TANGGAP MORFOFISIOLOGI BIBIT KAKAO
YANG DIBERI FOSFAT ALAM AYAMARU PAPUA,
ASAM HUMAT, INOKULASI FMA DAN
BAKTERI PELARUT FOSFAT
ANTONIUS SUPARNO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Tanggap Morfofisiologi Bibit Kakao yang Diberi Fosfat Alam Ayamaru Papua, Asam Humat, Inokulasi FMA, dan Bakteri Pelarut Fosfat adalah karya saya sendiri dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Mei 2008
Antonius Suparno
ii
ABSTRACT
ANTONIUS SUPARNO. Morpho-physiological Responses of Cacao Seedlings to The Addition of Papuan Ayamaru Phosphate Rock, Humic Acid, and the Inoculation AM Fungi and Phosphate Solubilizing Bacteria. Under direction of SUDIRMAN YAHYA, SUDRADJAT,YADI SETIADI, and KOMARRUDDIN IDRIS
The purposes of the study were to obtain a humic acid dosage, an arbuscular mycorrhizal fungi (AMF) inoculant, and a phosphate solubilizing bacteria (PSB) isolate for improving the response of cacao seedlings to Ayamaru phosphate rock (APR). The study was conducted in the Cikabayan research garden of IPB. F1 UAH cacao seeds from Coffee and Cacao Research Center, Jember, East Java was used and the seedlings were grown on Ultisol, acid soil from Jasinga in 20 cm x 30 cm sized polybag. The seedlings were grown under 60% of shading net for the period of four months.
The study consists of four trials which were set up in two-factor factorial trials using a Completely Randomized Design. The first factor in Trial I, II and III were APR dosages: 0, 0.5, 1.0, 1.5, and 2.0 g P2O5/seedling and 2.0 g P2O5 SP36/seedling used as comparison. The second factor in Trial I were AMF inoculation; in Trial II were PSB isolate inoculation; and in Trial III were four levels of humic acid dosages. The first factor in Trial IV was four levels of APR dosages: 0, 1.0, 2.0, and 4.0 g P2O5/seedling and 2.0 g P2O5 SP36/seedling used as comparison. While the second factor was combination treatments from the best result of Trial I, II, and III.
The best treatments obtained from Trial I, II and III as followed: AMF inoculation using Mycofer inoculant, inoculation with FT.3.2 isolate, and 3.10-3 mL of humic acid. Trial IV revealed that stem diameter, leaf number, and shoot/root ratio of the seedling with 2.0 g P2O5/seedling of APR was not significantly different than the seedling with 2.0 g P2O5 SP36/seedling. The response of most variables of seedlings to APR up to 4.0 g P2O5 in all combination treatments of the second factor on the Trial IV was linear. However, the treatments which involved AMF inoculation gave the better result as comparing to seedling without AMF. In term of shoot dry-weight, the effectiveness of Mycofer, humic acid and PSB was 104.29%, 4.38% and 4.24%; in term of P uptake was 191.00%, 33.20% and 18.31%; in term of root colonization was 682.82%, 10.26% and 6.17% ; in term of acid phosphatase activity was 30.07%, 7.58%, 7.34%, while in term of APR efficiency was 104.29%, 4.38%, and 4.24% respectively.
Combination treatments in this study were aimed to enhance available P and to lower exchangeable Al content of the media. Combination treatments with the increasing level of APR up to 2.0 g P2O5/seedling caused increasing the root colonization and acid phosphatase activity went a long with decreasing exchangeable Al content, but with the increasing level of APR up to 4.0 g P2O5/seedling caused decreasing the root colonization and acid phosphatase activity and on the contrary increasing exchangeable Al content of the media.
The response of the growth seedlings and P available content of the media up to 4.0 g P2O5 of APR was still increased linearly. The responses were improved by the application of humic acid, AMF and PSB inoculation. The synthesis of organic acids by PSB inoculation and the release of acid phosphatase by AMF to the media indicated that there were the external mechanism of P solubilization of phosphate rock.
Keywords: Theobroma cacao, Gigaspora margarita, Glomus etunicatum,
iii
RINGKASAN
ANTONIUS SUPARNO. Tanggap Morfofisiologi Bibit Kakao yang Diberi Fosfat Alam Ayamaru Papua, Asam Humat, Inokulasi FMA, dan Bakteri Pelarut Fosfat. Dibimbingan oleh SUDIRMAN YAHYA, SUDRADJAT, YADI SETIADI, dan KOMARUDDIN IDRIS.
Areal pengembangan kakao umumnya mengarah pada lahan dengan tingkat kesuburan tanah rendah. Salah satu kriteria yang menjadi hal penting dalam pengusahaan perkebunan kakao adalah penggunaan bibit bermutu. Bertolak dari hal tersebut maka penelitian ini dilakukan untuk mengkaji penggunaan fosfat alam Ayamaru Papua (FA) sebagai pupuk pada pembibitan kakao yang diinokulasi fungi mikoriza arbukula (FMA), bakteri pelarut fosfat (BPF), dan pemberian asam humat. Penelitian ini bertujuan untuk mendapatkan inokulum FMA, isolat BPF, tingkat dosis asam humat, maupun kombinasi ketiganya yang efektif meningkatkan daya guna FA sehingga diperoleh kualitas bibit kakao yang lebih baik.
Penelitian dilaksanakan dalam jaring naungan 60% selama empat bulan. Bibit kakao ditanam pada polibag 20 cm x 30 cm dengan media tanah Ultisol Jasinga. Benih kakao yang digunakan adalah UAH F1 dari Pusat Penelitian Kopi dan Kakao Jember, sedangkan FA berasal dari Distrik Ayamaru, Papua. Penelitian terdiri atas empat tahapan percobaan dan dilaksanakan menggunakan Rancangan Acak Lengkap dua faktor yang. Faktor pertama pada percobaan I, II, dan III adalah dosis FA: 0, 0.5, 1.0, 1.5, 2.0 g P2O5/bibit dan 2.0 g P2O5 SP36/bibit sebagai pembanding. Faktor kedua pada percobaan I adalah inokulasi FMA : tanpa FMA (Mo), FMA indigenous Manokwari (M1), dan FMA Mycofer (M2). Faktor kedua pada percobaan II adalah inokulasi BPF : tanpa BPF (Bo), isolat RJM.30.2 (B1), dan isolat FT.3.2 (B2). Faktor kedua pada percobaan III adalah tingkat dosis asam humat yaitu : 0, 1.10-3, 2.10-3, dan 3.10-3 mL asam humat/bibit. Pada percobaan IV, faktor pertama adalah dosis FA : 0, 1.0, 2.0, 4.0 g P2O5/bibit dan 2.0 g P2O5 SP36/bibit sebagai pembanding. Faktor kedua pada percobaan IV adalah kombinasi hasil terbaik dari percobaan I, II, dan III.
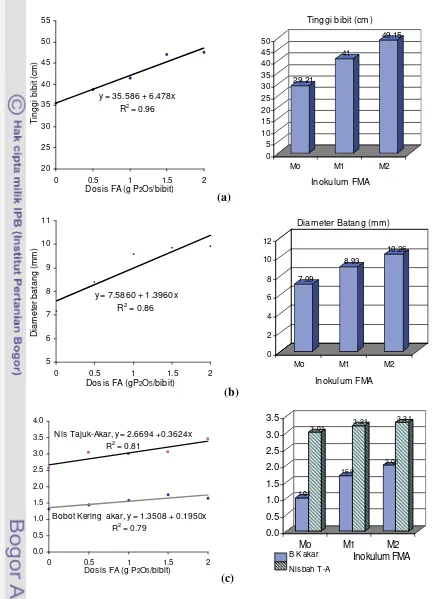
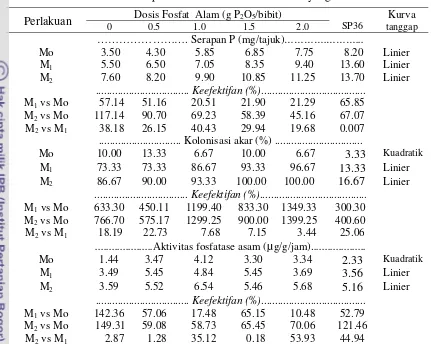
Hasil percobaan I menunjukkan bahwa tingkat dosis FA mulai berpengaruh pada bibit kakao pada umur 4 minggu setelah tanam (MST), sedangkan inokulasi FMA mulai menunjukkan pengaruhnya pada umur 8 MST. Interaksi antara tingkat dosis FA dan inokulasi FMA berpengaruh nyata pada bobot kering tajuk, jumlah daun, serapan P tajuk, kolonisasi akar, dan aktivitas fosfatase asam. Tanggap tinggi bibit, diameter batang, bobot kering akar, dan kadar Aldd media pada pemberian FA 2.0 gP2O5/bibit tidak berbeda nyata dengan pemberian 2.0 g P2O5 SP36/bibit. Hasil uji Ortogonal polinomial menunjukkan bahwa tanggap bibit kakao akibat tingkat dosis FA hingga 2.0 g P2O5/bibit adalah linier. Peningkatan dosis FA hingga 2.0 gP2O5/bibit meningkatkan pertumbuhan bibit kakao, dan pertumbuhan ini lebih meningkat apabila diinokulasi FMA. Pada semua peubah menunjukkan bahwa FMA Mycofer lebih efektif daripada FMA indigenous Manokwari. Pada dosis FA 2.0 g P2O5/bibit, apabila dibandingkan dengan bibit tanpa FMA, Mycofer lebih efektif meningkatkan bobot kering (127.55%) daripada FMA indigenous Manokwari (95.97%), sedangkan serapan P tajuk masing-masing meningkat sebesar 45.16% dan 21.29%.
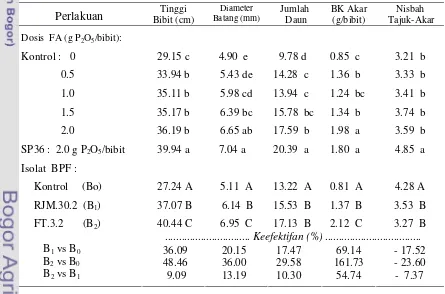
iv Tanggap diameter batang dan bobot kering akar bibit kakao pada dosis FA 2.0 gP2O5/bibit tidak berbeda nyata dengan pemberian 2.0 g P2O5 SP36/bibit, sedangkan tanggap tinggi, jumlah daun, dan nisbah tajuk-akar bibit tertinggi diperoleh pada pemberian 2.0 g P2O5 SP36/bibit. Hasil uji Ortogonal polinomial menunjukkan bahwa tanggap bibit kakao akibat tingkat dosis FA hingga 2.0 g P2O5/bibit adalah linier. Peningkatan dosis FA hingga 2.0 gP2O5/bibit meningkatkan pertumbuhan bibit kakao, dan pertumbuhan ini lebih meningkat bila diinokulasi BPF. Pada semua tingkat dosis FA, tanggap bobot kering tajuk memberikan hasil tertinggi bila diinokulasi isolat FT.3.2 dan yang terendah diperoleh pada bibit tanpa inokulasi BPF. Pada dosis FA 2.0 g P2O5/bibit, bila dibandingkan dengan kontrol, isolat RJM.30.2 meningkatkan bobot kering tajuk sebesar 48.41% dan kadar P tersedia 3.12%, sedangkan isolat FT.3.2 masing-masing 78.18% dan 9.36%. .
