divisi Spermatophyta; sub-divisi Angiosperma, dan mempunyai dua famili, yaitu Potamogetonaceae dan Hydro-charitaceae. Indonesia mempunyai 12 spesies lamun (Fortes 1990). Sebagian besar sedimen laut di dunia ditumbuhi lamun, kecuali di
Djamali 1995). Enhalus acoroides dan Thalassia hemprichii adalah dua spesies yang paling dominan di P. Kambing (Kiswara 2004).
Zona sebaran lamun di Teluk Banten mulai dari pantai sampai dengan tubir. Perbedaan zona sebaran lamun dijumpai dalam kom-posisi jenis lamun (vegetasi tunggal atau campuran) dan luasan tutupannya (jarang, sedang, tinggi atau sangat tinggi) (Kiswara 1992). Luasan tutupan lamun mempengaruhi kepadatan meiofauna.
Tujuan
Penelitian ini bertujuan meneliti distribusi vertikal meiofauna interstisiil pada padang lamun dengan luasan tutupan berbeda di Pulau Kambing, Teluk Banten.
Waktu dan Tempat
Penelitian ini dilaksanakan pada bulan Februari sampai dengan September 2007. Pengambilan contoh dilaksanakan pada bulan Mei 2007 di P. Kambing, Teluk Banten. Identifikasi dilakukan di Pusat Penelitian Oseangrafi (P20), LIPI, Jakarta.
BAHAN DAN METODE
Bahan dan Alat
Objek penelitian ini adalah meiofauna yang hidup di rongga antar butir sedimen (interstisiil). Bahan-bahan yang digunakan adalah contoh sedimen laut, formalin 4%, rose bengal, dan gliserol. Alat yang digunakan untuk penentuan stasiun adalah bingkai 50 x 50 cm2. Alat-alat yang digunakan untuk peng-ambilan contoh sedimen adalah core (p = 10 cm, d = 3 cm) dan (p = 30 cm, d = 5 cm) serta botol plastik. Alat-alat yang digunakan untuk menghitung parameter lingkungan adalah termometer, refraktometer, penguji tanah (soil tester), dan kertas tahan air (under water paper). Alat yang digunakan untuk identifi-kasi meiofauna adalah loop Irwin dan mikros-kop Leica ATC 2000 dan mikrosmikros-kop stereo Carton. Dokumentasi meiofauna menggu-nakan kamera Casio Exilim.
Deskripsi Tempat Pengambilan Contoh
Latar Belakang
Meiofauna adalah istilah yang digunakan untuk kelompok hewan yang berukuran antara 0.063-1 mm (Bouwman 1987, diacu dalam Susetiono 1999). Anggota meiofauna dapat digolongkan ke dalam dua kelompok, yaitu meiofauna tetap dan meiofauna sementara. Meiofauna tetap adalah hewan yang sepanjang siklus hidup ukuran tubuhnya selalu berada di dalam kisaran ukuran meiofauna (Coull 1988). Taksa meiofauna tetap adalah Gastrotricha, Gnathosmulida, Kinorhyncha, Loricifera, Tardigrada, Mystacocarida, dan beberapa wakil dari Rotifera, Nematoda, Poly-chaeta, Copepoda, Ostracoda, Turbellaria, A-cari, Hydrozoa, Nemertina, Entroprocta, Gas-tropoda, Aplacophora, Branchiopoda, Holo-thuroidea, Tunicata, Priapulida, Oligochaeta, dan Sipuncula (Coull 1988). Meiofauna sementara adalah hewan yang ketika juvenil, berdasarkan ukuran tubuhnya, dikelompokkan ke dalam meiofauna, tetapi ketika dewasa dikelompok ke dalam makrofauna. Juvenil dari makrofauna umumnya adalah meiofauna sementara (Coull 1988). Kata meiofauna berasal dari bahasa Yunani, yaitu meio yang berarti lebih kecil (Higgins & Thiel 1988). Ukuran meiofauna lebih kecil dari makro-fauna, yaitu hewan yang tersaring di saringan 1 mm (Higgins & Thiel 1988), tetapi lebih besar dari mikrofauna (Giere 1993).
Meiofauna dapat ditemukan hampir di semua habitat air, mulai dari air tawar, payau sampai dengan laut. Di wilayah laut, meiofauna dapat ditemukan mulai dari daerah intertidal sampai dengan laut dalam. Meio-fauna juga dapat ditemukan diberbagai sedi-men (interstisiil) mulai dari lumpur sampai dengan kerikil kasar. Meiofauna yang hidup di sedimen dapat melakukan distribusi vertikal. Organisme yang hidup di sedimen laut disebut sebagai organisme bentik atau bentos (Brusca & Brusca 1990). Beberapa meiofauna di-temukan di atas sedimen, seperti pada daun makroalga, bagian tubuh hewan (celah koral, cacing tabung, duri Echinodermata), dan lamun (Coull 1988).
Lamun tumbuh di sedimen laut. Lamun termasuk kedalam divisi Spermatophyta; sub-divisi Angiosperma, dan mempunyai dua famili, yaitu Potamogetonaceae dan Hydro-charitaceae. Indonesia mempunyai 12 spesies lamun (Fortes 1990). Sebagian besar sedimen laut di dunia ditumbuhi lamun, kecuali di
Djamali 1995). Enhalus acoroides dan Thalassia hemprichii adalah dua spesies yang paling dominan di P. Kambing (Kiswara 2004).
Zona sebaran lamun di Teluk Banten mulai dari pantai sampai dengan tubir. Perbedaan zona sebaran lamun dijumpai dalam kom-posisi jenis lamun (vegetasi tunggal atau campuran) dan luasan tutupannya (jarang, sedang, tinggi atau sangat tinggi) (Kiswara 1992). Luasan tutupan lamun mempengaruhi kepadatan meiofauna.
Tujuan
Penelitian ini bertujuan meneliti distribusi vertikal meiofauna interstisiil pada padang lamun dengan luasan tutupan berbeda di Pulau Kambing, Teluk Banten.
Waktu dan Tempat
Penelitian ini dilaksanakan pada bulan Februari sampai dengan September 2007. Pengambilan contoh dilaksanakan pada bulan Mei 2007 di P. Kambing, Teluk Banten. Identifikasi dilakukan di Pusat Penelitian Oseangrafi (P20), LIPI, Jakarta.
BAHAN DAN METODE
Bahan dan Alat
Objek penelitian ini adalah meiofauna yang hidup di rongga antar butir sedimen (interstisiil). Bahan-bahan yang digunakan adalah contoh sedimen laut, formalin 4%, rose bengal, dan gliserol. Alat yang digunakan untuk penentuan stasiun adalah bingkai 50 x 50 cm2. Alat-alat yang digunakan untuk peng-ambilan contoh sedimen adalah core (p = 10 cm, d = 3 cm) dan (p = 30 cm, d = 5 cm) serta botol plastik. Alat-alat yang digunakan untuk menghitung parameter lingkungan adalah termometer, refraktometer, penguji tanah (soil tester), dan kertas tahan air (under water paper). Alat yang digunakan untuk identifi-kasi meiofauna adalah loop Irwin dan mikros-kop Leica ATC 2000 dan mikrosmikros-kop stereo Carton. Dokumentasi meiofauna menggu-nakan kamera Casio Exilim.
Deskripsi Tempat Pengambilan Contoh