Hasil percobaan III menunjukkan bahwa baik tingkat dosis FA maupun asam humat mulai berpengaruh pada bibit kakao umur 4 MST. Interaksi antara tingkat dosis FA dan tingkat dosis asam humat berpengaruh nyata pada tinggi, diameter batang, bobot kering tajuk, kadar P total, P tersedia, dan Aldd media. Hasil uji Ortogonal polinomial menunjukkan bahwa tanggap bibit kakao akibat tingkat dosis FA hingga 2.0 gP2O5/bibit maupun tingkat dosis asam humat adalah linier. Pada dosis asam humat 3.10-3 mL, peningkatan dosis FA hingga 2.0 g P2O5/bibit secara linier meningkatkan kadar P tersedia sebesar 117% dan bobot kering tajuk 49.02%. Sedangkan pada dosis FA 2.0 g P2O5/bibit, peningkatan dosis asam humat hingga 3.10-3 mL/bibit secara linear meningkatkan kadar P tersedia sebesar 44.15% dan bobot kering tajuk 62.12%. Pada dosis FA 2.0 g P2O5/bibit, bila dibandingkan dengan kontrol, dosis 1.10-3, 2.10-3, dan 3.10-3 mL asam humat/bibit masing-masing memberikan kadar P tersedia berturut-turut 7.82%, 29.75%, dan 40.81%; dan bobot kering tajuk 39.13%, 45.27%, dan 70.08%.
Inokulum FMA terbaik dari percobaan I adalah FMA Mycofer, isolat terbaik dari percobaan II adalah isolat FT.3.2, sedangkan dosis asam humat terbaik dari percobaan III adalah 3.10-3 mL/bibit. Oleh karena itu faktor kedua percobaan IV adalah kombinasi: Mycofer + 3.10-3 mL asam humat (MH), Mycofer + isolat FT.3.2 (MB), isolat FT.3.2 + 3.10-3 mL asam humat (BH), dan Mycofer + isolat FT.3.2 + 3.10 -3
mL asam humat (MBH). Baik tingkat dosis FA maupun perlakuan kombinasi telah menunjukkan pengaruhnya pada bibit kakao umur 4 MST. Interaksi antara tingkat dosis FA dan kombinasi perlakuan berpengaruh nyata pada tanggap bobot kering tajuk, serapan P tajuk, aktivitas fosfatase asam, kolonisasi akar, pH media, P total, P tersedia, dan kadar Aldd media. Tanggap bibit kakao akibat dosis FA hingga 4.0 g P2O5/bibit pada semua perlakuan kombinasi untuk sebagian besar peubah adalah linier. Namun demikian, perlakuan kombinasi yang melibatkan inokulasi FMA memberikan hasil terbaik dibandingkan tanpa inokulasi FMA. Terhadap bobot kering tajuk keefektifan FMA Mycofer adalah 104.29%, asam humat 4.38%, dan bakteri pelarut fosfat 4.24%, terhadap serapan P tajuk berturut-turut 191.00%, 30.20% dan 18.31%, terhadap kolonisasi akar 681.82%, 10.26%, 6.17%, terhadap aktivitas fosfatase asam 30.07%, 7.58%, 7.34%, sedangkan terhadap efisiensi FA adalah 104.29%, 4.38%, dan 4.24%.
v
Tanggap pertumbuhan dan kadar P tersedia pada bibit terhadap FA hingga 4.0 g P2O5/bibit masih meningkat secara linier. Tanggap pertumbuhan bibit dan kadar P tersedia lebih ditingkatkan apabila diberi asam humat, inokulasi BPF dan FMA. Sintesis asam-asam organik oleh BPF dan pelepasan asam fosfatase ke media oleh akar bibit yang diinokulasi FMA mengidikasikan terjadinya mekanisme eksternal pelarutan FA.
Kata kunci: Theobroma cacao, Gigaspora margarita, Glomus etunicatum, Glomus manihotis, Glomus aggregatum, Acaulospora tuberculata,
Acaulospora scrobiculata
vi
@ Hak Cipta milik IPB tahun 2008
Hak Cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian,
penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
vii
TANGGAP MORFOFISIOLOGI BIBIT KAKAO
YANG DIBERI FOSFAT ALAM AYAMARU PAPUA,
ASAM HUMAT, INOKULASI FMA, DAN
BAKTERI PELARUT FOSFAT
Oleh
ANTONIUS SUPARNO
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Departemen Agronomi dan Hortikultura
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
viii Judul Disertasi : Tanggap Morfofisiologi Bibit Kakao yang Diberi Fosfat
Alam Ayamaru Papua, Asam Humat, Inokulasi FMA, dan Bakteri Pelarut Fosfat
Nama : Antonius Suparno NIM : A361040081
Disetujui Komisi Pembimbing
Prof. Dr. Ir. Sudirman Yahya, M.Sc Dr. Ir. Sudradjat, MS K e t u a Anggota
Dr. Ir. Yadi Setiadi, M.Sc Dr. Ir. Komaruddin Idris, MS Anggota Anggota
Diketahui
Ketua Program Studi Agronomi Dekan Sekolah Pascasarjana
Dr. Ir. Munif Ghulamahdi, MS Prof. Dr. Ir. Khairil A. Notodiputro, MS
ix
PRAKATA
Puji dan syukur penulis panjatkan ke Hadirat Tuhan atas segala hikmat dan karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Penelitian ini telah dilaksanakan sejak bulan Agustus 2006 di Kebun Percobaan Cikabayan IPB, Bogor. Disertasi ini memuat satu bab yang merupakan pengembangan dari naskah artikel yang diajukan ke jurnal ilmiah. Bab 3 berjudul Peranan FMA dalam Tanggap Bibit Kakao terhadap Fosfat Alam Ayamaru, dengan judul artikel Pertumbuhan dan Serapan P Bibit Kakao (Theobroma cacao L.) yang Diberi Fosfat Alam Papua dan Inokulasi Mikoriza sedang menunggu penerbitan di Jurnal Buletin Agronomi Vol.XXXVI No. 1 April Th. 2008.
Pada kesempatan ini, penulis mengucapkan terima kasih kepada :
1. Bapak Prof. Dr. Ir. Sudirman Yahya, M.Sc selaku Ketua Komisi Pembimbing, Bapak Dr. Ir. Sudradjat, MS, Bapak Dr. Ir. Yadi Setiadi, M.Sc, dan Bapak Dr. Ir. Komaruddin Idris, MS selaku Anggota Komisi Pembimbing atas segala perhatian dan bimbingannya selama penulis mempersiapkan dan melaksanakan penelitian hingga penulisan disertasi ini.
2. Staf Dosen pada Program Pascasarjana IPB atas bimbingannya dalam perkuliahan dan praktikum selama penulis mengikuti program S3.
3. Rektor Unversitas Negeri Papua Manokwari yang telah memberikan ijin kepada penulis untuk mengikuti program S3 di Program Studi Agronomi pada Program Pascasarjana IPB
4. Rektor IPB, Dekan Pascasarjana, dan Ketua Program Studi Agronomi IPB atas kesempatan yang diberikan kepada penulis untuk mengikuti program S3 dan penggunaan fasilitas yang tersedia
5. Manajemen Program Due-Like Unipa-Dikti atas beasiswa pendidikan yang diberikan kepada penulis, sehingga dapat membantu penulis dalam melaksanakan pendidikan S3
x
7. Rekan mahasiswa S2 dan S3 Pascasarjana IPB dan laboran atas bantuan dan kerjasama yang telah diberikan, kepada pak Abimanyu dan Ibu Yudi, pak Yulius, pak Iwan Sasli, pak Wasgito, pak Takdir, mbak Faiq, mbak Susan dan semua rekan dari Papua
8. Semua pihak yang tidak dapat penulis sebutkan seluruhnya yang telah membantu penulis selama melaksanakan pendidikan S3
Semoga segala perhatian, bantuan, dan pengorbanan bapak dan ibu serta saudara/saudari sekalian mendapatkan berkat dan imbalan dari Tuhan Yang Maha Kuasa.
Semoga karya ilmiah ini bermanfaat.
Bogor, Mei 2008
xi
RIWAYAT HIDUP
Penulis dilahirkan di Gunung Kidul pada tanggal 16 Juni 1964 sebagai anak ke enam dari tujuh bersaudara dari ayah P. Jatmodikromo dan M. Sadjem. Pendidikan sarjana ditempuh di Program Studi Agronomi, Fakultas Pertanian Universitas Cenderawasih Manokwari, lulus pada tahun 1989. Pada tahun 1995, penulis diterima di Program Studi Ilmu Tanaman pada Program Pascasarjana Universitas Padjadjaran (UNPAD) Bandung dan menamatkannya pada tahun 1998. Kesempatan untuk melanjutkan ke program doktor di Program Studi Agronomi pada Program Pascasarjana IPB diperoleh pada tahun 2004. Beasiswa pendidikan pascasarjana diperoleh dari Program Due-Like Unipa-Dikti tahun 2004.
Penulis bekerja sebagai staf pengajar pada Fakultas Pertanian Universitas Negeri Cenderawasih Manokwari sejak tahun 1991. Pada tahun 2001, Fakultas Pertanian ini telah dikembangkan menjadi Universitas Negeri Papua Manokwari.
Karya ilmiah berjudul Pengaruh Pemberian Fosfat Alam Ayamaru Papua dan Inokulasi Fungi Mikoriza Arbuskula (FMA) terhadap Pertumbuhan Bibit Kakao telah disajikan pada Seminar Nasional dan Kongres II Asosiasi Mikoriza Indonesia di Bogor pada bulan Juli 2007. Karya ilmiah lain berjudul Pengaruh Pemberian Fosfat Alam Ayamaru Papua dan Kombinasi Asam Humat, Inokulasi FMA dan Bakteri Pelarut Fosfat terhadap Pertumbuhan Bibit Kakao telah disajikan pada Simposium, Seminar Nasional dan Kongres IX Perhimpunan Agronomi Indonesia (PERAGI) di Unpad Bandung pada November 2007. Artikel lain berjudul Pertumbuhan dan Serapan P Bibit Kakao (Theobroma cacao L.) yang Diberi Fosfat Alam Papua dan Inokulasi Mikoriza akan diterbitkan pada Jurnal Buletin Agronomi Vol.XXXVI No. 1 April Th. 2008. Karya-karya ilmiah tersebut merupakan bagian dari program S3 penulis.
Di samping melaksanakan pendidikan S3, penulis telah mengikuti Pelatihan Identifikasi Jamur Mikoriza Arbuskula di Lembaga Ilmu Pengetahuan Indonesia (LIPI), Bidang Botani-Pusat Penelitian Biologi pada bulan Agustus 2007.
xii
DAFTAR ISI
Halaman
DAFTAR TABEL ……..……… vii
DAFTAR GAMBAR ………. ix
DAFTAR LAMPIRAN ………. xiii
I PENDAHULUAN Latar Belakang …….………. 1
Tujuan dan Manfaat ………. 11
Hipotesis ……….. 12
II TINJAUAN PUSTAKA Ketersediaan Fosfor Tanah ……….. 13
Peranan FMA terhadap Penyerapan Fosfor ………... 16
Fosfat Alam Ayamaru Papua ……… 19
Bakteri Pelarut Fosfat ……….. 23
Peranan Asam Humat ……… ………... 26
III TANGGAP BIBIT KAKAO YANG DIINOKULASI FMA PADA BERBAGAI DOSIS FOSFAT ALAM AYAMARU Pendahuluan ………. 31
Bahan dan Metode ……….. 33
Hasil ………. 38
Pembahasan ………. 51
Kesimpulan ………….……….. 58
IV TANGGAP BIBIT KAKAO YANG DIINOKULASI INOKULASI BAKTERI PELARUT FOSFAT PADA BERBAGAI DOSIS FOSFAT ALAM AYAMARU Pendahuluan ……….... 61
Bahan dan Metode ……….………... 63
Hasil ……… 68
Pembahasan …….………... 79
xiii V TANGGAP BIBIT KAKAO PADA BEBERAPA DOSIS FOSFAT
ALAM AYAMARU DAN ASAM HUMAT
Pendahuluan ……….... 86
Bahan dan Metode ..………. 88
Hasil ………..………... 92
Pembahasan ………. 104
Kesimpulan ………….………. 109
VI TANGGAP BIBIT KAKAO YANG DIINOKULASI FMA DAN BAKTERI PELARUT FOSFAT PADA BERBAGAI DOSIS FOSFAT ALAM AYAMARU YANG DIBERI ASAM HUMAT Pendahuluan ……… 112
Bahan dan Metode……… 115
Hasil ………..……….. 119
Pembahasan ……… 133
Kesimpulan ………….……… 139
VII PEMBAHASAN UMUM ………. 141
VIII KESIMPULAN DAN SARAN ………. 154
xiv
DAFTAR TABEL
Halaman
1 Karakteristik hasil analisis kandungan unsur dari asam humat dan
asam fulvat (Schnitzer 1997) ... 29 2 Tanggap tinggi, diameter batang, bobot kering akar, dan nisbah
tajuk-akar bibit kakao terhadap dosis FA pada inokulum FMA yang berbeda ... 39 3 Tanggap jumlah daun dan bobot kering tajuk bibit kakao terhadap dosis
fosfat alam pada inokulum FMA yang berbeda ... 43 4 Tanggap kadar P tajuk, pH media, kadar P tersedia, kadar P total, kadar
Aldd media bibit kakao terhadap dosis fosfat alam pada inokulum FMA yang berbeda ...
45 5 Tanggap serapan P tajuk, kolonisasi akar, dan kadar fosfatase asam akar
bibit kakao terhadap dosis FA dan inkulum FMA yang berbeda ... 48 6 Jenis-jenis asam organik yang dihasilkan oleh inkubasi isolat bakteri ... 69 7 Tanggap tinggi, diameter batang, jumlah daun, bobot kering akar, dan
nisbah tajuk-akar bibit kakao terhadap dosis fosfat alam pada isolat BPF yang berbeda ...
70 8 Tanggap kadar P tajuk, serapan P tajuk, dan pH media terhadap dosis
fosfat alam pada isolate BPF yang berbeda ... 74 9 Tanggap bobot kering tajuk, kadar P tersedia, kadar P total, dan kadar
Aldd media akibat dosis fosfat alam pada isolat BPF yang berbeda ... 76 10 Tanggap jumlah daun, bobot kering akar, dan nisbah tajuk-akar bibit
kakao terhadap dosis fosfat alam pada berbagai dosis asam humat ... 93 11 Tanggap tinggi dan diameter batang bibit kakao akibat dosis fosfat alam
pada berbagai dosis asam humat ... 96 12 Tanggap bobot kering tajuk, kadar P tersedia, kadar P total, dan
kadar Aldd media akibat dosis fosfat alam pada berbagai dosis
asam humat ... 98 13 Tanggap kadar P, serapan P tajuk bibit kakao, dan pH media terhadap
dosis fosfat alam pada berbagai dosis asam humat ... 102 14 Tanggap tinggi, diameter batang, jumlah daun, kadar P tajuk, bobot
kering akar, dan nisbah tajuk-akar bibit kakao terhadap dosis
fosfat alam pada berbagai kombinasi M, B, H ... 121 15 Uji Ortogonal polinomial tanggap bobot kering tajuk, serapan P tajuk,
kolonisasi akar, kadar fosfatase asam, kadar P tersedia, kadar P total, pH media, dan kadar Aldd akibat dosis fosfat alam pada berbagai
kombinasi M,B, H ... 127 16 Uji Ortogonal kontras bobot kering tajuk, serapan P tajuk,
kolonisasi akar, dan kadar fosfatase terhadap berbagai perlakuan
xv
17 Tanggap kadar P tersedia, kadar P total, pH media, dan kadar Aldd akibat dosis fosfat alam pada berbagai perlakuan kombinasi M, B, dan H ...
130 18 Uji Ortogonal kontras kadar P tersedia, kadar P total, pH, dan kadar Aldd
media akibat berbagai perlakuan kombinasi M, B, H ... 132 19 Efisiensi FA terhadap bobot kering tajuk bibit kakao pada dosis FA 4.0 g
xvi
DAFTAR GAMBAR
Halaman 1 Bagan alir tahapan penelitian ……… 10 2 Struktur infeksi FMA pada sel-sel kortek akar (Diouf et al. 2003) ... 17 3 Lokasi deposit fosfat alam Ayamaru Papua ... 21 4 Model proses-proses utama yang menunjukkan reduksi jerapan P dan
peningkatan ketersediaan P oleh aplikasi residu organik ke tanah (Haynes
& Mokolobate 2001)………... ………... 27 5 Pengaruh dosis fosfat alam dan jenis inokulum FMA terhadap tinggi (a),
diameter batang (b), bobot kering akar dan nisbah tajuk-akar (c)
bibit kakao ... 41 6 Penampilan bibit kakao yang diberi fosfat alam dan inokulasi FMA ……… 42 7 Tanggap Jumlah daun dan bobot kering tajuk akibat dosis fosfat alam pada
inokulasi FMA yang berbeda... 43 8 Pengaruh tingkat dosis fosfat alam dan ionokulasi FMA terhadap kadar P
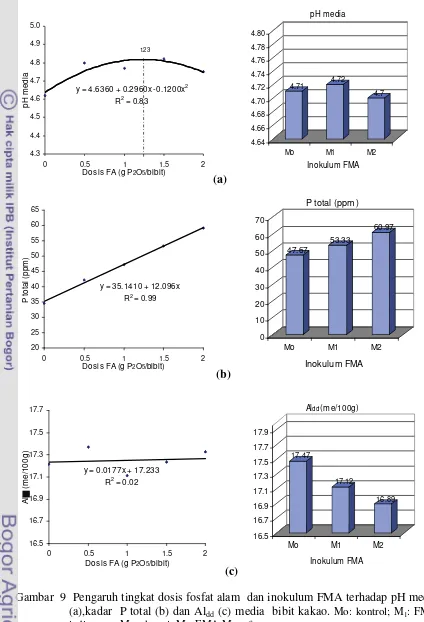
tajuk dan kadar P tersedia ... 46 9 Pengaruh tingkat dosis fosfat alam dan inokulum FMA terhadap pH media,
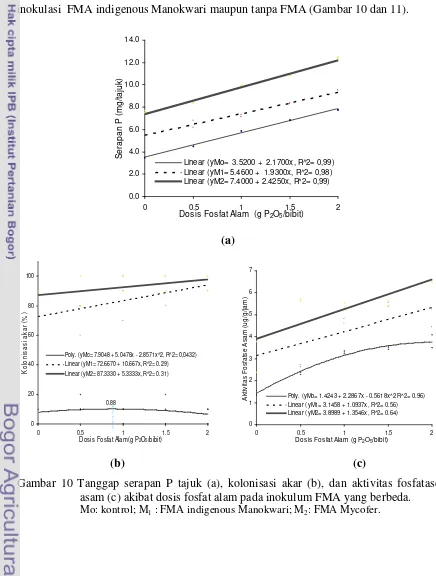
kadar P total, dan Aldd media bibit kakao ... 47 10 Tanggap serapan P tajuk, kolonisasi akar, dan aktivitas fosfatase asam
akibat dosis fosfat alam pada inokulum FMA yang berbeda... 49 11 Spora spesies FMA yang terdapat dalam inokulum Mycofer dan spora
spesies FMA indigenous Manokwari ... 50 12 Kolonisasi akar oleh FMA Mycofer dan FMA indigenous Manokwari …….. 51 13 Tanggap tinggi, diameter batang, dan jumlah daun bibit kakao terhadap
dosis fosfat alam pada isolat BPF yang berbeda... 71 14 Tanggap jumlah daun (a) dan bobot kering akar (b) bibit kakao terhadap
dosis fosfat alam pada isolat BPF yang berbeda... 72 15 Penampilan pertumbuhan bibit kakao yang diinokulasi BPF dan dosis
fosfat alam ... 73 16 Tanggap kadar P Tajuk, serapan P tajuk, dan pH media akibat dosis fosfat
alam pada isolat BPF yang berbeda. ... 75 17 Tanggap bobot kering tajuk, kadar P tersedia, kadar P total, dan kadar Aldd
media akibat dosis fosfat alam pada isolat BPF yang berbeda ... 78 18 Tanggap jumlah daun, bobot kering akar, dan nisbah tajuk-akar akibat
tingkat dosis fosfat dan dosis asam humat ... 94 19 Penampilan bibit kakao akibat pengaruh berbagai dosis asam humat dan
xvii
20 Tanggap tinggi dan diameter batang bibit kakao akibat dosis fosfat alam
pada berbagai dosis asam humat... 97 21 Tanggap bobot kering tajuk, kadar P tersedia, kadar P total, dan kadar Aldd
media akibat dosis FA pada berbagai dosis asam humat... ... 99 22 Kurva tiga dimensi tanggap bobot kering tajuk akibat dosis fosfat alam
pada berbagai dosis asam humat. …... 100 23 Kurva kontour tanggap bobot kering tajuk akibat dosis fosfat alam pada
berbagai dosis asam humat ... 101 24 Tanggap kadar P tajuk, serapan P tajuk, dan kadar Aldd mediaakibat
dosis fosfat alam pada berbagai dosisi asam humat ... 103 25 Penampilan bibit kakao akibat kombinasi M, B,H pada dosis P dan P36 , dan
akibat tingkat dosis fosfat alam Ayamaru pada kombinasi MBH ... ... 120 26 Kurva tanggap tinggi dan diameter batang bibit kakao terhadap dosis fosfat
alam dan histogram perlakuan kombinasi FMA (M), bakteri pelarut fosfat
(B), dan asam humat (H) ... 123 27 Kurva tanggap jumlah daun dan bobot kering akar bibit kakao terhadap
dosis fosfat alam dan histogram perlakuan kombinasi mikoriza (M), bakteri
pelarut fosfat (B), dan asam humat (H) ... 124 28 Kurva tanggap nisbah tajuk-akar dan kadar P tajuk terhadap dosis fosfat
alam dan histogram perlakuan kombinasi mikoriza (M), bakteri pelarut
fosfat (B), dan asam humat (H) ... 125 29 Kurva tanggap bobot kering tajuk, serapan P tajuk, kolonisasi akar dan
aktivitas fosfatase asam akibat dosis fosfat alam pada berbagai perlakuan
kombinasi M,B, H ... 127 30 Tanggap kadar P tersedia, kadar P total, pH, dan kadar Aldd media akibat
dosis fosfat alam pada berbagai kombinasi mikoriza (M), bakteri pelarut
fosfat (B), dan asam humat (H)... 131 31 Fenomena rizosfer akibat perlakuan pemberian fosfat alam Ayamaru (FA),
asam humat, inokulasi fungi mikoriza arbuskula (FMA), dan
xviii
DAFTAR LAMPIRAN
Halaman
1 Penghitungan jumlah spora dengan teknik penyaringan basah
(Gaderman & Nicholson 1963) ... 166
2 Metode Pengujian Most Probable Number FMA (Norris et al. 1992)…….. 167
3 Perhitungan kolonisasi akar dengan teknik pewarnaan Akar dari Phillips dan Hayman (1970) ... 168
4 Komposisi media Pikovskaya ... 169
5 Komposisi media kaldu nutrien ... 170
6 Hasil analisis fosfat alam Ayamaru Papua di Balai Penelitian Tanah Pusat Bogor tahun 2006 ... 171
7 Analisis kimia tanah Ultisol Jasinga ... 172
8 Metode analisis fosfatase asam (Tabatabai & Bremer 1969) ... 173
9a F-hitung analisis ragam peubah tanggap bibit kakao yang diinokulasi FMA pada berbagai dosis FA ………. 175
9b F-hitung analisis ragam peubah tanggap bibit kakao yang diinokulasi BPF pada berbagai dosis FA ……… 176
9c F-hitung analisis ragam peubah tanggap bibit kakao pada berbagai konsentrasi asam humat dan dosis FA ………... 177
9d F-hitung analisis ragam peubah tanggap bibit kakao yang diinokulasi kombinasi FMA, BPF, dan asam humat pada berbagai dosis FA ……….. 178
10a F-hitung analisis Ortogonal polinomial tanggap bibit kakao yang diinokulasi FMA pada berbagai dosis FA ………. 179
10b F-hitung analisis Ortogonal polinomial tanggap bibit kakao yang diinokulasi BPF pada berbagai dosis FA ……….. 180
10c F-hitung analisis Ortogonal polinomial tanggap bibit kakao yang diaplikasi asam humat pada berbagai dosis FA ……… 181
10d F-hitung analisis Ortogonal polinomial tanggap bibit kakao terhadap dosis fosfat alam pada berbagai kombinasi mikoriza (M), bakteri pelarut fosfat (B), dan asam humat (H)……… ………... 183
11 Nilai F-hitung Uji Ortogonal kontras perlakuan kombinasi FMA, bakteri pelarut posfat, dan Asam Humat ……….. 185
12a Deskripsi Glomus aggreratum Schenck & Smith (1982) ... 186
12b Deskripsi Acaulospora scrobiculata Trappe (1977)………... 187
I
PENDAHULUAN
Latar Belakang
Keberhasilan dalam budidaya pertanian terutama tanaman tahunan seperti kakao salah satunya ditentukan oleh kualitas bibit. Untuk mendapatkan bibit yang berkualitas diperlukan penanganan sejak awal baik dengan pemupukan, maupun dengan menginokulasikan agen hayati yang menguntungkan sehingga apabila bibit ditanam dan dipelihara di lapang dapat tumbuh dan berproduksi secara optimal.
Pengembangan perkebunan kakao sangat penting karena sektor ini berperan dalam menyediakan lapangan kerja, sumber pendapatan petani, dan sumber devisa negara (Hartemink 2003). Dalam perkembangannya, budidaya kakao di Indonesia mengalami peningkatan yang cukup pesat baik dari segi perluasan areal maupun produksinya. Luas perkebunan kakao pada tahun 2006 telah mencapai 1 191 742 ha dengan produksi mencapai 779 474 ton (Dirjen Perkebunan 2007).
2 meliputi Urea, TSP, SP36, dan KCl sedangkan agen hayati yang diinokulasikan adalah fungi mikoriza arbuskula (FMA) dan bakteri pelarut fosfat (BPF).
Pada saat penggunaan pupuk konvensional dibatasi oleh masalah biaya, fosfat
alam lokal dinilai berpotensi sebagai alternatif pupuk P (Goenadi et al. 2000; Arcand & Schneider 2006). Secara global aplikasi langsung fosfat alam meningkat
karena perkembangan yang cepat di bidang pertanian dan kebutuhan akan fosfat yang murah di negara berkembang (Mayhew 2003). Keuntungan penerapan teknologi ini adalah bahwa fosfat alam yang bermutu rendahpun dapat digunakan dan akan lebih murah (Notohadiprawiro 1989). Di samping itu tanah-tanah tropis yang mengandung Al dan Fe tinggi serta pH tanah yang rendah, fosfat alam bekerja lebih efektif daripada TSP karena P yang dikandungnya tidak mudah terjerap oleh Al dan Fe. Namun demikian faktor kendala utama dalam penggunaan fosfat alam (FA) secara langsung adalah pelepasan P yang lambat tersedia untuk memenuhi kecukupan P yang dibutuhkan tanaman (Vassilev et al. 2001). Oleh karena itu fosfat alam ini lebih sesuai apabila diaplikasikan pada tanaman tahunan.
3 Di Indonesia penggunaan fosfat alam sebagai pupuk masih sangat terbatas. Hal ini disebabkan bukan hanya endapan fosfat alam yang jumlahnya tidak banyak tetapi juga karena kadar P nya sangat bervariasi dari rendah sampai tinggi, dan umumnya tergolong rendah (Sabiham & Amat 1990). Di lain pihak terdapat lahan kering yang cukup luas yang memiliki sifat-sifat di antaranya kandungan unsur hara (terutama P, Ca dan Mg) yang rendah serta kemasaman tanah dan kadar Aldd tinggi. Pada umumnya fosfat alam dengan kandungan P dan Ca yang cukup tinggi diharapkan dapat menambah ketersediaan P bagi tanaman meskipun unsur tersebut dalam bentuk lambat tersedia. Idris (1995) dalam penelitiannya menunjukkan bahwa pemberian fosfat alam (Bogor dan Lamongan) nyata memperbaiki status P dan Ca tanah. Salah satu upaya untuk mengatasi kekurangan unsur hara dalam tanah adalah penggunaan fosfat alam yang berasal dari Distrik Ayamaru, Papua. Sampai sejauh ini belum dikembangkan secara optimal.
4 dibandingkan dengan TSP. Di samping itu fosfat alam bermanfaat ganda karena selain sebagai sumber P juga dapat meningkatkan pH karena dengan pemanasan cenderung menaikkan pH dari 6.19 (tanpa pemanasan) menjadi pH 6.88 dengan temperatur 600oC (Musaad et al. 1996).
Rendahnya tingkat kelarutan fosfat alam menjadi kendala dalam pemanfaatan secara langsung untuk bidang pertanian. Rumawas (1990) menyatakan bahwa pemanfaatan fosfat alam secara langsung atau dengan perlakuan minim dalam pertanian dapat ditempuh melalui tiga jalur, yaitu (1) secara fisik (penggilingan hingga 100 mesh, kalsinasi), (2) kimia (pelarutan dengan asam keras, asam humat), dan (3) mikrobiologik (mikoriza, mikroba pelarut fosfat). Bakteri pelarut fosfat dan asam humat berfungsi meningkatkan kelarutan fosfat, sedangkan penyerapan oleh tanaman dapat ditingkatkan dengan adanya simbiosis akar tanaman dengan fungi mikoriza.
5 karena itu tanggap tingkat pertumbuhan bibit kakao siap tanam biasanya lebih baik dibandingkan dengan tanaman tanpa inokulasi dan akan sangat berarti setelah di lapang (Yahya et al. 2000).
Beberapa hasil penelitian menunjukkan bahwa inokulasi bibit kakao dengan
Glomus fasciculatum menghasilkan serapan Ca, Mn, bobot kering tanaman dan
diameter batang yang lebih besar dibandingkan dengan Acaulospora delicata
(Widiastuti & Baon 1994). Pada bibit kakao yang diinokulasi dengan FMA campuran
Acaulospora sp., Gigaspora sp. dan Glomus sp. lebih tahan terhadap cekaman
kekeringan hingga 55% (Yahya et al. 2000).
Penggunaan bakteri pelarut fosfat (BPF) dalam bidang pertanian telah banyak dilakukan karena mempunyai peranan yang sangat besar dalam membantu penyediaan hara P bagi tanaman. Hal ini karena meskipun biasanya P total dalam tanah terdapat dalam jumlah yang banyak tetapi ketersediaannya bagi tanaman sangat rendah. Kondisi demikian sering menjadi faktor pembatas terhadap pertumbuhan tanaman (Mikanová & Nováková 2002). Tanaman hanya menyerap 10-25% P yang diberikan melalui pemupukan, sebagian besar mengakibatkan perubahan kimia dalam tanah menjadi bentuk tidak larut dan tidak tersedia bagi tanaman (Jumaniyazova et al. 2004). Bakteri pelarut fosfat mampu mengubah bentuk-bentuk fosfat yang tidak terlarut menjadi bentuk terlarut sehingga tersedia bagi tanaman antara lain melarutkan fosfat yang terikat oleh aluminium, besi maupun kalsium serta mampu memineralisasi fosfat organik misalnya fitat. Bakteri pelarut fosfat tersebut antara lain Azospirillum spp.,
6 mikroorganisme pelarut fosfat di antaranya dipengaruhi oleh jenis fosfat dalam media yang dapat dilarutkan (Mikanová & Nováková 2002).
Beberapa hasil penelitian menunjukkan kemampuan berbagai spesies bakteri dalam melarutkan senyawa fosfat sukar larut seperti trikalsium-fosfat, dikalsium-fosfat, hidroksiapatit, dan batuan fosfat (fosfat alam). Di antara genus bakteri yang memiliki kemampuan tersebut adalah Pseudomonas, Bacillus, Rhizobium, Burkholderia,
Achromobacter, Agrobacterium, Micrococcus, Flavobacterium, dan Erwinia.
Penggunaan bakteri pelarut fosfat tersebut sebagai inokulan secara simultan meningkatkan penyerapan P oleh tanaman dan meningkatkan hasil. Khususnya strain dari genus Pseudomonas, Bacillus dan Rhizobium merupakan pelarut fosfat yang paling kuat (Suh 2005). Karti (2003) menunjukkan bahwa isolat bakteri pelarut fosfat RJM.30.2, FT.3.2, FT.3.3, B8016495, dan B8016498 mempunyai potensi meningkatkan serapan P pada rumput S. splendida dan C. gayana, sedangkan Noor (2003) menunjukkan bahwa Pseudomonas fluorescens dapat meningkatkan kelarutan P pada tanah Ultisol yang diaplikasi fosfat alam Tunisia.
7 (pertukaran anion) dan adsorbsi spesifik yaitu pertukaran ikatan, yang secara simultan meliputi pengkelatan dan pembentukan komplek dalam larutan tanah.
Penggunakan senyawa humat pada tanah-tanah pertanian memberikan banyak keuntungan terutama pada tanah-tanah dengan kandungan bahan organik rendah dan merupakan bagian integral ekosistem pertanian yang memainkan peranan penting dalam siklus hara dan karbon secara global (MacCarthy 2003). Bahan humat diketahui dapat meningkatkan keefektifan fosfat alam karena mampu melepaskan PO43- dan Ca2+ dari fosfat alam yang sukar larut, mampu mengkomplek logam dari larutan dan menstimulir metabolisme mikrobia (Mayhew 2004). Aplikasi asam humat pada tanah berpengaruh terhadap penyerapan hara makro maupun mikro yaitu melalui pengaruhnya terhadap laju pelepasan unsur hara dari tanah (Lulakis & Petsas 1995), terutama Fe, Zn, dan Mn (Ayuso et al. 1996), juga mempengaruhi translokasi Fe dari akar ke tajuk (MacCharty et al. 1990).
8
Azospirillum brasiliense yang diaplikasikan bersama FMA dapat meningkatkan
sporulasi dan keefektifan FMA (Hanafiah 2001), meningkatkan aktivitas nitrogenase dan fiksasi N2 untuk membentuk N-NH3 yang digunakan oleh tanaman untuk meningkatkan aktivitas fotosintesis. Fotosintat yang meningkat dapat mendorong pertumbuhan FMA karena suplai C dari inang tercukupi. A. brasiliense mampu meningkatkan enzim perombak P-organik seperti fitat yang menyebabkan peningkatan P tersedia sehingga meningkatkan fungsi FMA dalam pengambilan P yang disuplai ke tanaman inang untuk pertumbuhannya. Inokulasi ganda Glomus intraradices dan
Bacillus subtilis secara nyata meningkatkan biomasa dan akumulasi N dan P dalam
jaringan tanaman bawang (Allium cepa L.), dan sedikitnya 75% P pada tanaman berasal dari penambahan fosfat alam (Toro et al. 1997).
Perlakuan ganda BPF dengan mikoriza dapat meningkatkan bobot kering tanaman. Interaksi antara dua FMA Glomus mosseae dan G. fasciculatum dengan bakteri pelarut fosfat Azospirillum sp., Pseudomonas sp. Basillus sp. dan Enterobacter
sp. dapat meningkatkan pertumbuhan dan serapan hara tanaman (Toro et al. 1996). Interaksi ganda yang positip terjadi karena adanya pengaruh fisiologis tanaman yang berkaitan dengan asimilasi C. Mikoriza mendapatkan sumber C dari tanaman sehingga dapat berkembang dengan baik.
Interaksi antara tanaman, FMA, dan bakteri pelarut fosfat bersifat spesifik, bergantung pada spesies/kultivar tanaman, spesies FMA, spesies/strain bakteri, dan lingkungannya. Azospirillum sp., Basillus sp., dan Enterobacter sp. berinteraksi positif dengan G. mosseae tetapi berinteraksi negatif dengan G. fasciculatum, sedangkan
9 sangat dipengaruhi oleh interaksi tanaman dan mikroba (Kaeppler et al. 2000). Oleh karena itu keefektifan bakteri pelarut fosfat, asam humat, dan FMA dalam meningkatkan ketersediaan P, selain bergantung pada jenis bakteri dan FMA itu sendiri, juga sangat bergantung pada jenis tanaman, dan jenis tanah (pupuk) serta interaksi ketiganya.
10 TANGGAP MORFOFISIOLOGI BIBIT KAKAO YANG DIBERI FOSFAT
ALAM AYAMARU PAPUA, ASAM HUMAT, INOKULASI FMA, DAN BAKTERI PELARUT FOSFAT
1. Tanggap bibit kakao yang diinokulasi FMA pada berbagai dosis fosfat alam Ayamaru
Tujuan :
Mendapatkan inokulum FMA efektif dan
dosis fosfat alam Ayamaru optimum pada
pembibitan kakao
Pendekatan :
Menguji FMA pada beberapa tingkat dosis fosfat alam Ayamaru yang diberikan pada bibit kakao
Hasil yang diharapkan:
Inokulum FMA yang efektif dan dosis optimum fosfat alam Ayamaru pada bibit kakao
2. Tanggap bibit kakao yang diinokulasi bakteri pelarut fosfat pada berbagai dosis fosfat alam Ayamaru
Tujuan :
Mendapatkan isolat yang efektif terhadap fosfat alam Ayamaru yang diberikan pada bibit kakao
Pendekatan :
Penguji bakteri pelarut fosfat pada bebe-rapa tingkat dosis fosfat alam Ayamaru yang diberikan pada bibit kako
Hasil yang diharapkan:
Diperolehisolat bakteri yang efektif dan dosis optimum fosfat alam Ayamaru pada bibit kakao
4.Tanggap bibit kakao yang diinokulasi FMA dan bakteri pelarut fosfat pada berbagai dosis fosfat alam Ayamaru yang diberi asam humat.
Tujuan :
Mendapatkan perlakuan kombinasi yang efektif pada bibit kakao yang diberi FA sehingga diperoleh kualitas bibit yang baik
Pendekatan :
Pengujian kombinasi FMA Mycofer, isolat FT.3.2, dan dosis 3.10-3 mL asam
humat/bibit pada pembibitan kakao yang diberi FA
Hasil yang diharapkan :
Kombinasi yang paling efektif untuk meningkatkan serapan P dan pertumbuhan bibit kakao yang diberi FA.
3. Tanggap bibit kakao pada beberapa tingkat dosis fosfat alam Ayamaru dan asam humat
Tujuan :
Mempelajari pengaruh tingkat dosis asam humat terhadap fosfat alam Ayamaru dan pengaruhnya pada bibit kakao
Pendekatan :
Menguji beberapa dosis asam humat pada beberapa tingkat dosis fosfat alam Ayamaru
Hasil yang diharapkan:
Diperoleh dosis asam humat yang efektif terhadap fosfat alam Ayamaru padabibit kakao
11 Percobaan pertama, kedua, dan ketiga dilaksanakan secara pararel, sedangkan percobaan keempat dilaksanakan setelah diperoleh hasil dari ketiga percobaan sebelumnya. Tanggap bibit kakao akibat tingkat dosis fosfat alam Ayamaru pada percobaan pertama, kedua, ketiga akan digunakan sebagai dasar penentuan tingkat dosis pada percobaan keempat.
Inokulum FMA terbaik hasil percobaan pertama, isolat bakteri pelarut fosfat terbaik hasil percobaan kedua, dan tingkat dosis asam humat yang paling efektif dari percobaan ketiga digunakan sebagai penyusun perlakuan kombinasi pada percobaan yang keempat.
Tujuan dan Manfaat
Penelitian ini bertujuan untuk :
1. Mempelajari pengaruh inokulasi FMA pada tanggap bibit kakao terhadap fosfat alam Ayamaru.
2. Mempelajari pengaruh inokulasi bakteri pelarut fosfat pada tanggap bibit kakao
terhadap fosfat alam Ayamaru.
3. Mempelajari pengaruh berbagai taraf dosis asam humat terhadap pelarutan P dan tanggap bibit kakao terhadap fosfat alam Ayamaru.
4. Mempelajari pengaruh perlakuan kombinasi FMA, bakteri pelarut fosfat, dan asam humat terhadap pelarutan P dan tanggap bibit kakao terhadap fosfat alam Ayamaru. 5. Mempelajari mekanisme pelarutan P oleh bakteri pelarut fosfat terhadap fosfat
alam Ayamaru pada pertumbuhan bibit kakao.
12 Dari penelitian ini diharapkan dapat meningkatkan nilai agronomis fosfat alam Ayamaru pada pembibitan kakao melalui aplikasi asam humat, inokulasi FMA dan bakteri pelarut.
Hipotesis
1. Inokulasi FMA pada bibit kakao meningkatkan keefektifan pemberian fosfat alam Ayamaru.
2. Keefektifan pemberian fosfat alam Ayamaru pada bibit kakao ditingkatkan apabila diinokulasi bakteri pelarut fosfat.
3. Keefektifan pemberian fosfat alam Ayamaru pada bibit kakao ditingkatkan apabila diaplikasi asam humat.
4. Keefektifan pemberian fosfat alam Ayamaru pada bibit kakao ditingkatkan apabila diberi perlakuan kombinasi di antara FMA, bakteri pelarut fosfat, dan asam humat. 5. Mekanisme utama pelarutan fosfat alam Ayamaru dan penurunan kadar Aldd media
disebabkan asam organik yang dihasilkan oleh bakteri pelarut fosfat.
II
TINJAUAN PUSTAKA
Ketersediaan Fosfor Tanah
Fosfor merupakan salah satu hara esensial tanaman yang ketersediaannya di tanah sangat kurang terutama pada tanah-tanah masam yang meliputi 30% tanah di dunia pada semua kontinen (Raghothama 1999). Pada permukaan bumi kandungan P bervariasi 0.005 – 0.15% (Havlin et al. 1999). Menurut Pusat Penelitian Tanah dan Agroklimat (1992) yang termasuk dalam tanah masam tersebut adalah termasuk dalam ordo Inceptisol, Ultisol, Entisol, Oxisol, dan Spodosol. Pada jenis-jenis tanah ini ketersediaan P rendah karena P terikat kuat oleh aluminium dan besi sehingga sulit tersedia bagi tanaman (Raghothama 1999).
Ketersediaan P dalam larutan tanah ditentukan oleh banyak faktor yang meliputi jenis tanah dan kemasaman, suhu tanah, kandungan air tanah, dan konsentrasi Ca (Schachtman et al. 1998). Pada beberapa jenis tanah kandungan P lebih tinggi dari yang diperlukan untuk pertumbuhan optimum tanaman, tetapi P ini dalam keadaan tidak tersedia bagi tanaman. Rata-rata konsentrasi P dalam larutan tanah adalah 0.05 ppm dan sangat bervariasi di antara jenis tanah. Konsentrasi P dalam larutan yang diperlukan oleh kebanyakan tanaman bervariasi dari 0.003 sampai 0.3 ppm dan bergantung pada spesies tanaman dan tingkat produksinya (Havlin et al. 1999), sedangkan kemampuan tanaman mengambil P dari larutan tanah juga bervariasi bergantung pada spesies (genotipe) dan dipengaruhi oleh interaksi antara tanaman dan mikroba (Kaeppler et al. 2000).
14 20% - 80% P dalam tanah terdapat dalam bentuk organik, yaitu asam phytic (inositol
heksafosfat) yang biasanya merupakan komponen utama. Walaupun mobil dalam
tanaman, P tidak mobil dalam tanah (McWilliams 2003). Rendahnya ketersediaan P pada sebagian besar tanah membatasi penyerapan oleh tanaman. Karena laju difusi P lambat (10-12 -10-15.cm2.s-1), maka laju penyerapan tanaman yang tinggi membuat zona deplesi/pengurasan P di sekitar akar (Schachtman et al. 1998; Smith 2002).
Fosfor yang diambil oleh tanaman dari larutan tanah berada dalam kesetimbangan dengan fosfat yang dijerap dalam mineral tanah dan koloid dalam fase tanah padat. Mekanisme pengambilan ditunjukkan oleh erapan P sebagai penyangga sejumlah P dalam larutan (Fox & Kamprath 1970, diacu dalam Smith et al. 2003). Fosfor organik dalam tanah tidak langsung tersedia untuk tanaman tetapi terlebih dahulu harus dimineralisasi menjadi P anorganik yang selanjutnya berada dalam reaksi kesetimbangan dalam larutan tanah (Reiseanuer 1966, diacu dalam Smith et al. 2003).
15 dalam larutan secara cepat terutama di sekitar akar. Beberapa spesies mengembangkan sistem percabangan akar yang tinggi dengan lebih banyak akar rambut sehingga lebih efisien dalam penyerapan P.
Bentuk keberadaan P dalam larutan tanah bergantung pada pH tanah. Apabila pH tanah di bawah 6.0 kebanyakan P berada dalam bentuk monovalen H2PO4-, sedangkan H3PO4- dan HPO42- hanya ada dalam proporsi kecil. Beberapa penelitian terhadap ketergantungan pH dalam pengambilan P oleh tanaman diketahui bahwa laju pengambilan tertinggi terjadi pada pH tanah 5.0 – 6.0 yang mana fosfat dominan dalam bentuk H2PO4-, sedangkan pada pH tanah 7.2 fosfat dominan dalam bentuk HPO42-. Tanaman mengambil HPO42- lebih lambat daripada H2PO4- (Schachtman et al. 1998; Havlin et al. 1999).
Ketika tanaman tercekam P area permukaan akar yang kontak dengan tanah mungkin juga ditingkatkan oleh tanaman dengan meningkatkan diameter akar (Ma et al. 2001) dan baik panjang maupun kerapatan rambut akar mungkin meningkat (Bates & Lycnh 2000). Beberapa tanaman mengembangkan bagian khusus atau akar-akar proteoid. Ekskresi karboksilat oleh akar tanaman bertujuan melepaskan P dari senyawa besi (Fe) dan aluminium (Al) tanah. Hal ini akan terlihat adanya peningkatan ekskresi dari akar pada tanaman yang tercekam P (Lambers & Poot 2002).
16 tanah, atau dengan kata lain mikoriza lebih berkembang pada kondisi P tersedia rendah, karena eksudat akar yang defisien P merangsang perkembangan mikoriza.
Peranan FMA terhadap Penyerapan Fosfor
Infeksi sistem perakaran pada sebagian besar tanaman oleh FMA terjadi pada 83% dikotil dan 79% monokotil dengan membentuk asosiasi simbiotik antara tanaman dengan mikoriza (Swift 2004), dengan keuntungan tanaman dapat mengambil P lebih besar dari larutan tanah (Smith 2002; Smith et al. 2003). Swift (2004) menyatakan bahwa salah satu pengaruh infeksi oleh fungi mikoriza arbuskula pada tanaman inangnya adalah peningkatan penyerapan P yang disebabkan oleh kemampuan mikoriza menyerap P dari tanah dan mentransfernya ke akar tanaman inang. Ketika tanaman kekurangan mineral seperti P atau N, hubungan simbiotik ini akan menguntungkan dan mendorong pertumbuhan tanaman (Morgan et al. 2005). Di samping meningkatkan serapan P, kolonisasi akar tanaman oleh FMA meningkatkan serapan tanaman terhadap N dengan menyerap amonium dan nitrat dari tanah (George et al. 1995). Oleh karena itu inokulasi tanaman dengan FMA merupakan cara efisien untuk penambahan P melalui pemupukan (FAO 2005) dan dapat mengurangi biaya pemupukan fosfat pada budidaya tanaman perkebunan (Rahim 2002).
Morfologi akar tanaman penting untuk memaksimalkan penyerapan P, sebab sistem perakaran dengan ratio area permukaan dan volume yang tinggi akan lebih efisien menjelajah volume tanah yang luas. Oleh karena itu mikoriza penting pada tanaman dalam penyerapan P, karena mikoriza ini meningkatkan kemampuan akar mengeksplorasi tanah lebih luas (Schachtman et al. 1998).
17 meningkatkan pertumbuhan tanaman (Rahim 2002). Hifa dari fungi mikoriza dapat menyebar hingga lebih dari 25 cm dari akar, dengan demikian akan meningkatkan kemampuan eksplorasi tanah untuk mendapatkan hara. Oleh karena itu aliran P pada akar yang berkoloni dengan fungi mikoriza 3 – 5 kali lebih cepat dibandingkan dengan akar yang tidak berkoloni dengan mikoriza dengan laju 10-11 mol m-1.s-1 (Smith & Read 1997). Pengambilan P oleh mikoriza selanjutnya ditranslokasikan melalui hifa, arbuskula ke sel-sel akar tanaman. Fungi mikoriza arbuskula (FMA) dicirikan oleh formasi struktur (arbuskular) percabangan di dalam sel kortek akar (Gambar 2).
Arbuskular dengan percabangan yang tinggi masuk ke dalam membran sel kortek akar tanaman inang, membentuk simbiotik dengan area permukaan yang sangat luas, yaitu membran peri-arbuskula (Smith et al. 2003). Fungi mikoriza arbuskula dan sel tanaman tetap dipisahkan oleh membran apoplastik dengan topologi mikoriza di bagian luar. Transfer P terjadi melewati ruang apoplastik sehingga melibatkan pembongkaran P di membran peri-arbuskular dan pemuatan ke dalam apoplas akar tanaman. Marchel (2004) menyatakan bahwa transfer fosfat terjadi pada struktur arbuskular dan gelembungan hifa.
18 Dalam simbiosisnya dengan tanaman, mikoriza menerima karbohidrat dan faktor-faktor pertumbuhan dari tanaman inang sebagai sumber energi untuk pertumbuhan dan perkembangannya sedangkan tanaman dapat meningkatkan serapan hara P dan unsur hara lainnya oleh adanya koloni akar dengan mikoriza (Muchovej 2002). Tanaman mensuplai fotoasimilat ke sistem perakaran bermikoriza 4% – 20% (Douds et al. 2000). Sel-sel kortek akar melepaskan karbohidrat ke permukaan simbiosis tanaman-mikoriza oleh aliran pasif yang distimulasi oleh adanya mikoriza (Bago et al. 2000).
Potensi mikoriza dalam membantu tanaman menyerap P bergantung pada kondisi P tanah. Swift (2004) menyatakan bahwa keuntungan yang tinggi dari simbiosis mikoriza dengan tanaman diperoleh pada tanah yang defisien P dan rendah pada tanah yang ketersediaan P-nya tinggi. Lebih lanjut dijelaskan bahwa apabila level P tanah lebih dari 140 mg/kg (140 ppm), maka infeksi mikoriza akan menurun, sedangkan apabila level P tanah 50 mg/kg (50 ppm) maka diperoleh perkembangan mikoriza yang tinggi. Rahim (2002) menyatakan bahwa keefektifan mikoriza berbeda untuk setiap tanaman dan kondisi lingkungannya.
19 Inokulasi mikoriza pada pembibitan tanaman sangat berarti ketika dipindahkan ke lapang karena dengan adanya simbiosis, tanaman lebih tahan menghadapi kondisi lingkungan tumbuh yang kurang menguntungkan (John 1996).
Penggunaan mikoriza dalam skala yang luas seringkali dibatasi oleh adanya hambatan dalam memproduksi inokulan FMA. Media perbanyakan FMA yang banyak digunakan adalah zeolith, tetapi sulit didapat dan harganya relatif mahal. Media campuran tanah Ultisol dan pasir bangunan (1:1) menunjukkan produksi jumlah spora dan derajat infeksi akar yang lebih tinggi dibandingkan dengan media zeolit (Anas & Tampubolon 2004).
Keefektifan inokulum bergantung pada jenis tanaman dan kondisi tanah. Namun demikian pH tanah merupakan faktor pembatas utama sedangkan struktur tanah dan bahan organik mungkin juga mempengaruhi kesesuaian tanah untuk mikoriza. Glomus intraradices spesies yang banyak dijumpai, sesuai pada pH tanah 6 – 9, sedangkan G.
etunicatum banyak dijumpai dan sesuai pada pH masam (John 2000). Inokulasi lebih
dari satu jenis mikoriza meningkatkan penyerapan P lebih tinggi dibandingkan dengan inokulasi tunggal (John 2000; Jansa et al. 2004). Di samping itu kolonisasi mikoriza juga dipengaruhi oleh level CO2, intensitas cahaya, lamanya cahaya, dan kualitas cahaya (Singh 2005).
Fosfat Alam Ayamaru Papua
20 2004). Kimiti dan Smithson (2002) menyatakan bahwa pada beberapa tahun terakhir penggunaan batuan fosfat sebagai sumber P secara langsung telah banyak dilakukan.
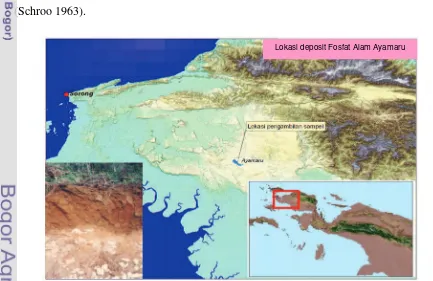
Di Indonesia endapan fosfat alam ditemukan tersebar di beberapa lokasi. Lokasi endapan fosfat alam sebagian besar terdapat di Pulau Jawa dan Madura, dan lainnya terdapat di Kalimantan, Sulawesi dan Papua. Total endapan di Jawa dan Madura diperkirakan 9.5 – 20 juta ton (Bisri & Permana 1991). Endapan fosfat alam di Papua terdapat di Distrik Ayamaru, Kabupaten Sorong Selatan pada ketinggian 300 – 400 m di atas permukaan laut, yang dipetakan meliputi luasan kurang lebih 100 000 ha (Reynders & Schultz 1958).
Diketahuinya tanah Ayamaru berkadar fosfat tinggi adalah berdasarkan hasil analisis fosfat dari contoh tanah oleh Schroo & Mouthaan (1960). Hasil analisis contoh dengan ekstraksi asam Fleischmann (50% H2SO4 + 50% HNO3) mendapatkan nilai 1% sampai 12% P2O5. Berbeda dengan endapan fosfat alam di lokasi lain yang berupa batuan, endapan fosfat yang terdapat di daerah Ayamaru berupa tanah. Hasil penelitian selanjutnya (Schroo 1963) menunjukkan bahwa mineral fosfat Ayamaru adalah mineral krandalit (CaAl3(PO4)2(OH)5.H2O) dengan ikatan nya disebut P-krandalit. Di Indonesia jenis endapan fosfat alam ini diketahui hanya terdapat di Distrik Ayamaru, Irian Jaya Barat (Gambar 3).
21 analisis adalah pasir, debu, gabungan debu dan liat, berturut-turut mengandung P2O5 sebesar 5.67%, 18.6% dan 18.55%.
Schroo & Mouthaan (1960) telah melakukan percobaan pot untuk mendapatkan keterangan apakah tanah fosfat alam Ayamaru dapat digunakan sebagai pupuk sumber P. Percobaan menggunakan tanaman jagung dan padi sebagai tanaman indikator. Tanah sebagai media tumbuh berasal dari Distrik Mindiptana, Merauke yang miskin P dan masam. Dari percobaan ini diperoleh kesimpulan bahwa pemberian 25 g fosfat alam Ayamaru mempunyai pengaruh yang sama dengan 100 mg P2O5 monokalsium fosfat untuk tanaman padi. Percobaan dengan menggunakan tanaman indikator jagung disimpulkan bahwa pemberian 50 g tanah Ayamaru mempunyai pengaruh yang sama dengan pemberian 100 mg P2O5 monokalsium fosfat. Hal ini menunjukkan bahwa tanah Ayamaru dapat menyediakan P2O5 sebanyak 0.2% – 0.4 % untuk tanaman (Schroo 1963).
[image:40.595.85.518.417.698.2]Lokasi deposit Fosfat Alam Ayamaru
22 Penggunaan fosfat alam sebagai pupuk langsung merupakan alternatif yang ekonomis dibandingkan dengan pupuk industri. Hasil penelitian Musaad et al. (1996) menunjukkan bahwa fosfat alam Ayamaru (krandalit) yang dipanaskan atau melalui proses termal (500 – 700oC) terjadi perubahan sifat fisik maupun kimia yang lebih baik, yaitu terjadi kenaikan pH, P-larut air, P-larut sitrat 2%, dan P-potensial. Pada pemanasan hingga temperatur 700oC, peningkatan P-larut air dan P potensial berturut-turut mencapai 320.64% dan 27.38%. Perlakuan kalsinasi terhadap beberapa fosfat alam krandalit dari Brasil menunjukkan dengan pemanasan 700oC kelarutan fosfat alam Tapira dalam Neutral Ammonium Citrate Extraction (NAC) meningkat 84%, fosfat alam Joquiá pada 500o C meningkat 60%, dan fosfat alam Catalão pada 700oC meningkat 44% (Francisco et al. 2007). Amat (1991) dari hasil penelitiannya menyimpulkan bahwa secara teknis tanah endapan fosfat alam Ayamaru dapat digunakan sebagai sumber pupuk P. Kelarutannya dapat ditingkatkan dengan penambahan bahan organik (pupuk kandang sapi dan ayam) dari 0.2% menjadi 0.55%. Hasil penelitian Akande et al. (2005) menunjukkan bahwa penambahan pupuk kandang ternak ayam pada pupuk P dari batuan fosfat dapat meningkatkan kelarutan P 2 -3 kali lebih besar daripada kontrol. Namun demikian oleh karena pelepasan P lambat, maka direkomendasikan bahwa pelepasan P dari fosfat alam mungkin dapat ditingkatkan dengan inokulasi mikroorganisme pelarut fosfat (Rumawas 1990).
23
Bakteri Pelarut Fosfat
Kemampuan mikroba melarutkan mineral fosfat yang sulit larut dalam tanah adalah proses yang sangat penting dalam ekosistem alami dan pada tanah-tanah pertanian. Di dalam tanah terdapat banyak fosfat, tetapi ketersediaannya bagi tanaman sangat rendah dan sering menjadi faktor pembatas terhadap pertumbuhan dan produksi tanaman (Wissuwa 2003). Oleh karena itu pada aplikasi pemupukan mineral dengan inokulasi mikroba pelarut fosfat dapat meningkatkan ketersediaan P pada tanah (Mikonová & Nováková 2002). Raja et al. (2002) menyatakan bahwa mikroorganisme dan fungi mikoriza memainkan peranan penting dalam metabolisme dan produktivitas tanaman.
Beberapa bakteri tanah terutama yang termasuk dalam genus Rhizobium (Suh 2005), Pseudomonas, Bacillus, Mycobacterium, Micrococcus, Flavobacterium,
Bacterium, Citrobacter, Enterobacter (Rao 1994; Siripin 2002), mempunyai
kemampuan mengubah kelarutan fosfat dalam tanah ke dalam bentuk terlarut dengan sekresi asam organik seperti sitrat, glutamat, laktat, oksalat, glioksilat, tartarat, ketobutirat, malat, fumarat, asetat, propionat, dan suksinat. Asam-asam ini membuat terputusnya bentuk-bentuk ikatan fosfat dengan mengkomplek Al dan Fe. Pada tanah-tanah tropis umumnya ketersediaan P untuk pertumbuhan tanaman rendah. Oleh karena itu bakteri pelarut fosfat sangat penting membantu menyediakan fosfat untuk pertumbuhan dan perkembangan tanaman.
24 biasanya diikuti dengan penurunan pH tanah yang cepat sehingga menyebabkan pelarutan Ca-P. Pada tanah masam, pelarutan AlPO4 melalui sekresi proton bersamaan dengan asimilasi NH4+ tanpa menghasilkan asam organik (Illmer et al. 1995).
Menurut Rao (1982) proses kelarutan fosfat yang sukar larut diawali dengan dihasilkannya asam-asam organik oleh mikroorganisme. Asam-asam organik yang disekresikan oleh mikroroganisme dapat melarutkan P. Kecepatan pelepasan P dari bentuk yang tidak tersedia menjadi bentuk yang tersedia dapat disebabkan oleh adanya pelepasan gas H2, CO2, H2S, dan CH2 sebagai akibat berlangsungnya proses reduksi dan dekomposisi bahan organik (Sabiham et al. 1983). Reaksi yang terjadi adalah :
1. Pembebasan P akibat produksi H2 (reduksi) : FePO4 Fe3+ + PO4
3-H2 2 H+ + e-
Fe3+ + e- Fe2+ +
FePO4 + H2 Fe2+ + H2PO4 -2. Pembebasan P akibat produksi CO2 (oksidatif) :
CO2 + H2O HCO3- + H+
Ca3(PO4)2 + 6HCO3- 3Ca (HCO3)2 + 2 PO43-
25 Reaksi pelarutan P akibat adanya gugus karboksil dapat ditunjukkan sebagai berikut :
1.
OH O- OH
M OH + R – C = O M OH O + H2PO4- H2PO4- R-C=O
2. R–C–O
OH O- O O
M H2PO4 + 3 R – C =O M–O-C– R + H2PO4 -H2PO4 O
R–C-O
kompleks-M asam organik kompleks- asam organik ortofosfat
M= Al3+ atau Fe3+
P organik dapat dimineralisasi atau dimobilisasi (Brady 1990) :
immobilisasi
Fe3+, Al3+, Ca2+
P-organik H2PO4- Fe-P, Al-P, Ca-P mineralisasi
Hasil penelitian Noor (2003) menunjukkan bahwa bakteri pelarut fosfat
Pseudomonas fluorescens pada 6 minggu setelah penanaman kedelai dapat
26 0.197, 0.201, 0.369, 0.188, 0.221, dan 0.240 berbeda nyata dengan kontrol 0.160. Isolat FT.3.3 efektif meningkatkan serapan P pada S. splendida sedangkan pada C. gayana peningkatan serapan P terbaik dihasilkan oleh isolat RJM.30.2 dan FT.3.2.
Peranan Asam Humat
Bahan organik yang terdapat di dalam tanah berasal dari sisa-sisa tanaman dan hewan. Bahan ini terdiri atas serasah, fraksi bahan organik ringan, biomassa mikroorganisme, bahan humat (asam humat, asam fulvat, dan humin), dan senyawa non-humat (asam amino, karbohidrat, lemak, lilin, resin). Bahan humat tanah dikelompokkan berdasarkan sifat kelarutannya menjadi empat kelompok, yaitu humin, asam fulvat, himatomelanik, dan asam humat.
Humin adalah fraksi humat tanah yang tidak larut baik dalam alkali maupun asam. Asam fulvat adalah fraksi humat yang larut baik dalam asam maupun alkali. Himatomelanik adalah bagian asam humat yang larut dalam alkohol. Asam humat adalah fraksi humat tanah yang larut dalam keadaan alkalin tetapi tidak larut dalam asam (Stevenson 1994). Asam humat dapat dibagi menjadi dua kelompok berdasarkan kelarutannya dengan elektrolit pada keadaan alkalin, yaitu (1) asam humat coklat, yaitu tidak menggumpal dan (2) asam humat abu-abu, yaitu mudah menggumpal dalam larutan elektrolit.
27 oksida, hidroksida, mineral, dan organik termasuk yang beracun untuk membentuk komplek yang larut air maupun yang tidak larut dalam air. Interaksi tersebut berupa interaksi pertukaran ion, jerapan permukaan, atau pengkelatan.
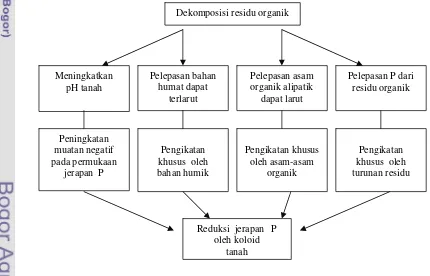
Asam humat hasil dekomposisi bahan organik berperan dalam meningkatkan ketersediaan P tanah (Tisdale et al. 1990) melalui (1) pembentukan senyawa komplek fosfohumat yang lebih mudah diserap tanaman, (2) pertukaran anion fosfat oleh anion organik, (3) terbungkusnya sesquioksida oleh humus, sehingga mengurangi kemampuan logam memfiksasi fosfat. Di samping itu bahan organik juga berpengaruh terhadap sifat fisik tanah seperti kapasitas menahan air, suhu dan sifat kimia seperti kapasitas tukar kation dan pH. Suatu model yang menunjukkan proses-proses yang dapat menginduksi penurunan pengikatan P dan meningkatkan ketersediaan P apabila residu bahan organik ditambahkan ke dalam tanah (Gambar 4).
Dekomposisi residu organik
Meningkatkan pH tanah
Pelepasan bahan humat dapat
terlarut
Pelepasan asam organik alipatik
dapat larut
Peningkatan muatan negatif pada permukaan
jerapan P
Pengikatan khusus oleh bahan humik
Pengikatan khusus oleh asam-asam
organik
Reduksi jerapan P oleh koloid
tanah
Pengikatan khusus oleh turunan residu Pelepasan P dari
[image:46.595.87.517.420.696.2]residu organik
28 Zimmer (2004) dan Mayhew (2004) menyatakan fungsi penting senyawa humat dalam sistem pertanian, yaitu merangsang pertumbuhan akar, aktivitas kehidupan biologi tanah (terutama populasi FMA), mengkelat mineral, memperbaiki serapan hara terutama P, N dan S, menurunkan kebutuhan pemupukan N, melarutkan mineral, memperbaiki struktur tanah, meningkatkan kapasitas air tanah. Hal ini dapat terjadi karena senyawa humat merupakan senyawa yang sangat aktif dalam tanah, dengan kapasitas tukar kation dan anion yang besar (Tan 2003).
Senyawa humat juga mempunyai kemampuan melarutkan dan mengkomplek mineral dari batuan fosfat. Pelarutan mineral dari batuan fosfat oleh aktivitas biologi akan lebih ditingkatkan dengan adanya senyawa humat (Mayhew 2004). Senyawa humat dapat meningkatkan keefektifan batuan fosfat karena pelarutan anion PO43- dan kation Ca2+ dari mineral batuan yang sukar larut, karena total pengasaman yang tinggi dan kemampuannya mengkomplek dan mengkelat larutan yang dihasilkan dan menstimulir metabolisme mikrobia tanah. Senyawa humat juga mengkelat Fe, Zn, Al, dan Cu dan mengkomplek elemen lainnya.
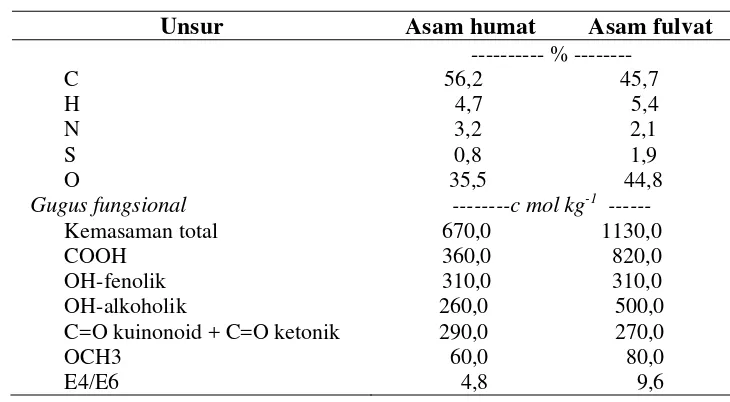
29 perbandingan komposisi asam humat dan asam fulvat yang telah dilakukan disajikan pada Tabel 1.
Tabel 1 Karakteristik hasil analisis kandungan unsur dari asam humat dan asam fulvat (Schnitzer 1997)
Unsur Asam humat Asam fulvat
C H N S O
Gugus fungsional
Kemasaman total COOH
OH-fenolik OH-alkoholik
C=O kuinonoid + C=O ketonik OCH3
E4/E6
--- % --- 56,2 45,7 4,7 5,4 3,2 2,1 0,8 1,9 35,5 44,8
---c mol kg-1
III
TANGGAP BIBIT KAKAO YANG DIINOKULASI
FMA PADA BERBAGAI DOSIS FOSFAT ALAM AYAMARU
ABSTRAK
Fosfor merupakan salah satu hara esensial tanaman. Oleh karena itu untuk mendapatkan pertumbuhan yang baik diperlukan pemupukan P atau juga inokulasi fungi mikoriza arbuskula (FMA). Inokulasi FMA diharapkan dapat meningkatkan efisiensi pupuk P dan meningkatkan serapan P tajuk. Penelitian ini bertujuan untuk mendapatkan inokulum FMA yang efektif meningkatkan daya guna fosfat alam Ayamaru (FA) pada bibit kakao.
Penelitian dilaksanakan di Kebun Percobaan Cikabayan IPB, Laboratorium Bioteknologi Hutan, Lab. Kimia dan Kesuburan Tanah, dan Lab. Fisiologi Tanaman IPB. Penelitian dilaksanakan dalam rancangan acak lengkap dua faktor. Faktor pertama adalah lima taraf dosis FA, yakni 0, 0.5, 1.0, 1.5, 2.0 g P2O5/bibit, dan 2.0 g P2O5 /bibit SP36, sebagai pembanding. Faktor kedua adalah inokulum FMA yang terdiri atas tanpa inokulasi FMA, FMA indigenous Manokwari, dan FMA Mycofer. Benih kakao yang digunakan adalah UAH F1 dari Pusat Penelitian Kopi dan Kakao Jember Jawa Timur. Pembibitan dilakukan dalam polibag 20 cm x 30 cm dengan media tanah masam Ultisol, Jasinga dan ditempatkan dalam jaring naungan 60% selama empat bulan.
Hasil penelitian menunjukkan bahwa pemberian FMA terbukti meningkatkan keefektifan fosfat alam Ayamaru. Pada bibit kakao yang diinokulasi FMA Mycofer, peningkatan dosis FA secara linier meningkatkan bobot kering tajuk sebesar 50.14% dan serapan P tajuk 64.88%, sedangkan dengan FMA indigenous Manokwari meningkat sebesar 66.30% dan serapan P tajuk 65.45%, sedangkan bibit tanpa FMA meningkat 73.56% dan serapan P sebesar 121.94%. Pada dosis FA 2.0 g P2O5, apabila dibandingkan dengan bibit tanpa FMA, inokulasi FMA Mycofer lebih efektif meningkatkan bobot kering tajuk (127.55%) daripada FMA indigenous Manokwari (95.97%), sedangkan serapan P tajuk meningkat 45.16% dan 21.29%.
FMA Mycofer terdiri atas Gigaspora margarita (INVAM-105), G. manihotis
(INDO-1), G. etunicatum (NPI-126), dan A. tuberculata (INDO-2), sedangkan hasil identifikasi morfologi spora FMA indigenous Manokwari terdiri atas Acaulospora tuberculata Janos & Trappe, A. scrobiculata Trappe, dan G. aggregatumSchenck & Smith.
ABSTRACT
31
phosphate rock (APR) for cacao seedlings. The study was conducted at the Experimental Farm of IPB, Cikabayan, Bogor, Forest Biotechnology Laboratory, Soil Chemistry and Fertility Laboratory and Plant Physiology Laboratory of IPB, Bogor.
The two factor-factorial experiment was set up in a Completely Randomized Design. The first factor was five levels of APR dosage: 0, 0.5, 1.0, 1.5, 2.0 g P2O5/seedling, and 2.0 g P2O5 SP36/seedling used as comparison; the second factor
was AMF inoculants, consisting of without AM fungi, Manokwari indigenous AMF and Mycofer. F1 UAH cacao seeds from Coffee and Cacao Research Center, Jember, East Java was used and the seedlings were grown on Ultisol, acid soil from Jasinga in 20 cm x 30 cm sized polybag. The seedlings were grown under 60% of shading net for the period of four months.
In this study, AMF inoculation, proved being able to improve the beneficial use of APR application on increasing the growth of cacao seedlings. When the seedlings were inoculated with AMF, the increase of APR dosage resulted in linear increasing of shoot dry-weight as much as 50.14% and of P uptake 64.88%, while inoculation of Manokwari indigenous AMF could increasing the shoot dry-weight as much as 66.30% and P uptake 65.45%, and on the non-mycorrhizal seedling the shoot dry-weight
was increased as much as 73.56% and P uptake 121.94%. On the APR dosage of 2.0 g
P2O5/seedling as comparing to the non-inoculated seedling, inoculation AMF using
Mycofer inoculants was found much more effective to increase the shoot dry-weight (127.55%) than using Manokwari indigenous AMF (95.97%), while P uptake increased 45.16% and 21.29%, respectively.
Mycofer inoculants contained Gigaspora margarita, (INVAM-105) G. manihotis
(INDO-1), G. etunicatum (NPI-126), dan A. tuberculata (INDO-2), while identification based on spore morphology found that Manokwari indigenous AMF were consisted of
Acaulospora tuberculata Janos & Trappe., A. scrobiculata Trappe, and G. aggregatum
Schenk & Smith.
Pendahuluan
32 Fosfat alam Ayamaru merupakan sumber fosfat alam yang terdapat di Distrik Ayamaru Papua dengan kadar P2O5 total 21%. Mineral fosfat alam Ayamaru adalah mineral krandalit (CaAl3(PO4)2(OH)5.H2O) dengan ikatan P-nya disebut P-krandalit (Schroo 1963). Di Indonesia endapan fosfat alam jenis ini diketahui hanya terdapat di Ayamaru Irian Jaya Barat
Asosiasi simbiotik antara tanaman dan Fungi Mikoriza Arbuskula (FMA) bersifat mutualistik yang meliputi 83% dikotil dan 79% monokotil (Swift 2004). Dalam simbiosisnya mikoriza dan tanaman, mikoriza menerima karbohidrat dan faktor-faktor pertumbuhan dari tanaman inang sebagai sumber energi untuk pertumbuhan dan perkembangannya, sedangkan tanaman dapat meningkatkan serapan hara P dan unsur hara lainnya oleh adanya koloni akar dengan mikoriza (Muchovej 2002). Tanaman mensuplai fotoasimilat ke sistem perakaran bermikoriza 4% – 20% (Douds et al.
33 Potensi mikoriza dalam membantu tanaman menyerap P bergantung pada kondisi P tanah. Swift (2004) menyatakan bahwa keuntungan yang tinggi dari simbiosis mikoriza dengan tanaman diperoleh pada tanah yang defisien P dan turun pada tanah yang ketersediaan P-nya tinggi. Lebih lanjut dijelaskan bahwa apabila level P tanah lebih dari 140 mg.kg-1 (140 ppm), maka infeksi mikoriza akan menurun, sedangkan apabila level P tanah 50 mg.kg-1 (50 ppm), maka